JOURNAL OFVIROLOGY, Apr. 1976, p. 92-102 Copyright ©1976 AmericanSocietyforMicrobiology
Vol. 18,No.1
PrintedinU.S.A.
Host Range
Temperature-Sensitive
Mutants
of
Herpes
Simplex
Virus
Type
2
D. WESTMORELAND AND F. RAPP*
Department ofMicrobiology, The Milton S. Hershey Medical Center, ThePennsylvaniaStateUniversity, College ofMedicine,Hershey,
Pennsylvania
17033Received forpublication22September1975
Twosmall-plaquemutantsofherpes simplex virus type2(HSV-2) (strain333),
whosegrowthat39Cwasblockedincertain celltypes
(cell-dependent
tempera-turesensitivity), werecompared with theparentalvirus inanumber of biologi-cal assays. One mutant (no. 69) was found to produce a large number of morphologically normal, but noninfectious,particles; undernonpermissive
con-ditions, these mutant particles were able to interfere with the replication of
wild-type HSV-2. The other mutant (no. 74), which is known to belong to a
different complementation group, appeared to direct little virus DNA synthesis, even atthepermissive temperature. ProgenyproductionandvirusDNA synthe-sis incells infected bymutant 74 were delayed in comparison with wild-type virus-infectedcells. Bothmutants werefoundtobe moresensitiveto UV
irradia-tionthan theparentalvirus; this wasespecially marked in thecase of mutant
74. Moreover, this mutant wasfoundto haveahightransformingefficiency at
much lower doses of irradiation than those needed to abolish the cytopathic
effect ofwildtype HSV-2.
Herpessimplex viruses (HSV) are capable of
adiversity ofinteractions with a wide range of
host cells. The consequences of infection
in-cludecell death (2, 16, 35) with or without virus
progenyproduction, virus persistence in a
la-tent, nonreplicative form (8, 32, 38), and cell
transformation with very little expression of virus-specified functions (10-13, 26, 30, 40).
There is, as yet, little information about the
natureof host-orvirus-specified control
mecha-nismsthatdeterminetheresponsetoinfection.
Indeed, despiteintensive study (16-18), many of
the crucial steps in HSV replication remain
unclear. A particularly profitable means of studying virus functions is the use of
condi-tional, lethal, usually temperature-sensitive
(ts) virus mutants. Such mutants have been
successfully employedintheconstructionof the
genetic map of HSV (5, 14, 36, 37, 39), and
recently detailed examination of biological
le-sionsresponsible for thetemperature-sensitive
phenotypehasbeguninseverallaboratories(1,
3, 15). Theproducts of infection and the ability
of tsmutantstotransform cells at the
nonper-missive temperature have been areas of most
extensive effort (26, 40).
Recently, Koment and Rapp (22-24)reported
the isolation ofa group of
temperature-sensi-tive mutants with rather different properties
from thosedescribed above. The
temperature-sensitivephenotype of these mutants was only
apparent in certain types of cell, conferring
"host range" characteristics upon the group.
Virus sensitivity to high (39 C) temperature
wasdemonstratedinhamsterand mouse
fibro-blast cultures butwas much less apparent in
epithelioidcellsorhumanfibroblasts.
Believing that these mutants might provide
a tool in the elucidation of virus-cell
interac-tion, their behavior in a number ofbiological
tests was compared with that of the parental
virus. Thework described inthis
communica-tionisanextensionof earlier studieson twoof
thetemperature-sensitive host range mutants, carried out to determine more specifically the
nature ofthe defectin virusproduction under
nonpermissive conditions and to compare other biological properties of mutant and wild-type viruses.
Ofparticularinterest was the transforming
ability ofthemutants. Ithas beenestablished
inmanylaboratories that, if the cytopathic
ef-fect of HSV is destroyed, the virus has the
capacitytotransform cells from hamsters, rats,
mice, and humans (10-13, 26, 40). It was
thought that mutants whose capacity for
pro-ductivegrowth was genetically reduced might
more readily reveal their transforming
poten-tial, andthis viewissupportedby
transforma-tionby tsmutants undernonpermissive
condi-92
on November 10, 2019 by guest
http://jvi.asm.org/
tions, whichhasbeen described from other
lab-oratories (26,40, 41).
A quantitative assay for biochemical
trans-formation has recently been developed inthis laboratory using a selection system for cells bearing virus-induced thymidine kinase (TK) (19-21), based on cell growth in HAT medium (25, 30) (Rapp and Buss, Intervirology, in press). This technique was used to determine
whether themutantshadabnormal
transform-ingability andwhether transforming potential could be correlated with any other biological property.
MATERIALS AND METHODS
Cells. Hamster embryo fibroblast (HEF) cells wereprepared from 12-day-old LSH strain hamster embryos (Lakeview Hamster Colony, New Field, N.J.), as described previously (9), and were sus-pended inmedium 199 supplemented with 0.225% sodiumbicarbonate solution and 10%fetal calf se-rumplus 100 U ofpenicillinand 100
g.g
of strepto-mycin per ml(completemedium 199). Primary HEF cells were added to plastic petri dishes (60 mm), plastic tissueculture flasks (25 cm2), or glass pre-scriptionbottles (16ounce[ca.0.473liter])as appro-priate.The cellswereused when confluentmonolay-ershadformed.
Rabbitkidney(RK)cells werepreparedfrom the kidneys of 21- to28-day-oldrabbits by the technique previously described(11)andwereseeded into plas-ticpetridishes (60 mm) inEagle mediumplus
so-diumbicarbonate,antibiotics, and fetal calfserum atthe concentrationsused with medium 199 (com-pleteEagle medium).
Human embryonic lung (HEL) cells were ob-tained from FlowLaboratories,Rockville, Md.,and were maintained in complete Dulbecco medium (Dulbecco medium plus 0.225% sodiumbicarbonate,
10%fetal calf serum, andantibiotics,asdescribed). AlsomaintainedincompleteDulbecco mediumwere
BSC-1 African greenmonkeykidneycells and 3T3-4E TK-mousecells. The latterwerephenotypically thymidinekinaseless(TK-)3T3cells obtained from Howard Green(MassachusettsInstituteof Technol-ogy, Boston, Mass.).
