CopyrightX)1977 AmericanSociety for Microbiology Printed inU.S.A.
Changes
in
Synthesis
of DNA-Binding
Proteins
During
the
Onset
of
Transformation in NRK Cells Transformed by a
Temperature-Sensitive
Mutant of Rous Sarcoma Virus
BRUCE E. MAGUNI* AND PRESTON H. DORSETT
Department ofAnatomy, University of Tennessee Center for the Health Sciences, Memphis, Tennessee38163,
and Department ofMicrobiology, University of Tennessee Center for the Health Sciences, Memphis, Tennessee 38163
Received for publication 23 November 1976
Synthesis
of cytoplasmic DNA-binding proteins was investigated after a shift
from the
nonpermissive to the permissive temperature in NRK cells
trans-formed
by a
temperature-sensitive mutant of Rous sarcoma virus [ts339(RSV)].
Cells were labeled for several generations in [3H]leucine and were
pulse-labeled
with
['5S]methionine
for 1 h at the
nonpermissive temperature
(390C)
and at the
permissive
temperature
(330C,
5 h after
shift from
390C).
Proteins binding to
sequential columns of
double-stranded and single-stranded DNA-cellulose were
examined by polyacrylamide gel electrophoresis in the presence of sodium
dodecyl sulfate, and the 5S/3H ratios were obtained for
each
column fraction
and
for individual
polypeptides. The protein fractions binding to single-stranded, but
not
double-stranded, DNA and eluting at high salt concentrations (greater than
0.60
MNaCl) showed elevated 35S/3H ratios. This
indicated increased synthesis
of these
proteins
within 5 h after the onset
of
transformation. The
majority of the
polypeptides in these fractions showed
increased synthesis as a consequence of
transformation. One prominent polypeptide among them
constituted
0.1%of
the
cytosol
protein
and had a
molecular
weight
of
93,000. We conclude that the
synthesis of
proteins
binding
tightly
tosingle-stranded
DNA
is
increased
early
after
the onset of transformation.
Cells
transformed
by
temperature-sensitive
(ts)
transformation
mutants
of Rous sarcoma
virus
(RSV) permit the study of sequential
events
that
occurduring
the course of
transfor-mation
(3, 11, 13, 14, 20, 27). Most
of these
studies have
focused
on
early
membrane-re-lated changes
that occur during the onset of
transformation in such systems.
Recent
experimental data
have
raised
the
possibility
that
viral-coded DNA-binding
pro-teins may
be
involved
inthe process
of
transfor-mation.
Cells
infected or
transformed
by
hu-man
adenovirus
types 2
and 5
produced
two
new
DNA-binding polypeptides (25, 21); Tenen
et
al.
(29)
demonstrated
that the
thermosensi-tive
defect
ina ts
A-type mutant
of simian virus
40 was in
the T
antigen. This
DNA-binding
protein (6, 12, 22, 26) attaches at or
nearthe
replication point of simian virus
40DNA
(22).
Although
afunctional
relationship between
transforming
ability
and
DNA-binding
capac-ity of
the
Tantigen has
notyet been
demon-strated,
the
possibility exists that the initial
'Present address: DepartmentofAnatomy, University
ofArizona,CollegeofMedicine,Tucson,AZ 85724.
action
of
the
transforming
gene
product
occurs onthe
cellular DNA.
DNA
affinity
chromatography has been
used
in
the
purification of procaryotic and eucaryotic
proteins whose
DNA-related
roles have been
elucidated
(1, 4, 10, 16, 23, 25, 30, 31).
It also has
been
useful
inselecting proteins that may
func-tion in DNA
replication, repair, or
recombina-tion (8).
The
genomes
of T-even
phage
and
herpes simplex
virus
contain a
high proportion
of genes involved with DNA
replication
(19, 23,
28)
and apparently code for a large number of
DNA-binding proteins (5, 10, 21). Hence, DNA
affinity chromatography
has
permitted
the
se-lection, without prior knowledge
of
their
func-tion,
of a group of proteins
that may function
inaspects
of
DNAreplication.
The aim of the
present investigation
was toexamine
alterations
inthe
synthesis
of
DNA-binding proteins after
ashift from the
nonper-missive to
the
permissive temperature,
ina ratcell
line
(NRK)
transformed by
a ts mutantof
the
B77 strainof
RSV,
ts339(RSV)
(9). These
cell cultures were incubated with radioactive
amino
acids from
h 5to
6after
temperature
469
on November 10, 2019 by guest
http://jvi.asm.org/
470 MAGUN AND DORSETT
shift. The
incorporationof radioactivity
intoDNA-binding proteins of shifted cultures was
compared
with that obtained from cultureswhich had been
treated similarly,
but whichhad
beenmaintained
atthe
nonpermissive
temperature.
The results of this
investigation
indicate
that the onset of transformation isac-companied
by an increased incorporation ofla-bel
into asmall
portionof the total
DNA-bind-ing
protein complement and that these proteins
bind
to
single-stranded DNA,
but not todouble-stranded DNA.
MATERIALS AND METHODS
Cells and culture. NRK cells and NRK cells transformed by ts339(RSV) wereobtained from W. K. Joklik, Duke University. The thermosensitive transformed cell line [ts339(RSV)NRK] had been established from a clone of transformed cells in soft agar (9).The cell lines werepropagatedinDulbecco minimal essential medium containing 10% heat-in-activated(560C, 30min)fetalcalf serum. The cells wereroutinelypropagatedasmonolayer cultures in 75-cm2 flasks.
