JOURNAL OF VIROLOGY,Sept. 1980,p.722-731 0022-538X/80/09-722/10$02.00/0
Vol.35,No.3
Correlation of RNA
Binding Affinity
of
Avian
Oncornavirus
p19
Proteins with the Extent of
Processing
of Virus
Genome
RNA in
Cells
J. P. LEIS,t* P. SCHEIBLE,ANDR. E.SMITH
DepartmentofMicrobiology and Immunology, Duke University Medical Center, Durham, NorthCarolina 27710
We
purified
thep19proteins
from thePrague C strain of Roussarcomavirus,avian myeloblastosis virus, B77 sarcoma virus, myeloblastosis-associated virus-2(0), and PR-E95-C virus and measured their binding affinities for 60S viral RNA by thenitrocellulose filterbinding technique. Theapparentassociationconstants of the p19
proteins
from Rous sarcoma virusPrague
C,
avianmyeloblastosis
virus,andB77 sarcomavirus forhomologous and
heterologous
60S RNAswere similar (1.5 x1011
to 2.6 x1011 liters/mol),
whereas those ofmyeloblastosis-associated
virus-2(0)
and PR-E 95-C virus were 10-fold lower. The sizes and relativeamountsof thevirus-specific polyadenylicacid-containing
RNAs in thecytoplasms
ofcellsinfected with RoussarcomavirusPragueC,
myeloblastosis-associated
virus-2(0),
and PR-E95-C virusweredeterminedby
fractionating
the RNAsonagarosegels containing methylmercury hydroxide, transferring
themtodiazobenzyloxymethyl
paper and hybridizing them to a 70-nucleotidecomple-mentary DNA
probe.
In cells infected with Rous sarcoma virusPrague C we detected 3.4 x 106-, 1.9 X106_,
and1.1 x106-dalton RNAs,
in PR-E95-C virus-infectedcells wedetected3.4 x106_,
1.9 x106_
and0.7x106-dalton
RNAs, and in cells infected withmyeloblastosis-associated
virus-2(0) we detected 3 x106_
and1.3x
106-dalton
RNAs. Each of these RNAspecies
contained RNAsequencesderived from the 5'terminus of
genome-length
RNA,
asevidencedby hybridiza-tionwith the 5' 70-nucleotidecomplementary
DNA. Theratios ofsubgenomic mRNA'stogenome-length
RNAs incells infected withmyeloblastosis-associated virus-2(0) and PR-E 95-C viruswerethree- tofive-foldhigher
than the ratio in cells infected with RoussarcomavirusPrague
C. These resultssuggestthatmore processing of viral RNAininfectedcells iscorrelated with lower binding affinities of thep19protein
for viralRNA,
andthey
areconsistent with thehypothesis
thatthe
p19
protein
controlsprocessing
of viral RNA in cells.Several
investigators
havepresented
evidence thatthereare atleast three viral mRNA's in thecytoplasm
of avian oncornavirus-infectedcells,
which have sedimentation coefficients of
38S,
28S, and 22S (3, 5, 10, 32).
Hybridizations
ofcomplementary
DNA(cDNA) probes
represent-ing
specific regions
of the viral genome haveshown that the 38S RNA represents the entire viral genome, the 28SRNA represents the
en-velope and sarcoma genes, and the 22S RNA represents the sarcoma gene. The first RNA serves as the mRNA for the
group-specific
an-tigens and the,B chain ofreverse
transcriptase
(7, 8, 14, 16, 17),thesecond RNA serves asthe mRNA for the
envelope
glycoprotein
(5, 18, 28, 30), and the third RNA mayserve asthe mRNA forthesarcomaprotein (2, 19, 20).
The28SandtPresent address: Department of Biochemistry, Case Western Reserve University, Cleveland, OH44106.
22S mRNA's contain at the 5' end an RNA sequence derived from the 5'
terminus
of 38S RNA (9, 15, 21, 32), suggesting that they ariseby
splicing
ofthe 38S RNA.The
enzymatic
mechanism for processingviralRNA incells is not known. However, we have observed thatpurified preparationsofRNase III cleave the 38S RNAof the PragueC strain of Rous sarcoma virus (RSV Pr-C)into fragments containing polyadenylic acid [poly(A)]; these fragments are similar in size to virus-specific RNAs isolated from infected cells. Moreover, one of the viral group-specific antigens, p19, bindsto
specific regions
of38SRNAand blocks cleavageof the RNAby purifiedRNase III (12). Theseresults suggestthatp19mayplaya rolein
controlling
in vivo processing bypreventingcleavage ofviral RNA by a RNase III-related splicingenzyme. Toprovide furthersupport for
722
on November 10, 2019 by guest
http://jvi.asm.org/
this hypothesis, we examinedtwo RNA tumor viruses which contain increased levels of the major envelopeglycoprotein in theirvirions (29; M. Linial,personal communication). ThePR-E 95-Cvirushas alteredmolecularweightforms of thep19 protein (24). In this paper we report that thep19proteins purified fromthese two viruses have a decreased affinity for 60S viral RNA. Furthermore, the levels of the subgenomic mRNA's found in the cytoplasm of cellsinfected with these viruses are greater than the levels found in theRSV Pr-C-infected cells.
MATERIALS AND METHODS
Reagents.
'Pi,
(carrier free), [3H]uridine (46.9Ci/ mmol), [3H]dCTP (25.6 Ci/mmol),[a-`2P]dCTP
(33 Ci/mmol),and [3H]dTTP (18.6Ci/mmol) were pur-chased from NewEnglandNuclear Corp. Nitrocellu-lose filters (type HA; 25 mm; pore size,0.45Im)
werepurchasedfromMilliporeCorp.,andglassfiber filters (typeE)werepurchasedfrom Gelman Instrument Co. Enzo-bond paperwaspurchasedfrom Enzo Biochem-ical Co. All other reagentswerepurchasedas previ-ously described (11).
Growth of viruses. Cloned RSV Pr-C and B77
sarcoma virus (subgroup C) were grown inchicken embryofibroblasts, asdescribedbySmith and
Bern-stein (25). The RNAsproduced by these virions were more than 97% a class RNA, as analyzed by gel
electrophoresis. Myeloblastosis-associated
virus-2(0)[MAV-2(0)],anontransforming virus of subgroup B,wasplaque purifiedand grown in chicken embryo fibroblastsasdescribedbySmithetal.(26). An avian
sarcomavirus(PR-E95-Cvirus)derivedbymultiple recombinationevents betweenRSV Pr-C and Rous-associated virus-0was agiftfrom MaxineLinial,Fred Hutchinson Cancer Research Center, Seattle,Wash. PR-E95-C virus isatransformingvirusofsubgroupE andwasgrowninquail embryofibroblasts. The virus titer in each casewas 106 infectious units per ml in
chronicallyinfected cells.