Virus.A strainof HSV-2designated333,isolated
from agenitallesionbyW. Rawls(whenatBaylor CollegeofMedicine, Houston, Tex.) andpassaged only in human cells, was the parental wild-type HSV-2 used in thisstudy. Two mutants of HSV-2 strain333, whose isolation andpreliminary charac-terizationhavebeenreportedrecently (22-24),were examined. Onemutant(no. 69)wasderivedfroma single smallplaqueproducedinRKcellsbya prepa-rationofwild-typevirusthathad beensubjectedto 3.6 x 104ergsof UV irradiation percm2. Thesecond mutant (no. 74)was obtained from a single small plaqueon RKcellsproduced byanHSV-2 strain333
preparation that had been grown in medium
con-taining 20 Fgg ofbromodeoxyuridine per ml as a mutagen.
Both mutants and the wild-type virus were plaque-purified four times prior to use. Virus stocks were prepared from monolayers of HEL cells that had been infected, using an input multiplicity of 0.5 PFU/cell, and weremaintained at 33 C until exten-sivecytopathic effects became apparent (36 to 40 h postinfection). At this point, the infected cells were harvested by a single cycle of freeze-thawing and were disrupted by ultrasonication, and the lysate wascentrifuged at 200xg for 10 min.The superna-tant was transferred to small vials and stored at -70C as stock virus.
Virus assay. Virus was titrated in RK cells by a plaque assay that has previously been described in detail (31). Briefly, confluent monolayers of RK cells inplastic petri dishes (60 mm) were inoculated with 0.1 mlof virus serially diluted in Tris buffer. After anadsorption period of 1 h at room temperature, 5 ml of complete Eagle medium containing 0.5% methylcellulose was added to each plate. Cultures wereincubated for 4 days at 33 C in a moist atmos-phereof 5%CO2inair, attheend of which time the cells were fixed in 5% formalin and were stained using 0.03% methylene blue solution, and virus plaques were counted with the aid of a dissecting microscope.Plaque diameter was measured with the aid of a Nikonprofile projector(model 6C) at x10 magnification; at least 200 plaques from not less than four plates were measured for each virus-cell system.Plaque "diameter" wasalwaystaken at the widest point inahorizontal direction.
Assay for DNA synthesis. The method used for DNAanalysis was similar to thatdescribed earlier by Crouch and Rapp (6). Confluent monolayers of HELcells in plastic bottles (25cm2)weretreated for 48hpriortothe experiment withcompletemedium 199,which had a reduced concentration of fetal calf serum(1%)toreduce cell DNA synthesis. The cells were then infected at an input multiplicity of ap-proximately5PFU/cellin 0.1mlof Tris buffer. After adsorption, the monolayers were rinsed with Tris buffer,and 5 ml ofcompletemedium 199 containing only2%fetal calfserum wasaddedtoeach bottle.
The incorporation of[methyl-3H]thymidine dur-ing8-h pulses wasassayed atdifferent times after infection. [Methyl-3Hlthymidine at aconcentration
of10,&Ci/ml(45Ci/mmol)wasaddeddirectlytothe
nutrient medium and, at the end of the labeling period,mediumwasdecanted from thecells, which were dislodged fromthe bottle by freeze-thawing. Cellsthathadbecomedetachedfrom themonolayer due to viruscytopathiceffectswererescued from the culture mediumbyasinglelow-speedcentrifugation andwereaddedbacktotheappropriatebottle. Har-vestedcells weredigestedfor a minimum of4h at
37C inamixtureof0.6mlof Pronase(10mg/ml);0.3
mlofSarkosyl (10% solution), and 0.3 mlofEDTA
(0.2 M, pH 8.1) in 0.015 M sodium chloride and 0.0015Msodiumcitrate (0.1x SSC),pH7.3.
Mock-infectedcultureswere alsousedtoprovide
a '4C-labeledDNAmarker in cesiumchloride gra-dients. Such culturesweretreatedbytheprocedure describedabove, except that[methyl-'4C]thymidine
at aconcentrationof0.1
;&Ci/ml
(54;&Ci/mmol)
re-placed[methyl-3H]thymidine, and thecultureswere
93
on November 10, 2019 by guest
http://jvi.asm.org/
94 WESTMORELAND AND RAPP
incubated for24hpriortoharvestingand digestion. After digestion, 0.3 ml of[3H]thymidine-labeled
material and0.1mlof[14C]thymidine-labeled
mate-rialwereaddedto3.8ml of cesiumchloride solution in 0.1x SSC (density = 1.7936 g/cm3) and
centri-fuged inaBeckman L3-50 ultracentrifugeat35,000 rpmfor60h usinga no.40-3fixed-anglerotor.After centrifugation, 8-drop fractionswerecollected from
the bottom of the gradientontoWhatman filter pa-perdisks. Thediskswerewashed threetimesin5%
trichloroacetic acid, once in 95%ethanol, andonce inacetone;theywereallowedtodryatroom temper-ature and were placed into 10 ml offormula 949
toluene-basedscintillation fluid (New England Nu-clear Corp.). Acid-insoluble radioactivity was
countedwithaBeckman LS-250 liquidscintillation counter,whichcould distinguish thetworadioactive labels. Countsinboth channelswere printedonto tapeand the gradient profileswereplotted with the
aid of a Hewlett-Packard 9820A calculator and
9862Agraph plotter.
UV irradiationof virus. Virus (diluted in phos-phate-buffered salinetoaconcentrationof107PFU/
ml)wasirradiated in plastic dishes(60 mm)witha
Thomas UV lamp withaGE-G8T5 bulbatadistance
such that the incident dosewas about4,200 ergs/s percm2.Irradiated viruswastitrated in RKcells,as
described above.
Virustransformation assay. Theability of wild-type and mutant viruses to mediate "biochemical transformation" of 3T3-4E TK- cells to TK+ cells
wasassayed by using selection forTK+cellsin HAT
medium (7, 25, 30). Mutant and wild-type virus stockswereinactivatedbyexposuretoUV light for differentperiods of time. Trypsinized 3T3-4E
TK-cellsinsuspension (4 x 106)weretreatedwith 2 x
106PFU ofirradiated virus (calculated from preirra-diation titer) in 4.0 ml of complete Dulbecco me-dium. The mixturewasshakenatroomtemperature for 2h, afterwhich0.3-mlaliquotsweredispensed
intoeach petri dish(12by60mm)containing4mlof complete Dulbecco medium and were incubated at 37C. Within 16 to 24 hafterplating, the Dulbecco medium wasreplaced with 5 mlof complete Dul-becco containing aminopterin (0.4 ,Ag/ml), glycine (10 ,ug/ml),hypoxanthine (15jLg/ml),andthymidine
(10 gtg/ml) (HAT medium). This medium was
re-placedeverythirdday for18days and,onday 21,the
plates were fixed using 5% formalin and were
stained in 0.3% methylene blue for 20 min. After staining, colonieswerecounted and themean
num-ber of coloniesperplatewascalculated.
