0022-538X/94/$04.00+0
Copyright© 1994,American SocietyforMicrobiology
Both NS3 and NS4A Are Required for Proteolytic Processing of
Hepatitis C Virus Nonstructural Proteins
CRISTINAFAILLA, LICIATOMEI,AND RAFFAELEDEFRANCESCO* Istituto di Ricerche diBiologia Molecolare "P. Angeletti"-Pomezia, 00040 Rome, Italy
Received 20 December 1993/Accepted 11 March 1994
The proteolytic cleavages at the NS3-NS4A, NS4A-NS4B, NS4B-NS5A, and NS5A-NS5B junctions of hepatitis C virus (HCV) polyproteinare effected bythe virus-encoded serineprotease containedwithinNS3. UsingtransientexpressioninHeLa cells of cDNAfragmentsthatcode forregionsof the HCVpolyprotein,we studiedwhetherviral functionsother thanNS3arerequired for proteolytic processingatthese sites.We found that,inadditiontoNS3,aC-terminal33-amino-acidsequenceof the NS4Aproteinisrequiredforcleavageat theNS3-NS4AandNS4B-NS5A sitesand thatitaccelerates therateofcleavage attheNS5A-NS5B junction. Inaddition,weshowthatNS4AcanactivatetheNS3proteasewhensuppliedintrans.Our datasuggestthat HCV NS4Amaybe thefunctionalanalogof flavivirus NS2Bandpestivirus plO proteins.
Hepatitis C virus (HCV) isthe major etiological agent of posttransfusion and sporadic community-acquired non-A, non-B hepatitis (11, 31). Inmostcases, HCVcauses a persis-tent liver infection that eventually develops into cirrhosis or hepatocellular carcinoma (10).
HCV contains a positive-stranded linear RNA genome of about 9.5 kb in length (12, 29, 41). On the basis of their genomicstructureand virionproperties, HCV, the pestivirus, andthe flavivirus have been classified as threegenera in the family Flaviviridae (19).Asin thecaseofflavi- and pestiviruses, HCV isasmall, enveloped virus, whose genomicRNAincludes asingleopenreading frame encodinga precursorpolyprotein that is cleaved co- or posttranslationally into mature viral polypeptides. The structural proteins are encoded by the N-terminal portion of thegenome,whereas the remainingpart
codes for the nonstructural proteins, whicharebelievedtobe
components of the viral replication machinery (for a recent
reviewonthe molecularbiology of HCV,seereference34). The map of the HCV polyprotein has been obtained by cell-free and cell culture expression studies (23, 24, 42). The geneorder that hasemergedfrom these studies is 5'-C-E1-E2-NS2-NS3-NS4A-NS4B-NSSA-NSSB-3'. C is a basic protein that binds RNA(39a), andit is postulated to be the nucleo-capsid protein.ElandE2areputative envelope glycoproteins. NS2throughNS5B representthe viral nonstructural proteins andcould befunctional analogsof thecorresponding proteins ofpesti-andflaviviruses (7, 13).
Processing of the polyprotein requires both host and viral proteases. The signal peptidase ofendoplasmic reticulum is responsible for generating the N termini of El, E2, and possibly NS2 (24). Whereas it has been recently established that anovel virus-encoded metalloprotease is responsible for the cleavage between NS2 and NS3 (21, 25), we and others have identified NS3as theproteaseresponsible forcleavages
at the NS3-NS4A, NS4A-NS4B, NS4B-NSSA, and NS5A-NS5B junctions (3, 15, 22, 42). Analysis of the amino acid sequence of theNS3proteinof HCVsuggestedthat this viral protein contains a trypsin-like serine protease domain that functions intheprocessing of the viralpolyprotein (35), as is
*Corresponding author. Phone: 93221. Fax:
39-6-910-93225.Electronic mail address: [email protected].
thecasefor thehomologousNS3proteinof flaviviruses(9)and p80ofpestiviruses (45).
The active siteofserineproteases invariably contains three residues, histidine, aspartate, and serine, that maintain the same relative spatial position in all the known structures of these enzymes. These residues constitute the catalytic triad: the histidine imidazolegroup abstracts the alcohol protonof the serine and transfers ittotheamine-leavinggroup,while the aspartate stabilizes the positive charge that develops on the histidine in the intermediatecomplex (36).The histidine1083,
aspartate 1107,and serine 1165 residues(numbered according
to their location in thepolyprotein of BK strain HCV [41]), found in the N-terminal domain of the NS3protein,arehighly conserved amongall HCV strainssequenced sofar and have been proposed to constitute the catalytic triad of the HCV protease (4, 7, 35).Consistent with this prediction, when the presumptive catalytic serine 1165 of the polyprotein was mutatedtoalanine, processingattheNS3-NS4A, NS4A-NS4B, NS4B-NS5A, and NS5A-NS5B sites was abolished (22, 42). Cleavageatthree of the four sites of the mutatedpolyprotein could be restored by supplyingfunctional NS3 intrans,while processingatNS3-NS4A occurredonlyincis(42).Thisfinding indicated that thecleavage of the latter site isan intramolec-ular event. Interestingly, the NS3-NS4A cleavage site differs from the others in that the processingtakesplace betweena threonine and a serine residue, whereas the remainder are cleaved between acysteine anda serine(22, 37).
Inflavivirus,NS3 isnecessary,butnotsufficient,for
process-ing at the nonstructural cleavage sites: in addition to the catalyticNS3domain,NS2B isrequiredforproteolytic activity (1, 6, 18, 33, 38). A stable physical interaction between NS3 and NS2B has been demonstrated (1, 18). Inthe caseof the othertwo genera of the family Flavivinidae, pestiviruses and HCV, it has been observed that the respective viral serine proteases,p80andNS3,canfunctionindependentlyof all the
regions of the polyprotein that are present upstream of the proteases themselves (22, 42, 45). However, in the case of bovine viral diarrhea virus (BVDV),amember of the
pestivi-rus genus, a region of the polyprotein outside p80 may be
necessarytoeffectcleavage bythe viral serineproteaseat the
p58-p75 site(45). Thisregionhastentativelybeenmappedto
aportionof thepolyprotein (13, 45)thatcorrespondstoHCV NS4B.
We initiated a study aimed to elucidate whether viral 3753
on November 9, 2019 by guest
http://jvi.asm.org/
functions other than NS3 are required for the proteolytic processingof thenonstructuralregion of theHCVpolyprotein. Using transient expression in HeLa cells of cDNAfragments that encoderegions of the HCVpolyprotein,we analyzed the cleavageactivityoftheNS3serineprotease oneach individual cleavage site. Inthispaper, we reportthat,inadditiontoNS3, a C-terminal domain of the NS4Aprotein is also required for efficient cleavage at the NS3-NS4A, NS4A-NS4B, NS4B-NS5A, and NS5A-NS5Bjunctions. In addition, we also show that NS4Acan act in trans to affectcleavage atthese sites.