Purificationof viralproteins.RSV Pr-Cp19and
p12,B77sarcomavirusp19,avianmyeloblastosisvirus (AMV) p19, and Escherichia coli RNase III were
purifiedaspreviouslydescribed(12).PR-E95-C virus
p19andp12andMAV-2(0)p19andp12werepurified
bythemethod of Green andBolognesi(4). The PR-E 95-C virusproducestwoforms ofp19,asidentifiedby sodiumdodecylsulfategel electrophoresis (24). Both formsmigratefaster than RSV Pr-Cor Rous-associ-ated virus-0p19onthesegels. Thetwop19proteins
wereseparatedfromoneanotherby chromatography
onphosphocellulose p11 (column volume,2ml)
equil-ibrated with a solution containing20 mM morpho-lineethanesulfonic acid buffer (pH 6.6), 0.2% Triton X-100, and 0.1 mMEDTA (buffer A). Thep19 proteins
were eluted from the column with a 16-ml linear gradient of0 to 0.5 M NaCl in buffer A and were detectedbyretention of 3P-labeled60SB77 RNA to nitrocellulose filters,asdescribedbelow. Two peaks of RNA-binding activitywereeluted from the column at
0.17and0.27MNaCl. Thep19protein eluting at 0.17
M NaCl contained70% of the totalprotein andwas used in theexperiments described below. These pro-tein fractionswereindistinguishable bysodium
dode-cyl sulfate gel electrophoresis at pH 8.8; they both yielded asingle polypeptide band.Also, thebinding
affinity of eachseparatedproteinfor60S PR-E 95-C virusRNAwasfoundtobe 1.8x 10"' liters/mol.
Preparation of labeled RNAs.
[3H]uridine-la-beled PR-E 95-C virus60S RNA (140 cpm/pmol of
nucleotide) and;2P-labeledMAV-2(0) 60S RNA(454 cpm/pmolofnucleotide)wereprepared bythe method ofSmithetal.(27).32P-labeled60S B77 RNA(1,296 cpm/pmol ofnucleotide) was a gift from A. Faras, UniversityofMinnesota MedicalSchool.
Preparation of virus-specific cDNA probes.
DNAcomplementary toviral RNAwas preparedas
describedbyLeisetal.(13),using60S RSV Pr-C RNA as the template and oligodeoxythymidylic acid1214 [oligo(dT)12-,4]astheprimer.Incubationwasfor16h
at 37°C. The reactions were stopped by adding a
twofoldexcessofEDTAtomagnesium ions, and the mixtureswere made0.1% in sodium dodecyl sulfate,
extracted withphenol,anddialyzedovernightat4°C against500 ml of2x SSC (lx SSC is0.15M NaCl
plus0.015Msodiumcitrate).The solutionwasbrought
to 0.3 M NaOH and incubated for 3 h at 30°C to hydrolyze RNA. The alkaliwasneutralized with
hy-drochloricacid,and the DNAwasdialyzedagainst500
ml of0.3MNaCl-20mMTris-hydrochloride(pH 7.5) overnightat4°C.These DNApreparationsweremore than 95%singlestranded,asdeterminedbydigestion
bynuclease S1. The32P-labeled70-nucleotide cDNA
probe(cDNA7o)waspreparedasdescribedbyLeiset al.(13),usingRSV Pr-C 38S RNA and
tRNAt,,
asthetemplate andprimer,respectively.After alkaline hy-drolysistoremoveRNA,thecDNAwasfractionated
byelectrophoresison 1.6%low-melting-pointagarose
gels calibrated withHaeIII-digestedsimian virus40
DNA.Bands of DNAwerelocatedbyautoradiography
of thewetgel,excised, and dissolved at65°Cin the buffer used fortwo-phasehybridization(31).
Preparation of cytoplasmic
poly(A)-contain-ingRNA.Poly(A)-containing RNA was isolated from RSV Pr-C-orMAV-2(0)-infected chicken embryo
fi-broblasts,uninfected chicken embryo fibroblasts, and PR-E95-Cvirus-infected quail embryo fibroblasts
es-sentially as described by Krzyzek et al. (9).
Virus-infected primarycellswereharvested from three roller bottles containing 8 x 108 cells and suspended in a solution containing 0.01 M Tris-hydrochloride (pH
8.0), 0.01 M NaCl, 0.015 M MgCl2, and 0.01 M
N-ethylmaleimide. Cellswerehomogenized, and the de-gree of cell breakage was monitored with a
phase-contrast microscope. Nuclei and cell debris were re-movedby successive centrifugations in aSorvallSS34
rotor at700xgfor4min and 3,000xg for 4 min at
4°C.Thesupernatantfractionwasdeproteinized four
timeswithanequalvolumeof phenol at50°C, and the RNAwasprecipitated with2volumes ofethanol. The RNAwasdissolvedin 2to 3ml of a solution containing
0.4MNaCl,2mMEDTA,10mMTris-hydrochloride (pH 7.5), and 0.5% sodium dodecyl sulfate, heated to 80°C for2min,rapidly cooled, and chromatographed
ona 10-mloligo(dT)-cellulose column at room
on November 10, 2019 by guest
http://jvi.asm.org/
724 LEIS, SCHEIBLE, AND SMITH
perature, whichwasequilibrated with the above-de-scribed buffer. The poly(A)-containing RNA which eluted from the column with 0.5% sodium dodecyl
sulfate-10 mM Tris-hydrochloride (pH 7.5)-10 mM NaCl was detected byabsorbancy at 260 nm. This
poly(A)-containingRNAwasbroughtto afinalNaCl
concentration of0.4 M, heated to 80°C for 2 min, cooledrapidly, and again fractionated on oligo(dT)-cellulose,asdescribed above. Thepoly(A)-containing
RNArecovered from the second columnusually rep-resented between1and 2% of the totalinput absorb-anceat 260 nm.The RNAwasadjustedto0.1MNaCl andprecipitatedwith2volumes of ethanol.32P-labeled
38S RSV RNA addedtothecellsexogenouslycould be recovered intact after the above-described proce-dure.
Fractionation ofpoly(A)-containing RNA by electrophoresis. The poly(A)-containing RNAwas
fractionated by electrophoresis through 1% agarose horizontal slabgels (30by16.5by0.3cm)containing
5mM methylmercury hydroxide(1). Electrophoresis
was at 150V for12 to 14hat4°C.Theelectrophoresis
bufferwascirculatedduringtherun.UnlabeledHeLa cell rRNA's were included as molecular weight
markers and were located by excising the marker tracks andstaining with ethidium bromide (1,ug/ml)
in0.5M ammoniumacetate.
Transfer of RNAto DBM paper.