Assayfor defective interfering particles. (i) Con-fluent monolayers of HEF cells in bottles (16 ounce)
wereinfectedatdifferentinput multiplicities,with aconstant amountofwild-typevirus andincreasing dilutions ofmutantorwild-type"interferingvirus."
After adsorption at room temperature, the cells
wereincubatedat39Cincompletemedium 199 for
30h,atwhich timethecellswereharvested and the
viruswasextracted and titrated,asdescribed above.
(ii) A constant amount of wild-type virus and increasing dilutions of "interfering" wild-type or mutant virus were mixed in a 0.1-ml quantity,
which was then addedto confluent monolayers of HEFcells in plates(60mm).After adsorption for1h
atroomtemperature,thecellswereincubatedat33 or 39C for 4 days under complete Eagle medium
containing 0.5% methylcellulose. After4 days the cellsheetwasfixed, stained, and examined for virus
plaques, asdescribed above.
Particle counts by electron microscopy. Virus
suspensions in medium 199wereprepared for elec-tron microscopy by the technique of Monroe and Brandt (29) andwereexaminedat75kVinaHitachi
HU12 electron microscope. Virus particles were
scoredas enveloped or nonenveloped andcoredor
emptied of any electron-dense core material. All
samples for electron microscopy were blind-coded
priortoexamination.
RESULTS
Host range, temperature sensitivity, and
plaque morphology. Theresultsofplating
ex-periments in different cell types, several of
which arein accordwithpreviously published
data (23, 24),aresummarizedin Table1.Plaque
size,although heterogeneous, fellwithina
sin-gle population for each virus in each cell type. Mutants 69 and 74, which produced smaller
TABLE 1. Plating efficiencies and mean plaque size at 33 and 39 C of wild-type HSV-2 strain 333 and of
mutants 69and 74a
Wild-type333 Mutant 69 Mutant74
Hostcell Plating effi- Mean plaque Platingeffi- Mean plaque Plating effi- Mean plaque ciency39C/ size (mm) ciency 39C/ size(mm) ciency 39C/ size (mm)
33C(%) 33C 39C 33C(%) 33C 39C 33C (%) 33C 39C
HEF 10 3.3 1.3 3.3 x 10-4 1.8 4.0 x 10-5 2.0 1.7
RK 100 2.5 2.0 10 2.0 0.5 15 1.5 0.5
HEL 100 2.5 2.8 11 2.4 0.6 30 2.4 0.8
BSC-1 (African 57 0.9 0.9 23 0.6 0.5 33 0.9 0.4
green monkey kidney line)
3T3-4E TK- <10-3 1.4 <10-2 0.9 <10-3 0.9
(mousecell line)
aTheplating efficiency ofvirus stocks(grownin HEL cells) wascomparedin different cell types, using the plaque assay
technique.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
RANGE TEMPERATURE-SENSITIVE
plaques than parental 333 under all
experimen-talconditions, were found to express a
tempera-ture-sensitive phenotype that was hostcell
de-pendent. Moreover, failure of the mutants to grow at 39 C was not merely due to a prolonged
virus growth cycle since infected HEF cells
could be maintained for up to 9 days at 39 C
without adetectable increase in plaque
num-ber. Mutant virus growth at 39 C was due to
leakage, rather than reversion to wild type.
Very little wild-type or mutant virus growth
wasobserved in 3T3-4E TK- cells at 39 C.
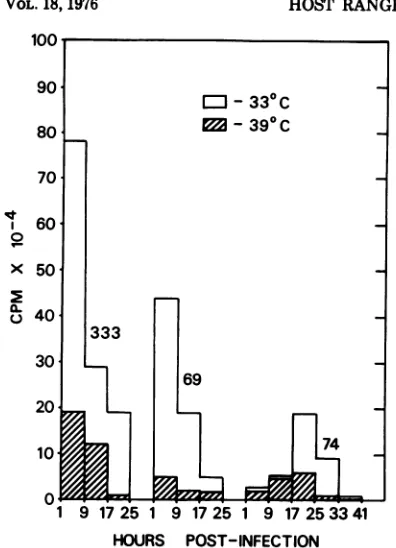
Figure 1 shows growth curves over the first
24hafter infection of wild-type 333, mutant 69,
andmutant 74 inHEFcells. The results further
demonstrate the temperature sensitivity of
both mutants in HEF cells and also suggest
that maturation of mutant 74 proceeds at a
slowerratethanthat of wild-typevirus,evenat
33C.
DNA studies. Because of their different
den-sities, HSV-2 DNA (density = 1.73g/cm3) (35)
andcellular DNA(density= 1.70g/cm3)canbe
differentiated by isopycnic bandinginCsCl
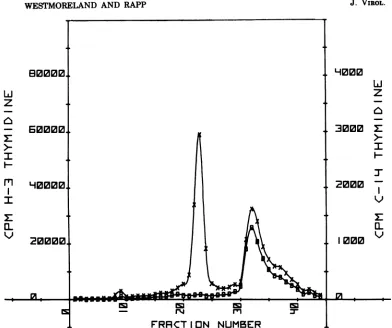
so-lution, as shown in Fig. 2. For each profile, the
total amount of [3H]thymidine incorporated
into virusand cellularDNAwascalculated by
summingthe radioactivityinthefractions
com-prising the virus and cell DNA peaks,
respec-tively.
The pattern ofvirus DNA synthesis during
the infectious cycle in HEF cells is shown in
Fig. 3 forwild-type 333 and the two mutants.
Wild-type virus was observed to synthesize
mostofitsDNAduring thefirst9hafter
infec-tion, a findingthat is consistent with earlier
reports (2, 34, 35). Thiswasthecasebothat 33 and 39C, although atthe highertemperature
less virus DNA was made, supporting the
slight temperature sensitivity of HSV-2 in
hamster cells(6).
Virus DNA synthesisin HEF cells infected
by mutant 69 occurred predominantly during
the first 9 h postinfection but, even at 33 C,
mutant 69 directed only half as much viral
DNA synthesis as did the wild-type 333. At
39C the discrepancy between wild-type and
mutant69DNAsynthesisisgreaterthan two-fold, but some mutant virus DNA synthesis
wasobserved.
