Vol.44,No. 1 JOURNALOFVIROLOGY,Oct.1982,P.311-320
0022-538X/82/100311-10$02.00/0
Copyright C 1982, AmericanSociety for Microbiology
Simple
Repeat
Sequence in
Epstein-Barr
Virus
DNA Is
Transcribed in
Latent
and
Productive Infections
MARKHELLER, VICKY VANSANTEN, AND ELLIOTT KIEFF*
DepartmentsofMedicine, Microbiology, Immunology, andVirology, KovlerViralOncologyLaboratory, The
Universityof Chicago, Chicago, Illinois 60637
Received22April1982/Accepted10 June 1982
The BamHI K region of Epstein-Barr virus DNA is transcribed in latently infected cells from Burkitt tumors and in growth-transformed B-lymphocytes latentlyinfected withEpstein-Barrvirus. We determined the nucleotidesequence ofa1,153-basepair Hinflfragmentin BamHIfragmentKfrom the B95-8 Epstein-Barr virus isolate. The fragment contains a remarkable 708-base pair simple sequence repeatarray, designatedIR3, which iscomposedofonly three nucleo-tidetriplet elements: GGG,GCA,andGGA. Thetripletsareorganizedinto three
repeatunits: GCAGGA, GCAGGAGGA, andGGGGCAGGA. Immediately 3'of
IR3 are tandem nearly perfect direct repeats of two different 24-base pair sequences. IR3 isconservedat acolinearpositionin the DNAsof other
Epstein-Barr virus isolates, and a homologous sequence maps at the same
location in the genome ofagenetically related baboonherpesvirus, herpesvirus papio. IR3 is transcribed from lefttorightinlatentlyinfected, growth-transformed IB4 cells. Itencodes partofa2.0-kilobase exonof the 3.7-kilobase cytoplasmic polyadenylated RNA previously detected in IB4 cells (van Santen et al., Proc. Natl. Acad. Sci. U.S.A.78:1930-1934, 1981). IR3 also encodespartsof 2.4- and 1.0-kilobase RNAs inproductively infected B95-8 cells.
Epstein-Barr virus (EBV) is the usual caus-ative agentof infectious mononucleosis. Latent EBV infection stimulates the growth of B-lym-phocytes in vitro and in vivo. The virusmaybe an important etiological factor in two human cancers,Burkittlymphomaandanaplastic naso-pharyngeal carcinoma (for reviews, see refer-ence12).
EBVDNAis alinear, double-stranded mole-cule ofapproximately 175,000 base pairs (bp) (Fig. 1;forreviews, seereferences 6and 7). At both endsof the moleculearedirectrepeatsofa 0.5-kilobase (kb) sequence, TR. Direct repeats ofa3,071-bp sequence, IR1, divide thegenome intoa9,000-bpuniqueregion,
Us,
anda 150,000-bpuniqueregion, UL (4, 4a).DirectrepeatsIR2 and IR4, of123 and 103 bp, respectively, map within the left and right ends, respectively, ofUL
(T.Dambaugh
and E.Kieff,
J.Virol.,
inpress).
There are three regions in EBV DNA which express cytoplasmic polyadenylated (poly(A)] RNAs in latently infected, growth-transformed cells(24, 25, 35, 49). These RNAs are encoded bythe large internal repeat(IRl) sequence and adjacentuniquesequences,by theBamHIRand K fragments at the center of UL, and by the EcoRI D het fragment at the right end of UL (Fig. 1). Atleast 50cytoplasmic poly(A) RNAs
are encoded by UL and Us in productively infected cells (22).
Under moderately stringent hybridization conditions (6xSSC[seebelow]and70°C)human and mouse cell DNAs hybridize to a 1.15-kb Hinflsubfragment ofthe BamHI Kfragmentof EBV DNA (M. Heller, A. Henderson, and E. Kieff, Proc. Natl. Acad. Sci. U.S.A., inpress). This observation, the transcription of the BamHI K fragment in latent and productive infections,and thesuggestionof rearrangements
in EBV DNA centered around the BamHI K
region (17) led us to examine this regionmore closely.Wereporthere the nucleotidesequence of the 1.15-kb Hinflfragmentfrom B95-8 EBV isolate. The fragment contains a remarkable sequence, designated IR3, which consists of interspersed direct repeatsofone6-bp andtwo 9-bp oligonucleotides. These oligonucleotides arecomposed ofonlythree nucleotide triplets.
The BamHI K
fragment
encodes a 3.7-kb cytoplasmic poly(A) RNA in latently infected, growth-transformedcells (49) and 2.2-, 1.9-, and 1.5-kb cytoplasmic poly(A) RNAs in produc-tively infected cells (22). Hybridizations with cDNA made from latently infected cell RNA have suggested that the 3.7-kb RNA is tran-scribed fromlefttorightin the genome orienta-tion indicated in Fig. 1 (49). We confirm the311
on November 10, 2019 by guest
http://jvi.asm.org/
111Ul ...
DL
0 10 30
10 40
U3 U4 l_
-IIIHI
0)040 60507860 070IOC80120 13090 100150 1601n0 170 xI2 b120x&d
50 60 7~0 80 90 100 110 120 130 140 150 160 170 XIO3b
IJhet. .J. A GlL, F CK.a E H C Dhit EcoRI
JhOd C 'V'V'V'V'V'VYH F '0'U'P'0Z' M'S' L E WYR' K 8 G D Eo'TWV1tb 81Bl Wl I1 A 'Ntt Bom HI
T -~
DonmP3HR-lvirolDNA DeIHonn895-8 nrd DNA
0 1 2 3 4
B. I I I
x103
---1
5 -beI
basspairs
BamHI *, *I
00 0
Itc
s
0
a
004
s 0
1414j4 4a
"'0 ea014
I1
1.1')I
I Is-a
I*Io
6
E .