To obtain sufficient cells, roller bottle cultures were utilized. Roller bottles (685 cm2) were seeded
with 1 x 107to 2 x 107ts339(RSV)NRKcells. NRK
roller cultures were prepared byseeding plastic bot-tles (490cm2)with5x 107to 10 x 107cells. Since the NRK cells did not attach well at390C, allcultures wereincubated for the initial 18 to 24 h at360C and then incubated at 390C. The culture fluid was
changedat 24- to 48-h intervals; thestatus ofthe
monolayer was monitored microscopically each day. Afteraconfluent monolayerhad beenobtained, the cultures were incubatedanadditional 48 h without
feeding to establish contact inhibition. The
ts339(RSV)NRK cells routinely established a
con-fluent monolayer within 3 to 4 days, whereas the
NRKcells routinelyrequired 6 to8days.
Cell culture reagents were purchased from Micro-biological Associates, Rockville,Md.
Preparation of cytosol. Cells were harvested by scraping, washed twice in cold phosphate-buffered
saline at low speed, and allowed to swell for30min in 2volumes of hypotonic buffer (1 mM MgCl2, 0.4
mM CaCl2, 0.5 mM dithiothreitol, 1 mM
phenyl-methylsulfonyl fluoride, and 25mM Tris-hydrochlo-ride, pH 7.9) at
00C.
The cells weredisrupted in a Douncehomogenizer with a "B" pestle, after which 2 volumesof0.05 MNaCl,0.1%Brij 58,1mM phenyl-methylsulfonyl fluoride, 1 mM EDTA, 10 mM /3-mercaptoethanol, 10% glycerol, and 10 mM Tris-hydrochloride, pH 8.1 (buffer A) was added. The homogenatewascentrifuged at 25,000xg for 15min and again at 100,000 x g for 1 h, after which the supernatantwaswithdrawn and made to contain 1.7 MNaCl. Polyethylene glycol-6000 was added to a final concentration of 10% for precipitation of DNA. Afterincubationat0°Cfor30min, the samples were clearedby centrifugation at 7,400 x g for 15 min. The resulting supernatant was dialyzed against threechanges of50volumesof buffer Aat4°C. Thesupernatant that formed after centrifugation at 20,000 xgfor 20 min wasdesignated as the cytosol fraction.
Preparation of DNA-cellulose columns. Native and denatured DNA-cellulose preparations were made bythe combined methods of Litman (16) and Alberts and Herrick (2). To prepare native DNA-cellulose, 50 mg of calfthymusDNA (Worthington
Biochemicals,Freehold, N.J.)wasdissolvedin 18ml of buffer B (0.01 MTris-hydrochloride, 1mMEDTA, pH 7.4).Six gramsof Cellex410(Bio-Rad Laborato-ries, Richmond, Calif.) was added, and the slurry
wasdriedat45°Covernight. The dried DNA-cellu-lose was pulverized and resuspended in ethanol,
followed byUV irradiation, as described (10). The dried native DNA-cellulose wasswollen for 2 h in buffer Bat4°Cand then washedbycentrifugation several timesinbuffer A before being packed into columns.Denatured DNA-cellulosewasprepared by heating 1.3 mg of calf thymus DNA per mlin0.05M NaOHat100°C for 10min,followed by rapid chilling
in ice water.After the pH had beenadjustedto7.2,1 gof Cellex 410wasaddedtoeach3mlof denatured DNA, andthe slurry was dried overnight at room temperature. The DNA-cellulosewas mixed again with denatured DNA and driedasbefore. The dena-tured DNA-cellulose was resuspended in buffer A and washed several timesbycentrifugationand
re-suspension. Native and denatured DNA-cellulose contained atleast 2 mg of DNA perpackedml of columnvolume. Swollen DNA-cellulose (denatured ornative)waspackedinto glasscolumns (0.7by 8 cm)containingavolume of3ml.Native DNA-cellu-lose was overlaid with 1 ml of plain Cellex 410, whichwasremoved before elution of bound protein toremoveproteinthat hadboundnonspecificallyto cellulose before elution of thespecifically bound pro-tein.
DNA-cellulose chromatography. Thirty to forty
milliliters of cytosol in buffer A was pumped
through the column ofnative DNA-cellulose at a
flowrateof12 ml/h.After it had beenloaded, the
column was flushed with 17 ml of buffer A. The upper plain cellulose pad was removed, suspended
in 5mlof water,andassayed for radioactivityinan aqueouscocktail. Less than1%of the total column-bound protein was adsorbed to the plain cellulose
pad. The native DNA-cellulose column was then eluted with successive 10-ml volumes of buffer A containing 0.1, 0.15, 0.25, 0.60, and 2.0 M NaCl. Eluateswereconcentratedto 1 ml in an ultrafiltra-tiondevice (Amicon MMC) containingaPM10 filter. Five percentsodium dodecyl sulfate (SDS), contain-ing5%
/3-mercaptoethanol,
wasaddedtothe concen-trated sample, which was then heated in boiling waterfor1minanddialyzed against electrophoresis sample buffer (0.01Mphosphate, pH 7.2,0.1%SDS,1% mercaptoethanol). Theradioactivity remaining inthe column afterthe 2.0 Msalt elutionwas mea-suredbysuspendingthecolumncontents indistilled waterand assayingaportion forradioactivity. All unretained cytosol from the native DNA-cellulose column was then pumped through a similar-sized column packed with denatured DNA-cellulose,
washed, and elutedasdescribedfor the native
on November 10, 2019 by guest
http://jvi.asm.org/
DNA-BINDING PROTEINS AND
471
cellulose columns. Eluted fractions were concen-trated and dialyzed against electrophoresis sample buffer.