Theresults obtainedsuggestthat mutant 74
directed relatively little virus DNA synthesis
(comparedwiththat directedbywild-typevirus ormutant69) for the first16h after infection of
HEF cells, although some progeny production
could be detectedatthistime.ThepeakofDNA
synthesis occurred between17 and25h
postin-fection,atboth33and39C. Considerablymore
virusDNAwasmadeatthe lowertemperature,
101
-j 5
x 10
101
101
0 4 8 12 16 20 24
HOURS POST-INFECTION
FIG. 1. Growth curves of HSV-2 strain 333, mu-tant69, andmutant 74 in HEFcells at 33 and 39 C. Cells were infectedat an input multiplicity of5.0 PFUlcell and were harvested, virus extracted, and
titrated,asdescribed.
but this amounted to only about 30% of the
amountofwild-type virus DNAmadeby HEF
cells infected under similar conditions.The
ob-servationthatmutant 74makesprogenyDNA
"late" intheinfectious cycleisconsistent with
the previous observation that the replication
18, 95
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.297.419.68.536.2]J. VIROL.
[image:5.509.72.464.54.382.2]96 WESTMORELAND AND RAPP
FIG. 2. Separation ofvirusand cellular DNA byCsCI equilibrium centrifugation. Thisrepresentative profile shows virus and cell DNAfromacultureofHEF cells that had beeninfectedwith HSV-2 strain 333 17 hpreviouslyand incubated at39C. [3H]TdR wasaddedat 9 hpostinfection, asdescribed. Symbols: x,
radioactivity profile obtainedfrom infected cells; 0, radioactivity profile from mock-infected cell sample labeled with[14C]TdR, whichwascocentrifugedwith materialfrom infectedcells.
cycle of the virus is longer than that ofmutant
69ortheparentalvirusat33 CinHEFcells.
Electron microscope studies. Theprogenyof infection of HEF cells (at 33 and 39C)by
wild-typeHSV-2 333 and bymutants69and 74were
harvested 24 h after infection andwere
exam-ined in the electron microscope. Virus with a
herpes-like morphology was seenin all
prepa-rations andwassubdividedinto particles with orwithoutanelectron-densecore(denoting the presence of virus DNA) and withorwithout a
distinguishableenvelope.
Itsoonbecameapparent, asshown inTable
2, whichrepresentsthe results fromanumber
of experiments, that even at the permissive temperatureprogenyofinfection bymutant69
hada veryhigh ratio ofparticletoinfectivity. It
was earlier indicated (Fig. 1) that 24 h after
infectionat33C theprogenytitersof infections
by wild-type 333 andmutant69werenot
sub-stantially different; thehigh ratio ofparticleto
infectivity indicates that cells infected by
mu-tant69produce alarge number ofvirus
parti-cles, mostof which are notinfectious. Similar
observations weremade when mutant 69 was
grown at 39 C in HEF cells, when ratios of
particletoinfectivityashighas amilliontoone
were notunusual. (The infectivity ofprogeny virus wasalwaysassayedat33CinRKcells.)
Theparticles producedbymutant69-infected
cellswereexamined morecloselytodetermine
whetherthe vastnumber of defective particles
was morphologically abnormal. No difference
wasfound betweenthe ratioofcored(probably
DNA containing) to noncored particles in the
progenyfromwild-typevirus-infected cells and in the progeny from mutant 69-infected cells.
Therewas aslightlylargerproportion of
nonen-veloped particlesin the progenyofmutant 69
infections comparedwiththose of wild-type
in-fections, butthis was notparticularly striking
and was insufficient to explain the defective
on November 10, 2019 by guest
http://jvi.asm.org/
MUTANTS
100
90
80
70
v 60
R0
X 50
e 40 30
201
10
1 9 17 25 1 9 17 25 1 9 17 253341 HOURS POST-INFECTION
FIG. 3. Pattern ofvirus DNA synthesis in HEF
cellsinfected with HSV-2-333,mutant69,ormutant 74andmaintainedat33or39C. Columnsrepresent total [3H]TdRincorporation in theviruspeak (from
CsCl equilibrium density gradient) during 8-h
pulses of labeled material.
TABLz 2. Productionof infectious and noninfectious
progenyby HSV-2 strain333andmutants69and74
inhamstercellsat33and 39Ca
Vi- Growth Particle/in- Nonenve- Nocore/ rus temp(C) fectivityra- loped/en- core
tio veloped
333 33 23-50 10-27 2-10
39 50-100 16-30 8-20
74 33 27-70 88-200 30-70
39 1.5-9x10' 100-300 50-100 69 33 2.3-10x10' 64-100 5-10
39
102-10'
40-200 10-50 6 Virus prepared under different experimentalcondi-tionswastitrated in RK cellsat33 C.
(i.e., morphologically normal but
noninfec-tious) nature ofthe greatmajority ofprogeny
produced bymutant69.
Virusproducedin HEFcellsat39Cshoweda
similarpattern.Therewas aslightincrease in
the proportion ofnonenveloped particles com-pared with the progeny of wild-type virus grown at 39C andno significantdifference in the ratio ofcoredtononcored particles. Thus,
there isno obvious morphological explanation
forthe defectivenatureof the bulk of the
prog-enyvirus produced by mutant69.
Progeny produced by mutant 74 at 33 C
ap-pearedtohave onlya veryslightlygreater ratio
ofparticle to infectivity than wild-type virus
progeny, and both the proportion of
nonenvel-oped and the proportion ofemptyparticleswere
somewhatgreaterthaninpreparationsof
wild-type virus. At39 Ctheratio of particle to infec-tivity ofthe progeny was significantly higher
than for wild-type virus, suggesting that
mu-tant 74, like mutant 69, produces "defective" progeny at 39C. Many of the noninfectious progenyofmutant 74appearedto be
nonenvel-oped, and therewas an increase inthe number
of"empty" nucleocapsids producedatthis
tem-perature.
The propensityofmutant 69forproducinga
large number of defective particles was also
demonstratedinpermissivecells. Virus stocks
grown inHELcellsat 33C hadahigherratioof
particletoinfectivity inthecase ofmutant69
thanfor wild-type or mutant 74virus. The
de-gree of defectiveness of virus in stocks of 69
dependeduponthehost cell in which thevirus
wastitrated (Table3).The observations bothat
33 and 39C further support the "host range"
propertiesof thetwo mutants.