Z :r 00 0
0°
,1(,r
0000o S
;L _
i 0 e -N
i < I
500 1000 bass pcairs
.77 kb Hpa:l
[image:2.496.48.444.38.520.2]I I .73kbAva3I/HpaIl
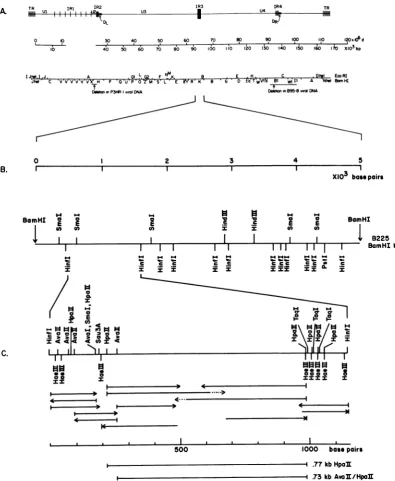
FIG. 1. Organizationof BamHI Kregionin EBV DNA. (A)Structure of the EBVgenome.ThelinearDNAis boundedby multipletandemdirectrepeats(TR)(0.5 kb). Tandeminternal directrepeatregions IR1(3,071bp)(4,
4a),IR2(123 bp),IR3(thispaper),and IR4(103bp)(Dambaugh andKieff,in press)separateregions of
unique-sequenceDNA(Ut toU5).EcoRIand BamHIrestrictionenzymesitesandthe locationsof DNA deletions in P3HR-1 andB95-8 virusisolatesareindicated(for reviews,seereferences 6and7).Thediscontinuityin themap coordinates in IR1 reflects variation in theaveragenumberofcopiesof the IR1 repeatamongdifferentEBV isolates.(B)RestrictionmapofthepDK2255.0-kb BamHI Krecombinant insert. A 32P-labeled terminal Hinfl-BamHI fragment (4.74 to 5.0 kb in pDK225 K) was hybridized to Southern blots of partial restriction
endonuclease digests of theBamHI Kinserttolocate the cleavage sites (44). (C)Restrictionmapof thepDK225
1.15-kbHinfl fragmentandsequencingstrategy.Restriction fragmentsweresequenced bychemicaldegradation
(28).The 5'-to-3'polarityand theextentof the sequenced strandsareindicated byarrows,and the labeled ends
areshownwithavertical mark. Partialsequenceinformation is indicated bydottedarrows.The restrictionmap
is derived from thesequence. The locations ofa776-bpHpaIIfragment anda735-bp AvaII-HpaII fragment
whichwereusedashybridizationprobesinthis studyareindicated below the scale.
312
lamHI
I
B225 amHI KC.
1-Bc
on November 10, 2019 by guest
http://jvi.asm.org/
SIMPLE REPEAT SEQUENCE IN EBV DNA 313
directionof transcription of this RNA and find that the IR3 repeat is transcribed in latently infectedandproductively infected cells.
MATERIALSANDMETHODS
Cellsandvirus. The procedures and materials used
for cell culture and purification of EBV from the
culture fluids of producer cell lines have been
de-scribed previously (10). Intracellular viral DNAs were partially purified from sodium dodecyl sulfate-protein-ase K lysates of latently infected cell cultures by
repeated cycles of isopycnic centrifugation (17).
AG876 (34), W91, and P3HR-1 (21) are virus-produc-ing cell lines that were derived by culture of African Burkitt tumor tissue or, in the case of W91, by
infection of marmoset lymphocytes with
tumor-de-rivedvirus. Raji isanAfrican Burkitt tumor cell line
latentlyinfected with EBV (37). Cherry and Lamont
arelatentlyinfected cell lines that were established by
culture of lymphocytes of patients with infectious
mononucleosis (38; W. Henle and G. Henle, personal
communication). B95-8is a virus-producing cell line
derived by in vitro immortalization of marmoset
lym-phocytes with virus of infectiousmononucleosis origin
(31). IB4isaclonedlatently infected cell line obtained
by in vitro immortalization of human neonatal
lympho-cytes with B95-8 virus (24). Loukes is a human
B-lymphocyte cell line derived by culture of tumor tissue
from a patient with American Burkitt's lymphoma.
Loukesis EBV genome negative and antigen negative
(38, 49). Herpesvirus papio (HVPapio) was obtained
from culture fluids ofavirus-producing baboon
lym-phoid cell line, BA65, established from circulating
lymphocytesof a normal baboon, Papiocynocephalus
(14,18).
Preparation of RNA. Cytoplasmic poly(A) RNA
from B95-8orIB4cells was isolated bytwocycles of
oligodeoxythymidylate [oligo(dT)] chromatography
(22, 49). RNAs examinedby Northern blot analysis
weredenatured at60°Cin50%oformamide and 2.2 M
formaldehyde, electrophoresedin
formaldehyde-agar-osegels, and transferred to nitrocellulose filters (22,
48, 49). Nuclear RNAwas isolatedfrom IB4 cellsas
describedpreviously (2).
Recombinantplasmid. pDK225is a pBR322
recom-binantplasmidcontaining the BamHI K fragment of
the B95-8 strain of EBV (5). The Sall F fragment of
B95-8 viral DNAcontained in theplasmidpDK153 (5)
was subcloned into the plasmid pKH47 (16). The
subcloneisplasmidVRD1.
S1nucleaseanalysis ofIB4 RNA. VRD1 DNAwas
linearized withEcoRIrestriction endonuclease,
dena-tured and strand separated by chromatography on
oligodeoxyadenylate and oligo(dT) columns as
de-scribed previously (16). Unlabeled strand-separated
DNA(50to100ng)washybridized at 50°C for 3 h to 50
,ugof IB4cytoplasmic poly(A) RNA or yeast RNA in
10,ul ofasolution containing50%formamide, 0.4 M
NaCl, 40 mM PIPES
[piperazine-N,N'-bis(2-ethane-sulfonic acid)] (pH 6.4), 1 mM EDTA, and 0.1%
sodium dodecyl sulfate. The hybridization solutions
werediluted into 0.3 ml ofS1 buffer, and the
single-stranded regions of the DNA were digested at 37°C
withS1 nuclease (3). RNA was destroyed by
incuba-tion in 0.1 MNaOHat70°C. The
RNA-complemen-tary DNA waselectrophoresedonalkaline1%agarose
gels (29), transferred tonitrocellulosefilters, and
hy-bridized to nick-translated probes (40) as described
below.
Restrictionendonudeases,gelelectrophoresis,
radio-activelabeling of DNA,andhybridizationto
nitroceliu-lose filters. Restriction enzymes were obtained from
Bethesda ResearchLaboratoriesorNewEngland
Bio-labs. Viral DNAs or recombinant DNA fragments
werelabeled by nicktranslation with[a-32P]dCTP(500
Ci/mmol)(Amersham Corp.) forhybridizations to
ni-trocellulose filters. The specific activity of the
[I2P]-DNAs
was 1 x 108to3 x 108cpml,ugofDNA.Forhybridization, nitrocellulosefilters were
preincu-batedfor 1 h in lOx Denhardthybridization solution
containing 0.2% polyvinylpyrrolidone, 0.2% bovine
serum albumin, and 0.2% Ficoll (9) and hybridized
with 32P-labeled probes in lx Denhardt solution as
described in the figure legends. The filters were
washed after hybridization at 65 to 70°C in 0.2%
sodium dodecyl sulfate anddecreasingconcentrations
of SSC (2, 0.5, and 0.1x SSC;1xSSCis 0.15 MNaCl
plus 0.015 M sodiumcitrate, pH 7.4) forseveral hours.