Scintillation spectrometry. Aqueous samples
wereadded to 10 ml of scintillation mixture, 1 part Triton X-100, and 2 parts toluene containing 2,5-diphenyloxazole (4.9 g/liter) and bis-MSB (0.1 g/ liter). Gels to be assayed for radioactivity were fro-zen ondryiceand stored at -20'C. The gels were sliced transversely into1-mmwafers with a Hoefer gel slicer (15 cm long). Each slice was incubated overnight with0.5 mlof hydrogen peroxide at450C
in atightly capped polyethylene vial. After 24 h, each vial received the scintillation cocktail and was counted in a scintillation spectrometer (Beckman LS230). Under theseconditions,3Hefficiencywas at least 22%. 35Sand 3H activity were determined
si-multaneously by using the channels ratio method with an externalstandard.
Protein determination. Protein was determined according to Lowry et al. (17) with bovine serum albumin asstandard. Because of the small amounts of proteinin the DNA-binding protein fractions, it wasnecessary to measure protein content bythe use
ofradioactivity. Allcultureswere incubated for at
least six generations in[3H]leucine (1.0
,.tCi/ml).
A small amount of cytosol protein was dialyzed against 0.1 N NaOH and assayed both for protein and tritium. Withthese conditions, 800 dpm of 3H perjig
of protein wasobtained, and this valuewas foundtobereproducible within 5% amongseveral experiments. Because of the small amounts of pro-tein used for electrophoresis, radioactivity assay proved to be the most accurate means of measuring protein concentration and was applied throughout this investigation.Polyacrylamide gel electrophoresis. Protein frac-tionsinsample bufferwereexaminedby polyacryl-amide gelelectrophoresis (PAGE)inthe presence of SDS accordingtoLaemmli(15). The separationgel was 12.5 cmlong and contained 8.75% acrylamide and 0.23% methylenebisacrylamide. Molecular-weight marker proteins were as previously de-scribed (18).
RESULTS
Phenotypic
changes aftertemperature
shift. The experiments described
inthe
presentinvestigation are
based
onthe induction of the
transformed
stateby alteration of
incubating
temperatures
from the nonpermissive
(390C)
tothe
permissive(33°C)
temperature. It wasnec-essary
first
toexaminecellular
properties
phe-notypic
of transformation
as atemporal
conse-quence oftemperature change.
When grown at
39°C,
thets339(RSV)NRK
cells formed
a confluentmonolayer
similar tothat of nontransformed cells
(Fig.
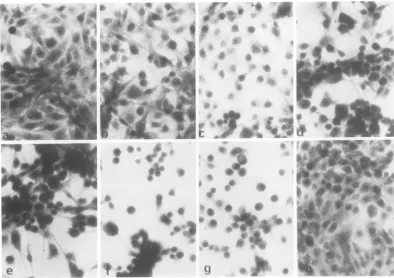
1). After thetemperature was
shifted
to33°C,
the cellsas-sumed a
transformed
phenotype,
which wasevident as
early
as 6 h after the temperatureshift
(Fig.
ic). Cells migrated
intoclustersand
acquired
arounded morphology instead of theflat, stellate shape that
wascharacteristic
oftheir
growth
at 39°C.Twenty-four
hours aftershift to
the
permissive
temperature,the
trans-formed appearance was seen in
its
extremeform. Cellsthat had
been
maintained
at39°C
throughout the
courseof the experiment
main-tained
their untransformed
appearance(Fig.
lh).
As
ts339(RSV)NRKcells
approachedcon-fluency
at39°C, their
rateof
DNAsynthesis
apparently
decreased,
asdemonstrated by
the
reduction in
incorporation
of[3H]thymidine
into acid-precipitable
material(Fig.
2).How-ever, if
parallel cultures
were shifted to33°C,
the
incorporation
of[3H]thymidine began
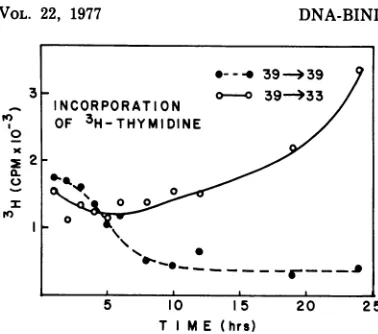
toin-creasewithin 6 h and continued to rise over a
24-h
time period.
Itthus
appeared that a shifttothepermissive temperature was able to
stimu-late DNA replication of
ts339(RSV)NRK
cells
within a few hours after the temperature
change.
Fractionation
ofDNA-binding
proteins.DNA-binding
proteins
wereobtained byperco-lation of cytosolthrough sequential columns
of
native
(double-stranded)
and denatured(sin-gle-stranded) calf thymus
DNA-cellulose,
afterwhich the bound proteins wereeluted
stepwise
with
increasing
saltconcentrations. Elution
profiles of the
DNA-binding
proteins fromna-tive DNA showed
discrete
peaks of protein,which were removed by each successive salt
wash(Fig. 3A). The 2.0 M NaCl wash contained
little
detectable
protein.The
nativeDNA-cellu-lose column bound approximately 4% of the
total
cytosol protein,
96% ofwhich wassubse-quently recovered
inthe NaCl
eluates
(Table 1). An additional 4% of the cytosol proteinad-sorbed
todenatured DNA,
and 90%of
theseproteins were
recovered
inthe NaCl
eluates(Fig.
3B, Table 1). In contrast to the elutionprofile from native
DNA-cellulose, the
2.0 MNaCl eluate from denatured DNA
comprised
asizable peak.
Approximately
half of thepro-teinsbinding to denatured DNA were removed
in
the
0.10 MNaCl
wash.
The sizes of the
DNA-binding protein fractions
weresimilar,
whethercells had been grown at
39°C
or had beenshifted to
33°C
for 6 hbeforeharvesting.
