JOURNAL OFVIROLOGY, Sept. 2008, p. 9171–9178 Vol. 82, No. 18 0022-538X/08/$08.00⫹0 doi:10.1128/JVI.00530-08
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Functional Analysis of the Complex
trans
-Activating Response
Element RNA Structure in Simian Immunodeficiency Virus
䌤
Mireille Centlivre, Bep Klaver, Ben Berkhout, and Atze T. Das*
Laboratory of Experimental Virology, Department of Medical Microbiology, Center for Infection and Immunity Amsterdam (CINIMA), Academic Medical Center of the University of Amsterdam,
Meibergdreef 15, 1105 AZ Amsterdam, The Netherlands
Received 10 March 2008/Accepted 27 June 2008
Transcription of human immunodeficiency virus (HIV) and simian immunodeficiency virus (SIV) is acti-vated through binding of the viral Tat protein to thetrans-activating response (TAR) element at the 5ⴕend of the nascent transcript. Whereas HIV type 1 (HIV-1) TAR folds a simple hairpin structure, the corresponding domains of HIV-2 and SIVmac exhibit a more complex structure composed of three stem-loops. This structural polymorphism may be attributed to additional functions of TAR in HIV-2/SIVmac replication. We recently constructed an SIVmac variant that does not require the Tat-TAR interaction for transcription. We used this variant to study additional roles of TAR in SIVmac replication and generated mutants with a truncated TAR structure. We demonstrate that partial or nearly complete removal of TAR does not impair viral transcription, RNA processing, and translation. Moreover, these deletions do not significantly affect virus replication in the PM1 T-cell line and macaque peripheral blood mononuclear cells. These results demonstrate that the complex TAR structure in SIVmac has no other essential function in virus replication in vitro besides its role in Tat-mediated activation of transcription.
Transcription of human and simian immunodeficiency vi-ruses (HIV and SIV, respectively) is activated by binding of the virally encoded Tattrans-activator protein to thetrans -activat-ing response element (TAR) at the 5⬘ends of the viral RNAs (3, 4, 20, 26, 28). Upon Tat binding to the nascent transcript, the positive transcriptional elongation factor (pTEFb) is re-cruited to the transcription complex at the 5⬘ long terminal repeat (LTR) promoter. Consequently, the processivity of the elongating RNA polymerase II is enhanced through phosphor-ylation of its C-terminal domain by the kinase component of pTEFb (see reference 5 for a review). The assembly of new transcription complexes is also stimulated by pTEFb, through the recruitment of TATA box binding protein to the LTR promoter (27, 39). TAR thus functions as an essential RNA enhancer element to promote the switch from low basal tran-scription to Tat-activated processive trantran-scription (11, 14).
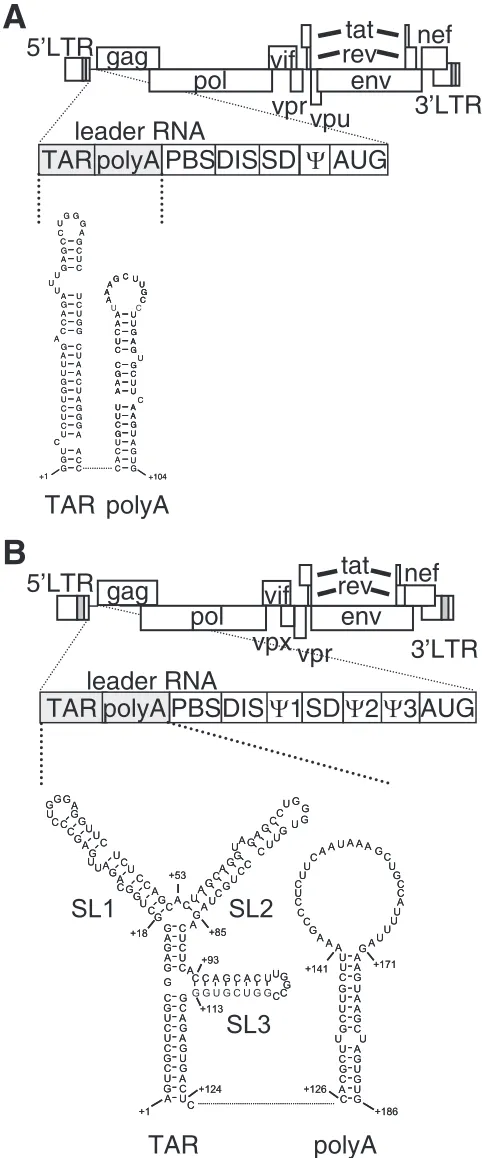
Most of the studies in which the Tat-TAR interaction was investigated were focused on HIV type 1 (HIV-1), and despite strong similarities between the trans-activation mechanisms and Tat proteins of HIV-1, HIV-2, and SIVmac, some clear differences are apparent. Whereas HIV-1 Tat can efficiently activate transcription of HIV-2 and the closely related virus SIVmac, HIV-2 Tat is a poor activator of HIV-1 transcription (28). This difference intrans-activation capacity may be a con-sequence of the structural polymorphism between HIV-1 and HIV-2/SIVmac TARs (Fig. 1). HIV-1 TAR RNA folds into a characteristic hairpin structure with a 3-nucleotide (nt) bulge and a 6-nt loop, which facilitate the binding of Tat and the cyclin T1 component of pTEFb, respectively (Fig. 1A). The
HIV-2 and SIVmac TAR elements exhibit a more complex RNA structure with three stem-loop domains (SL1 to SL3) (Fig. 1B) (8, 26). SL1 and SL2 contain a 2-nt bulge and a 6-nt loop and resemble the single HIV-1 stem-loop structure. Al-though both SL1 and SL2 are involved in Tat-mediated acti-vation of transcription (10, 25), SL1 seems to be more impor-tant than SL2 fortrans-activation (26, 40). This is possibly due to intrinsic folding kinetics of the two structures, as SL1 has a position more proximal to the transcription initiation site. Al-ternatively, the presence of a purine base in the SL2 bulge may be suboptimal for Tat binding. SL3 does not resemble the typical HIV-1 TAR structure and is not involved in Tattrans -activation (25).
The reason that HIV-2 and SIVmac have such a complex TAR structure with three stem-loop elements, whereas HIV-1 replicates efficiently with only a single TAR hairpin, is cur-rently unclear. Strong conservation of certain characteristics of the TAR structure in different HIV-2 and SIV isolates suggests that all three stem-loops are important for viral replication (8, 26). Possibly, TAR has other functions in the HIV-2/SIV life cycle in addition to its role in the activation of transcription, as previously suggested for HIV-1 (2, 5–7, 9, 13, 29). The best system with which to study additional TAR functions is that of the replicating virus. However, the analysis of TAR mutant viruses is complicated by the fact that TAR is essential for transcription. Since TAR mutations will therefore have a dom-inant negative effect on viral replication, it is difficult to study any effect on other steps in the viral life cycle.