DNAfragmentswerepurifiedfor DNAsequencingby
agarose or acrylamide gel electrophoresis, 5' or 3'
labeled with 32P-nucleotide by using polynucleotide
kinase or terminal deoxynucleotidyl transferase,
re-spectively, and sequenced by chemical degradation
(28). Determination of the DNA sequence from the
middle of theregion inFig.1Crequiredtheresolution
of sequence ladders up to 475 nucleotides from a
labeled endon5%or6%polyacrylamide gels.
RESULTS
Mapping and sequenceoftheHinflfragment.
pDK225 is a pBR322-BamHI K recombinant
plasmid from a BamHI recombinant fragment libraryof EBV B95-8 (5). The size of the BamHI Kfragment in B95-8 viralDNAand in pDK225 is 5.0 kb. The positions of the Hinfl, SmaI, HindIII, and PstI sites in pDK225 were deter-mined by partial restriction endonuclease map-ping (44). Fig. 1B isamapof BamHIfragmentK in the orientation corresponding to that of the EBVgenome (Fig. 1A).
The nucleotide sequence ofa1,153-bp Hinfl subfragmentofpDK225BamHIfragmentKwas
determined by chemical degradation of end-labeled DNA strandsbyusingthe strategy indi-cated in Fig.1C(28).The sequence is presented 5' to 3' in Fig. 2, in the same orientation as in Fig. 1.Theorientationwasverifiedby correlat-ing the SmaI restriction enzyme cleavage site within theHinflsequence(Fig.2)withtheSmaI sitesmappedin BamHIfragmentK(Fig. 1).The 1,153-bp Hinfl fragment hasabasecomposition of 69.5 mol% guanosine plus cytosine. The strand shown in Fig. 2 contains 54.9% guano-sine,26.3%adenosine,14.6%cytosine,andonly 4.2% thymidine. Several kinds ofrepeated
re-gionsare present in thefragment. Most striking is a region at nucleotides 290 to 979 which consists of direct repeats of a hexanucleotide
VOL. 44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
314 SANTEN, AND KIEFF
H1nfl
ENDI END3
AATCATGTCT GACGAGGGGCCAGGTACAGG ACCTGGAAAT GGCCTAGGAG AGAAGGGAGA CACATCTGGA CCAGAAGGCT CCGGCGGCAG TG2GACCTCAA 100
ENDIENDi
AGAAGAGGGG GTGATAACCA TGGACGAGGA CGGGGAAGAG GACGAGGACG AGGAGGCGGA AGACCAGGAG CCCCGGGCGG CTCAGGATCA GGGCCAAGAC 200 HpaII AvaIl 3 2 3 2 3 - 3 1 2 3
ENDI 3' SS
ATAGAGATGG TGTCCGGAGA CCCCAAAAAC GTCCAAGTTG CATTGGCTGC AAAGGGACCC ACGGTGGAAC AGGAGCAGGA GCAGGAGCGG GAGGGGCAGG 300 2 3 1 2 3 2 3 3 1 2 3 2 3 3 1 2 3 1 2 3 1 2 3 1 2 3 2 3 3 1 2 3 2 AGCAGGAGGG GCAGGAGCAG GAGGAGGGGC AGGAGCAGGA GGAGGGGCAG GAGGGGCAGG AGGGGCAGGA GGGGCAGGAG CAGGAGGAGG GGCAGGAGCA
400 3 3 1 2 3 1 2 3 1 32231233 23 322 1 2 3 2 3 3 1 2 3 1
GGAGGAGGGG CAGGAGGGGC AGGAGGGGCA GGAGCAGGAG GAGGGGCAGG AGCAGGAGGA GGGGCAGGAG GGGCAGGAGC AGGAGGAGGG GCAGGAGGGG
500
2 3 1 2 3 2 3 3 1 2 3 233 1 2 3 1 2 3 2 3 3 1 2 3 1 2 3 1 2 3 2 3 CAGGAGGGGC AGGAGCAGGA GGAGGGGCAG GAGCAGGAGG AGGGGCAGGA GGGGCAGGAG CAGGAGGAGG GGCAGGAGGG GCAGGAGGGGCAGGAGCAGG
600
3 1 2 32 2 3123 1 2 3 1 2 3 2 3 1 2 3 2 33 1 2 3 123 1 2 3 2 AGGAGGGGCA GGAGCAGGAG GGGCAGGAGG GGCAGGAGGG GCAGGAGCAG GAGGGGCAGG AGCAGGAGGA GGGGCAGGAG GGGCAGGAGG GGCAGGAGCA
700
3 1 2 32 3 1 2 3 2 3 1 3 22 3 1 2 3 1 2 3 2 3 1 2 3 1 2 3 2 3 1
GGAGGGGCAG GAGCAGGAGG GGCAGGAGCA GGAGGGGCAG GAGCAGGAGG GGCAGGAGGG GCAGGAGCAG GAGGGGCAGG AGGGGCAGGA GCAGGAGGGG
800
2 3 1 2 3 2 3 3 1 2 3 1 2 3 2 3 3 1 2 3 1 2 3 2 3 1 2 3 1 2 3 2 3 1 CAGGAGGGGC AGGAGCAGGA GGAGGGGCAG GAGGGGCAGG AGCAGGAGGA GGGGCAGGAG GGGCAGGAGC AGGAGGGGCA GGAGGGGCAG GAGCAGGAGG
900
2 3 1 2 3 2 3 2 3 1 2 3 2 3 3 1 2 3 2 3 1 2 3 2 3 HpaIl
GGCAGGAGGG GCAGGAGCAG GAGGGGCAGG AGGGGCAGGA GCAGGAGGAG GGGCAGGAGCAGGAGGGGCA GGAGCAGGAG GTGGAGGCCG GGGTCGAGGA
2a 2b
ENDi
2c ENDI
GGCAGTGGAG GCCGGGGTCG AGGAGGTAGT GGAGGCCGGG GTCGAGGAGG TAGTGGAGGCCGCCGGGG1
3b
GTGAAAGAGC CAGGGGGAGA GGTCGTGGAC
HInfI END3 GTGGAGAAAA GAGGCCCATG ANT
FIG. 2. Nucleotidesequenceof the1,153-bpHinflfragment.Thesequenceispresentedinthepolarity5'to3'
intheorientation of the map shown in Fig. 1. HpaIIandAvaIIrestriction enzymesitescleaved togenerate
fragments usedashybridization probesinthisstudyareindicated.Superscripts 1, 2,and 3areshownabove the
nucleotide triplets GGG, GCA,andGGA,respectively,atpositions272to979.ThisregionistheIR3repeated
sequencein EBV DNA.Repeatunits(2 3)and(233)areunderlinedinthesequenceandareinterspersedwith anotherrepeatelement, (1 2 3)n,where nvaries from 1 to4. Direct repeatregions areindicatedin boxes at
positions981to1068 and 1076to1123. Basepairmismatches in therepeatedsequencesareshownbyanasterisk
above the mismatched nucleotide (see text).Termination codonsareindicated in the first (END1)and third
(END3) translational reading frames;thesecondreadingframeiscompletelyopen.Apotential3'splicesiteisat
position 219.