PAGE
of
DNA-binding proteins. The
DNA-binding proteins
ineach
NaClfraction(except
the 2.0 M NaCl wash from native
DNA,
whichcontained insufficient
protein
forcharacteriza-tion)
were examinedby
PAGE. Each fractionwasfound to contain a
unique
polypeptide
frac-tion
when
compared
witheither
the otherDNA-binding
proteinfractions
or theunfrac-tionated
cytosol
proteins (Fig. 4).
Whenon November 10, 2019 by guest
http://jvi.asm.org/
472
MAGUN AND DORSETTV
1!9*
W
a
4_-Ft . r
4 4~~~~~~1.7
OWN
*#.>*'i
',
e
%
4it~~~~~~t;
* i.' * y t~~~~~~~~~~t~ * t* I
S
0._ :w. lb a
mn1~~~~~~~~~~~.4
.4
4
*
e
I*S
s~
ha,
U:
V
4
P,*0
~
'4k
.,i1
g
0
o
FIG. 1. Morphological changes ints339(RSV)NRK cells afterashift from 39to33°I. Slide culturesof
ts339(RSV)NRK cells were incubated at 39°C until a confluent monolayerwasobtained,after whichsomeof
the cultures were shifted to
33XC
and some remained at390C. Slide cultureswerefixed and stained with hematoxylin and eosin at the time ofshift to33XC(a), and at 3 h (b), 6 h (c), 9 h (d),12h (e),24 h(I), and 48h (g)after the shiftto33C.Similar culturesmaintainedat39°Cwerealsofixedandstained at thesametimes. These cultures all appeared similar to those in(h), which showsacellculture maintainedat39°Cthroughoutthe 48-h experiment.
pared
with the unfractionated
cytosol proteins,
most
of the DNA-binding proteins
wererich
inhigh-molecular-weight
polypeptides
(larger
than 80,000). Comparison of cytosol proteins
before
and
after
DNA-cellulose
chromatogra-phy
(Fig. 4)
showed
few changes, indicating
that the
DNA-binding
polypeptide
profiles
rep-resented
anamplified
view
of minor
compo-nents present in
the
whole cytosol. Although
polypeptide profiles obtained
from cells
shifted
from 39 to
330C
for 6 hshowed
some notabledifferences when
compared with
profiles of
pro-teins
obtained from cultures
maintained
at3900,
the
profiles obtained
atboth
tempera-tureswere
remarkably similar.
Incorporation
oflabeled
amino acids intoDNA-binding
proteins.
Ifchanges
in thesyn-thesis of
DNA-binding proteins
occurafter
ashift
tothe
permissive temperature,
itmight be
expected
that incorporation
of radioactive
amino
acids
intothose DNA-binding proteins
also
would
reflect changes. To
testthis
hypoth-esis,
twosetsof
ts339(RSV)NRK cells
werein-cubated with
[3H]leucine
at390C
for six
genera-tions
topermit uniform incorporation
of
[3H]leucine
into
cellular protein. One
setof
these
cultures
waspulse-labeled
with
[35S]methionine
from h
5 to 6after
ashift
to330C.
The
35S/3H
ratios
of
the
DNA-binding
pro-teins
synthesized
after the shift
to33WC
werecompared
with
the
35S/3H ratios
of
DNA-bind-ing proteins
from the other
setof cultures
thathad been
maintained
at390C
and
incubated
with
[35S]methionine
for 1 h.Comparison
oftheI5S/3H
ratiosfrom
the twoexperimental
groups,(35S/3H
at330C)/(35S/3H
at3900),
referred
tohenceforth
asR,should be
anindicator of
spe-cific
protein fractions
whose
rateof
synthesis
becomes altered after
atemperature shift.
In the
ts339(RSV)NRK
cells, a shift from 39 to330C
resulted
indecreased
Rvalues,
ranging
from 0.90 to 0.98for the
cytosol
andDNA-bind-ingprotein fractions, except for the 0.60
and
2.0MNaCl
eluates
fromdenatured
DNA(Table
2).Thelatter fractions had
elevated
Rvalues(1.36and 1.23,
respectively),
indicatingthat,
inthese$4
0.
0.
Pn.?.''
'12
.s
.-c..o
*0 51.
4.,. W.
.0 el
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.62.456.72.350.2]3
It) 0
x
a-I
QC
2
5 10 15
T I M E (hrs)
FIG. 2. Synthesis of DNA in ts339 cellsafterashift fromthe nonpermissil
missive temperature. ts339(RSV)NRK seeded in 60-mm culture plates and i 360C for12to18h.The culturesthenwe at39'C until themonolayers had become Fourteen hours afterthe culture medium replacedwithfreshmedium, one-halfof wereshiftedto33'C.Atthetimeofthe shift(zero time)andat1,2,3, 4, 5,6,E and 24 hafter thetemperatureshift,fou 33and390Cwereincubatedwith
[3H]thy
h. [3H]thymidine (5 uCi) in 0.1 ml o) buffered saline was addeddirectlyto e After the 1-h pulse-labeling period, th scraped from the dishes andpipetted i chloroacetic acid. Theacid-precipitabler collectedonglass-fiber filters, andthe; incorporatedperculturewasdetermined represents theaverage ofthefourculture turemediumwasnotchangedthroughout
fractions,
therewas anetincreasedtion of35S into protein after the to
shift.
In an attempt to determine whet
creased R values of the 0.60 and 2
fractions from denatured DNA occ
consequence ofthe onset oftransfc
thets339(RSV)NRK cells,asimilar
was performed with NRK cells th
been transformed by RSV. All DI
protein fractions showed decreased
ranging from 0.46 to0.65 (Table 2).
tothe increased R values of the0.6(
NaCl fractions from denatured DN
from the ts339(RSV)NRK cells, t
fractions inthe NRKcells exhibited
R values (0.46 and0.48). Since the
R valueisprobablyaconsequencea
enzymaticactivitydue to the lowerE
ture, the increaseinR value in the
ing protein fractions from thets339
cells after temperature shift should
increased synthetic
ratesof
someor all ofthe439 proteins inthese fractions.