We recently generated an SIVmac variant in which the Tat-TAR trans-activation mechanism is functionally replaced by the doxycycline (DOX)-inducible Tet-On gene regulation sys-tem (18). Transcription of this SIV-rtTA variant is not depen-dent on the Tat-TAR axis, which makes it an ideal tool to study additional TAR functions. In this study, new SIV-rtTA variants
* Corresponding author. Mailing address: Laboratory of Experi-mental Virology, Academic Medical Center, Room K3-115, Meiberg-dreef 15, 1105 AZ Amsterdam, The Netherlands. Phone: 31 20 566 3396. Fax: 31 20 691 6531. E-mail: [email protected].
䌤Published ahead of print on 2 July 2008.
9171
on November 8, 2019 by guest
http://jvi.asm.org/
were generated in which TAR was partially or completely deleted. We found that SIV-rtTA can replicate efficiently upon deletion of all three stem-loop domains of TAR, which dem-onstrates that TAR has no additional essential function in SIVmac replication in vitro.
MATERIALS AND METHODS
Construction of SIV-rtTA variants with deletions in TAR.Construction of the SIV-rtTA molecular clone was described previously (18). The variant used in this study contains the wild-typetatgene (19). Deletions in TAR were introduced in both the 5⬘LTR and 3⬘LTR of the plasmid pSIV-rtTA-Tatwt
through multiple steps of PCR mutagenesis. The primers used in this study are listed in Table 1. (i) Introduction of TAR deletions into 3ⴕLTR. The mutagenesis protocol involved two PCRs with primer sets specific for each TAR deletion (PCR1 and PCR2), followed by a common PCR (PCR3).
(a) Deletion of SL1. PCR1 was performed with primers 5WT⌬SL1 and TA016luc on the template pSIV-LTR-2⌬15tetO-TARwt
-luc (18) for the con-struction of the 1⌬2wtvariant, with primers 5Mut⌬SL1 and TA016luc on pSIV-LTR-2⌬15tetO-TARm
-luc (18) for the 1⌬2m
variant, and with primers 5⫹63⌬SL1 and TA016luc on pSIV-LTR-2⌬15tetO-TARm⫹63-luc (17) for the 1⌬2⫹63variant. PCR2 was performed with primers 5Nde1 and 3⌬SL1 on pSIV-LTR-2⌬15tetO-TARm-luc for all three variants.
(b)Deletion of SL2.PCR1 was performed with primers 5Nde1 and 3⌬SL2 on pSIV-LTR-2⌬15tetO-TARwt-luc for the 1wt2⌬ variant or on pSIV-LTR-2⌬15tetO-TARm
-luc for the 1m
2⌬variant. PCR2 was performed with primers 5⌬SL2 and TA016luc on pSIV-LTR-2⌬15tetO-TARm-luc for both variants.
(c)Deletion of SL3.PCR1 was performed with primers 5Nde1 and 3⌬SL3 on pSIV-LTR-2⌬15tetO-TARm⫹63-luc. PCR2 was performed with primers 5⌬SL3 and TA016luc on pSIV-LTR-2⌬15tetO-TARm⫹63-luc.
(d)Complete deletion of TAR.PCR1 was performed with primers 5Nde1 and 3⌬TAR on pSIV-LTR-2⌬15tetO-TARm
-luc. PCR2 was performed with primers 5⌬TAR and TA016luc on pSIV-LTR-2⌬15tetO-TARm-luc.
(e)Nearly complete deletion of TAR. PCR1 was performed with primers 5Nde1 and 3TAR10⌬on pSIV-LTR-2⌬15tetO-TARm-luc. PCR2 was performed with primers 5TAR10⌬and TA016luc on pSIV-LTR-2⌬15tetO-TARm
-luc. The PCR1 and PCR2 products for each TAR variant were purified, mixed, and used as a template in PCR3 with primers 5Nde1 and TA016luc. The PCR3 products were digested with EcoRI and XhoI and used to replace the corre-sponding fragments in pBS-3⬘SIV-rtTA, which contains the 3⬘half of the SIV-rtTA genome (18). We thus generated pBS-3⬘SIV-rtTA plasmids with different TAR deletions in the 3⬘LTR.
(ii) Introduction of TAR deletions into 5ⴕLTR.PCRs were performed with primers 5Nde1 and 3TARNar1, with the different TAR-deleted pBS-3⬘SIV-rtTA constructs as templates. The PCR products were digested with EcoRI and NarI and used to replace the corresponding fragment in the pKP-5⬘SIV construct, which contains the 5⬘half of the SIV-rtTA genome (18), resulting in pKP-5⬘SIV plasmids with the different TAR deletions in the 5⬘LTR.
(iii) Introduction of TAR deletions into SIV-rtTA molecular clone.The TAR modifications were introduced into the 3⬘LTR of the SIV-rtTA molecular clone through ligation of the XmaI-XhoI fragment of each pBS-3⬘SIV-rtTA TAR deletion variant into the XmaI- and XhoI-digested pSIV-rtTA-Tatwt
plasmid. The TAR modifications were subsequently also introduced into the 5⬘LTR by ligation of the NotI-NarI fragment of each pKP-5⬘SIV TAR deletion variant into NotI- and NarI-digested pSIV-rtTA with the corresponding modification in the 3⬘LTR.
RNA secondary structure prediction. Computer-assisted RNA secondary structure predictions were performed using the Mfold, version 3.2, algorithm (47) offered by the MBCMR Mfold server (http://mfold.burnet.edu.au). Stan-dard settings were used for all folding jobs (37°C and 1 M NaCl).
rtTA activity assay.In the pSIV-LTR-2⌬15tetO-TARm-luc plasmid, the ex-pression of firefly luciferase is under the control of the LTR-2⌬tetO promoter of SIV-rtTA (18). In this plasmid, the complete LTR region of SIV-rtTA (encom-passing U3, R, and U5) is present upstream of the luciferase gene. The EcoRI-XhoI fragment of each pSIV-rtTA TAR deletion variant, spanning the U3 promoter and the TAR region, was used to replace the corresponding sequences in the pSIV-LTR-2⌬15tetO-TARm-luc plasmid, which resulted in the TAR de-letion variants of this reporter construct. The plasmid pRL-CMV (Promega), in which the expression ofRenillaluciferase is controlled by the cytomegalovirus (CMV) immediate-early enhancer/promoter, was cotransfected into C33A cells to allow correction for differences in transfection efficiency.
