Copyright© 1994, American Society for Microbiology
Functional
Complementation
of Nuclear
Targeting-Defective
Mutants of Simian
Virus
40
Structural
Proteins
NORIO ISHII, AKIRANAKANISHI, MASAYASUYAMADA,t MICHAEL H. MACALALAD,ANDHARUMI KASAMATSU*
Department ofBiology and Molecular BiologyInstitute, University ofCalifomia,
Los
Angeles,
LosAngeles, California
90024Received 25 May 1994/Accepted 7 September 1994
Structural proteins of simian virus 40 (SV40), Vp2 and Vp3 (Vp2/3) and Vpl, carry individual nuclear
targeting signals,
Vp3198_2o6
(Vp2316-324)
andVpll,,
respectively, which are encoded in different readingframes of an overlapping region of the genome. How signals coordinate nuclear targeting during virion
morphogenesis was examined by usingSV40variants in which there is only onestructural geneforVplor
Vp2/3,nuclear targeting-defective mutantsthereof,VP2/3202TandVplAN5, ornonoverlappingSV40variants inwhichthegenesfor Vpl and Vp2/3areseparated,and mutant derivatives ofthegenecarryingeitherone or
bothmutations. Nucleartargetingwasassessedimmunocytochemicallyfollowingnuclearmicroinjection ofthe
variant DNAs. WhenVp2/3 and Vpl mutants with defects in the nuclear targeting signals were expressed
individually, the mutant proteins localized mostly to the cytoplasm. However, when mutant VP2/3202T was
coexpressed in the same cell along with wild-type Vpl, the mutant protein was effectively targeted-to the
nucleus. Likewise, theVpIAN5 mutant proteinwas transported into the nucleus when wild-type Vp2/3was
expressedin thesamecells. Theseresultssuggestthat while Vpl and Vp2/3have independent nuclear targeting signals, additional signals,suchasthosedefining protein-protein interactions,playaconcerted rolein nuclear
localization aiongwiththe nuclear targeting signals oftheindividual proteins.
During the late phase of simian virus 40(SV40) infection, cytoplasmically synthesized structural proteins,Vpl,Vp2, and Vp3, are transported into the nucleus within 60 min of their synthesis (23),where they assemble into maturevirions with SV40 minichromosomes (28). The fine structure of SV40
reveals thatthe virion is composed of 72 Vpl pentamers (1, 21), each of which is presumedtointeractwithonemolecule of
eitherVp2orVp3 (1, 21). Since theformation ofSV40 Vpl pentamersoccursinvitro(16),pentamerformationcanalsobe expectedtooccurin thecytoplasmsoonaftersynthesisduring SV40infection. The site atwhichVp2 andVp3interactwith
Vpl following synthesis is not known. They might be trans-portedindividuallytothenucleus,orthey mightinteract in the cytoplasm. On the basis ofgenetic andbiochemical data,we
have proposed that the viral structural proteins destined to form thematurevirion inthenucleus interact in thecytoplasm (19, 22).Asthe structuralproteinsareknowntocarry individ-ual nuclear targeting signals (as discussed below) which are responsible for their nuclear localization following their syn-thesis (7, 15, 17, 29, 30),this hypothesiscanbe tested experi-mentally. The phenotypic rescue of the targeting-defective Vpl, for example, is expected to occurwhen Vp2 or Vp3 is present.
Three structuralproteinsofSV40areencodedinapartially overlappingmanner on the genome. Thegenes of the minor structural proteins, Vp2 andVp3 (herein designated Vp2/3),
aretranslated in thesamereading frame,with theamino acid
sequence ofVp3corresponding to the carboxy-terminal two-thirds ofVp2.Thegenefor themajorcoatprotein, Vpl,begins
38 codons 5' totheVp2/3termination codonand is translated
*Correspondingauthor. Mailingaddress: Department ofBiology,
UniversityofCalifornia,LosAngeles,405Hilgard Ave.,LosAngeles, CA 90024. Phone:(310)825-3048. Fax:(310)206-7286.
tPresent address: Department of Animal Science, College of Agriculture, Kyoto University, Kyoto 606, Japan.
in a differentreading frame. Within the carboxy-terminal 35 residues ofVp2/3,thereexistaDNA-binding domain (6) and a Vpl-interactive determinant (15), in additionto theVp2/3
nucleartransportsignal (NTS)whichmapstoresidues 198 to
206ofVp3 (316to324 ofVp2). TheVp2/3 NTSisnecessary andsufficientto targettheproteinstothe nucleus(7, 9). The
nuclearlocalizationsignal (NLS) ofVpl hasbeen mapped by
deletion analysisofpoliovirus Vpl-SV40 Vplfusionproteins tothe amino-terminaleightresidues of Vpl(29),yetitcannot function as an independent NTS. When these residueswere conjugatedatthe carboxyterminustochickenserumalbumin,
theywereunabletopromote entryof the nonnuclearprotein
into the nucleus (5). How these two differentsignals, Vp2/3-NTS and Vpl-NLS, function in the nuclear targeting of the viralproteinsis notwell understood; hence,werefertothem
asnucleartargeting signals. Bothtargeting signals arewithin the overlapping region. As mutations in theoverlapping region
of Vp2/3 could inevitably introduce alterations in the Vpl
sequence,weconstructedanonoverlappingSV40(NO-SV40),
whoseVp2/3 genes are separatedfrom theVpl gene. In this study,wehave tested for thecomplementationof the nuclear targeting-defective Vpl (or Vp2/3) by Vp2/3 (or Vpl).