433 Isotopic labeling of individual DNA-binding
polypeptides. The DNA-binding protein
frac-tionsfrom the ts339(RSV)NRK cellsinthe
tem-perature shift experiment shown in Table 2
were analyzed by PAGE. Each gel was sliced
transversely into 1-mm slices, and each slice
wasanalyzed for 35S and 3H activity. The
35S/3H
ratioswereobtained for corresponding
polypep-tide bands from shifted and nonshifted
cul-tures, and anR value wascalculated for each
pair of correspondingbands.
20 2 5 The R values ofcorresponding bands ofthe
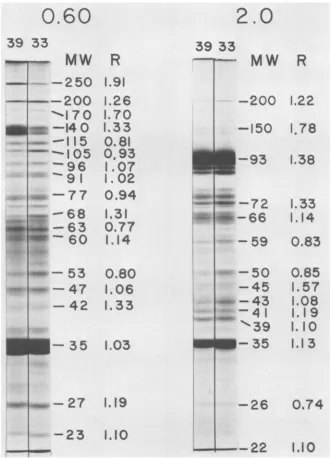
electropherograms
are shown in Fig. 5for the?(RSV)NRK 0.60 and 2.0 M eluates from denatured DNA.
'eto theper- The R values formostbands in these
fractions
cells were were greater than 1, indicating that an
in-tncubated
at creased incorporationof 355 intothesepolypep-!reincubated tides occurredafter the shiftto330C.Two poly-se confluent. peptides, which exhibited elevated R values in
thecultureens
Fig.
5,
werepresent
inrelatively large
temperature
amounts.One,
which hadanapparent
molecu-,p10,
12,18 larweight
of
35,000,
waspresent
in the0.25,
orculturesat 0.60, and 2.0 M eluates from denatured DNA
vmidinefor1 (Fig. 4)andcomprisedabout 0.75% of thetotal
f
phosphate- cytosol protein. The other major band, which'ach culture. hada molecular weight of 93,000, was eluted Le cells were from denatured DNAby2.0 MNaCland
com-nto cold tri-
prised
about 0.1% of the total cytosol protein.materialwas ItsR valuewas1.38.
radioactivity
Corresponding
bands of
otherDNA-binding
'es. The
cupl
polypeptides
weresimilarly
analyzed
for thettthe36h. native and denatured
DNA-binding
proteinfractions. Some DNA-binding polypeptides
ap-parently increased in quantity after a shift to
lincorpora-
thepermissive temperature (Fig. 4); whenthetemperature R values of these polypeptides were
deter-mined, allwerefoundtobeless than 0.90 (data
other the in- not shown),
indicating
that the appearance of.0 M NaCl these bands was probably not due to an
in-urred as a crease in their synthesis. The results for all
rmation in
DNA-binding
protein fractionsaresummarizedexperiment in Fig. 6. The
histogram
of R values for 159Lat had not
DNA-binding
polypeptides (as many as couldNA-binding
be measured accurately), demonstrates thatI R values, most
DNA-binding
polypeptide bandR valuesIncontrast were less than 1, with a mean of 0.90. The
0 and 2.0 M majority ofpolypeptides in Fig. 6 with R values
[A obtained greaterthan1.2werecontained in the 0.60 and
these same 2.0 M NaCl elutions from denatured DNA
d decreased (shown inFig. 5).
decrease in Uptakeof
[35S]methionine
atnonpermissivefdecreased and permissive temperatures. Alterations in
adtempera- the
35S/3H
ratios ofproteins could resultfromDNA-bind- temperature-related changes in the uptake of
l(RSV)NRK 35Sintothecells. If this didoccur,the R values
d be due to might indicate an altered availability of
o-- 39-INCORPORATION
OF 3H-THYMIDINE
, 0 0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.53.242.56.223.2]474 MAGUN AND DORSETT
I-J It a.
FRACTION NUMBER
FIG. 3. DNA-cellulose chromatography of cytosol proteins.Chromatography of50mgofcytosolprotein wasperformed sequentiallyonnative(a) and dena-tured(b) DNA-cellulose columnsasdescribedin Ma-terials andMethods. ts339(RSV)NRK cells had been incubated in 1.0 ,Ci of[3H]leucineperml foratleast six generations to uniformly label protein. The
amount of protein in each 1-ml fraction was mea-suredby scintillation counting using 800 dpm of 3H per pg of protein as a correction factor. At each
arrow,the molarity of the elution buffer (buffer A)
TABLE 1. Fractionation of DNA-binding proteins from ts339(RSV)NRK cellsgrownat39 and330Ca
NaCl Total
pro-Temp concnof Cytosol tein
Temp elution protein boundto
(OC) buffer (%)b column
(M) (%)
Native DNA-cel-lulose
39 0.10 1.19 27.8
33 0.10 1.22 26.7
39 0.15 1.19 27.8
33 0.15 1.32 28.6
39 0.25 1.07 25.1
33 0.25 1.18 25.8
39 0.60 0.58 13.5
33 0.60 0.59 13.0
39 2.0 0.09 2.1
33 2.0 0.10 2.2
Denatured DNA-cellulose
39 0.10 2.17 47.8
33 0.10 2.23 46.0
39 0.15 0.46 10.0
33 0.15 0.54 11.9
39 0.25 0.62 13.7
33 0.25 0.66 13.6
39 0.60 0.57 12.5
33 0.60 0.56 11.6
39 2.0 0.33 7.3
33 2.0 0.38 7.8
a DNA-cellulosechromatography wasperformed
asdescribed in Materials and Methods withcytosol protein extracted from ts339(RSV)NRK cells cul-tured eithercontinuouslyat390Corshifted for 6h to 330C beforeharvesting.