andtwononanucleotidesequences.These oligo-nucleotides arecomposed of only three triplets:
GGG, GCA, and GGA. A similar region at
nucleotides 272to286 isseparated from the long tripletarrayby thetriplet GCG atpositions 287
to289. The entirerepeated regionatnucleotides 272 to 979 is designated IR3. To indicate the formatof therepeatswithinIR3, the numerical designations 1, 2, and 3 arearbitrarily assigned
to the triplets GGG, GCA, and GGA,
respec-tively, andareindicatedassuperscriptsoverthe
corresponding triplets in Fig.2(within the limits of therepeated region only). The general struc-tureof therepeatcanberepresentedas(1 2 3),
where nvariesfrom 1 to4, interspersedwith (2
3) or(23 3). The elements (2 3)and (2 3 3 )are
underscoredtoindicate thepatternof intersper-sion of theseoligonucleotidesamongthetandem
repeats of the (1 2 3) nonanucleotide. A 6-bp
sequence, 5'-GGTGGA-3', is repeatednearthe
left andrightlimitsofIR3(positions263 to268
and980to985,respectively).
Two other classes ofdirectrepeatsareinthe
Hinfl sequence. They are indicated as boxed
regions in Fig. 2. One class consists of three
tandemrepeats ofa24-bp sequence (sequences 2a, 2b, and 2c) at positions 994 to 1,068. The
prototypeformissequence2b. A transition from
TtoCoccursatposition 1,003incopy2a,anda
3-bp insertion, GCC, is present at positions 1,062 to 1,064 in repeat2c (indicated by
aster-isks above the mismatched nucleotides). An
*** END1i END1I
TA GAGGA
3a
kCGTGAAAGAGCCAGG GGGGGAAGTC
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.63.430.79.397.2]SIMPLE REPEAT SEQUENCE IN EBV DNA 315
incompletecopyof thisrepeat sequence occurs between IR3 and 2aatpositions 981to993. The second class is composed oftwo tandem 24-bp sequences (sequences 3a and 3b) at positions 1,076 to 1,123(nonidentical basesare indicated by asterisks).
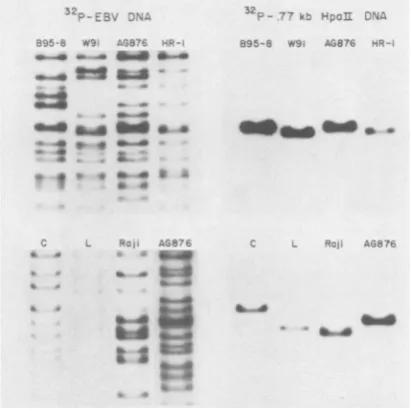
Conservation and variability of the IR3 region. Since IR3 has homology with a component of human and mouse cell DNAs (Heller et al., in press),it ispossible that IR3 has beenacquired by B95-8 virusthrough recombination with cell DNA. SomeEBVisolateswould then be expect-edtobe lackingIR3 ormight incorporateit into adifferent site. Analternative model is that IR3 isa componentofthe EBV genomefrom various sources. To determine whether the IR3 se-quence is a common feature in other EBV isolates, the 32P-labeled HpaIIfragment, which contains 57 nucleotides to the left of IR3, 708 nucleotides of IR3, and 11 nucleotides to the right ofIR3 (Fig. 1C and 2), washybridized to nitrocellulose filters containing the separated BamHI fragments ofthe W91,AG876, P3HR-1, Raji, Cherry, or Lamont isolates ofEBV (see above for adescription of the origins of these EBVstrains). The right panels of Fig. 3 show the results of this hybridization. The left panels present aportion of the sameblots whichwere rehybridized with32P-labeledEBV DNA to indi-cate all of the viral DNA fragments. The IR3 probe identifies a single fragment in all of the EBVstrains. The 32P-labeledAvaII-HpaII frag-ment(IR3plus 16 bptotheleft and11bptothe right of IR3 [Fig. 1C and 2]) and a pDK225 BamHI K probe also hybridize to the same fragments on other Southern blot strips of BamHIfragments ofeachofthe EBV DNAs (17 and data not shown). In each instance, the extentof hybridization oftheIR3-specific probe tothesingleBamHI Kfragmentwas proportion-al to the amount of viral DNA on the blot as judged from the hybridization with 32P-labeled EBVDNA.The results indicatethat much or all of the IR3 sequence is conserved within the BamHI Kfragment in each of these isolates.
The BamHI Kfragments vary insize among theEBVisolates(Fig. 3)asfollows: Cherry (5.2 kb) >AG876 (5.0 kb)> B95-8(5.0kb)> P3HR-1 (4.9kb)> W91(4.8 kb) > Lamont(4.8kb) >
Raji (4.7 kb). This variability reflects either differences among EBV isolates in the unique-sequence portions of BamHI fragment K or heterogeneity within the IR3 repeat sequence. Heterogeneity in the IR3 region might occur through intramolecular or intermolecular un-equal homologous recombination within the re-peat. Thevariability of IR3among EBV DNAs was investigated by hybridizing the 32P-labeled HpaII fragment (described above) to Southern blotsofthe separatedHpaIIrestriction enzyme
tW.-4_
-4_
.. Ibm... ...a
4b. * 44
.-F ' m
-7
3- 7 K u-,A
89 5- 8 019, A-,876 "
.! A".jR.7F
W.46
_-_>k....A
,A
'4..
FIG. 3. Conservationof IR3 sequences in DNAs of
different EBV strains. DNAs from virus (B95-8, W91, AG876, and P3HR-1) or from nonpermissively
infect-ed cells (Cherry [C], Lamont [LI, and Raji) were
digested with BamHI, electrophoresed on agarose
gels, andtransferred to nitrocellulose filters. The right
panels show the resultsof hybridization(in 6x SSC at
72°C) ofa32P-labeled,0.77-kbHpaII IR3 DNAprobe
to the filters. The left panels show the result of
rehybridization of the same filters to a 32P-labeled
EBV DNA probe. Only the BamHI K region of the
filters is shown.
fragments of EBV DNAs purified from virus, infected cells, or recombinant plasmids. The results established thatvariability in the size of IR3 does occur among isolates, but that IR3 is stableamongtheDNAmolecules of each isolate (Fig.4). TheIR3-HpaIIfragmentsof B95-8 viral DNA and the recombinant derivative pDK225 are0.77 kb. The fragments ofW91 viral DNA and therecombinant EcoRIBcosmid derivative are 0.68 kb, whereas the AG876 and P3HR-1 viral and the Lamont intracellular IR3-HpaII fragments are 0.80, 0.70, and 0.64 kb, respec-tively. The observed range of variation in the size of the HpaII fragment containing IR3 is about 0.16 kb (AG876 versus Lamont [Fig. 4]) and accounts for most or all ofthe variability betweenthe BamHI Kfragments ofthese DNAs (Fig. 3). Thedifference in size of theBamHI K fragmentsin theCherry and RajiDNAssuggests that more extensivevariation inthe IR3 region may occur.