MATERLALSAND METHODS
Cells and virus. HeLa cells, originally obtained from the AmericanType CultureCollection,were grown in Dulbecco's modified Eagle's medium containing 10% fetal calf serum (FCS). Vaccinia virus vTF7-3 (20) was grown in RK13 cells cultured in minimum essential medium containing 10% FCS. Construction of recombinantplasmids. HCV cDNAclones of the HCV-BK isolatewereusedfor allplasmid constructions (41). DNAfragments derived from HCVcDNA were inserted downstream of the 5' untranslated region of
encephalomyo-carditis virus and under a T7 promoter in the pCite-1 vector
(Novagen), in the appropriate translationalreading frame and followed by a termination codon. All the plasmids described belowwere obtained with standard recombinant DNA tech-nology (39).
Plasmid pCite(NS3) contains the portion of the HCV ge-nome between nucleotides 3351 and 5177 and therefore en-codestheportion of the viralpolyproteinbetweenamino acids 1007 and 1615. Construction of this plasmid has been de-scribed earlier(42).
pCite(NS4AB) bears a cDNA fragment corresponding to
nucleotides 5276 to 6225 of the HCV genome (amino acid residues 1649 to 1964 of the polyprotein).
pCite(NS5AB) contains the portion of the HCV genome
between nucleotides 6224and 9400(aminoacid residues 1965 to3010of the polyprotein).
pCite(NS4B/5A)wasobtainedbycloninginpCite-1 acDNA
fragment containing nucleotides 5653 to 7472.This construct
encodes the portion of the polyprotein between amino acid residues 1775 and2380.
pCite(NS3-AP) is a derivative of pCite(SX) (42) which contains HCVcDNA from nucleotides 3305 to 5373 (amino
acid residues 992 to 1680). A fragment derived from the
pBC12/RSV/APvectorandcodingforplacentalalkaline
phos-phatase (5)wasinserted downstream ofthis region,withcare
taken tomaintain thecorrect readingframe.
pCite(NS3/4A) contains a cDNA fragment comprising the
region between nucleotides3305and5465 ofHCV cDNA.The resultingconstructcodes for theportion of the HCV
polypro-tein between amino acids992 and 1711.
pCite(NS3/4A:S,
165-A)
encodes the same polypeptide,ex-ceptthat thecatalyticserine(S-1165)of NS3 has beenreplaced withan alanine.
pCite(NS3/4AAC) contains a cDNA fragment comprising
theregionbetween nucleotides 3305 and 5373 ofHCVcDNA.
The resultingconstructcodes therefore for theportion ofthe HCV polyproteinbetween amino acids 992 and 1680.
pCite(NS4A) bearsacDNAfragment spanningnucleotides
5276 to 5465 of the HCV genome(aminoacid residues 1649 to 1711).
pCite(NS4ABAN) contains the HCV sequence from nucle-otide 5367 to nucleotide 6225 (amino acid residues 1679 to
1964).
pCite(NS3AC)
is a 3-deletion product ofpCite(NS3) (42)
thatcontainsHCV cDNA from nucleotide3351 to nucleotide
4043 (aminoacid residues 1007to 1237).
Preparation of labelled extracts. HeLa cells seeded at a
density of 4 x 105 cells per 6-cm plate were infected with
vaccinia virus vTF7-3 at a multiplicity of 5 PFU per cell as
previouslydescribed
(42).
Afteradsorptionfor30 minat37°C,
3 ml of Dulbecco's modified Eagle's medium
supplemented
with 10% FCS was added. Cellswere incubated foran addi-tional30 minat
37°C.
Recombinantplasmid
DNAwasprecip-itated by the calcium phosphate method (39) and added
directly to each plate. When required, pUC19
plasmid
DNAwasused ascarrierDNAfortransfection.At 4h
posttransfec-tion, the medium was replaced with minimum essential
me-dium
lacking
methionine(GIBCO)
and thecellswerestarved for 1hat37°C.
Cellswerethen radiolabelled for 3 hwith 400jLCi of Tran35S-label
(ICN)
in 2 ml of minimum essential mediumlacking
methionine andsupplemented
with 2%dia-lyzed FCS. Cells were harvested and
resuspended
in IPB150(20 mM Tris-HCl
[pH 8],
150 mMNaCl,
1%Triton)
supple-mented with 1 mM
phenylmethylsulfonyl
fluoride,
1 mMEDTA, and 1 mMdithiothreitol.
Immunoprecipitations
wereconducted on denatured extracts with HCV
region-specific
rabbit
polyclonal
antisera as describedpreviously
(37, 42).
Analytical gel
electrophoresis
of theimmunoprecipitated
products was
performed
as describedpreviously (37).
Whennecessary,
quantitation
of radioactiveprotein
bandswascar-riedoutbydensitometric
scanning
of theautoradiograms
with the program Image(version 1.22)
on a Macintosh LC475personal computer equippedwith a
Sony
CCD videocamera.RESULTS
Expression plasmids and method of
analysis.
Previousstudies of HCV
polyprotein processing
indicatedthat NS3isaserine proteaseinvolved in the maturation of HCV
nonstruc-tural
proteins (3,
22,42). Although
it has been demonstrated that NS3 isabsolutely
required
for thecleavage
at theNS3-NS4A, NS4A-NS4B, NS4B-NSSA, and NS5A-NS5B
sites,
noevidence that the same
polypeptide
is per se sufficient forcleavage,
either in vitroor invivo,
has beenprovided.
Inorderto addressthismatter,weconstructedthe series of
expression
plasmids
describedinFig.
1.Appropriate fragments
of the HCV genome were cloned downstream of the T7
bacteriophage
promoterin thepCite-1
vector.Thisexpression
vector contains the
encephalomyocarditis
virus internalribo-some entry site
(27),
which promotes efficient andCAP-independent
translation of mRNA transcribed from the T7 promoter (16). In orderto obtain transientexpression
ofthe various portions of the HCVpolyprotein,
HeLa cells wereinfected with vTF7-3, a recombinant vaccinia virus that sup-ports the
synthesis
ofbacteriophage
T7polymerase
in thecytoplasmof infected cells
(20),
andwerethentransfectedwiththe
appropriate plasmids.
Cellproteins
weremetabolically
labelled with
[35S]methionine
as described in Materials and Methods. Viralproteins
andprocessing products
were then identifiedby
immunoprecipitation
of denatured whole-cellextracts with
region-specific polyclonal
antibodies to HCVNS3, NS4, and NSSA (37,
42)
and on the basis of theirmolecularweights.Itmustbe
pointed
out,however,
that in the absence of N-terminal sequenceanalysis
of eachprocessing
productwe cannot ruleoutthatnew
cleavage
siteshave beengenerated
inthegenetically
altered NS3 proteasesubstrates. NS4A isrequiredfor theNS3-NS4Acleavage.NS3generates the N terminus ofNS4Aby cleaving
itselfoff thepolyprotein
viaanintermolecularreaction(in cis) (42).