MATERIALSAND METHODS
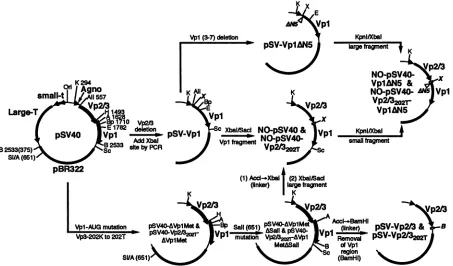
Plasmid construction. The strategy of the construction of the plasmids used in this study is diagrammed inFig. 1. All DNAmanipulationswere performed as described previously (25).
pSV40 containing the wild-type SV40 DNA inserted into
pBR322as ashuttlevectorhas beendescribed earlier (16). pSV-Vpl,in which theVp2andVp3codingsequenceswere deleted from pSV40, was constructed by ligating a 6.9-kb
KpnI-BpullO2I fragment and a 263-bpKpnI-AvaII fragment
frompSV40 with a225-bp AvaII-BpullO2I fragment
synthe-sizedby PCR.The 225-bp fragment,which lacks most of the 8209
on November 9, 2019 by guest
http://jvi.asm.org/
8210 ISHII ET AL.
K 294
Od
small-t| AII 557
L.arge-TVpt
B2533(375) s,By2533PCR
SI/A(651)
_r%%%%
x
E
Vpl
KpnID/bal
pSV-VpIMJ5 fragment
t
~~~/
~~NO-pSV40
VpIAN5 &
K NO-pSV40- Vpl
XV
Vp2t322r
Vp1IN5
0-PSV40
& PI KpnUXbalNO-pSV40- smallfragment
FH5ZZ A
(1)Accl-.Xbal (2) Xbal/Sacl
(linker) largefragment
K IK
Vp2I3 Vp24V213
AccI-.PBamHl
V4pl-AUJGmutatIon pSV40-AVplMet& Bp Sall(6S1) pSV40-AVp1MS (linker)_ S-p
V-32OKtO22T pS\40.V2rM2T- VP ASall&pSV40- 'Vpl pSV-Vp2/22 Vp3-202Kto202T aVmet P 1 MiUatboii VW2W
AVPI
Removalp2O2T
v rMev MetASall fBdVp1
S regIon
SI/A(65) (BamHl)
FIG. 1. ConstructionofSV40 andNO-SV40variant plasmidDNAs. Aschematicdiagram forthe construction of
pSV-Vpl,
pSV-VplAN5,
pSV-Vp2/3,PSV-VP2/3202T,NO-pSV40,NO-pSV40-VplAN5,
NO-pSV40-Vp2/3202T,
andNO-pSV40-Vp2/3202T-VplAN5
ispresented.All of the constructs aswellasthe intermediateplasmidsencodeSV40largeTantigen (Large-T),small tantigen (small-t),andagnoprotein (agno)and harbor all theregulatory elements(as shown forpSV40).Onlythepertinentportionof eachplasmid isshown. Thelocation for the deletion of the thirdtoseventh residues ofVplis markedbyaAN5.The numbersrepresent theSV40 nucleotidenumbers,andthose inparentheses
arethoseofpBR322. Abbreviations:Ori,originofreplication; A, AccI;All, Avall;B,BamHI; Bp,Bpu1102I;E,EcoRI; H, HindIII;K,
Kpnl;
Sc, SacI;SI,
Sall; X, XbaI.
Vp2/3region(SV40 nucleotides [nt]567to
1498),
was gener-ated by using pSV40 as a template and then digested withAvaIlandBpu1102I.Thesenseprimer,5'-GTCTlTTTATI1lC AGGTCCATGGTCTAGATGAAGATGGCCCCAACAA
AA-3', represents the SV40 sequence upstream ofnt567 and downstream ofnt 1948, and also includesanXbaIsite (under-lined)upstreamofthe startcodon ofVpl(boldfaceletters)to
facilitate subsequent manipulation. The antisenseprimer,
5'-CAAAGGAATTCTAGCCACACTGTAGCA-3', represents the SV40 sequence from nt 1792to 1766.
pSV-VplAN5containsa5-amino-acid deletion of residues 3
through7ofVpl, andwasconstructed byexchanginga288-bp
XbaI-EcoRI fragment of pSV-Vpl for a PCR-generated 273-bp
XbaI-EcoRI
fragment. The 273-bp fragment containing thedesignateddeletionwasgeneratedby usingpSV-Vpl as atemplate anddigested byXbaI andEcoRI.The sense primer
was 5'-TGCTCTAGATGAAGATGGCCGGAAGTTGTCC
AGGGGCA-3' (nt 563 to 1543 with deletion ofVp2/3coding sequences), and the antisense primer was as described above.
pSV-Vp2/3, in which only the Vp2/3 structural gene is present, and
PSV-VP2/3202T,
in which lysine 202 of Vp3 is mutated to threonine, were generated from pSV40-AVplMet and
pSV40202T-AVplMet,
respectively. Briefly,KpnI-Bpu
11021,
6.9- and 1.4-kb fragments of pSV40 were separated andthe 1.4-kbfragment was partially digested with HindIII toyield a 1.2-kb KpnI-HindIII fragment. The 6.9- and 1.2-kb
fragments were ligated together with a 217-bp mutated PCR
fragment togenerate pSV40-AVplMet and
PSV40202T-AVPl
Met, respectively. In these plasmids, two ATG codons at the
Vpl start region were eliminated by substituting two threo-nines (underlinedin the sequence thatfollows)without alter-ing the amino acid sequence ofVp2/3.Thesenseprimers,5'-T
AAAAGCTTACGAAGACGGCCCCAACAAAA(A/C)GA
AAAGGAAG-3' (nt 1490to1530),and the above-mentioned antisenseprimerwereused withpSV40as atemplateforPCR, and theresultingPCR fragmentsweredigestedwith HindlIl-Bpu1102Itoobtain the217-bp fragment.Inthesense primer,
the nucleotide sequence at the Vp3 lysine 202 (AAG) was
degenerate, leading either to the original lysine or to a
threonine substitution.Inordertofacilitate theuseof theAccI
site within the SV40 sequence, the AccIISalI site within
pBR322 (nt 651) of both constructs was destroyed to yield pSV40-AVplMetASalI andpSV40-Vp2/3202T-AVp1MetASalI, respectively, by digestionwithSall,followedby circularization after the repair reaction of the cleaved ends. BamHI linker
(5'-pCGGATCCG-3'; NewEngland Biolabs) was insertedat
the single AccI site of SV40 following the repair reaction.