Poly(A)-con-taining RNAwastransferred from the agarosegelsto
diazobenzyloxymethyl(DBM) paper (Enzo-bond),as describedby Wahletal.(31).RNAwasfragmentedas described by Wahletal. (31)tofacilitate transferto
the DBM paper.
Two-phasehybridization.The[32P]DNAprobes werehybridized to RNAcovalently boundto DBM paperasdescribed byWahletal. (31).The cDNA70
probe, whichwasisolated byagarose gel
electropho-resis,wasused without furtherpurification bymelting
the agarosegelfraction into thehybridizationbuffer
at65°C. ThecDNA70 probewas80-fold in excessof the virus RNA sequences on the DBM paper. For
sequential hybridization, the first DNA probe was removed from the paperbywashingwith 99% form-amide-10 mMTris-hydrochloride (pH 7.5)for20min
at65°C.Single-phase hybridizationswerecarriedout aspreviouslydescribed(13).
Nitrocellulose filter assay formeasuring bind-ing of RNA to proteins. The nitrocellulose filter
binding assaywas aspreviously described (12). The apparent association constants were calculated by plottingthe ratio of theamountof bound RNAtothe
amount offree RNA against the amount of bound
RNA,using the method of Scatchard(22),asmodified by Hizi et al. (6). The association constants were calculated from the negative slopes of the lines in theseplots andareexpressedinliters per mole.
RESULTS
Twoof thegroup-specificantigensof RSVp12 and p19, bind viral RNA (23). However, they differin theirbinding specificity; p12binds non-selectivelytoallpolyanions, whereasp19binds
preferentially
to regions of RNAs that containRNase III-sensitive sites (12). Furthermore, RSV p19 is abletoprotect38SRSV RNA from cleavage by RNase III in vitro, whereas p12
cannot.Thespecific RNA-bindingpropertiesof p19 have led us to postulate that this protein
mayaffect theamountof processing of
genome-length (38S) RNA into the smaller mRNA's which code for the envelope glycoprotein and thesarcomaprotein ininfectedcells.To provide furthersupportfor this hypothesis, wepurified p19 proteins from three avian oncornaviruses, RSV Pr-C, B77sarcomavirus, andAMV,which have similar levels of envelope glycoprotein in
their virions, as well as two avian
oncornavi-ruses, MAV-2(0) and PR-E 95-C virus, which
have higher levels of envelope glycoprotein in
their virions relativetolevels of the
group-spe-cific antigens. We measured the binding affinity ofthesep19 proteins for 60SviralRNA by using the nitrocellulose filter binding assay (6). The retention of32P-labeled 60S B77 sarcoma virus RNAtonitrocellulosefilters byMAV-2(0) and
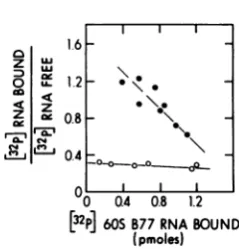
RSV Pr-C p19's isshown in Fig. 1. These data werereplotted by the method ofScatchard (22) (Fig. 2). The apparent association constant of RSV Pr-C p19 for60SB77RNA was 2.6x 1011
liters/mol, whereas the apparent association
constant of MAV-2(0) p19 was 10-fold lower (Table 1). By using the same technique, the apparent association constants for the p19's of B77sarcoma virus, AMV, and PR-E 95-C virus
werecalculated(Table 1). The binding affinities ofB77, AMV, and RSV Pr-C p19 proteins for 60S B77 RNA were very similar. Thus, there doesnotappear tobe anydetectable difference in the binding affinities of the p19 proteins of different virusesfor 60S B77RNA, as measured by this technique. These results are in
agree-a I I
, 20
clo
CLoll
__ III
0 [3 2 3 4 5 6
[32PJ60SB77 RNA ADDED
(Pmoles)
FIG. 1. Binding of 32P-labeled B77 RNA toRSV Pr-C and MAV-2(0)pl9proteins.Increasingamounts of32P-labeled60SRNA (1,272cpm/pmolof
nucleo-tide)wereincubatedwitheither2pgofRSVPr-Cp19 or0.8pgofMAV-2(0)p19, asdescribed in thetext. After15minat0°C, the reactionmixturewasdiluted
with I mlof10mM7ris-hydrochloride (pH 7.5)-10
mM NaCl andpassed throughanitrocellulosefilter. Symbols:0, binding of60S B77RNA toRSV Pr-C
pl9;0,bindingof60SB77 RNAtoMAV-2(0)p19. J. VIROL.
n,&I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.510.294.441.469.567.2]VOL. 35,
1.6
0
Z LU
fD0 U. 1.2
< z
Z = 0.8
,X,
I
0.4
_-0 0.4 0.8 1.2
[32p]
60S B77 RNA BOUND [image:4.510.96.216.61.186.2](pmoles)
FIG. 2. Scatchard plot of the data presented in Fig. 1. The amountoffree60S RNA was calculated bysubtracting theamountofbound 60S RNAfrom the total60SRNA. Symbols:0, binding of60SB77 RNAtoRSV Pr-Cp19; 0,binding of60S B77 RNA toMAV-2(0)p19.
TABLE 1. Association constantsof viral proteins for 60S B77 RNA
Protein Association
con-stant
(liters/mol)'
RSV Pr-C p19 2.6x10"
B77 sarcoma virusp19 2.0x101
AMVpl9 1.5xloI
PR-E95-C virusp19 1.9 x 1010
MAV-2(0) p19 2.5x10"'
RSV Pr-Cp12 1.2 x10"
PR-E 95-C virusp12 2.1 x1011
E.coli RNase III 4.5 x 1010
aThese valueswerecalculated from Scatchardplots
and represent theamountofbindingof 60S B77 RNA (1,272cpm/pmolofnucleotide)toRSV Pr-Cp19 (1.9
jg),
B77sarcomavirusp19
(1.1Mg),
AMVp19
(1.3Mg),
PR-E 95-Cvirusp19 (0.1,g),
MAV-2(0)p19 (0.18 tg),RSVPr-Cp12 (0.8,ug),PR-E95-C virusp12 (0.1,ug),
and E. coli RNase III(0.85U)asdescribed in thetext.
ment with results found previously with 60S RSV Pr-C RNA (12). Incontrast, theapparent association constants of PR-E 95-C virus and MAV-2(0) p19 proteins for 60S B77RNAwere similar to each other, but 10-fold lower than those oftheabove-described proteins (Table1). The PR-E 95-C virusprotein usedwasthe0.17 M eluantprotein from the phosphocellulose col-umn.