Conservation of IR3 in another herpesvirus. EBV and HVPapio are B-lymphotrophic pri-mate herpesviruses whichare endemic in their respective host species, i.e., humans and ba-boons(13-15).Fortypercentof the sequencesof
VOL.44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.253.459.77.283.2]Restriction enzyme maps of the BamHI K
re-gion of EBV DNA and the colinear rere-gion of HVPapio DNAareshown inFig. 5. The hybrid-ization results established that HVPapio DNA containsa sequencerelatedtoIR3 whichmaps
in EcoRI fragment G and XbaIfragment H and is colinear with EBV BamHI fragment K (Fig. 5).
m
.VPaPIO
-~a Y i:. C1S fl N
a a
I
D- Ua
x,-S3 a
H- 0
S
a
FIG. 4. Variability in the size of IR3 among EBV isolates. DNAs from (left to right) recombinant
plas-mid pDK225 containing the B95-8BamHIK fragment,
B95-8 virus, a recombinant cosmid containing W91
EcoRI fragment B, W91 virus, P3HR-1 virus, AG876 virus, AG876 transformant (a cell line of neonatal lymphocytes transformed in vitro by AG876 virus),
and the nonpermissively infected cell line Lamont
were digested with HpaII restriction enzyme. The
resultof hybridization of a32P-labeled,0.77-kbHpaII
probe in 6x SSC at 72°C to a Southern blot of the
separated HpaII fragments is shown. The
hybridiza-tion to higher-molecular-weight fragments in the
AG876 transformant and Lamont cell DNAs is due to partial resistance of these methylated DNAs to cleav-agebyHpaII.The ethidium bromide-stained gel insert
showsadenovirus type 2 EcoRI and pBR322
BamHI-Hinflfragment size markers.
EBV and HVPapio DNAs are well conservec
between the viruses (13, 15, 19). The
homolo-goussequencesarecolinear among the genomes and aredispersed alongalmost the fulllengthof the DNAs(20, 26). Thecolinear relatedness and common DNA organization indicate that these primateviruses evolvedfroma common
ances-tor. An IR3 AvaII-HpaIIprobewas hybridized to Southern blots of the separated restriction
enzyme fragments ofHVPapio DNA to deter-mine whetherahomologoussequence is present at a colinear point in this primate genome.
0
N'- a
B.
-LvPcpo
G
H
A M LE EcoRI
D XboI
B
U/__>._1 /-;t-"B- -nBomHl
FIG. 5. IR3-related sequences in another
herpesvi-rus. (A) Identical blot strips of viral DNA from the
relatedprimateherpesvirus HVPapiohybridized in 6x
SSC at 720C to 32P-labeled viral DNA, pDK225 BamHI fragment K, and AvaII-HpaII IR3 probes.
Viralfragments which hybridized to labeled pDK225
or IR3 are indicated. Designations of the fragment
namesandtheir map locations inHVPapioDNAhave
beendescribedpreviously(18, 20,26). (B) EBV DNA
BamHl K region and the colinear region from
HVPapioDNA.Thearrowsbelow the EBVDNAmap
indicate the location of the Sall Ffragment in viral
DNAandplasmid VRD1.
.1-"... .,.r .-.
rn) Ct:
a
Ak
", f..
4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.496.52.242.79.359.2] [image:6.496.257.444.159.550.2]SIMPLE REPEAT SEQUENCE IN EBV DNA 317
Transcription of ER3. Since a 3.7-kb poly(A) cytoplasmic RNA is transcribed from the BamHI Kfragment in latently infected, growth-transformed IB4 cells (24, 49), we investigated whether IR3 encodes part of this RNA. 32P_ labeled IR3 DNA hybridized to the 3.7-kb IB4 RNA on Northernblotsand also identified 5.8-and 4.5-kboligo(dT)-selected IB4 nuclearRNAs (Fig. 6A). The direction of transcription ofthe 3.7-kb IB4 RNA and thepotential slicing ofthis transcript in the IR3 regionwereexamined by Si nuclease analysis (3) of strand-separated DNA hybridized to IB4 RNA. The 7.9-kb Sall F fragment of EBV DNA (62 to67 megadaltons) which contains IR3was subclonedinto plasmid pKH47, and the separated DNA strands were isolated (16). Eachseparated strandwas hybrid-ized to IB4 cytoplasmic poly(A) RNA. After digestion with Si nuclease, the RNA-comple-mentary DNA waselectrophoresed on denatur-ingagarosegels and transferredtonitrocellulose filters. IR3 and SalI F probes hybridize to a
2.0-kb RNA-complementary fragmentfrom the SalI F strand opposite in polarity to the
se-quence shown in Fig. 2. Neither the SalI F nor
the IR3 probe hybridized to blotsfrom experi-ments done with the SalI F strand identical in polarity tothe sequence shown in Fig. 2 (data notshown). Thus, the 3.7-kbRNA is transcribed from lefttoright inEBVDNAand contains the IR3sequence(Fig. 2). Since Northern blot anal-ysis showed that thelatently infected cell RNA is 3.7 kb whereas the RNA-protected DNA segmentisonly 2.0kb, the datasuggest that the cytoplasmic poly(A) RNA encoded by IR3 is spliced. Nootherfragmentsweredetectedwith the SalI F probe. Therefore, either multiple splicing events join small exons from Sall F DNA which total 1.7 kb in size, orpart ofthe 3.7-kb RNA is transcribed from DNA which maps outside of SalIfragmentF.
Low-abundance cytoplasmic poly(A) 2.2-, 1.9- and 1.5-kb RNAs are expressed from the BamHIKregioninproductivelyinfected B95-8 cells (22). The IR3 probe did not hybridize to these RNAsonNorthernblots, but dididentify previously undetected 2.4- and 1.0-kb RNAs (Fig. 6C).
The RNAs detected in B95-8 and IB4 cells with the IR3 probe represent viral transcripts and are not cellular RNAs which might be identified due to homology between the IR3 sequence and cellular DNA. The pDK225 BamHI K probe did not identify any RNA on Northern blots of poly(A)RNAfrom Loukes,an EBV genome-negative B-lymphocyte cell line (49). Also, labeled cDNA made from Loukes cytoplasmicRNAdidnothybridizetoSouthern blots of viral or recombinant DNA fragments containingthe IR3region (datanot shown).