Finally, pSV-Vp2/3 and
PSV-VP2/32O2T
were constructed by theremoval of the0.9-kb BamHIfragment containing the Vplcodingsequence.
NO-pSV40 and
NO-pSV40-Vp2/3202T
were constructed from pSV40-AVplMetASalI and pSV40-Vp2/3202T-AVplMetASalI, respectively, by inserting an XbaI linker (5'-pCTCTA
GAG-3'; NewEngland Biolabs) at the AccI site. The XbaI-Sacl fragmentfrompSV-Vpl, containing thefull-length Vpl
codingsequence, wasinserted into the XbaI andSacl sitesto
generate NO-pSV40 and
NO-pSV40-Vp2/3202T.
NO-pSV40-VplAN5 and NO-pSV40-Vp2/32o2T-Vp1AN5
J. VIROL.
PBI
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.80.533.75.341.2]NUCLEAR TARGETING OF SV40 STRUCTURAL PROTEINS 8211 Vpl-NLS p
Vp Vp2t3NTS
3 7
D2TI
I I
3 to 7 Vp2t32
SV40
SV-Vp1la
SV-Vp1ANS 4
S 20
SV-Vp2/3 I.
SV-Vp2/202T
100
so
NO-SV40O
NO-SV40-Vp1AN5 Q
4o0
Uf%-VAirVnI52---- o 20
M%P-v4u-VPifF4202T
NO-SV40-Vp2t3202r
[image:3.612.94.272.69.294.2]VpIAN5
FIG. 2. Lategenestructureof the variantSV40DNAs. The Vp2/3 (E) and Vpl(U)genes areindicated. Vpl and Vp2/3 nucleartargeting signalsaremarked (arrows)as openand solidsegments,respectively, within each of the coding sequences. The Vp2/3202T mutation is indicatedas awhitevertical line within the nuclear targeting signal, andtheVp1AN5 deletion is indicated byanopenarrowhead belowthe nuclear targeting signal. The numbers adjacent to the coding
se-quencesindicatethe lastamino acid.
were generated by exchanging the XbaI-SacI fragments with
those derived frompSV-VplAN5.
Allconstructionswereverified bydideoxynucleotide
double-stranded plasmid sequencing with Sequenase version 2.0 (United StatesBiochemical). Oligonucleotides forPCRwere
synthesized by the Preparation Laboratory of the UCLA
MolecularBiologyInstitute.
Cells,microin.jection, andplaqueassay. Culture conditions for TC7 cells, a subline of African green monkey kidney epithelialcells, andmicroinjection procedures havebeen
de-scribed previously (7). The derivatives of SV40 DNAs for
microinjection were obtained by digesting the plasmids at BamHIsitestoeliminate thepBR322 regionand
recirculariz-ing the DNAs (3.3 ,ug/ml)with T4 DNA ligase (16.7 U/ml).
After theligation reaction,the DNAswereconcentrated with a Centricon 30 microconcentrator (Amicon),
phenol-chloro-formextracted,ethanolprecipitated,andresuspendedin phos-phate-buffered saline at a concentration of 50 ng/,ul. DNAs
weremicroinjected into the nucleus ofTC7 cells whichwere
grown oncoverslipsmarked foreasylocationofinjectedcells. At this concentration, each cell is expected to receive about 100DNAmolecules. For immunofluorescencestudies, 100 to 200 cells were microinjected with each of the DNAson the
same coverslips and each experiment was repeated at least twice.The procedurefor theplaqueassayhas been described previously(32).
Indirect immunofluorescence. The rabbit and guinea pig
antisera against purified Vp3 and Vpl have been described
previously(8, 18). After the indicated incubationperiod, the microinjected cells were fixed in methanol-acetone (1:1) at
-20°C, and stained first with hamster anti-large T antigen (1:50), guinea piganti-Vpl (1:50), and rabbitanti-Vp3 (1:50)
and then with fluorescein isothiocyanate-,
tetramethylrhoda-A.SV40 B.SV-Vpl
C.SV-Vp2/3 D.NO-SV40
K
.*~
.- K0 4 8 12 16 20 24
hpmj
IC
41 21 W.
101
101
0 4 8 12 16 20 24
hpmj
FIG. 3. Timecourse forthe appearance of large T antigen, Vpl, andVp2/3 in the variantDNA-injectedcells.TC7 cells were microin-jected with SV40DNA(A),SV-Vpl (B), SV-Vp2/3 (C), or NO-SV40 (D),incubated,and fixed at the indicated times (hpmj). Proteins were visualized by staining first with hamster anti-large T, guinea pig anti-Vpl, and rabbit anti-Vp3 antibodies and then with fluorescein isothiocyanate-goat anti-hamster IgG, tetramethylrhodamine isothio-cyanate-goatanti-guinea pig IgG, and cascadeblue-conjugated goat anti-rabbitIgG.Percentexpressionrepresents eitherthe percentage of T-antigen-positive cellsas afraction ofcellsexhibiting large T-antigen staining (0) in total injected cells or the percentage of Vpl- or Vp3-positive cells as afraction of cellsexhibiting either Vpl (U)or Vp3staining(E)amonglarge-T-antigen-positive cells.
mine isothiocyanate-, and cascade blue-conjugated goat
sec-ondaryimmunoglobulinGs (IgGs) (1:50), respectively. In the triple-antigen detection procedure, we sometimes noted that the Vp3 cascade blue stainingwas difficultto unambiguously
distinguish from the T-antigen fluorescein staining. The sub-cellular localization of theviralproteinswasascertainedbythe
indirect immunofluorescent procedure using anti-Vpl and
anti-Vp3 (see thelegendsofFig.4and5). Washedcoverslips
weremountedontoglassslides and examined byfluorescence
microscopyasdescribedpreviously (7).