Binding of PR-E 95-C virus pl9to PR-E 95-C virus 60S RNA. To ruleoutthe
possibil-ity
that the lowerbinding
affinities of MAV-2(0) andPR-E 95-C virusp19
proteins for 60S B77 RNA were related to virus-specific nucleotide sequences, werepeatedthebindingexperiments with 60S PR-E 95-Cvirus RNAand 60S MAV-2(0) RNA. Figure 3 showstheretention of3H-labeled 60S PR-E95-C virusRNA to nitrocel-lulose filtersinthepresence of PR-E 95-C virus p19andRSV Pr-Cp19. Since the specific
activ-p19
0
z .
z
0 < 15
0 10
E 6"
5
&t0
_- 0 20 40 60 80
[image:4.510.301.421.69.178.2][3H]
60S 95-C RNA ADDED (pmol.s)FIG. 3. Binding of 3H-labeled PR-E 95-C virus 60S RNA toPR-E 95-C virus andRSVPr-Cpl9proteins. Increasing amounts of3H-labeledPR-E 95-C virus 60S RNA (140 cpm/pmol of nucleotide) were incu-bated with either 2 pgofRSV Pr-C p19or0.1pgof PR-E 95-C virusp19, as described in the legend to Fig. 1.Symbols:S,binding of60SRNA to RSV Pr-C p19;0, binding of 60S RNA to PR-E 95-C virus p19.
ity of the
3H-labeled
PR-E 95-C virusRNA was 1/10that of the 32P-labeled B77 RNA used in the experiment described above(Fig.
1), the amountof RNA bound withincreasingamounts ofinput RNA reachedaplateau with both pro-teins. TheScatchardplots of the datainFig.3 areshown inFig.
4, and thecalculatedapparent association constants aresummarized in Table2. The
binding
affinity
of PR-E95-C virus p19for
homologous
RNA wasagain
10-fold lowerthan the binding affinity of the heterologous RSV protein for PR-E 95-C virus 60S RNA. A similar
relationship
wasfound for thebinding
ofMAV-2(0)
p19 and RSV Pr-Cp19 for 60SMAV-2(0) RNA(Table2).These resultsindicate that the lower binding affinitiesof PR-E95-C virus p19 andMAV-2(0) p19 for RNAare
properties
ofthese proteins andnotof the RNA used.
Binding
of otherproteins
to 60S RNA.The p12 protein is the other
group-specific
an-tigen which bindstoRNA.
Therefore,
we mea-sured thebinding
affinities of the PR-E 95-C virus andMAV-2(0) p12 proteins for viral RNA todetermine whether their associationconstantswere also lower. We found that the apparent associationconstantof PR-E 95-C virusp12 for
60SB77RNAwas2x 1011
liters/mol,
thesameasthe
binding
affinity
ofRSV Pr-Cp12 for B77RNA
(Table
1). Thebinding
affinity
ofMAV-2(0)p12 for 60S MAV-2(0) RNAwassimilar (1.1
x
10"
liters/mol) (Table 2). The binding affinityoffelineleukemia virus plO for 60S RSV RNA was 1.3 x
1011
liters/mol (12). Thus,wedid not detectdifferencesin thebindingaffinities of the p12proteinsfromtheseviruses. Inaseparate set of experiments, the apparent associationcon-stantof E.coliRNase III for60SB77RNAwas
determinedtobe4.5 x
10l'
liters/mol.
0 o\o
0\
0\%
C I'm -C,
on November 10, 2019 by guest
http://jvi.asm.org/
726 LEIS, SCHEIBLE, AND SMITH
0
z
'0
o
z
Qc
1u
o9
.n 1.
LU.
ui
ccU.S
z
cm
u
-o
0.
0.
0.
0.
-0 2.5 5 75 10 12.5
[3H] 60S 95-C RNA BOUND
[image:5.510.66.254.59.225.2](pmoles)
FIG. 4. Scatchard plot of the data presented in
Fig.3. Symbols: 0, binding of 60S PR-E95-C virus
RNAtoRSVPr-Cpl9;0,binding of 60S PR-E95-C
[image:5.510.287.433.231.458.2]virus RNAtoPR-E 95-C virusp19.
TABLE 2. Associationconstantsofviralproteins for PR-E95-C virus andMAV-2(0)60SRNAs
Association Protein 60S RNA constant (ii-ters/mol)' PR-E 95-Cvirus PR-E 95-C virus 1.8x
10'"
p19
RSV Pr-Cp19 PR-E 95-C virus 1.1 x 101
MAV-2(0) p19
RSV Pr-Cp19 MAV-2(0)p12
MAV-2(0) MAV-2(0) MAV-2(0)
The sizesof the viral RNAswereestimated by
plotting relative migration distances against logs ofmolecular weights, using 28S and 18S HeLa cellrRNA'sasmarkers. As Leeetal. (10) found, there were three RNA species in RSV
Pr-C-infected cells; the molecular weights of these RNAswereof 3.4x 106, 1.9 x 106, and1.1 x 106 (data not shown). When preparations of RSV Pr-C contained transformation-defective virus,
twoadditional RNA species (molecularweights, 3.0 x 106and 1.3x 106) werefound. The RNAs
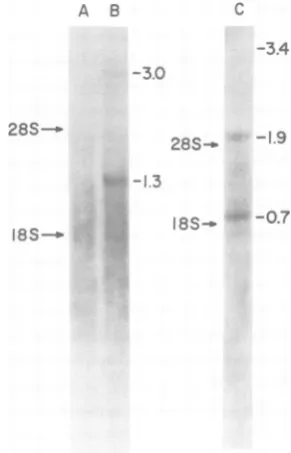
isolated from MAV-2(0)- and PR-E 95-C virus-infected cells are shown in Fig. 5.
MAV-2(0)-infectedcellscontainedtwoRNA species, which
A B C
-3.4 -3.0
28S-28S-b -1.9 1.3
18S--I1S
---0.7
4.1 x10'0
2.4x1011 1.1 x 1011
aThe valueswerecalculated from Scatchardplots
and represent the amount ofbinding of3H-labeled PR-E 95-C virus RNA (140 cpm/pmolofnucleotide) and3H-labeledMAV-2(0)60SRNA(454 cpm/pmolof
nucleotide) toPR-E 95-C virusp19 (0.1
jig),
RSV Pr-Cp19 (1.91g),
MAV-2(0) p19 (2.1,ug),andMAV-2(0) p12(0.3,ug),asdescribedin Table1,footnotea.Size ofvirus-specific RNAsisolated from the cytoplasm of infected cells. The data presented above show that the p19 proteins
pur-ified fromMAV-2(0) and PR-E 95-C virus have lowerbinding affinities for viral RNA than B77
sarcomavirus, RSV, and AMV p19 proteins. If
p19 suppresses the cleavage of the 38S RNA into subgenomic mRNA species in cells,the ratio
of smaller mRNA species to genome-length RNA should begreaterinMAV-2(0)- and PR-E 95-C virus-infected cells than in RSV Pr-C-in-fected cells. Weidentified the viral RNA species inRSVPr-C-, MAV-2(0)-, and PR-E 95-C virus-infected cells byfractionating the cytoplasmic
poly(A)-containing RNA on agarose gels
con-taining methylmercury hydroxide. After transfer
ontoDBMpaper,the virus-specific RNAswere
detected by hybridization to oligo(dT)-primed RSV Pr-C cDNA probes, as described above.