Weinitially
wantedon November 9, 2019 by guest
http://jvi.asm.org/
5' c
pCite(NS3) pCite(NS3AC) pCite(NS3/4AAC) pCite(NS3/4A)
pCite(NS3-AP) pCite(NS4A) pCite(NS4AB) pCite(NS4ABAN)
pCite(NS4B/5A) pCite(NS5AB)
pCite(NS3/4A:Scat-A)
E1 E2 NS2
190 383 810 1027
NS4
16581712 1973
Icat.A
FIG. 1. Schematic representation of the recombinant expression plasmids used in this study. The organization of viral polyprotein isshownat thetop.The vertical bars indicate theboundaries between the different proteins. The numbersatthetopof each bar indicate the position of the N-terminalamino acid of the following protein within the viral polyprotein.The HCV polyprotein portions expressed by the differentconstructs
areshownbelow; thenamesgiventothe corresponding plasmidsareshownonthe left. The rectangle marked AP indicatesaportion of placental
alkalinephosphatase.
tostudy whatviralstructuresarerequired for this intramolec-ularcleavagetooccur.
As a first step towards this goal, we analyzed the protein products of pCite(NS3/4A), a plasmid predicted to drive the expression of a polypeptide spanning the active protease
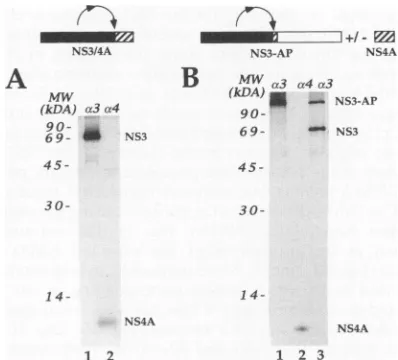
domain ofNS3 and endingatthe C terminus of NS4A (Fig. 1). As shown in Fig. 2A, transient expression of this plasmid in HeLa cellsyielded proteinsof70 kDa(lane 1) and 6 kDa (lane 2) that immunoprecipitated with NS3 and NS4 anti-sera,respectively. The NS3 protein generated by thisconstruct
migrates anomalously in comparison with wild-type NS3 be-cause it contains ashort stretchofunprocessed NS2at itsN terminus (42). The result of the experiment just described indicates that the NS3-dependent cleavage between NS3 and NS4A occurs efficiently in the absence of any other viral proteins.
We thenwished to determine what the minimalportionof NS4Arequiredforthisciscleavagetooccuris. To this end,we constructedpCite(NS3/4AAC),inwhich atermination codon wasinserted after the 23 N-terminalaminoacids of NS4A(Fig. 1). This latter plasmid yielded a protein, recognized by the anti-NS3antisera, thatcorresponds inmolecular weightto an uncleavedprecursor(datanotshown). However,theverysmall difference between the molecularweightof this latterprotein
precursor and that of mature NS3 made it very difficult to
establish whetherproteolytic cleavagewascompletelyoronly partially abolished. In order to resolve the possible cleavage products from theunprocessed precursor on sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), we decided toappendaheterologoussequence tothe N-terminal 23 residues ofNS4A. A cDNAfragment encodinga soluble form ofaplacentalalkaline phosphatase (5)wasthus fused in frame at the 3' end ofthe coding region of the pCite(NS3/ 4AAC) construct (Fig. 1). Transfection with the resulting plasmid, pCite(NS3-AP), resulted in the accumulation of a 130-kDa uncleaved proteinprecursor that wasrecognized by the anti-NS3 antiserum (Fig. 2B,lane 1).
NS3/4A
A
MW
(kDA) ct3A4
90_ ..
6 9- S NS3 45
-3
0-14
-NS4A
114~~~~+
A+/-KN53-AP NS4A
B
;DA} a3 a4 a3(kDA)
a_-
NS3-AP
90-69- d NS3
45-
30-
14-NS4A
[image:3.612.139.493.76.286.2]1 2 1 2 3
FIG. 2. Processingin cisatthe NS3-NS4Ajunction.Vaccinia virus vT7F-3-infected HeLa cellsweretransfectedasdescribed in Materials and Methods. Cellswere labelled with [5S]methioninefor 3h, and SDS-denatured lysates were immunoprecipitated with antisera to specific regions of HCV polyprotein. The positions of the relevant HCV proteins immunoprecipitated with anti-HCV antibodies are
indicated. a3 and a4 indicate antiseraagainstNS3 andNS4, respec-tively. MW,molecularweightstandards.(A) Lysatesfrom cells trans-fected withpCite(NS3/4A) wereimmunoprecipitated with anti-NS3 (lane 1)and anti-NS4(lane 2).Theimmunoprecipitated productswere
resolvedon anSDS-14% PAGE. (B) Lysates from cells transfected withpCite(NS3-AP)alone(lane 1)orcotransfectedwithpCite(NS4A) (lanes2and3)wereimmunoprecipitatedwith anti-NS3(lanes1 and3) and anti-NS4 (lane 2). The immunoprecipitated products were
re-solvedon anSDS-14% PAGE. NS5
2420
XI& XI& m m
il...'.. 3'
A B A B
NS3
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.346.545.408.588.2]A
2 \SS,:;S V NS4ABAN
NS4ABAN _ +/- E
NS3 NS4A
MW ax4 (kDA)
9 0
-a3 a4
69- NS3
-aY3 a4 MW a5A
(kDA)
9g0-NS3
B
VS4BI5A NS4B/5A
NtS4B/5A NS3 NS4A
aS3nSA
69 % N53- _ N53
-45- NS4B/5A NS4B/5A NS5A
NS5AB - +1
-NS5AB NS3 NS4A
a3a5A cx3 a aSA
-NS5AB
6 9- NS3 _ NS3
-- NS5A
45
-30- 30
-30- _ NS4ABAN - NS4ABAN
- NS4B
14 -1 4
-NS4A
2 3
1 4
-NS4A
1 2 3 4 5 6
NS4A
2 3
4 5
4 5 6
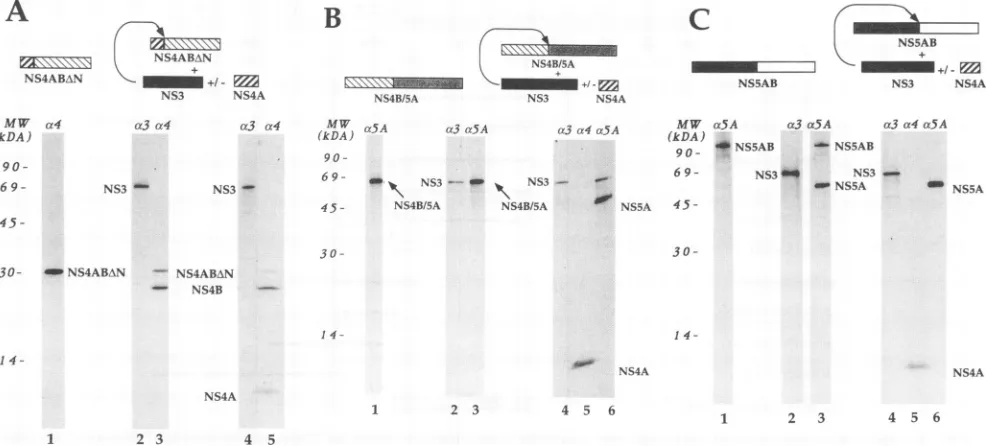
FIG. 3. Processing of the HCV polyprotein substrate intrans.(A)PlasmidpCite(NS4ABAN)wastransfected alone (lane 1), with pCite(NS3)
(lanes2and3),orwithboth pCite(NS3) and pCite(NS4A) (lanes 4 and 5). Cell lysateswereimmunoprecipitated with anti-NS3 (lanes 2 and 4)
andanti-NS4 (lanes 1, 3,and5). (B) PlasmidpCite(NS4B/5A)wastransfected alone (lane 1), with pCite(NS3) (lanes 2 and 3), orwithboth
pCite(NS3)and pCite(NS4A) (lanes 4, 5, and 6). Cell lysateswere immunoprecipitated with anti-NS3 (lanes 2 and 4), anti-NS4 (lane 5), and
anti-NS5A(lanes1and3). (C) Plasmid pCite(NS5AB)wastransfected alone (lane 1), with pCite(NS3) (lanes 2 and 3),orwithboth pCite(NS3)
andpCite(NS4A) (lanes 4, 5,and6).Cell lysateswereimmunoprecipitated with anti-NS3 (lanes 2 and 4), anti-NS4 (lane 5), and anti-NS5A (lanes
1, 3, and 6). c3, c4, and ox5A indicate antisera against NS3, NS4, and NS5A, respectively. The positions of the relevant HCV proteins
immunoprecipitated with anti-HCV antibodiesareindicated. MW,molecular weight standards.