RESULTS
Construction ofSV40and its variant plasmids. The struc-turesof the late genes ofSV40and the variantDNAs used in
thisstudyarediagrammedinFig.2. Allconstructsharborboth
largeT-antigenand smallt-antigengenes,
regulatory elements,
and anintact agno gene. SV40variant
plasmids
pSV-Vpl
orpSV-Vp2/3 contain only one structural gene, Vpl or Vp2/3, respectively, andpSV-VplAN5 and
PSV-VP2/3202T
carry mu-tations in their nucleartargeting
signals.
InSV-VplAN5,
the3rd through 7th amino acids
(Pro-Thr-Lys-Arg-Lys)
of Vplwhich constitute the Vpl NLS weredeleted.
pSV-Vp2/3
andPSV-VP2/3202T,
which containsathreonine substitution atVp3
lysine 202 and has been shown to be critical for nuclear
localization, lack the Vpl coding sequence because ATG
initiating codons in the Vpl
reading
framewere mutated. Anonoverlapping
SV40 variantplasmid, NO-pSV40,
was alsomade to separate
Vp2/3
andVpl
coding
sequences with ashort spacerregionbetween thetwo
coding
segments.Nucleartargeting
signal
mutations inVpl
and/orVp2/3
wereintro-II m
-I1
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.329.549.77.278.2]8212 ISHII ET AL.
Vpl
SV40
SV-Vpl
SV-Vpl AN5
SV-Vp2/3
[image:4.612.152.465.72.532.2]SV-VP2/3202T
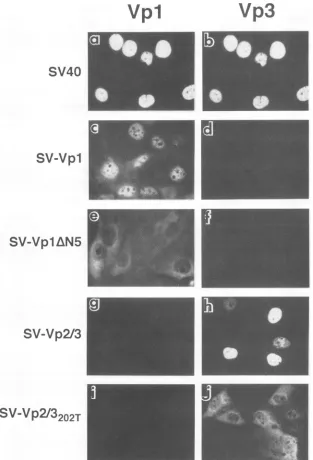
FIG. 4. Subcellular localization ofsingularly expressed capsid proteins. Cellsweremicroinjectedwithplasmid-derivedSV40DNA(aandb),
SV-Vpl (candd),SV-VplAN5(e andf), SV-Vp2/3(gandh),orSV-Vp2/3202T(iandj)asdescribedinMaterialsandMethods.Cellswerefixed 24hpmj, doubly stained firstwithguineapig anti-Vpl andrabbitanti-Vp3 antibodies andthen withtetramethylrhodamine isothiocyanate-goat
anti-guinea pig IgG andcascadeblue-conjugatedgoatanti-rabbitIgG.Panels a,c,e, g,and i representanti-Vplstaining,andpanelsb, d, f, h,and
jrepresentanti-Vp3stainingofthecorresponding field of cells.
duced to generate
NO-SV40-Vp2/3202T,
NO-SV40-VplAN5,orNO-SV40-Vp2/32o2T-Vp1AN5.
Time course for the viralgene products encoded by SV40 variantDNAs.Recircularized SV40 DNA as well as its variant DNAs, SV-Vpl, SV-Vp2/3, andNO-SV40, were microinjected into cells, and the appearance of large T antigen and the
corresponding structuralproteins was assessed cytochemically
as afunction ofthe time following injection (Fig. 3). In the SV40 DNA-injected cells, large T antigen was observed in
mostcellsby2hpostmicroinjection (hpmj), and Vpl and Vp3 were observedby4 to 5 hpmj (Fig. 3A). Most proteins were
observedtoaccumulatein the nucleus (see Fig.4a and b and
7). LargeTantigenexpressed from the variant DNAs followed
atimecoursesimilartothat ofSV40,whereas the fraction of cellspositive forVpl orVp2/3wasvariable,dependingonthe
constructs(Fig.3B, C, and D). In cells injected with SV-Vplor
SV-Vp2/3, aproportion of eitherVpl- orVp2/3-positive cells reached a plateau level after 24 hpmj (Fig. 3B and C). Less than half of the T-antigen-positive cells in
SV-Vp2/3-intro-duced cells showed Vp2/3 staining (Fig. 3C). Nonetheless, when the Vpl orVp2/3 staining was detectable at any given
time point, all staining was found unambiguously in the
Vp3
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
Vp1
NO-SVV40
NO-SV40-Vp1 AN5
NO-SV40-VP2/3202T
[image:5.612.157.471.70.462.2]NO-SV40-
VP2/3202T-Vp1AN5
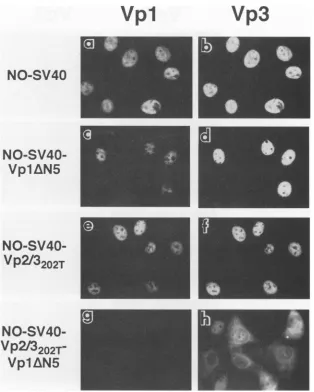
FIG. 5. Subcellular localization of capsid proteins expressed from NO-SV40 DNAs. Cells microinjected with NO-SV40 DNA (a and b),
NO-SV40-Vp1AN5 (c and d),NO-SV40-Vp2/3202T(e andf),or
NO-SV40-Vp2/3202T-Vp1AN5
(g and h) were processed at 24 hpmj as described inthelegendtoFig.4. Panels a, c, e, and g representanti-Vpl staining, and panels b, d, f, and h represent anti-Vp3 staining of thecorrespondingfieldof cells.
nucleus (Fig. 4c and h). No Vp2/3 or Vpl was detected in
SV-Vpl-orSV-Vp2/3-injected cells, respectively (Fig. 3B and C and 4d andg).NO-SV40 also showed efficientexpressionof
largeTantigenandthe structuralproteins,withVp2/3 expres-sionlagging behindthatof Vpl(Fig. 3D).On the basis of these
observations, all analyses were performed at 24 hpmj in the
following experiments.