FIG. 5. Sizes of virus-specific poly(A)-containing
RNAs from the cytoplasm of infected cells. RNAs
wereisolatedfromthecytoplasm of uninfectedand MAV-2(0)-infected chicken embryo fibroblasts and PR-E 95-C virus-infected quail embryo fibroblasts, fractionated on 1% agarose gels containing 5 mM methylmercury hydroxide, andtransferred to DBM
paper asdescribedin thetext;10 nmolofeach RNA
wasappliedtotheagarosegels. Virus-specificRNA
wasdetected by incubating thepaperwith 2 x 106
cpmof oligo(dT) -primedRSVPr-C cDNAfor48hat 650C,washedasdescribedbyWahl etal.(31), dried,
and subjected to autoradiography with Kodak X-OMat X-rayfilm, using Lightning-Plus screens at -700Cfor 7days. The PR-E 95-C virus RNA was
fractionatedonaseparateagarose gel. Thearrows
indicated thepositions ofthe 28S and 18S HeLacell
rRNA's. LaneA, CytoplasmicRNAfrom uninfected
chickenembryo fibroblasts;laneB, cytoplasmicRNA from MAV-2(0)-infectedchickenembryo fibroblasts;
lane C, cytoplasmicRNAfromPR-E 95-C virus-in-fectedquail embryo fibroblasts.
I
.0- *'
-1.8_ 0
0
!4
\
00
12 O0 0 O\ _
O.. _~0--'
.2~~~~~~0-01
1=0
1-. 1J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.510.65.257.301.399.2]had molecular weights of 3.0x 106and 1.3x 106 (Fig. 5, lane B).MAV-2(0) isanontransforming
virus withabtypeRNAgenome(26). Thelarger
RNA species represents genome-length RNA, whereas the smaller speciesrepresentsenvelope mRNA(see below). None of these RNA species
wasdetected in uninfected cells (Fig.5,lane A).
In PR-E 95-C virus-infected cells, we detected
three RNA species, which had molecular weights of 3.4x 106, 1.9x106, and0.7x 106 (Fig. 5, lane C). Since PR-E 95-C viruswas derived
fromRSVPr-C,weassumed that the three RNA
species corresponded to the RSV PR-CRNAs ofcomparable sizes. For clarity, Table 3shows the different virus RNAs detected and their probablecoding capabilities.
Amounts of virus-specific RNAsisolated from the cytoplasm of infected cells. The
amountof each RNA species detectedonDBM paper was measured by hybridization to the cDNA70probe, which represented the 70-nucleo-tidesequencefrom the
tRNAtp
DNAinitiationsitetowithin 30nucleotides of the 5' terminus ofRSVPr-C RNA. This cDNA is
complemen-tary to partofthespliced 5'-terminalsequence
and ispresentonlyoncein eachmRNA species
(9). Therefore, theamountof radioactivity hy-bridizedtoeach RNAspecies should reflect the
amount of the RNA present. The RSV Pr-C cDNA70 probe used in these experimentswas3.4
and94% resistant tonuclease S1 after hybridi-zation in theabsence andpresenceofexcess60S
RSV Pr-CRNA, respectively. Inaseparate
ex-periment, 40% of the RSV Pr-C cDNA7oprobe
was found to be resistant to nuclease S1 after hybridizing to 60S MAV-2(0) RNA. When this
experiment was repeated with
cDNA1oo,
whichcontained thecDNA7osequenceplus the 30
nu-cleotides at the 5' terminus of 38S RSV Pr-C RNA, 51% of the DNA probe was resistantto
nuclease Si. These results indicate that MAV-2(0) and RSV Pr-C share in part the same 5'
terminal sequence; 65% of the cDNA7o probe
wasresistanttonucleaseS1afterhybridization
to60S PR-E 95-C virusRNA.
The cDNA7o probe (in 80-fold excess) was
hybridized topoly(A)-containing RNA coupled
toDBM paper, asdescribed above. The paper
waswashedthoroughly andcutintostrips, and the radioactivity was measured with a liquid
scintillationspectrometer.We detected thesame
i_6X
;'Lo
2:1 2:
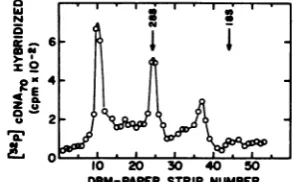
5 DSM-PAPER STRIPNUMBERFIG. 6. Hybridization of cDNA7o to RSV Pr-C RNA from infected cells. Chicken embryo fibroblasts were infected with freshly cloned stock of RSV Pr-C,
[image:6.510.281.431.245.336.2]ar~d the cytoplasmic poly(A)-containing RNA was isolated and fractionatedas described in the legend to Fig. 5; 10 nmol of RNA was fractionated on the agarose gel. The amount of virus-specific RNA pres-ent on the DBMpaper wasdetermined by hybridizing to 1.2 x10Pcpm of thecDNA70 probe(2,500)cpm/pmol) for 72 h at65°0, washing, cutting into 2-mm strips, and counting in a liquid scintillation spectrometer.
TABLE 3. Amountofvirus-specificRNA isolatedfrominfectedcells Relativeabundanceh
Molwtof Virus Codingcapacitya RSV
Pr-C
PR-ERNA(X106) RSVPr-
~~~~~~and
tdRSV 95-C vi-MAV-2(0)
C Pr_c" rwus
3.4 RSV Pr-C gag,pol,env,src 1 1 1
PR-E95-C
3.0 tdRSV Pr-C
gag,pol,env
1.1 1MAV-2(0)
1.9 RSV Pr-C env,src 0.7 0.3 2.3
PR-E 95-C
1.3 tdRSV Pr-C env 0.3 2.1
MAV-2(0)
1.1 RSV Pr-C src 0.3 0.6
0.7 PR-E95-C src 2.1
aAbbreviations:gag,group-specific antigens;pol,RNA-dependentDNApolymerase;env,
envelope
glycopro-tein; src, sarcoma geneproduct.
b The relative abundances of RNAswerecalculated from thehybridizationdata
presented
inFig.
6through
8.TheRSV Pr-C and PRE 95-C virus3.4x106-daltonRNAsandtheMAV-2(0)3.0x 106-dalton RNAwereset
equalto 1.