C33A cells were cultured in 2-cm2
wells and transfected with 20 ng
pSIV-A
B
TAR
leader RNA
polyA PBSDIS
Ψ
1
SD
Ψ
2
Ψ
3
AUG
gag
env
nef
rev
tat
vpx
vif
vpr
pol
5’LTR
3’LTR
SL1
SL2
SL3
G G A G A G A A C U C U CAC C A G C A C UUGG C C G C U G G C A G A U UG A G C C C U GGGA G G UU C U CU C CA
GC CUAG CAG
G UA GAG
CCU G GUG G U U C C C U G C U A G A GU C GC U C U GC GC
A G A G U G A C U +124 +18 +85 +53 +93 +113 G CU U A A A A G U C A C G C U UG C U U A A G C U A G U G U G +126 +186 +171 C A G C C C U
A AU
CU UC
G AU UU UA C G A A C G U C A +1 +141 G G A G A G A A C U C U CA
C C A G C A C UUGG C C G G U G C U G G G C U G G C A G A U UG A G C C C U GGGA G G UU C U CU C CA
GC CUAG CAG
G UA GAG
CCU G GUG G U U C C C U G C U A G A GU C GC U C U GC GC
A G A G U G A C U +124 +18 +85 +53 +93 +113 G CU U A A A A G U C A C G C U UG C U U A A G C U A G U G U G +126 +186 +171 C A G C C C U
A AU
CU UC
G AU UU UA C G A A C G U C A +1 +141
TAR
polyA
TAR polyA PBSDIS SD
Ψ
AUG
[image:2.585.42.283.71.650.2]leader RNA
pol
gag
env
nef
rev
tat
vpr
vif
vpu
5’LTR
3’LTR
A C G G G A C C C U G G G A A C C C U U G G G A C U UU U G C G G G G G A A A A A C C C U U U U U U U G G A C C U U G G A C C U A U A A U U A C G U G A A C U A A C G U U G C A G U A U G U C A C G G U C C +1 +104 A C G G G A C C C U G G G A A C C C U U G G G A C U UU U G C G G G G G A A A A A C C C U U U U U U U G G A C C U U G G A C C U A U A C G G A C C U A U A A U U A C G U G A A C U U A C G U G A A C U U A A C G U U G C A A C G U U G C A G U A U G U C A G U A U G U C A C G G U C C +1 +104TAR polyA
FIG. 1. Comparison of HIV-1 and SIVmac239 TAR structures. Sche-matic representations of the HIV-1 (A) and SIVmac239 (B) genomes are shown. The 5⬘LTR, 3⬘LTR, and open reading frames are indicated. The untranslated leader RNAs of HIV-1 (positions⫹1 to⫹335) and SIVmac239 (positions⫹1 to⫹534) consist of several regulatory domains, including the TAR, poly(A) hairpin, primer-binding site (PBS), dimerization initiation site (DIS), splice donor (SD), RNA packaging signal (⌿), and start codon ofgag
(AUG) (22, 31). TAR and poly(A) hairpin folding is depicted.
on November 8, 2019 by guest
http://jvi.asm.org/
LTR-2⌬15tetO-TAR-luc construct (TARm, TARm⫹63, TARwt, or TAR deletion variant), 0.5 ng pRL-CMV, and 0.4 ng rtTA expression plasmid pCMV-rtTAF86Y A209T(19) or 0 to 50 ng SIV Tat expression plasmid pcDNA3-SIV-Tat (18). pBluescript was added to the transfection mix as carrier DNA to a total of 1g of DNA. After transfection, C33A cells were cultured for 48 h with 0 to 1,000 ng/ml DOX. Cells were lysed in passive lysis buffer, and firefly andRenilla
luciferase activities were determined with a dual-luciferase assay (Promega). The expression of firefly andRenillaluciferases was within the linear range, and no squelching effects were observed. The promoter activity was calculated as the ratio between the firefly andRenillaluciferase activities and corrected for be-tween-session variation (41).
Isolation and Northern blot analysis of viral RNA.For the isolation of intra-cellular viral RNA, 293T cells were transfected with 5g of the pSIV-rtTA constructs by calcium phosphate precipitation and cultured in 10-cm2
wells with 1,000 ng/ml DOX (19). After 48 h, cells were washed with phosphate-buffered saline (PBS), lysed in 350l RLT buffer (Qiagen), and homogenized with a QIAshredder column (Qiagen). Total cellular RNA was isolated with an RNeasy kit (Qiagen), and contaminating DNA was removed with RNase-free DNase during isolation (Qiagen). Upon electrophoresis of 5g RNA in a 1% agarose gel in 1⫻morpholinepropanesulfonic acid (MOPS) buffer (40 mM MOPS, 10 mM sodium acetate, pH 7.0) with 6.5% formaldehyde at 100 V, RNAs were transferred onto a positively charged nylon membrane (Boehringer Mannheim) overnight by means of capillary force. RNAs were linked to the membrane in a UV cross-linker (Stratagene). An NdeI fragment spanning part of thertTAgene and the U3 region of SIV-rtTA was labeled with [␣-32P]dCTP by use of a High Prime DNA labeling kit (Roche). Prehybridization and hybridization of the membrane with the probe were done in ULTRAhyb buffer (Ambion) at 55°C for 1 and 16 h, respectively. The membrane was then washed at room temperature in low-stringency buffer (2⫻SSC–0.2% sodium dodecyl sulfate [1⫻SSC is 0.15 M NaCl plus 0.015 M sodium citrate]) and at 55°C in high-stringency buffer (0.1⫻SSC–0.2% sodium dodecyl sulfate). Images were obtained using a Storm 860 phosphorimager (Amersham Biosciences), and data analysis was performed with the ImageQuant software package. The sizes of SIV-rtTA RNA fragments were estimated from their migration on the Northern blot, using 18S and 28S rRNAs as size markers.
Cell and virus cultures.293T cells were transfected with 5g of the pSIV-rtTA constructs by calcium phosphate precipitation and cultured in 10-cm2wells with 1,000 ng/ml DOX as previously described (19). Culture supernatants were harvested 48 h after transfection and filtered through 0.45-m-pore-size filters. Virus production was determined by CA-p27 enzyme-linked immunosorbent assay (Coulter).
The PM1 T-cell line was cultured at 37°C and 5% CO2in RPMI 1640 con-taining 10% fetal bovine serum, 100 units/ml penicillin, and 100 units/ml strep-tomycin. To assay virus replication, PM1 cells were infected by incubation with
an equal amount of each SIV-rtTA variant, corresponding to 1 ng CA-p27, for 4 h at 37°C. Cells were then extensively washed with PBS and cultured in 2-cm2 wells in complete RPMI medium with 1,000 ng/ml DOX. Cell-free supernatants were collected and viral replication was monitored with a real-time PCR-based reverse transcriptase (RT) assay (18). Each experiment was performed in trip-licate.