IProtein contentwasmeasured by scintillation spectrometry asdescribed in Materials and Meth-ods. Fifty milligrams ofcytosol proteinwasloaded ontocolumns.
cTotal proteinwascalculatedasthe quantity of proteinrecovered in thecombined salt fractionsplus theproteinthat remained onthe column after the last salt wash (see MaterialsandMethods).
wasincreasedasshown. Allfractions between suc-cessive arrows werepooled for analysis by PAGEas
describedin MaterialsandMethods. The fractions elutedafter application ofthe 2.0 MNaCl bufferto
denatured DNA-cellulose were also analyzed by PAGE.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.59.445.57.542.2]PROTEINS ELUTED FROM
NATIVE DNA
0.10 0.15
in..
-I
0 m
son
vmwl
-!
m
_wmm_
IS-MA
0.2 5 0.
_
_...a-_._
_~
a
A B A B A B A
.60
A.
PROTEINS ELUTED FROM
DENATURED DNA
0.10 0.15 0.25 0.60
_ -_
mho
50
B A B ARB ARB A
2.0
Ili
22 A B
FIG. 4. Electropherograms of cytosolandDNA-bindingprotein fractions. ThecytosolandDNA-binding protein fractions fromts339(RSV)NRK cellseithermaintainedat390C (A)orshiftedto330C (B) for 6hwere examined byPAGE in thepresenceofSDS. Proteinfractionsfrom the experiment shown in Table1 were
concentratedand analyzed by PAGE. Samples containedfrom20to 70 pgof proteinpergel. The cytosol
fractionwasexaminedbefore (X)andafter (Y)the DNA-binding protein had been removed chromatographi-cally. (A)and(B) ofeachpair of gelscontain identicalamountsof protein.
[35S]methionine
rather than alteredsynthesis
of proteins. Since identical concentrations of
[35S]methionine existed in the culture fluid
afterthe1-hlabeling periodatboth 39and 330C
(6 h after temperature shift), it appears that
entrance of35S into ts339(RSV)NRK cells was
unaffectedby the temperature shift (Table 3).
DISCUSSION
The use of the ts339 mutant of RSV in a
mammalian cell system offers several
advan-tages over the use of the wild-type virus in
avian cells tostudy transformation (see
refer-ence 9). These are: (i) host cells show relative
genetic homogeneity, (ii)in most cases,
trans-formedcells donotproduce viral particles, and
(iii) normal(untransformed) cells donotappear
tocontainendogenous avian-sarcoma leukosis
viralgenes. Anadditionalvalue of thissystem
is the opportunity to study early molecular
changes during the onset of transformation
afteratemperature shift.
Among the
cytosol
proteins ofts339(RSV)-NRKcells, those that haveanaffinity for DNA
were found to comprise a small subset, and
mostcouldnotbe detectedamongthe profile of
total cytosol
polypeptides
on SDS gels.Chro-matographyonnative and denatured calf
thy-mus DNA-cellulose separated these proteins
intoseveral hundred polypeptides. An
accumu-lation ofbiochemical and genetic evidence has
indicated thatmanyDNA-bindingproteinscan
be associated with DNA-related functions in vivo, and that nonspecific (ionic) interaction
ofprotein withDNAis minimizedatthe NaCl
concentrations used for protein adsorption (2).
Although it remains unknown which
DNA-binding polypeptides separated in the current
investigation do in fact exhibit DNA-related
functions within thecell, the method of
analy-sisappearstobeafeasiblemeanstoselect and screen an assortmentofcandidate proteins for
their involvement in early aspects of growth
andtransformation.
CYTOSOL
_ _.
NE!
._
_
X Y
150
K
0
r-80 m 0 c
60x
-4 40
130
0a~
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.49.445.76.355.2]476 MAGUN AND DORSETT
TABLE 2. Fractionationof DNA-bindingproteins
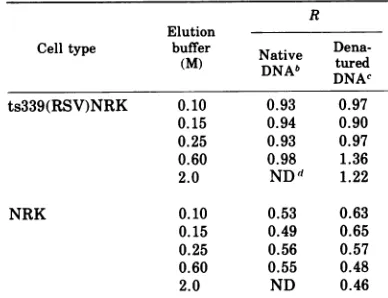
from thecytosolofts339(RSV)NRK and NRKcells atthepermissiveand nonpermissive temperature
R Elution
Cell type buffer
Native
Dena-(M)
DNAb
turedDNAc
ts339(RSV)NRK 0.10 0.93 0.97
0.15 0.94 0.90
0.25 0.93 0.97
0.60 0.98 1.36
2.0 ND 1.22
NRK 0.10 0.53 0.63
0.15 0.49 0.65
0.25 0.56 0.57
0.60 0.55 0.48
2.0 ND 0.46
a Roller bottle cell cultures [either
ts339(RSV)-NRKorNRK]weregrownat390Cfor6daysin 50ml of medium containing50
kuCi
of[3H]leucine per cul-ture, after which one-half of the cultures were shiftedto330C. Five hoursafter temperatureshift,the medium wasreducedto 5mlperbottle, and150
ACi
of [35S]methionine was added to the mediumcontaining[3H]leucine. The cultures were pulse-la-beled for1 hbefore harvesting. The other cultures were treated identically, but were maintained at
390C
throughout all labelingprocedures. Acytosol fraction was obtained from each group of cultures and was subjected to DNA-cellulose chromatogra-phy. Analiquot of each column fractionwas ana-lyzed for both3Hand35Sactivity. Each samplewas countedto a minimumof 25,000accumulatedcounts ineach channel.bR = (35S/3Hat
3300)/(35S/3H
at39CC)forcolumn fractions eluted from native DNA-cellulose. Forts339(RSV)NRK cytosol,R =0.95; for NRKcytosol,
R =0.55.
cR was determined for column fraction eluted fromdenatured DNA-cellulose.
dND, Notdone.