'td, Transformation defective.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.510.57.455.468.610.2]728 LEIS, SCHEIBLE, AND SMITH
O6 S -
-z
4(4
2
[image:7.510.66.238.62.175.2]DMPPRSTRIP NUMBER
FIG. 7. Hybridization of cDNA70 to MAV-2(0)
RNA from infected cells. The 32P-labeled cDNA70
probe (8x 104 cpm; 1,000cpm/pmol)washybridized
tothe DBM paper shown inFig.5,laneB, for72 hat
65°C, and theamountofradioactivity hybridizedto viral RNAwasdeterminedasdescribed in thelegend
toFig.6.
0.o
a
Jr -z
10 20 30 40 50 DBM-PAPER STRIP NUMBER
FIG. 8. Hybridization of cDNA70to PR-E 95-C
vi-rusRNAfrom infectedcells. The32P-labeledcDNA70
(8x104cpm;1,000 cpm/pmol)washybridizedto the DBMpapershown inFig. 5,laneC,for72 h at65°C,
and theamountof radioactivity hybridizedtoviral RNA wasdetermined asdescribed in thelegendto Fig.6.
size classes of viral RNAs withcDNA7o and with oligo(dT)-primed cDNA (Fig. 6 through 8). Cells infected withafreshly cloned stock ofRSV
Pr-C which was free of transformation-defective
virus contained three RNAspecies(Fig. 6).The
3.4x
106'-,
1.9x106-, and1.1x 106-daltonspecieswere presentin aratio of 1:0.7:0.4 (Table 3). A
similaranalysis of the RNAs isolated from cells infected withRSVPr-Ccontaining transforma-tion-defective virus yielded aratio of 1:1.1:0.3:
0.3:0.6 for the 3.4 x 106_, 3.0 x 106_, 1.9 x 106_,
1.3 x 106-, and 1.1 x 106-dalton RNA species (datanotshown). In eachcase,themost abun-dant RNAspecies detectedwasgenome-length
RNA(Table 3).
Incontrast,incellsinfected with either PR-E
95-C virus or MAV-2(0), the genome-length
RNAwastheleastabundantspecies. The ratio
of theMAV-2(0)-specificRNAswas1:2.1 for the
3.0 x 106-and 1.3 x 106-dalton species (Fig. 7). The ratio of the PR-E95-Cvirus-specific RNAs
was1:2.3:2.1forthe3.4x 106_,1.9x106_,and 0.7
X 106-dalton species (Fig. 8). These results are
alsosummarized inTable 3. By comparing the
amountsofenvelopemRNA's and theamounts of sarcoma mRNA's to genome-length RNAs (Table 3), we found that the subgenomic mRNA's ofMAV-2(0) andPR-E 95-C virus are three- tofivefoldmoreabundantrelative to ge-nomic RNAthan thesubgenomic mRNA's from RSV Pr-C-infected cells. Ifwe use the ratio of the distribution ofRSV Pr-C RNAs containing
the
transformation-defective
virus RNAs (seeabove), the MAV-2(0) and PR-E 95-C virus subgenomic mRNA'sarefourto eightfold more abundant than genome-length RNA, respec-tively.
DISCUSSION
Therearethree knownpathways for the uti-lization of 38S RSV Pr-C RNA after transcrip-tion in cells:(i) it is packaged in virionsas a60S
RNA
complex;
(ii) it is translated directly toexpress the group-specific antigens and the RNA-dependent DNA polymerase; and (iii) it is processed into 28S and22S RNAs for translation of the
envelope
and thesarcomageneproducts, respectively. The 38S,28S, and 22S RNAs have molecular weights of3.4 x106,
1.9X106,
and1.1 x 106, respectively,and havebeen isolated from polysomes from infected cells (10). Hybridiza-tion studies with gene-specific probeshave in-dicated that the 28SRNA representsthe3' one-half of38SRNA, whereas the22S RNA repre-sents the 3' one-third of 38S RNA (5, 32). A unique feature of the processing of the 38S RNA intothe28Sand 22S RNAs hasbeen thefinding thatatleast100nucleotides from the 5' terminus of38S RNAaresplicedontothe smaller RNAs (8, 15, 21, 32).The recognition that there are two pools of viral mRNA's in cells (unprocessed and proc-essed RNAs) has raisedaquestionas towhether there is amechanism that controls the flowof RNA fromthe formertothelatter. Thistypeof control may beimportant for viruses, since ge-nome-length38SRNA is used both forprogeny and for translation ofsomeof the viralproteins. We have proposed previously that the viral protein p19 controls the level of processing of 38S RNA into 28S and 22S RNAs in cells by binding to processing sites on 38S RNA and preventing theircleavage bytheprocessing en-zyme(12).Thebindingof
p19
tothe virus RNA may represent one of the first steps in virion assembly.The evidence which supports this hy-pothesisis asfollows.(i)RNase IIIcleaves38S RNA at sitessimilar to theprocessing sites. We have found that E. coli RNase III cleaves 38S RSV Pr-C RNA in vitro intopoly(A)-containingRNAspeciesof 1.8 x 106and 1.0 x
106
daltons, asanalyzed on 1%0)
IN1
4- I 1
2
-J. VIROL.
vf
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.510.89.233.259.348.2]729
agarosegels containingmethylmercury
hydrox-ide (S. Johnson,
unpublished
data). Inaddition,
RNase III cleaves 35S MAV-2(0) RNA (3.0 x
106 daltons) into poly(A)-containing RNA spe-cies of1.05 x 106 and0.86 x 106 daltons.The in vitro-produced RNA speciesareapproximately 100,000 daltons (350 nucleotides) smaller than the virus-specific mRNA's detected ininfected cells(Fig.5through 7), with the exception of the smallest MAV-2(0) RNA. Based on this size difference, we have
speculated
that RNase III recognizessplicing cleavage siteson viralRNAbut
probably
doesnotcatalyze
theligation
re-action. RNase III has also been showntocleave other viral RNAs. For instance, vesicular sto-matitis virus40S RNA is cleaved by RNase III into five RNAspecies whichare similarin size tothe knownvesicular stomatitis virus mRNA's found in cells(33). RNase III also cleaves polio-virus RNA. The cleavage sites appear to be
intercistronic,
asdeterminedbyoligonucleotide
mapping.AnRNase IIIfragment that encodes
the
poliovirus
polymerase
has beenisolated andtranslated in vitroto
yield
apolypeptide
the size of the viruspolymerase (M. Stewart,
R.Crouch,
and J. Maizel,
Virology,
inpress).
Apparently,
E. coli RNase III
recognizes
some feature of viral RNAs(probably
secondary structure) that ispresent inprocessing sites.(ii) p19 blocks the cleavage of 38S RNA by RNase III (12). In agreement with the results describedabove, AMV p19 has been shown by direct observation inanelectronmicroscopeto
indto
poliovirus
RNAatsitestowhichRNaseIII binds (R. Lenket al., personal
communica-tion).