The data described thus far suggest that an intact NS4A protein might benecessary for theNS3 protease to cleave at theNS3-NS4A site. There areessentially two possible expla-nations for this observation: either the integrity of NS4A is required to maintain the appropriate conformation at the NS3-NS4Ajunction,or NS4A acts as a cofactor forthe NS3 protease activity. Whereas it would be difficult to prove the formerhypothesis,wereasoned that ifthe latterwerethecase,
then wemight be able to observe cleavage at the NS3-NS4A boundary of the NS3/alkalinephosphatase chimeraproviding intact NS4Aintrans. We thereforetransfected, together with thepCite(NS3-AP) construct,aplasmiddrivingtheexpression
of intact NS4A [pCite(NS4A), Fig. 1]. The cotransfection
resulted in the production of the expected 6-kDa NS4A protein(Fig. 2B, lane2). More noticeably, in addition tosome
uncleaved precursor, a protein corresponding in size to the
expectedcleavage product (70 kDa;seeabove)wasspecifically recognizedby the anti-NS3 antisera (Fig. 2B, lane 3). These
results indicate that NS3 and NS4A are both required for
cleavage at the NS3-NS4Ajunctionand thatNS4A iscapable ofactingintrans to effectcleavage at this site.
NS4A is alsoan effector ofcleavage by NS3 at the
NS4B-NS5A and NS5A-NS5B junctions. In order to establish
whether NS4A is involved as a cofactor of NS3 protease
activity on the remaining cleavage sites, we constructed pCite(NS4ABAN), pCite(NS4B/5A), andpCite(NS5AB) (Fig. 1). Eachplasmid wasthen transfected aloneor cotransfected withpCite(NS3) aloneorwith thecombination ofpCite(NS3) andpCite(NS4A).
Transfection withplasmid pCite(NS4ABAN) results in the
synthesis of a protein of 30 kDa that is recognized by the
anti-NS4 antiserum(Fig. 3A,lane 1).Thisproteincontainsthe
C-terminal 33 residues of NS4Aand most of NS4B (Fig. 1).
Coexpression of the NS3 protein (Fig. 3A, lanes 2 and 3) resulted in thepartialproteolytic conversion of sucha30-kDa protein intoasmallerproductshowing electrophoretic mobil-ity similar to thatof theauthentic NS4B(26kDa[42]). When
NS4Awascoexpressedincombinationwith NS3(Fig. 3A,lane 4),wecouldobserveareproducible increasein theefficiencyof precursorprocessing (from -65 to -90%),in additiontothe 6-kDa protein encoded by the cotransfected pCite(NS4A) plasmid (Fig. 3A, lane5).
When we transfected plasmid pCite(NS4B/5A), the ex-pected 68-kDa protein productwas detected by immunopre-cipitation with anti-NS5A antiserum (Fig. 3B, lane 1). This protein containsmostof the NS4B and NS5Asequences(Fig. 1). Interestingly, whenNS3wascoexpressedwith thisprotein (Fig. 3B,lane 2),the same 68-kDa productwas immunopre-cipitated by the anti-NS5A antiserum and we found no evi-denceofproteolytic cleavage (Fig. 3B,lane3).However,when NS3wasexpressedincombination with NS4A (Fig. 3B,lanes 4 and5),wecouldobserve theoccurrence ofpartialcleavage ofthe 68-kDaproteinandtheproductionofa46-kDaprotein product that was recognized by the anti-NS5A antisera (Fig. 3B, lane6). It should bepointedouthere thatourpolyclonal anti-NS4 antiserumrecognizesNS4A farmoreefficientlythan NS4B. In particular, the portion of NS4B encoded by the pCite(NS4B/5A) construct appears not tobe immunoprecipi-tated, either when fused to NS5A or when cleaved off the precursor (Fig.3B,lane5;seealsoFig. 4,lane4, andFig. 5A, lanes 2 and5).
Last, we analyzed the activity of the NS3 protease on an HCVpolyprotein portion containingthecleavagesite between NS5A and NS5B. Plasmid pCite(NS5AB) supported the ex-pressionofa 115-kDaproteinthatwasspecifically recognized bythe anti-NS5A antiserum (Fig. 3C, lane1). WhenNS3was
45-Y3ut4a5A MW a5A (kDA WMNS5AB
- NS5A
1
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.61.554.78.301.2]coexpressed
with this latterprotein,
which contains the NS5A-NS5Bjunction (Fig. 3C,
lane2),
we observed partialproteo-lytic cleavage and production ofa polypeptide with the
elec-trophoretic mobility and immunological properties expected
for the mature NS5A (Fig. 3C, lane 3).This finding indicates
that the cleavage between NS5A and NS5B can occur in the
absence ofNS4A.However,when NS4Awascoexpressedwith NS3 andthe NS5A-NS5B precursor (Fig. 3C, lanes4 and 5), the
cleavage efficiency
increasedsubstantially, sincean NS5A-NS5B precursorwas no longerobservable (Fig. 3C,lane 6).NS4Aretains itsactivityintranswhen uncleavedfrom NS3. In a
previous
paper, we reported that the mutation of thecatalytic
serine of NS3 within thefull-length HCVpolyproteinabrogated processingat the NS3-NS4A, NS4A-NS4B,
NS4B-NS5A,and NS5A-NS5B sites(42). Wild-typeNS3 providedin
trans could
effectively
restore cleavage at the NS4A-NS4B,NS4B-NS5A, and NS5A-NS5B junctions but was unable to
cleave between NS3 and NS4A. This
finding
suggestedthat this lattercleavagecanonlyoccurincis,i.e., intramolecularly.Asa consequence, mature NS4A was not detectable byimmuno-precipitationinourtrans-complementation experiment.These
results are inapparent contradictionwith the data presented here: if the NS3proteaseneeds tobe activatedby NS4A, how could
processing
atseveral sitesberestoredby complementingthe mutated polyproteinwith NS3 alone and notwith NS4A?