Peripheral blood mononuclear cells (PBMC) were isolated from cynomolgus macaques (36) and cultured as described previously (17). Cells were activated with 2g/ml phytohemagglutinin for 2 days and infected with equal amounts of virus (10 ng CA-p27) for 16 h as described for PM1 cells. Cells were maintained with 100 units/ml recombinant interleukin-2 and 1,000 ng/ml DOX following infection. The RT levels in cell-free supernatants were determined to monitor viral replication.
RESULTS
Construction of SIV-rtTA variants in which TAR is partially or completely deleted.We previously constructed a condition-ally live SIVmac239 variant that replicates exclusively in the presence of DOX (18). In this SIV-rtTA variant, the Tat-TAR transcription activation mechanism was functionally replaced by the DOX-inducible Tet-On gene regulation system (Fig. 2A). Briefly, the Tat-TAR axis was inactivated through muta-tion of the bulge and loop sequences in the SL1 and SL2 domains of TAR, which prevents the binding of Tat and cyclin T1 (18). The Tet-On system was incorporated into the SIV-mac239 genome by insertion of thertTAtranscriptional acti-vator gene in place of thenefgene andtetoperator sites (teto) in the LTR promoter (19). In the presence of DOX, rtTA can bind to teto and activate transcription from the downstream transcription initiation site. Transcription and replication of SIV-rtTA are thus critically dependent on DOX. Although SIV-rtTA does not require Tat for transcription, mutational inactivation of Tat abolished viral replication (18). This result indicated that Tat has an additional function in SIVmac rep-lication, and the wild-typetatgene was therefore maintained in SIV-rtTA. Upon long-term culturing, SIV-rtTA acquired an A-to-G substitution at position⫹63 in the SL2 domain of TAR that strongly improved viral replication. This mutation did not restore Tat binding, and transcription of the new SIV-rtTA variant was still fully dependent on DOX (17).
Since SIV-rtTA does not need TAR for transcription from the LTR promoter, it is an ideal tool for studying additional TAR functions in SIVmac replication. We therefore con-structed novel SIV-rtTA variants with different TAR configu-rations (Fig. 2B). Compared to wild-type SIVmac239 (TARwt), SIV-rtTA carries mutations in the bulge and loop sequences in both SL1 and SL2 (TARm). In this SIV-rtTA context, we deleted either SL1 (1⌬2mvariant) or SL2 (1m2⌬). We also used
the improved SIV-rtTA variant with the additional TAR mu-tation (TARm⫹63), in which either SL1 (1⌬2⫹63) or SL3 (1m2⫹633⌬) was deleted. We also generated variants in which a short 8-bp stem-loop structure, formed by the first and last 10 nt of TAR, was left (TAR10⌬) or in which the complete TAR sequence was deleted (⌬TAR). As controls, we constructed variants with a wild-type TAR (TARwt) and derivatives with a deletion of SL1 (1⌬2wt) or SL2 (1wt2⌬).
Efficient gene expression upon partial or complete TAR deletion.To determine the effects of partial and complete TAR deletion on viral gene expression, we generated promoter-re-porter constructs in which the TAR mutant LTR promoter of the different SIV-rtTA variants was placed upstream of a
lu-TABLE 1. Primers used in this study
Primer Sequence (5⬘–3⬘)
5Nde1 ...GAGGCATATGTTAGATACCCAG TA016luc ...GGTTCCATCTTCCAGCGG
3TARNar1...CTGTTCAGGCGCCAATCTGCTAGGGATTTT CCTG
5WT⌬SL1 ...ATTCAGTCGCTCTGCGGAGAG-SL1 deletion-CT AGCAGGTAGAGCCTGGG
5Mut⌬SL1...ATTCAGTCGCTCTGCGGAGAG-SL1 deletion-CT AGCAGGAAGAGCATTGG
5⫹63⌬SL1 ...ATTCAGTCGCTCTGCGGAGAG-SL1 deletion-CT AGCAGGAGGAGCATTGG
3⌬SL1...CTCCGCAGAGCGACTGAATACAGAG 5⌬SL2...CTCACCAGCACTTGGCCGG
3⌬SL2...CCGGCCAAGTGCTGGTGAGAG-SL2 deletion-TGCTGGAGAGAACCTCC
5⌬SL3...GTTCCCTGCTAGACTCTCA-SL3 deletion-GCAG AGTGACTCCACGCTTG
3⌬SL3...GAGAGTCTAGCAGGGAACACC
5TAR10⌬...CGCTCTGTATTCAGTCGCTCTGC-TAR deletion-AGAGTGACTCCACGCTTGCTTG
3TAR10⌬...GCAGAGCGACTGAATACAGAGCGAAATG 5⌬TAR...CACTGCATTTCGCTCTGTATTCC-TAR
deletion-ACGCTTGCTTGCTTAAAGCCCT 3⌬TAR...GGAATACAGAGCGAAATGCAGTG
VOL. 82, 2008 ROLE OF TAR IN SIVmac REPLICATION 9173
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.45.285.77.307.2]9174
on November 8, 2019 by guest
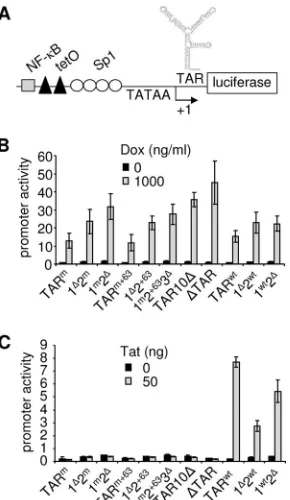
ciferase reporter gene (Fig. 3A). To assay DOX responsive-ness, C33A cells were transfected with the LTR-luciferase con-structs and an rtTA-expressing plasmid and cultured with or without DOX. The luciferase level measured 48 h after trans-fection reflects the efficiency of viral gene expression. All con-structs showed a low level of luciferase expression in the ab-sence of DOX and a high level with DOX (Fig. 3B). Deletion of SL1 (compare 1⌬2mwith TARm, 1⌬2⫹63with TARm⫹63, and 1⌬2wtwith TARwt), SL2 (compare 1m2⌬with TARmand 1wt2⌬
with TARwt), or SL3 (compare 1m2⫹633⌬with TARm⫹63) re-sulted in an increased level of gene expression. This can be explained by destabilization of the TAR structure, which has been reported to have an inhibitory effect on mRNA transla-tion (24, 42, 43). In agreement with this, the highest luciferase levels were observed upon nearly complete or complete dele-tion of TAR (TAR10⌬and⌬TAR).