(iii) RSV Pr-C p19 binds to avariety of un-processedprecursorRNAs with apparent asso-ciationconstants
ranging
from 109to1010 liters/
mol, whereas it doesnotbind or bindsweakly (apparent associationconstants,
107
liters/mol) tomature orprocessed
RNAs(12).(iv)p19is the first protein to be translated in response to 38S RNA since it is at the amino terminusof thegroup-specific antigenprecursor polypeptide.
Toprovide furthersupport forarole for p19 in regulating processing of 38S viral RNA, we studied theRNA-binding properties of RSV Pr-C,MAV-2(0), and PR-E 95-C virusp19proteins and analyzed the sizes and amounts of virus-specific poly(A)-containing mRNA's from the cytoplasm of cells infected with these viruses. The other viruses have higher levels of envelope glycoproteinin theirvirions than RSV Pr-C (24, 29). If p19 inhibits the processing of genome-length RNAs to
smaller
RNAs, wepredict that (i) the p19 proteins from MAV-2(0) and PR-E95-C virus have lowerbinding affinities for viral RNA than RSV Pr-C p19, (ii) there is no de-tectable difference in the binding affinities for 60S RNA by the p12 proteins from all three viruses,and(iii) the levelsofallof the processed mRNA'sareincreasedrelativetogenome-length RNA in MAV-2(0)- and PR-E 95-C virus-in-fected cells compared with RSV Pr-C-infected cells.
These predictions have been verified. We found that(i)theMAV-2(0) and the PR-E 95-C virus p19 proteins have 10-fold lower binding affinities for viral RNA thanp19 purified from RSV Pr-C, (ii) there is no difference in the binding affinities for RNA of the p12 proteins from thedifferent viruses, and (iii) in MAV-2(0)-and PR-E95-C virus-infected cells, the level of processed mRNA's is increased relative to ge-nome-length RNA
compared
with RSV Pr-C-infected cells.We detected three viral RNAs in RSV Pr-C-infected
cells;
these RNAs had molecular weights of3.4 x106,
1.9 X106,
and 1.1 x106,
whichwasinagreementwith the results of Lee et al. (10). In MAV-2(0)-infected cells we de-tected 3.0 x
106-
and 1.3 x106-dalton
RNA species. The former is genome-length RNA, whereas the latter RNA isthe envelope mRNA, basedonthefollowing
evidence. Translation of thetwoRNAs inalow-background reticulocyte cell-freeprotein-synthesizing
systemfollowed by immunoprecipitation and sodiumdodecyl
sul-fate gel
electrophoresis
indicated that the3.0 x106-dalton
RNA gave rise to a 76,000-daltonpolypeptide whichwasrecognized by antiserum tothe avian sarcoma virus
group-specific
anti-gens.
The 1.3 x106-dalton
RNA gaverise toa60,000-dalton
polypeptide
that wasrecognized
by antisera raised
against
AMVenvelope
gly-coprotein
(Scheible, unpublished
data). We de-tected3.4x 106-,1.9x106_,
and0.7x106-dalton
species in PR-E 95-C virus-infected cells. It is likely thatthese RNAscorrespondtothe RNAs found in RSV Pr-C-infectedcells since (i) PR-E 95-C virusisderivedfromRSV
Pr-C, (ii)
it isatransforming
virus, and(iii)
the number andsizes of the RNA
species
found in PR-E 95-C virus-infected cells are similar to the number and sizes of the RSV Pr-C RNAs. Each of the subgenomic RNA speciesdescribed above con-tains RNAsequencesderivedfrom the 5' and 3' termini ofgenome-lengthRNA.We measured the concentration of mRNA speciesisolated from chronically infected cells byhybridizationto acDNA7oprobe,which was complementary to part of the
spliced
5' RNA sequence and was present at a level ofonlyone copy per RNA.Thus,the amount of radioactive 35,on November 10, 2019 by guest
http://jvi.asm.org/
730
LEIS, SCHEIBLE, AND SMITHprobehybridizedshould havebeenproportional tothe number ofpoly(A)-containing RNA mol-eculespresentregardless of the size oftheRNA. Wehave expressed thesedataas aratio of the amountsof thedifferent size classes of RNAto avoidproblems in determining absoluteamounts of mRNA's incells. To determine the absolute amountsof mRNA's in infectedcells,wewould require information about thenumber of viable cellsinculture,the numberofparticlesproduced percell, the ratio of infectioustononinfectious virus particles, the recovery of RNA from in-fected cells, the number of virus-specific RNA molecules containing poly(A), and the number ofproviral equivalents per cell. In the present workwe examined thesteady-state level of ge-nomic and subgenomic virus mRNA's in chron-ically infected cells. This approachprovided an estimateof the relative abundance of virus-spe-cific mRNA's in cells chronically infected with threedifferent viruses. We found three-to five-fold more subgenomic mRNA's than genome-length RNAs in MAV-2(0)- and PR-E 95-C vi-rus-infected cells than in RSV Pr-C-infected cells. Therefore, it appears that there is en-hanced processing of genome-length to subge-nome-length mRNA's in MAV-2(0)- and PR-E 95-C virus-infected cells. This correlates with the existence ofp19 proteins with lower binding affinities for viral RNA in these two strains. Theseresultsareconsistent with thehypothesis thatthe p19 protein controls processing of viral RNA incells. Further workalong these lines will involve a correlation of the appearance of p19 with theappearanceof subgenomicmRNA mol-ecules
early
after infection of cells withdifferent viruses.ACKNOWLEDGMENTS
Weespeciallythank Maxine Linial forproviding prepara-tionsof PR-E 95-C virus. We also thank Sue Nebes and Olin Fox for technical assistance and Linda VanEldik forproviding purifiedMAV-2(0)polypeptides.
Thisinvestigationwassupportedby research grant PCM 77-00809fromthe National ScienceFoundation, bygrant VC-310from the American CancerSociety,andbyPublic Health Service grant CA12323from the National Institutes of Health. P.S. wassupportedby Public Health Service Training Grant 5T32 CA 09111from the National InstitutesofHealth.
LITERATURE CITED
1. Bailey,J., and N. Davidson.1976.Methylmercuryasa reversibledenaturing agent for agarose gel electropho-resis.Anal.Biochem.70:75-85.
2. Beemon, K., and T. Hunter.1977.In vitro translation yieldsapossibleRous sarcomavirus src geneproduct. Proc.Natl. Acad. Sci.U.S.A.74:3302-3306.
3. Brugge, J., A. Purchio, and R. Erikson. 1977. Virus-specificRNAspecies presentinthecytoplasmof Rous sarcomavirus-infected chickencells.Virology 83:16-26. 4. Green,R., and D.Bolognesi.1974.Isolationofproteins bygelfiltration in 6Mguanidinium chloride:application toRNAtumorviruses.Anal.Biochem. 28:479-490.