Wild-typevirushadessentially thesame
ra-tioofparticletoinfectivity (approximately10to
25particles/PFU) when titratedinRK, HEL,or
BSC-1 cells. InHEFcellsaslightly higherratio
ofparticle toinfectivitywasobserved at39C,
reflectinganapproximately 10-fold decreasein
plating efficiencyof HSV-2atelevated temper-aturesinthese cells. BSC-1 cellsappearedtobe permissivefor all three virusestested, atboth temperatures, and the lowest ratios of particle to infectivity were observed for all viruses in
these cells. RKcells, althoughmorepermissive
than HEF cells formutantvirusgrowth,
partic-ularlyat39C,wereless permissive than HEL
orBSC-1 cells. 3T34E TK- cells supported
lit-tle wild-typeor mutant virusgrowthat39C.
Assay for defective interfering particles.
Since stocks ofmutant 74, and more
particu-larly mutant69, were knowntocontainmany
virusparticles thatwereunabletoreplicateat
39C, a phenomenon that was especially
marked in HEF cells, experiments were
de-signed to determine whether the presence of
mutantviruswould interfere withreplication ofwild-type HSV-2-333 at 39C in HEF cells.
Defectiveinterferingparticleshavebeen
previ-ouslyreportedinHSV stocksprepared byvirus passage at high input
multiplicities
(4) andalso between wild-type MPdk+ and
MPdk-strainsofHSVgrowingin canine cells(33).
EJ -330C
_ -390C
333
69
74
h=
L-
I=1P
no
W_40~_-AZZZ-UM2W
VOL. 97
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.509.53.251.53.327.2]98 WESTMORELAND AND RAPP
TABLE 3. Relative plating efficiency of wild-type andmutantHSV-2 indifferent typesofcellsexpressedas
ratios ofparticle toinfectivitya
Platingefficiency of:
Cells 333wild type ts mutant 69 ts mutant74
33C 39C 33C 39C 33C 39C
HEF 17-25 250-480 200-300 7-9 x 107 40-110 9 x 105
Baby RK 10-18 15-30 100-200 1,500-1,900 20-50 60-100
HEL 17-25 17-25 80-100 750-900 20-50 50-80
BSC-1 cells 9-12 10-18 20-30 100-150 4-10 10-15
3T3-4E TK- cells 30-40 >104 5,000-8,000 >106 90-110 >104
aVirus infectivity was titrated using a plaque assay in which monolayers of cells weremaintained in
medium containing 0.5% methylcellulose for 4 days prior to the counting of virus plaques. Particle counting ofthe stock virus used in each experiment (grown in HEL cells at 33 C) was by electron microscope examination, as described in the text.
Such interference could be demonstrated
(Ta-ble 4). Wild-type and both mutantviruses
se-verely depressed the yield of HSV-2-333 when
the "interfering" virus was added at aninput
multiplicityof 20 PFU/cell. Decreased
concen-tration of interfering virus decreased the reduc-tion in progeny synthesis, but this effect was
least marked whenmutant69wasthe
interfer-ing agent.Itseemsentirelyreasonableto sup-pose that the large number of defective
parti-cles inpreparationsof this viruswas
responsi-ble for interference with normal replication of
HSV-2-333 at39C inHEF. Similar results to
those shown in Table 4were obtainedusing a
constantinput multiplicityof0.2PFU of HSV-2-333 percell and similar multiplicitiesof
"in-terferingvirus."
Interference by mutant defective particles
was also demonstrated using a plaque
reduc-tion technique (Table 5). At 33 C both large
(presumablywildtype) and small(presumably
mutant) plaques were counted, and the total
number of plaques was expressed. At 39C, it
was again observed that mutants 69 and 74
interfered with the development of wild-type
plaques, and again this effectwasmoremarked
forpreparationsofmutant69. Interference
be-tweenviruseswasnotapparentat33C, possi-bly becauseatthistemperatureconditions are
muchmorefavorable for virus growth, which is
hencefavoredoverabortive infection and
inter-ference.
Biochemicaltransformation of3T3-4E TK-cellsbyHSV-2 strain 333 andmutants69and 74.The effect of UV irradiationonthe
infectiv-ityofmutantand wild-type HSV-2 is shown in
Fig. 4. Wild-type strain 333 was found to be
more resistant to UV irradiation than either mutant, with 74 the most sensitive, the same
amount of irradiation causing a 1 to 2 log1o
greaterdrop in titer ofmutant74than of
paren-tal 333.
TABLE 4. Assay for defective interfering particles in stocksofmutantvirus,using virusyield as indicator Input multiplicity of interfering virus "Interfering virus" (PFU/cell)
20 2 0.2
333(Wild type)
Challenge: 2.0 2.0x 105 6.0 x 105 1.0x 106 PFU/cella (20)b (60) (100)
Challenge: 0.2 2.0 x 105 3.0 x 105 1.0x 106
PFU/cellc (20) (30) (100)
Mutant 69
Challenge: 2.0 4.0x 104 2.0x 105 3.0 x105
PFU/cella (4) (20) (30)
Challenge: 0.2 3.4 x 104 5.0x 105 8.0x106
PFU/cellc (3.4) (50) (80) Mutant 74
Challenge: 2.0 1.6 x 106 9.0x 105 1.0x 106
PFU/cella (16) (90) (100)
Challenge: 0.2 1.4 x 105 7.0 x 105 2.0x 106
PFU/cellc (14) (70) (200)
aAll cultures werechallengedwithHSV-2-333wild-type
virus at aninputmultiplicityof 2.0PFU/cell.Control yield =1.0x106fPFU/ml (20PFU/cell). Titers represent the yield of HSV-2(wild-type large plaques). Virus was titrated in RKcells asdescribedinthetext.
bNumbers in parentheses are percentages of control
yield.
eAll cultureswerechallengedwithHSV-2-333wild-type
virus at aninputmultiplicityof0.2PFU/cell.Controlyield = 106PFU/ml(20PFU/cell).Other conditions were similar tothose describedinfootnotea.
3T3-4E TK-cells were exposedto virusthat
had been irradiated for0 to 10 minand, after21
days of incubation in selective HAT medium,
the number ofcolonies that had formedonthe
12 replicate plates for each virus sample was
counted.Ineachexperiment, the reversion
fre-quencyof3T3-4ETK-cellsto a TK+phenotype
was estimated from the number of colonies
formingon12replicate plates containing cells
that had been exposed to UV-irradiated Tris
buffer. Anunfortunate feature of this
transfor-mationassayisthat the reversionfrequency of 3T3-4E TK- cellswas found tobe rather high J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.509.263.455.258.423.2]TABLE 5. Assay for defectiveinterfering particles in mutant stocks using plaque reduction as indicator
Temp No. of
plaques
(C) 10-2 10-3 10-4 10-5 10-6 10-7 333 33 TNTC TNTC TNTC 162 113 96 39 TNTC TNTC 174 164 119 98 69 33 TNTC TNTC TNTC 136 118 112
39 26 40 49 57 85 117
74 33 TNTC TNTC TNTC 142 137 116
39 55 57 61 111 115 125
Aconstant amount of wild-type 333 virus was plated which produced an expected number of plaques=90to 100/ dish at both 33 and 39 C. TNTC, Too numerous to count.