DISCUSSION
IR3 and a closely related sequence are con-servedat acolinear position in the genomes of HVPapioand all EBVisolates examined. At its simplest level of organization, IR3 is a tandem repeat array composed of only three nucleotide triplet elements: GGG, GCA, and GGA. The triplets are further organized as a hexanucleo-tidesequence, GCAGGA (H), and two
nonanu-A. IB4 POLY B. 52P-IR3 2P-SaII F C 895
aRNA i. A RN
CYTO NUT CYTr
_ CY.To_
>-t a.
-745
-
U.6
~-i.45
-4.0
-- .92
-POLY
A
O
-.51
-FIG. 6. Transcription of IR3 sequences in B95-8
and IB4 cells. (A) Northern blots(48) of cytoplasmic
and nuclear poly(A) RNA from nonpermissively
in-fected IB4 cells hybridized to a 32P-labeled
AvaIl-HpaIIIR3probe.Hybridizations were done at 52°C in
5x SSC and50%oformamide. The positions of 5.2-kb
(28S) and2.0-kb (18S)rRNAsize markersare shown
(>). (Theband which isslightly smaller than the 4.5-kb
nuclear RNA is an artifact anddid not hybridize to the
IR3 probe in several other hybridizations.) (B) S1
nucleaseanalysis of RNA from IB4 cells. Unlabeled
separatedDNA strands of a Sall F-pKH47
recombi-nant plasmid, VRD1 (62 to 67 megadaltons in EBV
DNA [Fig. 1]), were prepared by chromatography of
denatured linearized plasmid DNA on
oligodeoxya-denylate and oligo(dT) columns. A 50-ng amount of
the strand complementary to the sequence shown in
Fig. 2 was hybridized to 50 ,ug of IB4 cytoplasmic
poly(A) RNAoryeast RNA, digested withSi
nucle-ase,incubated in NaOH, electrophoresed on
denatur-ing agarose gels, and transferred to nitrocellulose.
Identical blots of theRNA-complementary DNA were
hybridized to32P-labeledIR3 andSall Fprobesin 6x
SSC at 72°C. The background hybridization near the
topof the blots probably represents reassociatedSall
F DNA formed due to slight contamination of the
RNA-coding strand with the complementary strand.
Size markersaresingle-strandedHinflfragments from
the pDK286 EBV recombinantplasmid (5). (C)
Auto-radiogram of a Northern blot of oligo(dT)-selected
cytoplasmic RNA from permissively infected B95-8
cells which was hybridized to an IR3 probe as
de-scribedfor (A).
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.496.257.451.194.371.2]cleotide sequences, GCAGGAGGA (Ni) and
GGGGCAGGA (N2). Atahigher level of
organi-zation, H or Ni sequences are interspersed
between N2 sequences ortandemcopies of N2.
There is no apparenthigher order ofsymmetry
orrepeat unit in IR3. IR3probably has evolved into an extended repeated region as a conse-quenceof unequalcrossoverwhichexpandedan
ancestralsetofsequencesrelatedtoH, Ni, and N2 (43).The polymorphism in size of IR3among
EBV isolates probably has been generated by this samemechanism. Thepreservation of
sym-metrythrough therepeatarraycould reflect the tendency of unequal crossoverto maintain the
fundamental repeat pattern.
Repeat sequences, such as IR3 and the two
tandem24-bprepeats, could be recognition sites for a protein. The origin and function of IR3
might be predicted bysequencerelatedness with
a known genetic element. We compared the
nucleotidesequence ofIR3 toeucaryotic cellu-lar and viral nucleotide sequences recorded at
the Nucleic Acid Sequence Database (8). Ran-dom perfect 9 to 12-bp matches were found in many sequences, including coding and
noncod-ing regions of mRNA and interspersed and mid-dlerepetitive (Alu family) elements (23).
Homol-ogyof theseother DNAstoIR3 does notextend beyond 12 bp since therelatedsequences arenot
tandemly repeated.
Other simple repeated sequences have been
describedpreviously. Examples includetandem
5-bp sequences in the recombinational class
switch region of immunoglobulin heavy-chain
genes (33); tandem repeats in the terminal a
sequence ofherpes simplex virus type 1 DNA
which mediates site-specific inversion events
(32); repeated dinucleotide arrays inintron 2 of
human gamma globin genes (42), in the
inter-genic spacer of sea urchin histone DNA (47),
and itn an "evolutionarily conserved" repeated
sequence family (30); an irregular repeat se-quence(CnT)matthetermini of linear extrachro-mosomal rDNA in Dictyostelium discoideum (11); and polymorphic tandemarrays of related 14to 15-bp sequencesin the5'-flanking region
of the human insulin gene(1). Unlike IR3,none
of these simple repeats encodes cytoplasmic RNA.
Northern blothybridizations showed thatthe
IR3 region encodes different cytoplasmic poly(A) RNAs in permissively infected B95-8
cells and in latently infected IB4 cells. In IB4 cells, IR3 is part ofa 2.0-kb exon, transcribed
left to right, which encodes part ofthe 3.7-kb
RNA identified on Northern blots (49; Fig. 6). IB4cell 5.8- and 4.5-kb nuclearpoly(A) RNAs were identified by an IR3 probe (Fig. 6). The
larger nuclear RNAs may be precursors ofthe
3.7-kb cytoplasmic poly(A)RNA. Sincewe
ex-amined onlysteady-state RNAin IB4 cells, our analysis cannot establish processing relation-shipsamongtheseRNAs. The DNAsequence in Fig. 2, correspondingto the RNA strand of the 2.0-kb exon, has a
Wor
potential splice accept-orsite, 5'-TGTCCGGAG-3', whichhas one mis-matched nucleotide (indicatedby the asterisk), compared with theconsensus sequence 5'-PyN-PyPyPyNPyAG-3' (41). Sites corresponding to theconsensus sequencesfortranscription initia-tion (M. Goldberg, Ph.D. thesis, Stanford Uni-versity, Stanford,Calif. 1979) and polyadenyla-tion(36) are not present. Itisnot knownwhether thesplice site is utilized in transcriptional proc-essing of viral RNA in IB4 cells. Termination codons occur in two of the three translational reading frames in the putative intron preceding the splice site (Fig. 2). The remaining reading frame has no stop codons through the entire Hinfl sequence.Itis likely that the 3.7-kb cytoplasmicpoly(A) RNAis translated intoaprotein.Polyribosomal poly(A)RNA(presumably mRNA) is expressed from the sameregion ofEBV DNA in IB4 cells and in latently infected cell lines established from Burkitt tumors (24, 25, 35). If the IR3 sequence in the 3.7-kb RNA is translated into protein, the nucleotide sequence (Fig. 2)in the three possible reading frames predicts a simple coding domain of more than 200 amino acid residues composed ofonly (i) serine, arginine, and glycine; (ii) glycine and alanine; or (iii) glutamine, glutamate, and glycine.Thevariation in length of IR3 among EBV isolates predicts that the putative translation products of IR3 mRNAs would correspondingly differin size.