The Tat responsiveness of the constructs was determined by transfection of C33A cells with the LTR-luciferase constructs and a Tat-expressing plasmid. As previously observed, the TARmand TARm⫹63constructs did not show any Tat stimu-lation (Fig. 3C), which is in agreement with the finding that SIV Tat cannot bind to TARmand TARm⫹63RNAs (17, 18). As expected, derivatives of the TARm and TARm⫹63 con-structs in which either SL1 (1⌬2m, 1⌬2⫹63), SL2 (1m2⌬), or SL3 (1m2⫹633⌬) was deleted and the variants with a complete or
nearly complete TAR deletion also did not respond to Tat. Only the control constructs with wild-type TAR (TARwt) and its variants in which either SL1 or SL2 was deleted (1⌬2wt and 1wt2⌬) showed luciferase expression with Tat. The 1⌬2wt and 1wt2⌬variants exhibited weaker Tattrans-activation capac-ities than that of the TARwtconstruct, which is in agreement with previous studies demonstrating that both SL1 and SL2 contribute to optimal Tat activation of transcription (10, 25). The contribution of SL1 is quantitatively more important than that of SL2, which could be due to hairpin characteristics (e.g., the presence of a purine in the SL2 bulge) (26, 40). These results demonstrate that partial or complete deletion of TAR allows efficient, DOX-dependent, Tat-independent gene ex-pression from the SIV-rtTA promoter.
TAR deletions do not affect SIV-rtTA production.To assess whether partial or complete deletion of TAR affects the pro-duction of viral RNA and proteins, we introduced the TAR deletions into both the 5⬘LTR and 3⬘LTR of SIV-rtTA. 293T cells were transfected with the SIV-rtTA variants and cultured in the presence of DOX for 48 h. Northern blot analysis of the viral RNAs isolated from the transfected cells revealed no significant quantitative differences in unspliced, singly spliced, and multispliced RNAs of the original and TAR-deleted SIV-rtTA variants (Fig. 4). The observed differences in transcript size are due to the TAR deletion in both the 5⬘LTR and
3⬘LTR. This result demonstrates that the partial or complete deletion of TAR in SIV-rtTA does not significantly affect the production and processing of viral RNA. Virus production was determined by measurement of CA-p27 (Fig. 5) and RT activ-ity (not shown) in the culture supernatant. Both analyses re-vealed similarly high levels of virus production for all SIV-rtTA variants. Whereas increased reporter gene expression was ob-served upon TAR truncation (Fig. 3B), CA-p27 production of the different TAR-truncated SIV-rtTA variants did not show this trend, possibly due to the presence of the complete un-translated leader in these viral RNAs (Fig. 5). These results demonstrate that partial or complete deletion of TAR does not impair SIV-rtTA virion production.
TAR-deleted SIV-rtTA variants replicate efficiently. To study the impact of partial and complete TAR deletion on virus replication, PM1 T cells were infected with the SIV-rtTA variants and viral replication was monitored by measurement of the RT level in the culture supernatant. SIV-rtTA-TARm⫹63replicated more efficiently than the original SIV-rtTA-TARmconstruct and almost as efficiently as the parental strains SIVmac239 and SIVmac239⌬Nef (Fig. 6A). Remark-ably, the SIV-rtTA-TARm derivatives in which either SL1 (1⌬2m) or SL2 (1m2⌬) was deleted replicated as efficiently as
the TARm⫹63variant (Fig. 6B). Deletion of SL1 (1⌬2⫹63) or SL3 (1m2⫹633⌬) in the TARm⫹63variant did not affect viral replication. The TAR10⌬variant, in which TAR was almost completely deleted, also replicated efficiently (Fig. 6C). Only the⌬TAR variant with a complete TAR deletion ex-hibited a reduced replication potential. This ⌬TAR muta-tion alters the sequence around the transcripmuta-tion start site, which may affect the selection of the initiation site. Se-quence analysis of the 5⬘ ends of the viral transcripts did indeed reveal that transcription of SIV-rtTA-⌬TAR fre-quently started downstream of the original⫹1 position (2 or more nt downstream [data not shown]). This aberrant initi-ation of transcription may explain the reduced repliciniti-ation capacity of the⌬TAR variant. In the TAR10⌬ variant, in which only the first and last 10 nt of the TAR sequence were maintained, transcription initiated at the original⫹1 posi-tion, and this virus did indeed replicate efficiently. These results demonstrate that partial or nearly complete deletion of TAR does not impair replication of SIV-rtTA in PM1 cells. In agreement with this, sequence analysis of these TAR-deleted variants upon prolonged culturing (up to 200 days) revealed that the introduced mutations were stably maintained, and none of the variants reverted to a wild-type TAR structure (data not shown).
In addition, we assessed replication of the TAR10⌬ and TARm⫹63variants in PBMC isolated from cynomolgus ma-caques (Fig. 6D). As observed with PM1 cells, these variants
FIG. 2. Mutation of the TAR element in the DOX-inducible SIV-rtTA variant. (A) In the DOX-dependent SIV-rtTA variant, the Tat-TAR transcription mechanism was functionally replaced by the Tet-On gene regulation mechanism. TAR was inactivated by mutation of the bulge and loop sequences (TARm). ThertTAgene was inserted at the site of thenefgene, andtetobinding sites were introduced in both the 5⬘LTR and 3⬘LTR promoter regions. (B) We generated a set of partial to complete TAR deletion variants. The constructs used in this study have a wild-type (TARwt), mutated (TARm; bulge and loop mutations are shown with black circles), or evolved (TARm⫹63;⫹63A-Gsubstitution is shown with a gray circle) TAR element. We made novel variants in which either SL1 (1⌬2WT, 1⌬2m, and 1⌬2⫹63), SL2 (1wt2⌬and 1m2⌬), or SL3 (1m2⫹633⌬) was deleted. In
addition, we made a⌬TAR variant, in which all TAR sequences were deleted (not shown), and a TAR10⌬variant, in which TAR was truncated to a 20-nt hairpin corresponding to nt⫹1 to⫹10 and⫹114 to⫹124.
VOL. 82, 2008 ROLE OF TAR IN SIVmac REPLICATION 9175
on November 8, 2019 by guest
http://jvi.asm.org/
replicated with similar efficiencies in macaque PBMC, which demonstrates that the nearly complete deletion of TAR also does not affect replication of SIV-rtTA in primary cells. Both DOX-controlled viruses replicated somewhat less efficiently than wild-type SIVmac239 and the SIVmac239⌬Nef variant, which were included for comparison.