0 us
z
LL
0
(A 0o cr:
0 2 4 6 8 10
MINUTES ULTRA-VIOLET IRRADIATION
FIG. 4. Survival of HSV-2 strain 333 and
mu-tants 69 and 74after exposure to increasing doses of UVlight.Symbols: 0,HSV-2 strain 333(wildtype); 0,mutant69; 0,mutant74.
andvariable; nevertheless, theresultsreported
hereareregularlyreproducible. Reversion
fre-quency wascalculated for each experiment and
subtracted from frequency of foci on
virus-treated plates to give the "transformation
fre-quency."
Unirradiated virus-induced cytopathology
and nocolonieswere seenafterexposureof cells
to unirradiated wild-type virus or mutant 69.
Cells infectedwithunirradiatedmutant 74did
giverise to rarecolonies,buttheseoccurredat
amuchlowerfrequencythan thereversion
fre-quency of 3T34E TK- cells (approximately
1-5) and theirsignificance is obscure.
WithincreasingUVdose,astheinfectivityof
virus preparations decreased, the frequency
withwhich transformedcolonieswereobserved
increased (Fig. 5). The data in Fig. 5have all
been corrected for the mean number of foci
16 14 12 10 8 6 4 w_ 2
-J
0
cc
12
3 0
U 10
U.
0
c 8
z
c4
'u
2 0
10 8 6 4
2 0
0 2 4 6 8 10
MINUTES U/V IRRADIATION
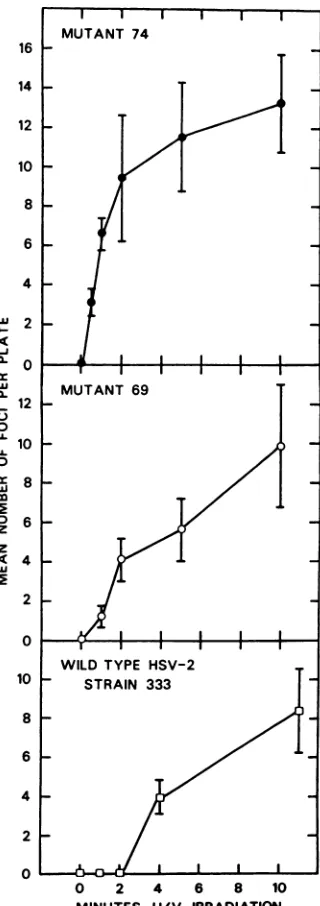
FIG. 5. Biochemical transformation of 3T34E
TK- by HSV-2-333 and mutants 69 and74. Mean numberof foci per plate was corrected for the mean numberof focioncontrol (non-virustreated) plates. The results represent themeanofthreeexperiments, andthe barsgive the standarderrorabout themean.
99
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.509.54.253.219.491.2]100 WESTMORELAND AND RAPP
observed on control plates. This varied from
0.17to 2.5 indifferent experiments. Significant
transformation was not observed with
wild-type HSV-2-333 until the preparation had
re-ceived at least 2 min of UV irradiation(4.8 x
105ergs/cm2); thereafter, the observed number
oftransformed foci increased. The maximum
frequency oftransformation events (i.e.,
num-bers of foci corrected for meanfoci oncontrol
plates) isapproximately3 x 1O-4.
Mutant 69, which was found tobe more
sensi-tive to UVirradiationthanthe parentalvirus,
induced the formation of significant numbers of transformed foci at lower UV doses than
wild-type 333, and aslightly higher maximum
fre-quency oftransformationwasobserved.
A more striking difference in transforming
ability was shown by mutant 74. This virus, which was the most sensitive to UV
irradia-tion, wasable to transform significant numbers
of cells at alow (12 x 104ergs/cm2) UV dose,
and thefrequencyof transformation after
expo-suretothis virusincreasedrapidlytoaplateau
levelthat was equal to, orslightlyhigherthan,
the maximum transformation frequency
ob-served withwild-typevirusormutant 69.This
plateauwasmaintained for UV dosesfrom2to
10min(4.8 x 105 to2.4 x 106 ergs/cm2). Virus
infectivity droppedtoundetectablelevels after
only5 x 105ergsof irradiation per cm2.
The data presented in Fig. 5 represent the
mean ofthree experiments. Morerecently, two
further experiments, using virus that had been
irradiated for as much as 15min,have provided
similar results.
Fociobserved intransformationexperiments
consisted ofhomogeneous populationsofcells,
but atleast two different morphological types
wereseenindifferent foci. One type was aflat,
epithelioidcell that gave rise to large, appar-ently single-layered colonies. The second cell
type wasdistinctly fibroblastoid andgave rise
to multilayered foci. These observations are
similartothose reportedinanearlier study of
HSVtransformation of mouse 3T3 cells (13), in
which theHATselection systemhad notbeen employed.
DISCUSSION
In the present study, two mutants,
previ-ously reported to bedefective for replication in
HEF cells at 39 C, were compared with each
other andwiththeparental virus for a number
of biological properties. These mutants were
chosen becausethey were found to be members
oftwodistinctcomplementationgroups (23) and
were not as leaky as other
temperature-sensi-tivehostrange mutants isolated (23).
Preliminary
characterization of themutantshaddemonstrated that their temperature
sensi-tivity was notduetothermolability ofthe vi-rion(R.W. Koment, unpublished data)ortoa
block in virusadsorptionorpenetrationat39 C
(23). Thiswasconsistent with resultsreported in this communication, demonstrating that a
number ofvirus-directed changes occurred in
HEF cells infected at 39C. Some virus DNA
synthesistook place, host cell DNA
synthesis
was turned off(datanotpresented), andafew
virus particles wereproduced, although these
particlesweremainly noninfectious. The
obser-vationthatmutant69producesmany
morpho-logically normal particles of greatly reduced infectivityis similartothat madebySchafferet al. (37), who reported a C groupts mutantof
HSV-1 that produced apparently
morphologi-cally
normalparticlesofgreatlyreducedinfec-tivity.