Vp2/3 and Vpl nuclear targeting signals function in an
interdependentmanner.Theimmunostainingpatternsfor the
structuralproteinsinafieldof cellsareshown in
Fig.
4, 5,and6; and the distribution and subcellular localization of the individualproteinsaresummarized inFig.7. InSV-Vpl (Fig.
4c and d)- and SV-Vp2/3 (Fig. 4g and h)-injected cells, the
corresponding structural proteins
efficiently
localized to the nucleus(Fig.7)asinSV40DNA-injectedcells(Fig.4aandb),
demonstrating that SV40 Vpl and Vp2/3 contain their own nuclear targeting signals and are able to reach the nucleus
independently. In
SV-VplAN5-injected
cells, the truncatedVp1AN5
proteinwasfound in thecytoplasm(Fig.
4eand7),
as was theVp2/3202T
mutant protein inSV-Vp2/3202T-injected
cells(Fig. 4jand7).The alteredsubcellularstaining patternsin the cellsexpressingmutantproteins confirmedtheimportance
of theN-terminal amino acid stretch ofVpl (29)and thelysine
202residue ofVp3for their nuclear localization (7,
30).
To study how the independent Vp2/3 and Vpl nuclear
targeting signals encodedwithin the
overlapping
generegion
cooperate with each other in the process of the nuclear
targetingof viralproteins,we nextconstructedan
NO-pSV40,
in which the Vp2/3 and Vpl genes are
designed
to be in tandem. NO-pSV40 DNA, following the removal oftheplas-mid DNA, wasjust as capable ofplaque formation as SV40
DNA(datanotshown). UsingNO-SV40 and its variantDNAs,
weexamined the function of eachsignalinthe presenceof the
corresponding counterpart structural
protein. Vpl
andVp3
expressedfromNO-SV40localizedtothe nucleusas
expected
(Fig.5aand b and7).While themutant
VP2/3202T
inSV-Vp2/
3202T-injected
cellswasineffective in nucleartargeting(Fig.
4j),itwas effectively localized to the nucleus in the presence of full-length Vpl
(NO-SV40-Vp2/3202T)
(Fig. 5f
and7).
Like-wise,thetruncatedVplwas
effectively
targeted
tothe nucleusVp3
on November 9, 2019 by guest
http://jvi.asm.org/
8214 ISHII ET AL.
Vp1
Vp3
SV-Vpl
SV-Vp2/3
SV-Vpl
AN5
SV-Vp2/3
SV-Vpl
SV-VP2/3202T
SV-Vpl
AN5
Sv-VP2/3202T
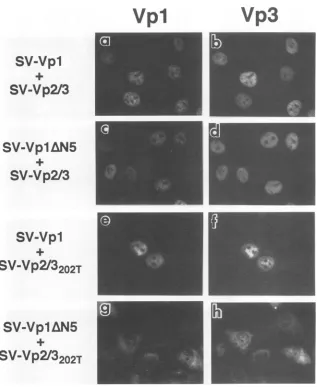
FIG. 6. SubcellularlocalizationofVpl andVp2/3 expressedfromacombinationof DNAs.CellsmicroinjectedwithSV-Vpl andSV-Vp2/3
DNAs(a and b),SV-Vp1AN5 andSV-Vp2/3(candd), SV-VplandSV-Vp2/3202T(eandf),orSV-Vp1AN5and
SV-Vp2/3202T
(gandh)were processedat24hpmjasdescribedin thelegendtoFig.4. Panels a, c, e, and g representanti-Vpl staining, andpanelsb, d,f, and h represent anti-Vp3 staining of thecorrespondingfield of cells.inthe presence ofVp2/3 (Fig.5c and7), althoughthemutant
Vplalonecouldonlypartiallylocalize tothenucleus(Fig.4e and 7). A similar complementation was observed when a
combination of DNAs, SV-VplAN5 and SV-Vp2/3 (Fig. 6c and d and7)orSV-Vpl and
SV-Vp2/3202T
(Fig.6e and f and7),wasintroduced into the cells, and the subcellular localiza-tion of the mutant viral proteins was indistinguishable from thatseenincells in whichthe wild-type Vpl gene andwild-type
Vp2/3 gene were coexpressed (Fig. 6a through f). Thus, the nucleartargeting defect in one structural protein was comple-mentedby the presence of another structural protein.
In the double mutant (NO-SV40-Vp2/3202T-Vp1AN5)-in-jected cells,the Vp2/3-NTS mutation maintained the SV-Vp2/
3202T phenotype (Fig. 7)
andshowedcytoplasmic
andperinu-clear fluorescence (Fig.
5h).
The truncated Vpl was notobserved in any compartment of the microinjected cells (Fig.
5g
and7).
Whenthetime course for the appearance of largeTantigen,
Vp2/3,
and Vplwasmonitored in thedoublemutant-introducedcells, large Tantigen and mutant Vp2/3, but not
Vpl,were clearly detected up to 24 hpmj (data notshown).