DISCUSSION
In this study, we analyzed the role of the complex TAR RNA element in SIVmac replication. For this purpose, we used a SIV-rtTA variant that does not depend on TAR for transcrip-tion, which allowed us to address the issue of whether TAR has an additional function in SIVmac replication. We demon-strated that partial or nearly complete deletion of TAR does not affect viral transcription, RNA processing, production, or replication. Therefore, we conclude that TAR has no essential function in SIVmac replication in vitro apart from its role in Tat-mediated activation of transcription.
We recently showed that TAR deletion is also compatible with replication of a similar DOX-dependent HIV-1 variant (HIV-rtTA) (15). While SIVmac has an⬃124-nt TAR element that folds into a complex three-stem-loop domain, the HIV-1 TAR element is a single hairpin of only ⬃57 nt (Fig. 1).
Apparently, this relatively simple TAR structure suffices for HIV-1 replication. The strong conservation of the complex TAR structure in different SIV isolates suggested that all three stem-loop elements may play a certain role in SIV replication. Previous studies (10, 25) and our observations (Fig. 3C) did indeed reveal that efficient Tat-mediated activation of tran-scription involves both SL1 and SL2. No role for SL3 in the Tat
trans-activation function has been reported (25). In this study, we showed that none of the stem-loop domains has an essential function in any other replication step. However, it cannot be excluded that SL1, SL2, and/or SL3 may have an accessory function in the SIVmac life cycle under specific conditions or in specific cell types in vivo, since replication of the TAR-deleted SIV-rtTA variants was assayed only in cell culture systems (32).
[image:6.585.92.237.67.317.2]TAR is part of the repeat (R) sequence present at the 5⬘and 3⬘ ends of the viral genome. This region is essential for the first-strand transfer step of reverse transcription, in which the
[image:6.585.330.508.68.376.2]FIG. 3. Gene expression is not affected by TAR deletions. (A) Pro-moter-reporter constructs were generated in which the firefly lucifer-ase gene was under the control of the original or TAR-mutated SIV-rtTA LTR promoter. (B) To determine DOX responsiveness, C33A cells were transfected with the reporter constructs and an rtTA-ex-pressing plasmid and cultured with 0 or 1,000 ng/ml DOX. (C) To determine Tat responsiveness, C33A cells were transfected with the reporter constructs and 0 or 50 ng of Tat-expressing plasmid. (B and C) A CMV promoter–Renilla luciferase construct was cotransfected to allow correction for differences in transfection efficiency. The ratios of the firefly andRenillaluciferase activities measured 2 days after trans-fection reflect the LTR promoter activity. The error bars represent the standard deviations for three experiments.
FIG. 4. TAR deletions do not affect viral RNA production. 293T cells were transfected with the SIV-rtTA variants and cultured with 1,000 ng/ml DOX for 48 h. Total intracellular RNA was isolated and analyzed by Northern blot analysis with an SIV-specific probe. (A) The positions of multispliced (msRNA), singly spliced (ssRNA), and un-spliced (usRNA) RNAs are indicated on the blot. The size differences between the SIVmac239, SIVmac239⌬Nef, and SIV-rtTA RNAs are due to deletion ofnef sequences, introduction ofrtTAand teto se-quences, and/or deletion of TAR sequences. (B) Ethidium bromide staining of the corresponding agarose gel demonstrates that the ob-served differences in RNA signals are due to differences in RNA loading. The 18S (⬃1.9 kb) and 28S (⬃5.0 kb) rRNAs were used as size markers.
on November 8, 2019 by guest
http://jvi.asm.org/
copy DNA is translocated from the 5⬘ to the 3⬘ R sequence. Whereas the R region is 176 nt in wild-type SIVmac239, the efficient replication of the TAR10⌬variant demonstrates that a 72-nt R sequence suffices for this function. This observation is in agreement with previous HIV-1 studies demonstrating that strand transfer is efficient with shortened R regions of only 30 nt (12) and 39 nt (15).
We previously showed that HIV-1 transcripts require a per-fect hairpin structure at the extreme 5⬘end to avoid the pres-ence of an unpaired 5⬘ dangling end (15). TAR fulfills this prerequisite in wild-type HIV-1, but a truncated TAR hairpin or an unrelated hairpin also suffices in the context of HIV-rtTA. All SIV-rtTA TAR-mutated transcripts can fold a base-paired stem structure at the 5⬘ end, and even the severely truncated 8-bp hairpin of the TAR10⌬variant supports effi-cient replication. However, complete deletion of TAR, as in the⌬TAR variant, shifted the transcription start site further downstream. These aberrantly initiated transcripts start within the poly(A) hairpin and will not fold into a stable stem-loop structure at the 5⬘end (Fig. 1B), resulting in the presence of unpaired nucleotides. As recently observed for HIV-1, such unpaired nucleotides may interact with downstream RNA se-quences and affect the overall leader RNA folding (30, 37, 44). The secondary structure of this leader RNA region is impor-tant for several processes in the lentiviral life cycle, such as dimerization, packaging, splicing, and polyadenylation (1, 16, 21–23, 33–35, 38, 45, 46). Any modification of the leader RNA structure will likely hamper viral replication, which may ex-plain the delayed replication of the⌬TAR variant.
The reduced replication of the original SIV-rtTA variant (TARm) may also be due to aberrant folding of the leader RNA. Possibly, the introduced bulge and loop mutations de-stabilize the local TAR folding or generate a new sequence with complementarity to downstream sequences, which could trigger an interaction between TAR sequences and a down-stream leader domain. The beneficial⫹63A-Gmutation that is selected during SIV-rtTA replication may prevent this un-wanted interaction and thus restore viral replication (17). Al-though details of such a misfolding scenario are currently lack-ing, this scenario is supported by the observation that the precise removal of SL1 (1⌬2mvariant), SL2 (1m2⌬), or all SL
sequences (TAR10⌬) similarly improves SIV-rtTA replication. SIV-rtTA can be used as a tool in SIV biology studies and vaccine research because both the level and duration of repli-cation can be controlled by DOX administration. We previ-ously demonstrated that SIV-rtTA requires wild-type Tat
pro-tein for replication, although gene expression is strictly controlled by the incorporated Tet-On system and not affected by Tat inactivation. This result suggests that Tat, unlike TAR, may have an important second function in the SIVmac life cycle, in addition to its transcriptional role. For this reason, the SIV-rtTA variant used in this study encodes the wild-type Tat protein. Reversion of the bulge and loop mutations in TARm, which had been introduced to prevent Tat and cyclin T1 bind-ing, would restore the Tat-TAR mechanism of transcription control. However, such a repair route requires multiple nucle-otide substitutions in the bulge and loop of TAR, which are not likely to occur. Indeed, we never observed restoration of the Tat-TAR axis in multiple long-term cultures of SIV-rtTA.