A possible explanation of this paradox is that NS4A is still
capable
ofacting
asan effector of the NS3 protease in trans,even when uncleavedfrom the C terminus ofanonfunctional NS3
protein.
To test this hypothesis, we carried out theexperiments
described below.We inserted a mutation in plasmid
pCite(NS3/4A)
that resulted in thereplacementofthecatalyticserine of NS3 withan alanine. The mutated construct is called pCite(NS3/4A: S1
65-A)
(Fig. 1).
When transfected in vTF7-3-infectedHeLacells,
thisplasmid
drives the expressionofa 75-kDapolypep-tide
containing
both themutated NS3andNS4A(Fig. 4,lanes 3 and4).
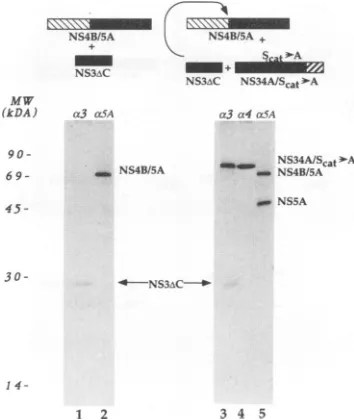
WecotransfectedpCite(NS3/4A:Sl
165-A) in combi-nation with pCite(NS4B/5A) and with pCite(NS3AC). The latterplasmid(Fig.
1)encodesaC-terminal deletionmutantof NS3 that retains full proteolytic activity (16a). The results of theexperiments
are showninFig.
4.Anti-NS3 antiserumimmunoprecipitatedtwomajor species,
of75 kDaand 29 kDa
(Fig.
4, lane3), correspondingrespec-tively
to the uncleaved NS3-NS4A precursor bearing the mutation in thecatalytic
serine of NS3 and to the active C-terminal deletion mutant of NS3. As expected, only the formerprotein
wasrecognized
bytheanti-NS4 antiserum(Fig.4, lane 4). Most interestingly, immunoprecipitation with
anti-NS5A antiserum revealed that the NS4B-NS5A protein had
undergone proteolytic cleavage (Fig.
4, lane 5) nearly asefficiently
asin theexperiment
showninFig.
3B,lane 6. These results suggest that NS4Adoesnot needtobe cleaved offthe NS3protein
in order to act as an effector of other NS3 moleculesintrans.The C-terminal portion ofNS4A is sufficient to act as an effector ofNS3
proteolytic activity.
The results described thus far indicate that NS4A isindispensable
for NS3cleavageatthe NS3-NS4A and NS4B-NS5A sites. In addition, NS4Aaug-ments
significantly
the rate ofproteolysis
at the NS5A-NS5Bcleavage
site.Conversely,
the rate ofcleavage of the proteinsubstrate encoded by the
pCite(NS4ABAN)
plasmid, which contains the NS4A-NS4Bjunction
but onlythe 33C-terminal residuesofNS4A,seems tobe littleaffectedbythe presence of NS4A. There are two possible interpretations of thisexperi-mental evidence: either the rate at which the NS4A-NS4B
cleavage
site ofthe HCVpolyprotein
isprocessed byNS3 doesNS4B/5A NS4B/5A +
NS3AC ~ ;
NS3iC NS34A/Scat EA
MW (kDA)
9 0
-6 9-45
-a3 cxSA4 a3a4a54
_NS4B/5A NS34A/Scat -A _ NBSA
-w NS5A
3 0
-
14-1 2 3 4 5
FIG. 4. NS4Aretains itsactivitywhen uncleavedfrom NS3. Plas-midpCite(NS4B/5A)wastransfected withpCite(NS3AC)(lanes1 and 2)orwith bothpCite(NS3AC)and
pCite(NS3/4A:S,
65-A) (lanes3 to 5).Cell lysateswere immunoprecipitatedwith anti-NS3 (lanes 1 and 3),anti-NS4(lane 4),and anti NS5A(lanes2 and5). 03,a4, and a.5A indicate antisera against NS3, NS4, and NS5A, respectively. The positions of the relevant HCV proteins immunoprecipitated with anti-HCV antibodiesare indicated. MW, molecularweightstandards.not depend on the presence of NS4A, or the C-terminal portion of NS4A that is contained in the protein encoded by thepCite(NS4ABAN) plasmidissufficientto act as aneffector ofthe NS3 proteolytic activity. In orderto address this issue, we performed the experiments described below.
As described earlier, pCite(NS4B/5A) codes for a protein precursorwhosecleavageisstrictly dependentonthepresence of the NS4Aprotein. We reasoned that if the C-terminal 33 amino acids ofNS4A were indeedsufficient to modulate the proteolytic activity of NS3, then the coexpression of the polypeptide encoded by the pCite(NS4ABAN) plasmid with NS3 should resultinthecleavageofthe NS4B-NS5Ajunction.