When thetwoSV40variantDNAs, SV-VplAN5andSV-Vp2/
3202T'
werecoinjected,
both mutantproteins
were observedprimarily in the cytoplasm (Fig. 6g and h and 7). Thus, the absence of Vp1AN5 in the double mutant-injected cells ap-pears to reflect a repression ofVpl synthesiswhen the two
genesarecontiguousonthe sameDNAmolecule.
DISCUSSION
Inthis study,the interrelationship of the nucleartargeting signals of SV40 structural proteins Vpl, Vp2, and Vp3 was
tested by using variant SV40s in which only one of the structural proteins was present or by using a NO-SV40 in which the genes forVpl and Vp2/3 are separated. NO-SV40
expressedlargeTantigen,Vpl, andVp2/3, producedaplaque,
and thus could complete a lytic cycle. In the variant SV40 DNAs, the nuclear targeting signal-defective mutant protein
Vp2/3202T
orVp1AN5,whenexpressed alone,remained in thecytoplasm but when awild-type copyof the other structural
proteinwassuppliedandexpressed,either in cisorin trans, the
mutant protein then localized to the nucleus. These results demonstrate thatwhile the nucleartargeting signaldetermines J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.147.464.70.457.2]the subcellular localization ofeachprotein and is responsible for the protein nuclear localization following its synthesis in
the
cytoplasm,
a functional defect of thesignalinonecapsid protein can be complemented by supplying the other capsidprotein
carrying anintactsignal.The presenceof at least one functionalsignal
in cis or trans is essential for the observedcomplementation.
Theseresults suggestthat additional signals suchasthosemediating
theprotein-proteininteractionsplayaconcertedrole
along
with the nucleartargeting signal duringSV40
morphogenesis.
In the study presented here, we usedanti-Vp3
which recognizes both Vp2 and Vp3 (18); thus, towhat extent the Vp3 staining represents Vp2 molecules in
which
118 residues unique to Vp2 including myristylatedglycine
(27)
at its aminoterminus cannotbe evaluated.There are a number of ways in whichspecific proteinscan
accumulate in the nucleus. The nuclear accumulation of
pro-teins
harboring
nuclear targeting signals hasbeen well docu-mented(11).
Proteinscontainingfunctionallyweaksignalscaneffectively
localizetothe nucleuswhenmorethanafewsignalsare present, as reported for theweakly active SV40
large-T-antigen
NLS-pyruvate kinase fusion proteins (24), defectivesignals coupled
to mouse IgG(20),
orbovine serumalbumin(12).
Proteins containingnonucleartargeting signalhave alsobeen shown to localize to the nucleus ifthey interact with
import-proficient
shuttleproteins
(for
areview,seereference14).
Forexample,
localization of the adenovirus DNApoly-merase is facilitated
by
another viralprotein,
pTP(33),
andNLS-deficient
subfragments
ofhepatitis
delta antigen canbetransported
into the nucleusby NLS-containing
full-lengthantigens
by
complexing throughleucine zipper sequences(31).Although
thecontributionofcooperative targeting by multiplesignals
and thepiggyback
transport toprotein
nuclearaccu-mulationhas been inferred
(14),
directproof hasbeenlacking.Our mutational
analysis
indicated thatonly
one wild-typenuclear
targeting signal
of the two is sufficient for the piggy-backtargeting
ofSV40proteins.
Reported
evidenceindicatesthatthehost transportmachin-eriesappeartoinfluencethe nuclear
targeting
of viralproteins
of
polyomaviruses.
Like SV40 structuralproteins,
Vpl andVp2/3
ofrelated murinepolyomavirus
possessdefined
target-ingsignals
that functioninmammaliancells(3, 4).
However,ininsect
cells,
polyomavirus Vp2/3
wasineffectively
targeted tothe nucleus but became
predominantly
nuclear inthe presence ofpolyomavirus Vpl
(10, 13).
BecauseVpl
effectively
accu-mulated in the
nucleus, Vpl-Vp2/3
interaction appears tomediate
Vp2/3
nuclear localization. Sincewhatwasobserved in hostcells
during
infectionby
polyomaviruses (10, 13, 19,
22,23,
26)
doesnot reveal the role of individualprotein
compo-nentsfor nucleartargeting,
these observations ofinsect cells(10, 13)
as well as of CV-1 cells via vaccinia virusvector-expressed
viralproteins
(26)
have been used to argue for a dominant role ofVpl
in nucleartargeting
ofVp2/3
inpoly-omavirus infection
(10,
13,
26).
In this report, we havecon-firmedourearlier observation that SV40
Vp2/3
isabletoreach the nucleusindependently
ofVpl
(7, 17). Currently,
whatcauses this difference other than the difference in hosts in which the recombinant viral
proteins
areexpressed
is not known.The
experimental
evidencepresented
here is in agreement with thehypothesis
that the SV40 subvirionassembly
takesplace
in thecytoplasm
(19, 22).
A similaridea
hasrecently
been
suggested
forpolyomavirus
on the basis ofprotein-protein
interaction studies(2)
and results obtainedby
coex-pression
ofpolyomavirus Vpl
andVp2/3
inmouse and insect cells(10, 13).
Whatremains tobe demonstrated is the identi-ties ofmutually
interactiveprotein
determinants and their roleDNA bod=.I
SV40O .----..f. aVp1
SV-Vp1---a-Vp'
SV-VpIAN5.---. a-Vpl
SV-Vp2/.--- aVp3 SV-VP2/3M2 .--- a-Vp3
NO-SV40
aVpl-a-Vp3
NO-SV40-VpI ---a-Vp{
a-Vp1-p
NO-SV-Vpl } P2 .{
a-Vp3
NO-SV40Vp2l't 3
{-a-Vp3--SV-Vp1M~ f.E a-vp3
sv-ps
'''~~'''''''a-Vpi!