[image:7.585.335.510.68.428.2]Nev-FIG. 5. TAR deletions do not impair virus production. 293T cells were transfected with the SIV-rtTA variants and cultured for 2 days with 1,000 ng/ml DOX. The virus level in the culture supernatant was determined by CA-p27 enzyme-linked immunosorbent assay. The er-ror bars represent the standard deviations for three experiments.
FIG. 6. Effects of TAR deletions on virus replication. (A to C) PM1 cells were infected with SIV-rtTA variants (1 ng CA-p27) and cultured with 1,000 ng/ml DOX. Replication of the original (TARm) and evolved (TARm⫹63) SIV-rtTA variants and of wild-type and Nef-de-leted SIVmac239 (A), of variants with a partial deletion of TAR (B), and of variants with a nearly complete to complete TAR deletion (C) is shown. Viral replication was monitored by quantification of the RT level in the culture supernatant. Each experiment was performed in triplicate, and data for a representative experiment are shown. Similar results were obtained when the infected cells were cultured with 100 ng/ml DOX and when virus cultures were started by transfection of either 1 or 5g of the proviral plasmids (data not shown). (D) PBMC isolated from cynomolgus macaques were infected with the TAR10⌬ and TARm⫹63variants and with wild-type and Nef-deleted SIVmac239 (10 ng CA-p27) and then cultured with 1,000 ng/ml DOX.
VOL. 82, 2008 ROLE OF TAR IN SIVmac REPLICATION 9177
on November 8, 2019 by guest
http://jvi.asm.org/
[image:7.585.102.224.69.150.2]ertheless, the likelihood of this unwanted evolution route is further reduced by the deletion of TAR sequences, and it seems highly unlikely that the TAR10⌬variant can revert to a Tat-TAR transcription mechanism.
ACKNOWLEDGMENTS
We thank Alex Harwig, Nicolas Legrand, Marcel Ooms, and Monique Vink for discussions, technical support, and comments on the manuscript. We thank Neil Almond and Mark Page (National Institute for Biological Standards and Control, United Kingdom) for providing macaque PBMC.
Mireille Centlivre was supported by postdoctoral fellowships from the FRM (Fondation pour la Recherche Medicale) and the EU (Marie Curie fellowship MEIF-CT-2007-039689). This research was spon-sored by the Dutch AIDS Foundation (AIDS Fonds grant 2005022) and the International AIDS Vaccine Initiative (IAVI).
REFERENCES
1.Abbink, T. E., and B. Berkhout.2007. RNA structure modulates splicing efficiency at the HIV-1 major splice donor. J. Virol.82:3090–3098. 2.Andersen, E. S., S. A. Contera, B. Knudsen, C. K. Damgaard, F.
Besen-bacher, and J. Kjems.2004. Role of the trans-activation response element in dimerization of HIV-1 RNA. J. Biol. Chem.279:22243–22249.
3.Arya, S. K., B. Beaver, L. Jagodzinski, B. Ensoli, P. J. Kanki, J. Albert, E. M. Fenyo, G. Biberfeld, J. F. Zagury, F. Laure, M. Essex, E. Norrby, F. Wong Staal, and R. C. Gallo.1987. New human and simian HIV-related retrovi-ruses possess functional transactivator (tat) gene. Nature328:548–550. 4.Arya, S. K., C. Guo, S. F. Josephs, and F. Wong-Staal.1985. Trans-activator
gene of human T-lymphotropic virus type III (HTLV-III). Science229:69–73. 5.Bannwarth, S., and A. Gatignol.2005. HIV-1 TAR RNA: the target of
molec-ular interactions between the virus and its host. Curr. HIV Res.3:61–71. 6.Bennasser, Y., S. Y. Le, M. L. Yeung, and K. T. Jeang.2004. HIV-1 encoded
candidate micro-RNAs and their cellular targets. Retrovirology1:43. 7.Bennasser, Y., M. L. Yeung, and K. T. Jeang.2006. HIV-1 TAR RNA
subverts RNA interference in transfected cells through sequestration of TAR RNA-binding protein, TRBP. J. Biol. Chem.281:27674–27678. 8.Berkhout, B.1992. Structural features in TAR RNA of human and simian
immu-nodeficiency viruses: a phylogenetic analysis. Nucleic Acids Res.20:27–31. 9.Berkhout, B.2000. Multiple biological roles associated with the repeat (R)
region of the HIV-1 RNA genome. Adv. Pharmacol.48:29–73.
10.Berkhout, B., A. Gatignol, J. Silver, and K. T. Jeang.1990. Efficient trans-activation by the HIV-2 Tat protein requires a duplicated TAR RNA struc-ture. Nucleic Acids Res.18:1839–1846.
11.Berkhout, B., R. H. Silverman, and K. T. Jeang.1989. Tat trans-activates the human immunodeficiency virus through a nascent RNA target. Cell59:273–282. 12.Berkhout, B., J. van Wamel, and B. Klaver.1995. Requirements for DNA
strand transfer during reverse transcription in mutant HIV-1 virions. J. Mol. Biol.252:59–69.
13.Berkhout, B., N. L. Vastenhouw, B. I. Klasens, and H. Huthoff.2001. Struc-tural features in the HIV-1 repeat region facilitate strand transfer during reverse transcription. RNA7:1097–1114.
14.Cullen, B. R.1986. Trans-activation of human immunodeficiency virus oc-curs via a bimodal mechanism. Cell46:973–982.
15.Das, A. T., A. Harwig, M. M. Vrolijk, and B. Berkhout.2007. The TAR hairpin of human immunodeficiency virus type 1 can be deleted when not required for Tat-mediated activation of transcription. J. Virol.81:7742–7748. 16.Das, A. T., B. Klaver, and B. Berkhout.1999. A hairpin structure in the R region of the human immunodeficiency virus type 1 RNA genome is instru-mental in polyadenylation site selection. J. Virol.73:81–91.
17.Das, A. T., B. Klaver, M. Centlivre, A. Harwig, M. Ooms, M. Page, N. Almond, F. Yuan, M. Piatak, Jr., J. D. Lifson, and B. Berkhout.2008. Optimization of the doxycycline-dependent simian immunodeficiency virus through in vitro evolution. Retrovirology5:44.
18.Das, A. T., B. Klaver, A. Harwig, M. Vink, M. Ooms, M. Centlivre, and B. Berkhout.2007. Construction of a doxycycline-dependent simian immuno-deficiency virus reveals a nontranscriptional function of Tat in viral replica-tion. J. Virol.81:11159–11169.
19.Das, A. T., X. Zhou, M. Vink, B. Klaver, K. Verhoef, G. Marzio, and B. Berkhout.2004. Viral evolution as a tool to improve the tetracycline-regu-lated gene expression system. J. Biol. Chem.279:18776–18782.