Growth-transformed cells latently infected with EBV contain a nuclear antigen, EBNA (39). EBNA varies in sizeamongdifferent latent-ly infected cell lines, and the relative sizes of EBNA in Namalwa, B95-8, P3HR-1, and Raji cells(46) correlate with the relative sizesof IR3 in each of these viralDNAs.Therefore,IR3 may encodepart ofEBNA. It should be noted that variation intheIR3tripletarraywhichpreserves the repeat pattern (expected from an unequal crossover mechanism) will maintain a unique translationalreading frame for codons3' toIR3. It is of interest in this regard that the single divergent region in IR3 at positions 287 to 289 (Fig.2)preservestripletsymmetryandtherefore the translational reading frame. Genetic and protein polymorphisms occurwithin the repeti-tivesequencesoftheglycine-alanine-serine sim-pleamino acid-coding domain of silk fibroin(27, 45). Alleles of silk fibroin differ in length as a consequence of unequal recombination ex-changes withinthe repetitive coding domain of the gene. These alleles encode fibroins which differcorrespondingly in size.
on November 10, 2019 by guest
http://jvi.asm.org/
SIMPLE REPEAT SEQUENCE IN EBV DNA 319 ACKNOWLEDGMENTS
We thank Victor Raczkowski for subcloning the Sall F
fragment from pDK153 into pKH47. We also thank Mary Hummel forthegenerousgift of nitrocellulose filters contain-ing B95-8 cytoplasmic poly(A) RNA. Chris Beiseland Susan Fennewaldwerehelpful in thecomputeranalysisof the DNA
sequence.
Thiswork wassupported by Public Health Servicegrants
CA17281 and CA 19264from the National Institutes ofHealth andbygrantMV 32 G from the American Cancer Society. V.V.S. isapredoctoraltrainee(CA 09268).E.K. isafaculty research awardee of the AmericanCancer Society.
LITERATURE CITED
1. Beil, G. I.,M.J. Selby,and W.J.Rutter. 1982. Thehighly polymorphic region nearthe human insulingeneis
com-posed of simple tandemly repeated sequences. Nature (London)295:31-35.
2. Berk,A. J.,F. Lee,T.Harrison, J. William,and P. A. Sharp. 1979. Pre-early adenovirus 5geneproduct
regu-lates synthesis of early viral messenger RNAs. Cell 17:935-944.
3. Berk,A.J.,and P. Sharp. 1977. Sizingandmappingof early adenovirus mRNAs by gel electrophoresis ofS1 endonuclease-digested hybrids.Cell 12:721-732. 4. Cheung, A.,and E. Kieff. 1981.Epstein-Barrvirus DNA.
X. Direct repeat within the internal direct repeat of Epstein-Barrvirus DNA. J. Virol. 40:501-507.
4a.Cheung, A.,and E. KIeff. 1982.Longinternal directrepeat inEpstein-Barrvirus DNA.J. Virol. 44:286-294. 5. Dambaugh, T., C. Beisel, M. Hummel, W. King, S.
Fennewald,A.Cheung,M.Heller,N.Raab-Traub,and E. Kieff. 1980. Epstein-Barr virus DNA. VII. Molecular cloningand detailedmappingof EBV(B95-8)DNA. Proc. Natl. Acad. Sci. U.S.A. 77:2999-3003.
6. Dambaugh, T.,M. Heller, N.Raab-Traub, W. King,A. Cheung, C. Beisel, M. Hummel, V. van Santen, S. Fennewald, and E. Kieff. 1980. DNAs of Epstein-Barr virus andherpesviruspapio,p.85-90. In A.Nahmias,W. Dondle, and R.Schinazi(ed).,Thehumanherpesvirus. Elsevier/NorthHollandPublishing Co.,New York. 7. Dambaugh, T.,N.Raab-Traub,M.Heller,C.Beisel,M.
Hummel,A.Cheung,S.Fennewald,W.King,and E. Kieff. 1980. Variations among isolates ofEpstein-Barr virus. Ann. N.Y. Acad. Sci. 602:711-719.
8. Dayhoff,M.O.,H. R.Chen,L. T.Hunt,W.C.Barker, L.-S.Yeh,D.G.George,and B. C. Orcutt. 1981. Nucleic acidsequencedatabase,version 13. National Biomedical Research Foundation, Georgetown University Medical Center, Washington, D.C.
9. Denhardt,D. T. 1966. A membrane-filtertechniquefor the detection ofcomplementary DNA. Biochem. Biophys. Res.Commun. 23:641-646.
10. Dolyniuk, M.,R.Pritchett,and E. Kieff. 1976. Proteins of Epstein-Barr virus. I. Analysis of the polypeptides of
purified enveloped Epstein-Barr virus. J. Virol. 17:935-949.
11. Emery, H. S., and A. M. Weiner. 1981. An irregular satellite sequence is found at the termini of the linear extrachromosomal rDNA in Dictyostelium discoideum.
Cell 26:411-419.
12. Epstein,M.A.,and B.Achong (ed.).1979. The Epstein-Barr virus.Springer-Verlag, Berlin.
13. Falk, L., F. Deinhardt, M. Nonoyama,L. G.Wolfe, C. Bergholz,B. Lapin, L. Yakovleva, V. Agrba,G. Henle, andW. Henle. 1976.Propertiesofababoonlymphotropic herpesvirusrelatedtoEpstein-Barrvirus. Int. J. Cancer 18:798-807.
14. Gerber, P., S. S. Kalter,G. Schidlovsky,W.D.Peterson, Jr., and M. D. Daniel. 1977. Biologic and antigenic characteristics of Epstein-Barr virus-related
herpesvi-rusesofchimpanzeesandbaboons.Int.J. Cancer
20:448-459.
15. Gerber, P.,R.F.Pritchett,and E. D. Kieff. 1976. Antigens
and DNA ofachimpanzee agent relatedtoEpstein-Barr virus. J. Virol. 19:1090-1099.
16. Hayashi, K. 1980. Acloning vehiclesuitable for strand separation. Gene 11:109-115.
17. Heller, M., T. Dambaugh, and E. Kieff. 1981. Epstein-Barr virus DNA. IX.Variation among viral DNAs from producer and nonproducer infected cells. J. Virol. 38:632-648.
18. Heller, M., P. Gerber, and E. Kieff. 1981. Herpesvirus papio DNA is similar in organization to Epstein-Barr virus DNA.J.Virol. 37:698-709.