SV-VpIAN5 w p a-Vpl
SV-Vp2M f a-vp3E
SV-Vpl0 --- CEaP3p SV-Vp2f3m0T --- CE-Vp3 SV-Vpl^N5E a-,Vpl
SV-Vp2O3mT2 ... a-Vp3
o 20 40 60 80 100
%PosiiveCells
FIG. 7. Subcellular distribution of Vpl and Vp2/3. Cells were injected with either individual variantDNAs or acombination oftwo variantDNAs asshownatthe left. Thenumber ofcellsexhibiting Vpl or Vp2/3 (100%) was classified as N (-) or C (O), which showed distinctnuclear orcytoplasmicstaining, respectively, and N>C ( )or C>N(u),inwhichstainingwasobservedinboth compartments with thestaining predominantly in the nucleusorinthecytoplasm, respec-tively. Cellswereprocessedasdescribed in thelegendstoFig.4to 6.
in the subvirion
assembly
andprotein nuclear targetingpro-cesses. Withinthecarboxyl35 residues ofSV40Vp2/3, three
signals, a Vp2/3 NTS, a Vpl-interactive determinant, and a
DNA-binding
domain, are present (6, 7, 15, 17, 30). Inpolyomavirus Vp2/3, a Vp2/3 NTS isat thecarboxyl end (4)
anda
Vpl-interactive
determinanthasbeenrecently mapped
withinresidues140to181 of theVp3(2). Polyomavirus Vp2/3
is shorter than SV40 Vp2/3 by 28 residues inwhich both a
Vpl-interactive
determinant and aDNA-binding
domain ofSV40
Vp2/3
arepresent.Thelocationfor theVpl-interactive
determinant of
polyomavirus Vp3
raisesapossibility
thatSV40Vp3
has an additional domain thathelps
function in theprotein-protein interaction withSV40Vpl.
Alternatively,
the interactivedeterminantpreviouslyidentifiedby
deletionanal-ysis
(15)
couldhave arisen fromconformationalchange
oftheproteinduetothe truncationof the
carboxyl
13 residues. The latterpossibility
hasbeensuggested by
Barouch and Harrison(2)
onthebasis oftheaminoacidhomology
found in residuesbetween170 and 181 of
polyomavirus Vp3
and 174 and 185 ofSV40
Vp3.
This,
however,
is ruled outby
results from ourlaboratory
indicating
that theVpl-interactive
determinant and theDNA-binding
domain ofVp2/3,
both of which occupy the last 13 residues ofVp2/3,
arefunctionally
separable
from each other. Mutantsthatgreatly impair
DNA-binding
activity
canstill interact with Vpl
(8a). Although
theVp2/3-interactive
determinant ofVpl hasnotbeen
identified,
itexcludes resi-dues 3 to 7 of Vpl, whose deletion resulted in the loss of nucleartargeting,
butwascomplemented by
Vp2/3
presumably
through
theprotein-protein
interaction. We canlearn
how additionalsignals
defining
theprotein-protein
interactioncon-77.
I I
I
FM
.m
MMM7
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.323.558.74.320.2]8216 ISHII ET AL.
tribute to the nuclear
targeting
ofvirionproteins
when theVp2/3-interactive
determinantofVpl
is identified.An
intriguing
observationwasthat when theNO-SV40-Vp2/32o2T-VplAN5
DNAwasinjected,
the truncatedVpl
wasnot detected in any compartment of the cells(Fig.
5g
and 7).Cytoplasmic Vp1AN5
andVp2/3202T
were detected in the samecells when the two genes on separate DNA molecules werecoinjected (Fig. 6g). Thus,
the observedVpl repression
occurred when the two mutant genes were
arranged
in cis. Since NO-SV40 isviable,
we assume that the amount ofallstructural
proteins
expressed
in the NO-SV40 DNA-intro-duced celliscomparable
to that in the normal SV40DNA-introduced cell. Whether the reduced level of mutant Vpl
observed in the double
mutant-injected
cellsreflectsan exper-imental artifact or aregulatory
role ofVp2/3
inVpl
produc-tion remains tobe tested.
ACKNOWLEDGMENTS
Weare
grateful
toJohnN.Bradyforsupplyinghamsteranti-SV40antiserum andtoEric Y.Chen and AndresL.Medinaforassistancein some ofthe stepsofplasmid construction.We also thank ArnoldJ. Berkand DavidA. Deanfor criticalreadingof themanuscript.
This workwassupportedbyPublic HealthService grantCA50574 and inpartbyfundsprovided bythe Committee onResearch of the AcademicSenate of theUniversityofCalifornia,LosAngeles.
REFERENCES
1. Baker,T.S.,J.Drak, and M.Bina. 1988. Reconstructionof the three-dimensionalstructureofsimian virus40and visualizationof the chromatincore.Proc.Natl. Acad. Sci. USA85:422-426. 2. Barouch,D.H.,andS. C.Harrison.1994.Interactionsamongthe
majorand minorcoatproteinsofpolyomavirus.J.Virol. 68:3982-3989.
3. Chang,D.,J.I.Haynes,J.N.Brady,and R. A.Consigli.1992.The
useofadditive and subtractiveapproachestoexaminethe nuclear localization sequenceof the polyomavirusmajor capsid protein
VP1.Virology189:821-827.
4.
Chang,
D.,
J. I.Haynes, J.N. Brady,and R. A. Consigli. 1992. Identificationofanuclearlocalizationsequenceinthepolyoma-viruscapsid proteinVP2.Virology191:978-983.
5.
Chelsky,
D.,R.Ralph,andG.Jonak.1989.Sequence requirementsforsynthetic peptide-mediatedtranslocationtothe nucleus. Mol. Cell. Biol.9:2487-2492.