5. Hayward, W. 1977. Size and genetic content of viral RNAs in avianoncornavirus-infected cells. J. Virol. 24: 47-63.
6. Hizi, A., J. Leis, and W.Joklik. 1977. The RNA-de-pendent DNA polymerase of avian sarcoma virus B77. III.Binding of viral and non-viral RNAs to theafi,/2 andaforms of enzyme. J. Biol. Chem. 252:6878-6884. 7.Jamjoon, G.,R.Naso,and R.Arlinghaus. 1977.
Fur-thercharacterization of intracellular precursor polypro-teins ofRauscher leukemia virus. Virology 78:11-34. 8. Kerr, I., U. Olshevsky, H. Lodish, and D. Baltimore.
1976.Translation of murine leukemia virus RNA in cell-free systems from animal cells. J. Virol. 18:627-635. 9. Krzyzek, R., M.Collett, A. Lau, M. Perdue, J. Leis,
and A. Faras. 1978. Evidence for splicing of avian sarcoma virus 5' terminal genomic sequences onto viral-specific mRNA in infectedcells. Proc. Natl. Acad. Sci. U.S.A. 75:4327-4331.
10.Lee, J.,H.Varmus,and J.Bishop. 1979. Virus-specific messenger RNAs in permissive cells infected by avian sarcomavirus.J. Biol. Chem.254:8015-8022. 11. Leis, J. 1976. RNA-dependent DNA polymerase activity
ofRNA tumor viruses. VI. Processive mode of action of avianmyeloblastosis virus polymerase. J. Virol. 19:932-939.
12. Leis, J., J. McGinnis, and R. Green. 1978. Rous sar-coma virus p19 binds to specific double-stranded regions ofviral RNA: effect of p19 on cleavage of viral RNA by RNase III.Virology 84:87-98.
13.Leis, J., R.Smith,P.Dierks,J.Parsons,M.Collett, and A.Faras. 1978.Invitrotranscriptionof reconsti-tuted 35SRNA-tRNA,,ptemplate-primercomplexes by the avian oncornavirus DNApolymerase.Effect of tem-peratureonthesize of the DNAtranscripts.Virology 85:28-42.
14.McGinnis, J., A. Hizi, R. Smith,and J. Leis. 1978. Translation ofa180,000 daltonRSVprecursor polypep-tidecontainingreversetranscriptaseandgroup-specific antigens in response to34S viral RNA.Virology84: 518-522.
15.Mellon, P., and P.Duesberg. 1977. Subgenomic cellular Rous sarcoma virus RNAs contain oligonucleotides fromthe 3' half and the 5'terminus of virion RNA. Nature(London)270:631-634.
16. Oppermann,H.,J.Bishop,H.Varmus,andL. Lev-intow. 1977. Ajoint productof the genes gag andpol ofaviansarcomavirus:apossibleprecursor ofreverse
transcriptase.Cell12:993-1005.
17. Paterson, B.,D.Marciani,and T.Papas. 1977. Cell-freesynthesisof the precursorpolypeptide for avian myeloblastosis virus DNA polymerase. Proc. Natl. Acad. Sci. U.S.A. 74:4951-4954.
18. Pawson,T.,R.Harvey,and A. Smith.1977.The size of Roussarcomavirus mRNAs active incell-free trans-lation. Nature(London)268:416-420.
19. Purchio,A.,E.Erikson,J.Brugge,and R.Erikson. 1978. Identification ofapolypeptide encodedby the aviansarcoma virus srcgene. Proc.Natl. Acad. Sci. U.S.A. 75:1567-1571.
20. Purchio,A.,E.Erikson,andR.Erikson.1977. Trans-lation of 35S and ofsubgenomicregionsof avian
sar-comavirus RNA.Proc. Natl. Acad.Sci. U.S.A. 74:4661-4665.
21. Rothenberg, E., D. Donoghue, and D. Baltimore. 1978.Analysisofa5'leadersequenceonmurine leu-kemia virus21S RNA:heteroduplexmappingwithlong reversetranscriptaseproducts.Cell13:435-451. 22. Scatchard,G.1949.Theattractionofproteinsforsmall
moleculesand ions.Ann.N. Y. Acad. Sci.51:660-672. 23. Sen, A.,andG. Todaro. 1977.The genome associated
specific RNAbinding proteinsofavian andmammalian type-Cviruses.Cell10:91-99.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
24. Shaikh, R., M. Linial, S. Brown, A. Sen, and R.
Ei-senman.1979.Recombinant avian oncoviruses.II.
Al-terations in thegagproteinsandevidence forintragenic recombination.Virology92:463-481.
25. Smith, R., and E. Berstein.1973.Productionand puri-fication oflargeamountsof Roua sarcomavirus.Appl. Microbiol. 25:346-353.
26. Smith, R., LDavids, andP. Neiman. 1976.Comparison ofanavianosteopetrosisvirus withanavian lympho-matosis virusby RNA-DNA hybridization.J. Virol.17: 160-167.
27. Smith, R., S. Nebes,and J. Leis. 1977.Production of largeamountsof35S RNAandcomplementary DNA from avian RNAtumor viruses. Anal. Biochem. 77: 226-234.
28. Stacey, D., V.Allfrey, and H. Hanafusa.1977. Microin-jection analysis of envelope messenger activities of avian leukosisvirus RNAs.Proc. Natl.Acad. Sci.U.S.A. 74:1614-1618.
29. VanEldik, L., and R. Smith.1978.Isolationand
char-acterization of the envelope glycoprotein ofanavian
osteopetrosisvirus: effect of hostcell onantigenic
reac-tivity. Virology90:80-89.
30. Van Zaane, D., A. Gielkens, W. Hesaelink, and H. Bloemers. 1977. Identification of Rausher murine leu-kemia virus-specific mRNAs forthe synthesis of
gag-andenv-geneproducts. Proc. Natl. Acad. Sci. U.S.A. 74:1855-1859.
31. Wahl, G., M. Stern, and G. Stark.1979.Efficient
trans-feroflarge DNA fragments fromagarosegelsto diazo-benzyloxymethyl-paper and rapid hybridizationby
us-ingdextran sulfate. Proc. Natl. Acad. Sci. U.S.A.76: 3683-3687.
32. Weiss,S., H. Varmus, and H. Bishop. 1977.Thesize andgeneticcomposition ofvirus-specific RNAs inthe cytoplasm ofcells producing avian sarcoma-leukosis
virus.Cell 12:983-992.
33. Wertz, G.,and N. Davis. 1979.RNaseIII cleaves vesic-ular stomatitisvirusgenome-lengthRNAs but failsto
cleave viral mRNA's. J. Virol. 30:108-115. VOL. 35, 1980