Schaffer and co-workers (37) were also ableto
correlate the production of cored and empty
particleswith the DNAphenotype of the virus.
To some extent, similar observations were
made in thisstudy usingmutant 74.This
mu-tant, even atpermissive temperatures in HEF
cells,hadadelayedonsetofDNAsynthesisand
at39 C made very little virus DNA. Particles
produced in cells infected by mutant 74 showed
a preponderance of nonenveloped empty
nu-cleocapsidsat39C.
The "host range" properties of mutants 69
and 74 appearto form a gradualpattern from
fully permissive cells (BSC-1) to extremely
re-strictive cells (HEF). This growth restriction in
different cells isdemonstrated at33 C but, at
39C, it is much more marked. Itseems
reason-able to conclude that the mutants have
diffi-culty in replicating in certain cells and that the
additionalstressofhigh temperature (HSV-2 is
knowntogrowpoorlyatelevated temperatures
[6]) prevents virus growth almost completely.
This conclusion could have importantparallels
invivo, particularly with regardtolatency. It
may be that certain cells ofthe host are
natu-rally resistant to HSV growth or even that
there are certain naturally occurring HSV
strainswhose growth isrestricted in some cells,
e.g., of nervoustissue. Such strain differences
would not be without precedent; Epstein-Barr
virus for example, another herpesvirus of
hu-mans,appearstohave at least twoforms, one of
which canreplicate inlymphocytes,the other of
which cannotreplicate but can transform
hu-mancord bloodlymphocytes (27, 28). It is even
possible, of course, that such strain differences
ofEpstein-Barr virus are due to detectivess,"
likemutants 69 and 74.
The high transforming frequency observed J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
withmutant 74 after relatively low doses of UV
irradiation was a particularly interesting
re-sult in this context. It had been thought possi-ble that mutant 69, which produced a very large number of defective particles, might
prove to be aparticularly efficient transforming
virus. This was not found to be the case, as
extensive cytopathic effects were observed
when this virus was added to cells after low UV doses, suggesting that, although unable to
rep-licate, the majority of mutant 69 particles are
able to induce cell death and, thus, preclude transformation. Mutant 74, although it did not produce a large number of defective particles,
was abnormally sensitive to UV damage. It
seems probable that this property, together
withthe naturally delayed growth cycle of the
virus, tipped the balance toward cell
transfor-mation (rather than lysis). It should be
men-tioned again,however, that there are a number
of shortcomings with the transformation
sys-tem used in this study. The assay was based
strictly on quantitative focus formation, and colonies were not examined for the presence of herpes-specific antigens. Considerable
varia-tion was seen in the number of fociproduced in
different experiments, and the reversion
fre-quency of the 3T3-4E TK- cells was high (1 x
105to 5 x 10-5). Similartransformation
experi-ments using L(TK-) cells (obtained from W.
Munyon) gave transformation frequencies too
low tobe quantified, but again a greater
trans-formingefficiency was observed using mutant
74 than mutant 69 or wild-type 333 virus.
Takentogether, the data on the transforming
potentialof the three virusestestedlead to the
conclusionthat great care must be exercised in
the development ofattenuated herpes simplex
virus vaccines. Mutant 74 is attenuated, both
forgrowth incell culture and for virulence in
mice(22),and on this criterion would be
consid-eredasuitablevaccinecandidate. However,by
selecting forgrowth attenuation, selection has
alsobeen made foraviruswhosecytolytic
prop-ertiesaremorethanusually sensitive to
exoge-nousdamage, but whose transforming
capabili-tiesremain intact. This situation would clearly
be most undesirable inavirusvaccine.
ACKNOWLEDGMENTS
Thiswork was supported by contract no. NOICP53516 within TheVirusCancerProgram of the National Cancer Institute and fellowship IARC/R 622 awarded to Diana Westmoreland fromthe WorldHealthOrganization, Inter-national Agency forResearch on Cancer.
Wethank Ronald Glaser andRoss Farrugia for the elec-tronmicroscopy and itsevaluation and Jenny Kilmer for excellent technical assistance.
LITERATURE CITED
1. Aron,G.M., D. M.Purifoy,and P. A.Schaffer. 1975. DNA synthesis and DNA polymerase activity of
herpes simplex virus type 1 temperature-sensitive
mutants. J.Virol. 16:498-507.
2. Ben-Porat, T., and A. S. Kaplan. 1973. Replication-biochemical aspects, p. 163. In A. S. Kaplan (ed.), The herpesviruses. AcademicPressInc., New York. 3. Benyesh-Melnick,M., P.A.Schaffer,R.J.Courtney,
J. Esparza, andS. Kimura. 1975. Viralgene
func-tionsexpressedanddetected by temperature sensi-tivemutants ofherpes simplexvirus. Cold Spring
Harbor Symp. Quant. Biol.39.731-746.
4. Bronson, D. L., G. R.Dreesman, N.Biswal, and M. Benyesh-Melnick. 1973. Defectivevirionsof herpes simplexviruses.Intervirology1:141-153.
5. Brown,S.M., D. A. Ritchie, and J.H.Subak-Sharpe.
1973. Genetic studies ofHSV-1. Isolation ofts mu-tants, their arrangement into complementation
groupsand recombinationanalysis leadingto a
link-age map. J.Gen. Virol. 18:329-346.
6. Crouch,N.A., and F.Rapp. 1972.Cell-dependent
dif-ferencesintheproduction of infectious herpes sim-plex virus at asupraoptimaltemperature. J. Virol.
9:223-230.
7. Davis, D. B., W. Munyon, R. Buchsbaum, and R.
Chawda. 1974. Virustype-specific thymidinekinase incellsbiochemically transformedby herpessimplex virustypes 1and2.J. Virol.13:140-145.
8. Docherty, J. J.,andM.Chopan.1974.The latentherpes simplexvirus.Bacteriol.Rev. 38:337-355.
9. Duff,R., andF.Rapp.1970.Quantitative
characteris-tics of the transformation of hamster cells by
PARA(defectivesimian virus40)-adenovirus7.J.
Vi-rol.5:568-577.
10. Duff, R., andF.Rapp.1971.Oncogenic transformation ofhamstercells after exposure to herpes simplex virustype 2. Nature(London)NewBiol.233:48-50.
11. Duff, R., and F. Rapp. 1971. Properties of hamster
embryo fibroblasts transformedinvitro after expo-sure to ultraviolet-irradiated herpes simplex virus type 2. J.Virol.8:469-477.