6. Clever,J.,D. A.Dean,andH.Kasamatsu. 1993.Identificationof aDNAbindingdomain in simian virus 40capsidproteins Vp2and
Vp3.J.Biol.Chem.268:20877-20883.
7. Clever,J.,and H.Kasamatsu. 1991.Simian virus40 Vp2/3small structural proteins harbor their own nuclear transport signal.
Virology181:78-90.
8. Clever,J.,M.Yamada,andH. Kasamatsu.1991.Importof simian virus 40virionsthroughnuclear porecomplexes.Proc.Natl. Acad. Sci.USA88:7333-7337.
8a.Dean,D.A.,etal.Unpublisheddata.
9. Dean, D. A., and H. Kasamatsu. 1994. Signal- and
energy-dependent nuclear transport of SV40 Vp3 by isolated nuclei: establishmentofa filtrationassay for nuclearprotein import. J. Biol. Chem.269:4910-4916.
10. Delos,S.E.,L.Montross,R. B.Moreland,and R. L.Garcea. 1993.
Expressionofthe polyomavirus VP2and VP3proteinsin insect cells: coexpression with the major capsid protein VP1 alters VP2/VP3 subcellular localization.Virology194:393-398. 11. Dingwall, C., and R. A. Laskey. 1992. The nuclear membrane.
Science 258:942-947.
12.
Dworetzky,
S. I.,R. E. Lanford,and C. M. Feldherr. 1988. Theeffects of variations in the number and sequence of targeting signalsonnuclear uptake.J. Cell Biol. 107:1279-1287.
13. Forstova, J., N. Krauzewicz, S. Wallace, A. J. Street, S. M. Dilworth, S. Beard, and B. E. Griffin. 1993. Cooperation of structuralproteins duringlate events in the lifecycleof polyoma-virus. J. Virol. 67:1405-1413.
14. Garcia-Bustos, J., J. Heitman, and M. N. Hall. 1991. Nuclear protein localization. Biochim. Biophys.Acta 1071:83-101. 15. Gharakhanian, E., and H. Kasamatsu. 1990. Two independent
signals,anuclear localizationsignal andaVpl-interaction signal, reside within thecarboxy-35 amino acids of SV40 Vp3. Virology 178:62-71.
16. Gharakhanian, E., J. Takahashi, J. Clever, and H. Kasamatsu. 1988. In vitro assay for protein-protein interaction: carboxyl-terminal 40 residues of simian virus 40 structural protein Vp3 containadeterminant for interactionwithVpl.Proc. Natl. Acad. Sci.USA85:6607-6611.
17. Gharakhanian,E., J. Takahashi, and H. Kasamatsu. 1987. The carboxyl 35 amino acids of SV40 Vp3 areessential for its nuclear accumulation.Virology 157:440-448.
18. Kasamatsu, H.,andA.Nehorayan. 1979. Intracellularlocalization of viral polypeptides during simian virus 40 infection. J. Virol. 32:648-660.
19. Kasamatsu, H.,and A.Nehorayan.1979. Vplaffects intracellular localization ofVp3polypeptide during simian virus40 infection. Proc.Natl. Acad. Sci.USA 76:2808-2812.
20. Lanford,R.E., C. M. Feldherr, R. G. White, R. G. Dunham, and P. Kanda. 1990. Comparison of diverse transport signals in synthetic peptide-induced nucleartransport. Exp. Cell Res. 186: 32-38.
21. Liddington,R.C.,Y.Yan,J. Moulai, R.Sahli,T. L.Benjamin, and S. C. Harrison. 1991. Structure of simian virus 40 at 3.8-A resolution.Nature (London) 354:278-284.
22. Lin, W.,T.Hata,and H. Kasamatsu.1984.Subcellular distribution of viral structural proteins during simian virus 40 infection. J. Virol.50:363-371.
23. Lin, W.,J.L.Shurgot,and H.Kasamatsu. 1986. Thesynthesisand transportofSV40 structuralproteins. Virology154:108-120. 24. Roberts, B. L., W.D. Richardson, and A. E. Smith. 1987. The
effect ofproteincontextonnuclearlocationsignal function. Cell 50:465-475.
25. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning:alaboratory manual, 2nd ed. Cold SpringHarbor Labo-ratory,ColdSpring Harbor, N.Y.
26. Stamatos,N.M., S. Chakrabarti, B. Moss, and J. D. Hare. 1987. Expression of polyomavirus virion proteins by a vaccinia virus vector:associationof VP1andVP2 with thenuclear framework.J. Virol.61:516-525.
27. Streuli, C.H.,andE. Griffin.1987. Myristic acidis coupledto a structural proteinofpolyoma virus andSV40. Nature (London) 329:619-622.
28. Tooze, J. (ed.). 1981. DNA tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
29. Wychowski,C.,D.Benichou,and M.Girard. 1986. Adomainof SV40capsid polypeptideVP1thatspecifies migrationintothe cell nucleus. EMBOJ.5:2569-2576.
30. Wychowski, C.,D.Benichou,and M.Girard. 1987.The intranu-clear location of simian virus 40 polypeptides VP2 and VP3 dependsonaspecificaminoacid sequence. J. Virol. 61:3862-3869. 31. Xia,Y.-P., C.-T. Yeh, J.-H. Ou, and M. M. C. Lai. 1992. Charac-terization ofnuclear targeting signal ofhepatitis delta antigen: nucleartransportas aprotein complex.J.Virol. 66:914-921. 32. Yamada, M., and H. Kasamatsu. 1993. Role of nuclear pore
complexinsimian virus 40 nucleartargeting.J.Virol.67:119-130. 33. Zhao, L.-J., and R. Padmanabhan. 1988. Nuclear transport of adenovirus DNA polymerase is facilitated by interaction with preterminal protein. Cell 55:1005-1015.
J. VIROL.