Thepropertiesof theproteinencodedby pCite(NS4ABAN)as
an effector of NS3 were thus compared with those of the
polypeptide encoded by plasmid pCite(NS4AB). This latter
plasmid contains, inaddition to the NS4Bregioncommonto
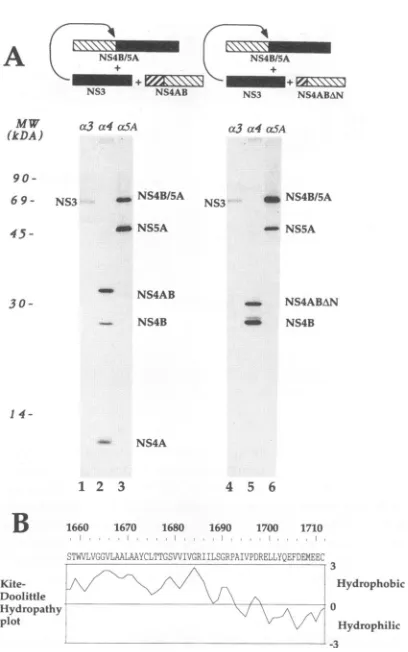
bothconstructs, the entire NS4Acoding region (Fig. 1). The results of an experiment in which pCite(NS4B/5A) was co-transfected with both pCite(NS3) and pCite(NS4AB) are shown inFig.SA, lanes 1 to3.Whenthe NS3protein (Fig.5A,
lane 1) is expressed in combination with the polypeptide
encodedby pCite(NS4AB), the latterwas partiallycleaved to
yield the two expected fragments of 26 and 6 kDa (Fig. SA,
lane2).Moreover,theNS4B-NS5Aproteinprecursor wasalso
efficiently processed (Fig. SA, lane3). Lanes4 to 6ofFig. 5A
show the results of similar cotransfection experiments, but in which pCite(NS4AB)wasreplaced bypCite(NS4ABzAN). The
expressionof the NS3 protease (Fig. 5A, lane4) in
combina-tion with theproductsofthepCite(NS4ABAN)construct(Fig. 5A,lane5;seealsoFig.3A,lane3)resulted in thecleavageof
thepolypeptide containingthe NS4B-NS5Ajunction (Fig. 5A,
lane 6). This last result suggests that a domain of NS4A contained within the 33 most C-terminal amino acids of the
protein may besufficient to modulate the activity of the NS3
protease.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.356.533.78.288.2]A NS4B/5A NS4B/5A
-+NS3i03NS\ANS4AB NS3S3+ NS4ABNNS4BN
MW a3a4mLSA
(kDA)
90
-6 9
-
45-30
-NS3 _ NS4B/5A
_ NS5A
- NS4AB NS4B
a5A
NS3 _NS4B/5A
-NSSA
-14
-NS4A
1 2 3
B
Kite-Doolittle
Hydropathv plot
1660 1670 1680
4 5 6 1690 1700 1710
Hydrophobic
.:0
- , Iydrophilic
[image:6.612.72.276.67.393.2]--3
FIG. 5. The C-terminal domain of NS4A is sufficient to activate NS3. (A) PlasmidpCite(NS4B/5A) was transfected withpCite(NS3)
and with eitherpCite(NS4AB) (lanesIto3)orwithpCite(NS4ABAN) (lanes 4 to 6). Cell lysateswere immunoprecipitated with anti-NS3 (lanes1and4),anti-NS4(lanes2 and5),and anti-NS5A(lanes3 and
6). c3, a4,and otSA indicate antiseraagainst NS3, NS4, and NS5A, respectively.Thepositionsof the relevant HCVproteins immunopre-cipitated with anti-HCV antibodies are indicated. MW, molecular
weight standards. (B) Hydropathy plotof NS4A. Theaverage hydro-phobicityindexwasobtained with thealgorithmofKyteand Doolittle withasearchlength of 8 amino acids (32).
DISCUSSION
The evidence presented here shows that, in our transient expression system,theproteasedomain of HCV NS3requires the participation of an additional virus-encoded protein, NS4A, in order to efficiently cleave the substrates contained withintheHCV polyprotein.NS4Acanactas aneffector of the NS3 proteolytic activity when supplied in trans, and further-more a 33-amino-acid domain located at the C terminus of NS4A seemstobe sufficient tocomplement the NS3 activity. Therequirement foratwo-component proteasethusseemsto
be commonto all members of the Flaviviridae(see below). In flaviviruses, NS2B is required to facilitate efficient pro-teolytic processing of the nonstructural region of the viral polyprotein by NS3 (1, 6, 18, 33, 38). In contrast to HCV NS4A,flavivirus NS2B is locatedatthe N terminus of NS3. In spite of theirdifferent topological arrangements, HCV NS4A and flavivirus NS2B share several features: both proteins are required aseffectors ofproteaseactivity,canfunction in cisor in trans, and are liberated from their respective catalytic domainsbyanintramolecularcleavage. A 40-amino-acid
seg-mentof NS2Bconservedamongallflaviviruses has been shown
tobesufficienttotrans-complement the NS3proteolytic activ-ity in dengue (17) andyellow fever (8) viruses. Ahydropathy
plot of the NS2B protein shows that this 40-amino-acid seg-mentisahydrophilic domain surrounded bytwohydrophobic
regions (17). Similarly, the 33-amino-acid region of HCV NS4A, identified as being sufficient for the activation of the NS3 protease, spansa hydrophilic, C-terminal domain and is
preceded byaveryhydrophobicN-terminalportion (Fig. 5B).
Thegeneticorganizationand theprocessingofthe nonstruc-tural region of the polyprotein of BVDV, a member of the
pestivirus genus, parallels closely that of the corresponding
region of HCV: thep80serine protease, which is the functional homolog of NS3, has been shown by Wiskerchen and Collett
(45) to be required for processing of the nonstructural
pro-teins. Cleavage bythis proteaseactivityattheNandC termini ofp80 itselfoccurs intramolecularly (in cis). Cleavage at the remaining nonstructural sites can occur in trans. The region
correspondingtoNS5 of flavivirus isprocessed,asinHCV,to
two distinct polypeptides, p58 and p75. The segment of the
polyprotein following p80, equivalenttoNS4,generatesasmall
N-terminal hydrophobic protein,
piO,
and possibly a 32-kDa protein that has not yet been characterized in detail. It iscurrently thought that p58 and p75 are the functional
ho-mologs of HCV NS5A and NS5B, whereas plO and the
uncharacterized 32-kDa factorcorrespondtoHCV NS4A and
NS4B polypeptides, respectively. Interestingly, p80 protease
activityalonewasshowntobeinsufficient for thecleavageofa
substratecontainingthep58-p75junction. Conversely, expres-sion of a construct encoding, in addition to the entire p80
protein, plO and part of the 32-kDa protein, led to efficient
cleavage of the same site. In view of these data and of the
evidencepresented here,we suggestthat BVDVplOis
neces-sary to complement the p80 serine protease activity and
therefore is the functional homologof HCV NS4A. plO had
previouslybeendisregardedas aBVDVprotease cofactor(13,
45). This conclusion was based on the observation that
pro-cessingofapolyproteinthat hadbeen mutated in thecatalytic
serine ofp80wasrescuedatseveralsitesby
trans-complemen-tation withaprotein containinganintact protease domain.In
thisexperiment,plOwas notgenerated,but the
corresponding
sequence remained uncleaved from themutantp80
(45);
thisfinding indicated that thep80-plO cleavage can onlyoccur in
cisandwastakenasanargument toruleoutthe involvement
ofplOasapossible p80cofactorrequiredforcleavage atthe
remainingsites.However, it hasemergedfrom theexperiment
described inFig.4thatHCV NS4Acan complementtheNS3 protease activity in trans even when uncleaved from an NS3 molecule that has been mutated in the catalytic serine. It is therefore alsolikely, by analogy,thatBVDVplOdoesnotneed tobe released fromamutatedp80inorderto actintrans as an
effector of functional protease molecules.
The mechanism by which HCV NS4A activates the NS3-contained serine protease is not yet clear. The two proteins
could associate inacomplex toform an activeheterodimeric protease.Indeed,astablecomplexbetween NS3anda
protein
containingNS4Aandpartof NS4B hasbeen observedin vitro
(25). A similar complex has been observed in flaviviruses
between NS3 and NS2B (1, 8). A related possibility is that NS4A is required to target NS3 to the correct cellular
com-partment for maximal protease activity. Alternatively, NS4A could act as a virus-encoded chaperonin aiding the correct
folding of the protease domain
(14).