20.Dayton, A. I., J. G. Sodroski, C. A. Rosen, W. C. Goh, and W. A. Haseltine. 1986. The trans-activator gene of the human T cell lymphotropic virus type III is required for replication. Cell44:941–947.
21.Dirac, A. M., H. Huthoff, J. Kjems, and B. Berkhout. 2001. The dimer initiation site hairpin mediates dimerization of the human immunodeficiency virus, type 2 RNA genome. J. Biol. Chem.276:32345–32352.
22.Dirac, A. M., H. Huthoff, J. Kjems, and B. Berkhout.2002. Regulated HIV-2
RNA dimerization by means of alternative RNA conformations. Nucleic Acids Res.30:2647–2655.
23.Dirac, A. M., H. Huthoff, J. Kjems, and B. Berkhout.2002. Requirements for RNA heterodimerization of the human immunodeficiency virus type 1 (HIV-1) and HIV-2 genomes. J. Gen. Virol.83:2533–2542.
24.Dorin, D., M. C. Bonnet, S. Bannwarth, A. Gatignol, E. F. Meurs, and C. Vaquero.2003. The TAR RNA-binding protein, TRBP, stimulates the ex-pression of TAR-containing RNAs in vitro and in vivo independently of its ability to inhibit the dsRNA-dependent kinase PKR. J. Biol. Chem.278: 4440–4448.
25.Emerman, M., M. Guyader, L. Montagnier, D. Baltimore, and M. A. Muesing.1987. The specificity of the human immunodeficiency virus type 2 transactivator is different from that of human immunodeficiency virus type 1. EMBO J.6:3755–3760.
26.Fenrick, R., M. H. Malim, J. Hauber, S. Y. Le, J. Maizel, and B. R. Cullen. 1989. Functional analysis of the Tat transactivator of human immunodefi-ciency virus type 2. J. Virol.63:5006–5012.
27.Gatignol, A., A. Buckler-White, B. Berkhout, and K. T. Jeang.1991. Char-acterization of a human TAR RNA-binding protein that activates the HIV-1 LTR. Science251:1597–1600.
28.Guyader, M., M. Emerman, P. Sonigo, F. Clavel, L. Montagnier, and M. Alizon.1987. Genome organization and transactivation of the human immu-nodeficiency virus type 2. Nature326:662–669.
29.Harrich, D., C. W. Hooker, and E. Parry.2000. The human immunodefi-ciency virus type 1 TAR RNA upper stem-loop plays distinct roles in reverse transcription and RNA packaging. J. Virol.74:5639–5646.
30.Huthoff, H., and B. Berkhout.2001. Mutations in the TAR hairpin affect the equilibrium between alternative conformations of the HIV-1 leader RNA. Nucleic Acids Res.29:2594–2600.
31.Huthoff, H., and B. Berkhout.2001. Two alternating structures for the HIV-1 leader RNA. RNA7:143–157.
32.Jones, K. L., S. Sonza, and J. Mak.2008. Primary T-lymphocytes rescue the replication of HIV-1 DIS RNA mutants in part by facilitating reverse tran-scription. Nucleic Acids Res.36:1578–1588.
33.Jossinet, F., J. S. Lodmell, C. Ehresmann, B. Ehresmann, and R. Marquet. 2001. Identification of thein vitroHIV-2/SIV RNA dimerization site reveals striking differences with HIV-1. J. Biol. Chem.276:5598–5604.
34.Lanchy, J. M., and J. S. Lodmell.2002. Alternate usage of two dimerization initiation sites in HIV-2 viral RNA in vitro. J. Mol. Biol.319:637–648. 35.Lanchy, J. M., C. A. Rentz, J. D. Ivanovitch, and J. S. Lodmell.2003.
Elements located upstream and downstream of the major splice donor site influence the ability of HIV-2 leader RNA to dimerize in vitro. Biochemistry 42:2634–2642.
36.Mills, K. H., A. L. Barnard, M. Williams, M. Page, C. Ling, E. J. Stott, P. Silvera, F. Taffs, A. S. Kingsman, S. E. Adams, N. Almond, P. A. Kitchin, and W. C. Jones.1991. Vaccine-induced CD4⫹T cells against the simian immu-nodeficiency virus gag protein. Epitope specificity and relevance to protec-tive immunity. J. Immunol.147:3560–3567.
37.Ooms, M., H. Huthoff, R. Russell, C. Liang, and B. Berkhout.2004. A riboswitch regulates RNA dimerization and packaging in human immuno-deficiency virus type 1 virions. J. Virol.78:10814–10819.
38.Patel, J., S. W. Wang, E. Izmailova, and A. Aldovini.2003. The simian immunodeficiency virus 5⬘ untranslated leader sequence plays a role in intracellular viral protein accumulation and in RNA packaging. J. Virol. 77:6284–6292.
39.Raha, T., S. W. Cheng, and M. R. Green.2005. HIV-1 Tat stimulates transcription complex assembly through recruitment of TBP in the absence of TAFs. PLoS Biol.3:e44.
40.Rhim, H., and A. P. Rice.1994. Functional significance of the dinucleotide bulge in stem-loop1 and stem-loop2 of HIV-2 TAR RNA. Virology202:202–211. 41.Ruijter, J. M., H. H. Thygesen, O. J. Schoneveld, A. T. Das, B. Berkhout, and
W. H. Lamers.2006. Factor correction as a tool to eliminate between-session variation in replicate experiments: application to molecular biology and retrovirology. Retrovirology3:1–8.
42.SenGupta, D. N., B. Berkhout, A. Gatignol, A. M. Zhou, and R. H. Silver-man.1990. Direct evidence for translational regulation by leader RNA and Tat protein of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA87:7492–7496.
43.Svitkin, Y. V., A. Pause, and N. Sonenberg.1994. LA autoantigen alleviates translational repression by the 5⬘leader sequence of the human immunode-ficiency virus type 1 RNA. J. Virol.68:7001–7007.
44.Vrolijk, M. M., M. Ooms, A. Harwig, A. T. Das, and B. Berkhout.2008. Destabilization of the TAR hairpin affects the structure and function of the HIV-1 leader RNA. Nucleic Acids Res.36:4352–4363.
45.Wang, S. W., and A. Aldovini.2002. RNA incorporation is critical for ret-roviral particle integrity after cell membrane assembly of Gag complexes. J. Virol.76:11853–11865.
46.Whitney, J. B., and M. A. Wainberg.2006. Impaired RNA incorporation and dimerization in live attenuated leader-variants of SIVmac239. Retrovirology3:96. 47.Zuker, M.2003. Mfold web server for nucleic acid folding and hybridization
prediction. Nucleic Acids Res.31:3406–3415.
on November 8, 2019 by guest
http://jvi.asm.org/