The results presented in this paper
demon-strate that an increased incorporation of
[35S]methionine
intoa small number ofDNA-binding
polypeptides
occurred as a result of atemperature shift from the
nonpermissive
tothe
permissive
temperature. This increased in-corporation was found to occur in proteinsbind-ing tightly to
single-stranded
DNA, but notdouble-stranded DNA, and was notdetected in
untransformed NRK cells subjected to
tempera-ture shift.
The procedure of
continuous
labeling in[3H]leucine
and short-term labeling in[35S]methionine
allows the 35S/3H ratios to beused
as a meansof comparing
the amount ofrecently synthesized
protein with the totalamount
synthesized for
individualspecies of
protein. Alterations in this ratio may occur as a result of: (i) an altered rate of de novo synthesis
(transcriptional
ortranslational
control), or (ii)a decreased rate of degradation(proteolytic
con-trol). As yet, we have not determined which of
these
mechanisms isresponsible
for thein-creased
incorporation of[35S]methionine
intosingle-stranded
DNA-binding proteinsduring
transformation.
Most of the
DNA-binding
polypeptides
inthetwo
fractions
binding tightly
tosingle-stranded
DNA
exhibited
increased 35S/3H ratios. If theincreased
incorporation
of[35S]methionine
doesinfactrepresent an increased
synthesis
ofthoseproteins, one explanation might be the
exis-tence of a coordinate control of the
synthesis
orcellular
handling of
a group of proteinswith
similar
orinterrelated
functions. Anotherex-planation
might be that some of thepolypep-tides
that
appear inthe
gels
asminor
bandsareactually degradation products of
thelarge
poly-peptide
of molecular
weight 93,000,
which
com-prises
morethan
60%of the
protein
in thatfunction.
Most
of the
single-stranded
DNA-binding
proteins
of procaryotic
cells have been
impli-cated
in some aspectof
DNAreplication
(8).
Therefore,
it is notsurprising that
anincrease
in
synthesis
of this
groupof proteins
precedes
orcoincides with the
onsetof
DNAsynthesis
inshifted
ts339(RSV)NRK cells
asdemonstrated
in
this
paper.We
arecurrently investigating
the possibility that increased
synthesis
of these
proteins may occur as a
result
of the
onsetof
DNA
synthesis
and might
therefore be
inde-pendent
of transformation
perse.Recently,
wehave
determined that the
93,000-dalton
poly-peptide
shows
anincreased
incorporation
of
[35S]methionine within h
1after the
shift
to [image:8.504.62.256.108.257.2]the permissive
temperature(in
preparation).
TABLE 3. Percentageof 35S remaining in labeling medium after1-hpulse ofts339(RSV)NRK cells
Labeling
temp0
35S
(final/initial)b(_C)
39 0.62
33 0.63
aCultures were labeled for 1 h at either 39 or
330C.
Those cultures labeled at330C
had been shifted from 39to330C 5 hbeforelabeling was be-gun.h35S was determined by withdrawing aportion ofmediumcontaining [35S]methionine for scintilla-tion counting before labeling cells (initial 35S) and afterlabeling cells for 1 heither at 39 or
330C
(final35S). Experimental protocol was identical to that described inTable 2.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.270.461.521.562.2]0.60
2.0
39
33
39 33
,-_
MW
R
-
2 50
1.91
-200 1.26
NI
7
0
1.70
r-H14
0
1.33
--115
0.81
---i05
0
93
-96
1.07
-91
1.02
77
0.94
e.
*L_68
1.31
_-63
0.77
-6
0
1.14
a
...N
--53
0.80
m
-47
1.06
-
42
1.33
1.03
27
1.19
-23
1.10
'".
MW
R
-200
1.22
-150
1078
1-93
1.38
-72
-66
1.33
1.14
-59
0.83
-50
-45
43
41
W-35
0.85
1?57
1.08
1.1
9
1.
10
1.13
-26
0.74
--22
1.10
FIG. 5. PAGEelectropherograms of ts339(RSV)NRK DNA-bindingproteinsobtainedintheexperiment describedinTable 2. Theprofiles ofthe DNA-bindingproteinselatedfromdenaturedDNAwith0.60 MNaCl and 2.0 M NaClareshown, along with the R values andmolecular weights (x1O-30 MW) for the major correspondingpeaks. The molecular weights were estimatedfrom companiongels containing proteins of knownmolecular weights (see MaterialsandMethods).
Thisindicates that synthesis ofthispolypeptide
precedes DNA synthesis by several hours and
that it isoneof the earliestevents to be
corre-lated withtheonsetoftransformation.