In this regard, it is ofinterestthat severalbacterial serine proteases,including
ax-lytic
protease andsubtilisin,possesspropeptidesthat share several
properties with HCV NS4A and flavivirus NS2B: they are
necessary for proteaseactivity,canactivate the proteaseeither
on November 9, 2019 by guest
http://jvi.asm.org/
in cis orin trans, and are cleaved off the protease domain by intramolecular proteolytic reactions (26, 40, 47). These propeptidesare requiredtoassist the folding of theirprotease counterpart,and whenfolding has been completed the propep-tide becomes dispensable and no longer required for
proteo-lytic activity (2, 47). Alternatively, NS4A may act as a chap-eroninthat facilitates theinteraction of NS3 with the substrate, forexample by locally unfolding the polyprotein precursor, in order tomake the cleavage site accessible to theprotease.
In other viral systems, examples in which dimerization is required for protease activity (30) or in which the protease
activity is modulated by interaction with additional polypep-tides exist. In cowpea mosaic virus, the specificity of the 24-kDa protease for trans cleavage of the capsid precursor is altered bythe virus-encoded 32-kDaprotein (43). The3Cand 3CD proteases encoded by poliovirus differ incleavage speci-ficity forcapsid precursor, the specificity being determined by the interaction of the 3D region with the substrate (28, 46). Moreover, the adenovirus cysteine protease has been recently shown tobe activated via the formation of a disulfide bridge with an11-amino-acid peptide derived from the viral protein pVI (44).
Further studieson the structures and the functions of both HCV NS3 and NS4A, aswell asof their interactions with the other components of the viral polyprotein, are required to
solve the issue of how the virus-encoded serine protease is
activated by a second viral polypeptide. These studies will
eventuallyshed lightonthe temporal and spatialregulation of
HCV gene expression.
ACKNOWLEDGMENTS
WethankR. Cortese,J.Jiricni, andN. LaMonica forcritical review, and we thank all IRBM coworkers for helpful discussion. We also thank P. Neuner for oligonucleotide synthesis and Y. Cully for graphics.
REFERENCES
1. Arias, C. F., F. Preugschat, and J. H. Strauss. 1993. Dengue 2 virus NS2B andNS3 formastable complex that can cleave NS3 within the helicase domain. Virology 193:888-899.
2. Baker, D.,J. L. Sohl, and D. A. Agard. 1992. A protein-folding reaction underkinetic control. Nature (London) 356:263-265. 3. Bartenschlager,R., L. Ahlborn-Laake, J. Mous, and H. Jacobsen.
1993. Nonstructural protein 3 of the hepatitis C virus encodes a
serine-type proteinase required for cleavage at the NS3/4 and NS4/5 junctions. J. Virol.67:3835-3844.
4. Bazan,J. F., and R. J. Fletterick. 1989. Detection ofatrypsin-like
serine protease domain in flavivirus and pestivirus. Virology 171:637-639.
5. Berger,J., J. Hauber,R. Hauber, R. Geiger, and B. Cullen. 1988. Secreted placental alkaline phosphatase: a powerful new
quanti-tative indicator of gene expression in eukaryotic cells. Gene 66:1-10.
6. Chambers,T. J., A. Grakoui,and C. M. Rice. 1991. Processing of the yellow fever nonstructural polyprotein: a catalytically active
NS3 proteinase domain and NS2B are required for cleavages at
dibasicsites. J. Virol.65:6042-6050.
7. Chambers, T. J., C. Hahn, R. Galler, and C. M. Rice. 1990. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol.44:649-688.
8. Chambers, T. J., A. Nestorowicz, S. M. Amberg, and C. M.Rice. 1993. Mutagenesisof the yellow fever virus NS2Bprotein: effects
onproteolyticprocessing,NS2B-NS3complex formation, andviral replication. J. Virol. 67:6797-6807.
9. Chambers, T. J., R. C.Weir, A. Grakoui, D. W. McCourt, F. F.
Bazan, R. J. Fletterick, and C. M. Rice. 1990. Evidence that the
N-terminaldomain of nonstructuralproteinNS3from yellow fever virusis aserineproteaseresponsibleforsite-specific cleavages in theviral polyprotein. Proc.Natl. Acad. Sci. USA87:8898-8902.
10. Chien, D., Q.-L. Choo, A. Tabrizi, C. Kuo, J. McFarland, K. Berger, C. Lee, J. Shuster, T. Nguyen, D. Moyer, M. M. Tong, S. Furuta, M. Omata, G. Tegtmeyer, H. Alter, E.Schiff, L. Jeffers, M. Houghton, and G. Kuo. 1992. Diagnosis of hepatitis C virus (HCV) infection using an immunodominantchimeric polyprotein to capture circulating antibodies: reevaluation of the role of HCV in liver disease. Proc. Natl. Acad. Sci. USA 89:10011-10015. 11. Choo, Q.-L., G. Kuo, A. J. Weiner, L. R. D. W. Bradley, and M.
Houghton. 1989. Isolation of a cDNA clone derived from a blood-borne non-A non-B viral hepatitis genome. Science 244: 359-362.
12. Choo, Q.-I., K. H. Richman, J. H. Han, K. Berger, C. Lee, C. Dong, C. Gallegos, D. Coit, A. Medina-Selby, P. J. Barr, A. J. Weiner, D. W. Bradley,G. Kuo, and M. Houghton. 1991. Genetic organi-zation and diversity of the hepatitis C virus. Proc. Natl. Acad. Sci. USA 88:2451-2455.
13. Collett, M. 1992. Molecular genetics of pestiviruses. Comp. Im-munol. Microbiol. Infect. Dis. 15:145-154.
14. Craig, E. 1993. Chaperones: helpers along the pathways to protein folding. Science 260:1902-1904.
15. Eckart, M. R., M. Selby, F. Masiarz, C. Lee, K. Berger, K. Crawford, C. Kuo, G. Kuo, M. Houghton, and Q.-L. Choo. 1993. The hepatitis C virus encodes a serine protease involved in processing of the putative nonstructural proteins from the viral polyprotein precursor. Biochem. Biophys. Res. Commun. 192: 399-406.
16. Elroy-Stein,O.,T. R. Fuerst, and B. Moss. 1989. Cap-independent translation of mRNA conferred by encephalomyocarditis virus 5' sequence improves the performance of the vaccinia/bacteriophage T7 hybrid expression system. Proc. Natl. Acad. Sci. USA 86:6126-6130.
16a.Failla, C., and R. De Francesco. Unpublished data.
17. Falgout, B., R. H. Miller, and C. Lai. 1993. Deletion analysis of dengue virus type 4 nonstructural protein
NS2B:
identification of a domain required for NS2B-NS3 proteinase activity. J. Virol. 67:2034-2042.18. Falgout, B., M. Pethel, Y.-M. Zhang, and C.-J. Lai. 1991. Both nonstructural proteins NS2B and NS3 are required for the pro-teolytic processing of dengue virus nonstructural proteins. J. Virol. 65:2467-2475.
19. Francki, R. I., C. M. Fauquet, D. L. Knudson, and F. Brown. 1991. Classification and nomenclature of viruses. Arch. Virol. Suppl. 2:223-233.
20. Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss. 1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizes bacteriophage T7 RNA polymerase. Proc. Natl.Acad. Sci. USA 83:8122-8126.