12. Duff, R., andF.Rapp.1973.Oncogenic transformation ofhamsterembryo cellsafter exposuretoinactivated herpessimplexvirustype1.J.Virol.12:209-217. 13. Duff, R., and F. Rapp. 1975. Quantitative assay for
transformation of3T3cellsbyherpessimplexvirus type 2. J.Virol.15:490-496.
14. Esparza,J.D.,J. M.Purifoy,P. A.Schaffer, and M. Benyesh-Melnick. 1974.Isolation, complementation
andpreliminaryphenotypiccharacterizationof
tem-perature-sensitive mutantsofherpessimplexvirus type 2.Virology 57:554-565.
15. Halliburton,I.W.,andM.C.Timbury. 1973. Charac-terization of temperature-sensitivemutantsof herpes
simplex virus type 2. Growth and DNAsynthesis. Virology54:60-68.
16. Honess,R. W.,and B.Roizman. 1973. Proteins
speci-fiedby herpes simplex virus. XI. Identification and relative molar rates ofsynthesis ofstructural and nonstructural herpesvirus polypeptides in the
in-fectedcell. J.Virol.12:1347-1365.
17. Honess, R. W., and B.Roizman. 1974.Regulation of
macromolecular synthesis of three groups of viral proteins. J. Virol.14:8-19.
18. Honess, R. W., andB. Roizman. 1975. Regulation of herpesvirus macromolecular synthesis: sequential
transition ofpolypeptide synthesis requires func-tional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A.72:1276-1280.
19. Kit,S.,and D. R.Dubbs. 1963. Acquisitionof
thymi-dine kinaseactivitybyherpessimplexinfectedmouse fibroblast cells. Biochem. Biophys. Res. Commun.
11:55-59.
20. Kit, S., W. C. Leung, G. N.Jorgensen, D.Trkula,and D. R. Dubbs. 1975. Thymidinekinase isozymes of normaland virus infected cells. ColdSpring Harbor
101
on November 10, 2019 by guest
http://jvi.asm.org/
102 WESTMORELAND AND RAPP
Symp. Quant. Biol.39:703-715.
21. Klemperer, H. G., G. R.Haynes, W. H. Shedden,and D. H. Watson. 1967. A virus specificthymidine ki-nase inBHK-21 cells infected with herpes simplex virus.Virology 31:120-128.
22. Koment, R. W., and F. Rapp. 1975. In vivo characteris-tics oftemperature-sensitivehostrange mutants of herpes simplex virus type 2.Intervirology5:10-20. 23. Koment,R. W., and F. Rapp.1975.
Temperature-sensi-tive hostrange mutants of herpes simplex virus type 2.J. Virol.15:812-819.
24. Koment, R. W., and F. Rapp. 1975. Variation in
suscep-tibilityofdifferentcell types totemperature-sensitive
host range mutants of herpes simplex virustype 2. Virology 64:164-169.
25. Littlefield, J. 1965. Studies on thymidine kinase in cultured mouse fibroblasts. Biochim. Biophys. Acta 95:14-22.
26. Macnab, J. C. M. 1974. Transformation of rat embryo
cellsbytemperature sensitive mutants of herpes
sim-plexvirus. J. Gen.Virol. 24:143-153.
27. Miller, G., J.Robinson, and L. Heston. 1975. Immortal-izing andnon-immortalizingstrains ofEpstein-Barr virus. Cold Spring Harbor Symp. Quant. Biol.
39:773-781.
28. Miller, G., J. Robinson, L. Heston, and M. Lipman. 1974. Differencesbetween laboratory strains of
Ep-stein-Barrvirus based onimmortalization,abortive infection and interference. Proc. Natl. Acad. Sci.
U.S.A. 71:4006-4010.
29. Monroe, J. H., and P. M. Brandt. 1970. Rapid semi-quantitative method for screening large numbers of virus samples bynegative staining electron micros-copy.Appl.Microbiol.20:259-262.
30. Munyon, W., E.Kraiselburd,D. Davies, and J. Mann. 1971.Transferofthymidine kinase to thymidine ki-naseless L cells by infection with ultraviolet-irradi-ated herpessimplex virus. J. Virol. 7:813-820. 31. Rapp,F. 1963. Variants ofherpes simplex virus:
isola-tion,characterization, and factors influencing plaque
formation.J.Bacteriol.86:985-991.
32. Rapp, F., and M. A.Jerkofsky. 1973. Persistent and latentinfections, p. 271-289.InA. S. Kaplan (ed.), Theherpesviruses.AcademicPressInc.,New York. 33. Roizman, B. 1965. Abortiveinfectionof canine cellsby herpes simplexvirus. HI.The interference of
condi-tionallethalviruswithanextended hostrange mu-tant.Virology27:113-117.
34. Roizman, B., andP. R. Roane.1964. The multiplica-tion of herpes simplex virus. II. The relationship
between proteinsynthesisand theduplicationof viral DNA ininfectedHEp-2 cells.Virology22:262-269. 35. Roizman, B., P. G. Spear, and E. D. Kieff. 1973.
Herpessimplexviruses1and2:abiochemical defini-tion. Perspect.Virol.8:129-169.
36. Schaffer,P.A., G. M.Aron,N. Biswal,andM.
Ben-yesh-Melnick. 1973. Temperature-sensitivemutants
ofherpessimplexvirustype1:isolation, complemen-tionandpartial characterization. Virology52:57-71. 37. Schaffer, P.A.,J. P.Brunschwig, R. M. McCombs,
andM.Benyesh-Melnick.1974. Electronmicroscope studies oftemperature sensitive mutants ofherpes simplexvirustype1.Virology62:442-457.
38. Stevens, J. G., and M. L. Coor. 1973. Latentherpes
simplex in sensoryganglia. Perspect. Virol. 8:171-188.
39. Subak-Sharpe, J. H., S.M.Brown,D.A.Ritchie,M.C. Timbury,J. C. M.Macnab, H.S. Marsden, and J. Hay. 1975. Geneticandbiochemicalstudies with her-pesvirus. Cold Spring Harbor Symp. Quant. Biol. 39.717-730.
40. Takahashi, M.,and K. Yaminishi. 1974.
Transforma-tionof hamsterembryo andhumanembryocellsby temperature sensitive mutants ofherpessimplex vi-rus type 2.Virology61:306-311.
41. Wilkie,N.M., J.B.Clements,J. C.M.Macnab,and J.
H.Subak-Sharpe.1975.The structure andbiological
properties ofherpessimplexvirusDNA.ColdSpring
HarborSymp. Quant. Biol.39:657-666.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/