19. Heller, M., P. Gerber, and E. Kieff. 1982. The DNA of herpesvirus pan, a third member of the Epstein-Barr virus-herpesviruspapio group. J. Virol. 41:931-939. 20. Heller, M., and E. Kieff. 1981. Colinearity between the
DNAs of Epstein-Barr virus and herpesvirus papio. J. Virol. 37:821-826.
21. Hinuma, Y., M. Konn, J. Yamaguchi, D.J. Wudarski, J.R.Blakeslee, Jr.,andJ.T.Grace, Jr. 1967. Immunoflu-orescenceand herpes-typevirusparticlesin the P3HR-1 Burkittlymphomacell line. J. Virol. 1:1045-1051. 22. Hummel, M., andE.Kieff. 1982.Epstein-Barrvirus RNA.
VIII. Viral RNA inpermissively infected B95-8 cells. J. Virol. 43:262-272.
23.Jelinek, W. R., T. P. Toomey, L.Leinwand,C. Duncan, P. A.Biro,P. V.Choudary, S. Weissman, C. M.Rubin, C. M.Houck,P. L.Deininger,andC.W. Schmid. 1980. Ubiquitous, interspersed repeatedsequencesin mammali-angenomes.Proc. Natl. Acad.Sci. U.S.A. 77:1398-1402. 24. King, W., A. L. Thomas-Poweil, N. Raab-Traub, M. Hawke,and E.Kieff.1980. Epstein-Barrvirus RNA.V. Viral RNA in arestringently infected, growth-transformed cell line. J. Virol. 36:506-518.
25. King, W., V. van Santen, and E.Kieff. 1981.Epstein-Barr virus RNA. VI. Viral RNA inrestringently and abortively infectedRaji cells. J. Virol. 38:649460.
26. Lee, Y.S.,M.Nonoyama, andH. Rabin.1981. Colinear relationshipsofherpesviruspapio DNAtoEpstein-Barr virus DNA. Virology 110:248-252.
27. Lizardi,P. M.1979.Geneticpolymorphism ofsilkfibroin studiedbytwo-dimensional translation pausefingerprints. Cell 18:581-589.
28. Maxam,A. M., andW. Gilbert. 1980.Sequencing end-labeled DNA with base-specific chemical cleavages. MethodsEnzymol.65:499-560.
29. McDonnell,M.W., M. N.Simon,andF. W.Studier. 1977. Analysis ofrestrictionfragments ofT7 DNA and determi-nation of molecularweights byelectrophoresisin neutral andalkalinegels.J. Mol.Biol. 110:119-146.
30. Miesfeld, R.,M.Krystal,and N. Arnheim.1981. A mem-berofa newrepeatedsequencefamilywhich is conserved throughout eukaryotic evolution is found between the human 8and 13globingenes.Nucleic Acids Res. 9:5931-5947.
31. Miller,G., and M. Lipman. 1973. Release ofinfectious Epstein-Barrvirusby transformed marmosetleukocytes. Proc. Natl.Acad. Sci.U.S.A.70:190-194.
32. Mocarski, E. S., and B. Rolzman. 1981. Site-specific inversionsequenceof theherpes simplexvirus genome: domain and structural features. Proc. Natl. Acad. Sci. U.S.A. 78:7047-7051.
33. Nikaido, T., S.Nakal,and T.Honjo.1981.Switchregion ofimmunoglobulinCILgeneiscomposed ofsimple tandem repetitive sequences. Nature (London) 292:845-848. 34. Pizzo,P.A.,I. T.Magrath, S. K.Chattopadhyay,R.J.
Biggar,andP.Gerber. 1978.A newtumor-derived
trans-forming strain of Epstein-Barr virus. Nature (London) 272:629-631.
35. Powell,A. L.T.,W. King,and E. Kieff.1979. Epstein-Barrvirus-specificRNA.III.Mapping of the DNA encod-ing viralRNAin restringent infection. J. Virol. 29:261-274.
36. Proudfoot,N.J., and G. G. Brownlee. 1976. 3'non-coding region sequencesineukaryotic messengerRNA. Nature (London) 263:211-214.
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
37. Pulvertaft, R. J. V. 1964. Cytology ofBurkitt's tumor (African lymphoma).Lanceti:238-240.
38. Raab-Traub, N.,T.Dambaugh,andE. Kieff. 1980.DNA ofEpstein-Barr virus.VII.Analysisandmolecular epide-miology of the "additional" DNA oftwoBurkitttumor isolates ofEBV.Cell22:257-267.
39. Reedman,B. M., and G. Klein. 1973. Cellularlocalization ofEpstein-Barr virus (EBV)-associated complement-fix-ingantigen of producer and non-producer lymphoblastoid celllines. Int. J. Cancer 11:499-520.
40. Rigby, P. W. J., M.Dieckmann, C. Rhodes,andP.Berg. 1977. Labeling deoxyribonucleic acid to high specific activityinvitro nicktranslation with DNApolymeraseI. J. Mol. Biol. 113:237-251.
41. Sharp, P. A. 1981. Speculations onRNA splicing. Cell 23:643-646.
42. SUghtom, J. C., A. E. Blechl, and 0. Smithies. 1980.
HumanfetalG-y- and Ay-globingenes:complete
nucleo-tide sequences suggest that DNA can be exchanged
between these duplicatedgenes.Cell21:627-638. 43. Smith,G. P.1976.EvolutionofrepeatedDNAsequences
by unequalcrossover.Science 191:528-535.
44. Smith,H.O.,andM.Birnstiel. 1976. Asimple method for DNArestriction sitemapping. Nucleic Acids Res. 3:2387-2398.
45. Sprague, K. V., M. B. Roth, R. F.Manning, and P. L.
Gage.1979.Alleles of the fibroingenecoding forproteins of differentlengths.Cell17:407-413.
46. Strnad, B. C., T. C. Schuster, R. F. Hopkins, R. H. Neubauer, and H.Rabin. 1981. Identification ofan
Ep-stein-Barr virus nuclear antigen by fluoroimmunoelectro-phoresis and radioimmunoelectrophoresis. J. Virol. 38:996-1004.
47. Sures, I., J. Lowry, and L. H. Kedes. 1978. The DNA
sequencesofseaurchin(S.purpuratus)H2A, H2B, and
H3 histonecodingandspacerregions. Cell 15:1033-1044. 48. Thomas,P. A.1980.Hybridization of denatured RNA and
smallDNAfragments transferredtonitrocellulose. Proc. Nati. Acad. Sci. U.S.A. 77:5201-5205.
49. vanSanten,V., A.Cheung,andE. Kieff. 1981. Epstein-Barrvirus RNA. VII. Size and direction oftranscription of virus specified cytoplasmic RNA inacellline trans-formedbyEBV. Proc.Nati. Acad. Sci. U.S.A. 78:1930-1934.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/