21. Grakoui, A., D. W. McCourt, C. Wychowski, S. M. Feinstone, and C. M. Rice. 1993. A second hepatitis C virus-encoded proteinase.
Proc. Natl. Acad. Sci. USA 90:10583-10587.
22. Grakoui, A., D. W. McCourt, C. Wychowski, S. M. Feinstone, and
C. M. Rice. 1993. Characterization of the hepatitis C virus-encoded serine proteinase: determination of proteinase-depen-dent polyprotein cleavage sites. J. Virol. 67:2832-2843.
23. Grakoui, A., C. Wychowski, C. Lin, S. M. Feinstone, and C. M. Rice. 1993. Expression and identification of hepatitis C virus polyprotein cleavage products. J. Virol. 67:1385-1395.
24. Hijikata, M., N. Kato, Y. Ootusyama, M. Nakagawa, and K. Shimotohno. 1991. Gene mapping of the putative structural region
of the hepatitis C virus genome by in vitro processing analysis. Proc. Natl. Acad. Sci. USA 88:5547-5551.
25. Hijikata, M., H. Mizushima, T. Akagi, S. Mori, N. Kakiuchi, N. Kato, T. Tanaka, K. Kimura, and K. Shimotohno. 1993. Two
distinct proteinase activities required for the processing of a putative nonstructural precursor protein of hepatitis C virus. J. Virol. 67:4665-4675.
26. Ikemura, I., and M. Inouye. 1988. In vitro processing of pro-subtilisin in
Eschenichia
coli. J. Biol. Chem. 263:12959-12963. 27. Jang, S. K., M. V. Davies, R. J. Kaufman, and E. Wimmer. 1989.Initiation of protein synthesis by internal entry of ribosomes into the 5' nontranslated region of encephalomyocarditis virus RNA in vivo. J. Virol. 63:1651-1660.
28. Jore, J., B. D. Geus, R. J. Jackson, P. H. Pouwels, and B. E.
on November 9, 2019 by guest
http://jvi.asm.org/
Enger-Valk. 1988. Poliovirus protein 3CD is the activeprotease for processing of theprecursorprotein Plin vitro.J. Gen.Virol. 69:1627-1636.
29. Kato, M., M. Hijikata, Y. Ootsuyama, M. Nakagawa, S. Ohkoshi, T. Sugimura, and K. Shimotohno. 1990. Molecular cloning of human hepatitis C virus genome from Japanese patients with non-A non-Bhepatitis. Proc. Natl. Acad. Sci. USA 87:9524-9528. 30. Katoh, I., Y. Ikawa, and Y. Yoshinaka. 1989.Retrovirusproteinase characterizedas adimericaspartic proteinase. J. Virol. 63:2226-2232.
31. Kuo,G.,Q.-L. Choo,H. J.Alter,G. L. Gitnick, A. G. Redecker, R. H. Purcell, T. Myamura, J. L. Dienstag, M. J. Alter, C. E. Syevens, G. E. Tagtmeier, F. Bonino, M. Colombo, W.-S. Lee, C. Kuo, K. Berger, J. R. Shister, L. R.Overby, D. W. Bradley, and M. Houghton. 1989. An assay forcirculating antibodies to a major etiologic virusof human non-A non-Bhepatitis. Science 244:362-364.
32. Kyte, J., andR. F.Doolittle.1982. Asingle method for displaying thehydropathic character ofaprotein. J. Mol. Biol. 157:105-132. 33. Lin, C., S. M. Amberg, T. J. Chambers, and C. M. Rice. 1993. Cleavageat anovel site in the NS4Aregion bytheyellow fever virus NS2B-3 proteinase is a prerequisite for processing at the downstream4A/4Bsignalasesite. J. Virol. 67:2327-2335. 34. Matsuura, Y., and T. Miyamura. 1993. The molecularbiologyof
hepatitis C virus. Semin.Virol.4:297-304.
35. Miller, R. H., and R. H. Purcell. 1990. Hepatitis C virus shares amino acidsequencesimilaritywithpestivirusesand flavivirusesas well asmembers of twoplantvirus subgroups. Proc. Natl. Acad. Sci. USA 87:2057-2061.
36. Phillips, M. A., and R. J. Fletterick. 1992. Proteases. Curr.Opin. Struct. Biol. 2:713-720.
37. Pizzi, E.,A.Tramontano, L.Tomei,N. LaMonica, C.Failla,M. Sardana, T. Wood,andR. De Francesco. 1994. Molecular model ofthe specificitypocket of the hepatitis Cvirusprotease: impli-cations for substrate recognition. Proc. Natl. Acad. Sci. USA 91:888-892.
38. Preugschat, F., C.-W. Yao, and J. H. Strauss. 1990. In vitro processing of denguevirus type 2 nonstructuralproteins NS2A, NS2B,andNS3. J. Virol. 64:4364-4374.
39. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning:alaboratory manual, 2nd ed. Cold Spring Harbor Labo-ratoryPress,Cold Spring Harbor,N.Y.
39a.Santolini, E., G. Migliacchio, and N. La Monica. 1994. Biosynthe-sis and biochemical properties of the hepatitis C virus core
protein.J.Virol. 68:3631-3641.
40. Silen, J. L., and D. A. Agard. 1989. The lyticproteasepro-region doesnotrequireaphysicallinkagetoactivate theproteasedomain in vivo. Nature(London)341:462-464.
41. Takamizawa, A., C. Mori, I. Fuke, S.Manabe, S. Murakami, J. Fujita,E.Onoshi, T.Andoh,I. Yoshida,and H.Okayama. 1991. Structure and organization of the hepatitis C virus genome isolated from human carriers. J. Virol. 65:1105-1113.
42. Tomei, L., C. Failla, E. Santolini, R. DeFrancesco, and N. La Monica.1993.NS3isaserineproteaserequired for processing of hepatitisC viruspolyprotein.J.Virol. 67:4017-4026.
43. Vos, P., J. Verver, M. Jaegle, J. Wellink, A. V. Kammen, and R. Goldbach. 1988. Two viral proteins involved in the proteolytic processingof thecowpeamosaic viruspolyproteins. Nucleic Acids Res.15:1967-1985.
44. Webster, A., R. T. Hay, and G. Kemp. 1993. The adenovirus protease isactivated by avirus-coded disulphide-linked peptide. Cell 72:97-104.
45. Wiskerchen, M., and M. S. Collett. 1991.Pestivirusgene expres-sion: protein p80 of bovine viral diarrhea virus is a proteinase involved inpolyprotein processing. Virology184:341-350. 46. Ypma-Wong, M.F., P. G. Dewalt, V. H. Johnson, J. G. Lamb, and
B.L.Semler. 1988.Protein 3CD is the majorpoliovirus proteinase responsible for cleavage of the P1 capsid precursor. Virology 166:265-270.
47. Zhu, X.,Y.Ohta,F.Jordan,andM. Inouye.1989. Pro-sequence of subtilisin can guide the refoldingof denatured subtilisin in an
intermolecularprocess.Nature(London)339:483-484.