Although we have used theterm
"cytoplas-micDNA-binding protein" throughout this
in-vestigation, it should be pointedout thatmany
of theproteins separated in this investigation
may be compartmentalized to the nucleus in
vivo. Hypotonic swelling of the cells, followed
__
ao
*1.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.504.83.414.80.538.2]R value
FIG. 6. Histogram ofRvaluesfor159 correspond-ingDNA-binding polypeptidesobtainedfromthe
ex-perimentdescribed in Table2withts339(RSV)NRK cells. AllDNA-bindingprotein fractions were
ana-lyzed byPAGE, afterwhichgels were sliced
trans-verselyfor scintillationspectrometry. R valueswere
obtainedforcorresponding peaks from shifted and nonshiftedcultures. Arrow denotesmean.
by Dounce
homogenization,
could beresponsi-ble
for the release
ofsoluble proteins fromnu-clear to
cytoplasmic
compartments.However,
Choe and
Rose
(7) found thatatleasthalf of thecytoplasmic
DNA-binding
proteins of WI-38cellsweretransported into nuclei within1
h,
inthe
absence
of DNAsynthesis. These
resultswould
indicate
that thecytoplasmic
DNA-bind-ing
proteins
maymigrate
tothe nucleus
andperform
DNA-related functions there. How-ever, itremainsto bedetermined whether theproteins
whose synthesis
isaltered
after theonsetoftransformation inthe present
investi-gation
do in fact haveDNA-related roles.
ACKNOWLEDGMENTS
WethankFrances Byrd for excellent technical
assist-ance. This workwas supportedby Public Health Service
grantCA-17183 from the National Cancer Institute. LITERATURE CITED
1. Alberts, B. M.1970.Function ofgene32protein,anew protein essential for the genetic recombination and replication of T4 bacteriophage DNA. Fed. Proc. 29:1154.
2. Alberts, B.,and G.Herrick.1971.DNA-cellulose chro-matographyp.198-217. InS. P.Colowick and N.0.
Kaplan (ed.), Methodsinenzymology, vol. 21,partD. Academic Press Inc., New York.
3. Anderson, W. B., G. Johnson, and I. Pastan. 1973. Transformation of chick embryo fibroblasts by wild-typeand temperature-sensitive Roussarcomavirus alters adenylate cyclase activity. Proc. Natl. Acad. Sci. U.S. A. 70:1055-1059.
4. Arndt-Jovin, D., T. M. Jovin, W. Bahr, A.-M. Fris-chauf,andM.Marquardt.1975.Covalent attachment of DNA toagarose.Eur.J.Biochem. 54:411418. 5. Bayliss, G. J., H. S. Marsden, and J. Hay.1975.Herpes
simplex virus proteins:DNA-binding proteinsin in-fected cells and in the virus structure. Virology 68:124-134.
6. Carroll,R.B.,L.Hager,and R. Dulbecco. 1974. Sim-ian virus 40 TAntigenbindsto DNA. Proc. Natl.
Acad. Sci. U. S. A. 71:3754-3757.
7. Choe, B.-K.,and N. R. Rose. 1974. Synthesisof
DNA-bindingproteinsduringthe cellcycleofWI-38 cells.
Exp.Cell Res.83:271-280.
8. Geider,K. 1976.Molecular aspects of DNAreplication
inEscherichia coli systems. Curr.TopMicrobiol. Im-munol. 74:55-112.
9. Graf, T.,andR. R. Friis. 1973.Differentialexpression
oftransformation inrat andchicken cells infected with an avian sarcoma virus ts mutant. Virology
56:369-374.
10. Huang, W.-M.,and J. M. Buchanan. 1974.Synergistic
interactions of T4early proteinsconcerned with their
binding to DNA. Proc. Natl. Acad. Sci. U. S. A. 71:2226-2230.
11. Hynes, R. L., and J. A. Wyke. 1975. Alterations in surfaceproteinsinchicken cellstransformedby tem-perature-sensitive mutantsofRoussarcomavirus. Virology64:492-504.
12. Jessel,D.,J.Hudson,T.Landau, D. Tenen, and D. M. Livingston. 1975. Interaction of partially purified simianvirus 40 Tantigenwithcircular viral DNA molecules. Proc. Natl. Acad. Sci. U. S. A. 72:1960-1964.
13. Kawai,S.,and H. Hanafusa. 1971. The effects of
recip-rocal changes in temperature on the transformed state of cells infected with a Rous sarcoma virus
mutant.Virology46:470479.
14. Kletzien,R.F.,and J. F.Perdue. 1974.Sugartransport inchickembryo fibroblasts.II.Alterations in
trans-portfollowingtransformationbya temperature-sen-sitive mutant of the Rous sarcoma virus. J. Biol. Chem.249:3375-3382.
15. Laemmli, U. K. 1970. Cleavageofstructuralproteins duringtheassembly of the head of bacteriophage T4. Nature(London)227:680-685.
16. Litman, R. M. 1968. Adeoxyribonucleicacid
polymer-asefromMicrococcus luteus (Micrococcus lysodeikti-cus) isolated on deoxyribonucleic acid-cellulose. J. Biol.Chem. 243:6222-6233.
17. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Proteinmeasurement with the Folin phenolreagent. J.Biol. Chem. 193:265-275. 18. Magun,B. E.,R. R.Burgess, and H. P. Rusch. 1975.
Nuclearphosphoproteins ofPhysarumpolycephalum. Arch. Biochem.Biophys.170:49-60.
19. Matthews, C. K. 1971. Bacteriophage biochemistry.
Am.Chem. Soc. Monogr no. 166. VanNostrand and Reinhold,New York.
20. Otten, J.,J. P.Bader,G. S. Johnson, and I. Pastan. 1972.A mutation in a Roussarcoma virus gene that controls adenosine 3',5'-monophosphate levels and transformation. J. Biol. Chem. 247:1632-1633. 21. Purifoy, D. J. M., and K. L. Powell. 1976.DNA-binding
proteins induced by herpes simplex virus type 2 in HEp-2 cells. J. Virol. 19:717-731.
22. Reed, S., J.Ferguson, R. Davis, and G. Stark. 1975. T Antigen binds to simian virus 40 DNA at the origin of replication. Proc. Natl. Acad. Sci. U. S. A. 72:1605-1609.
23. Schaffer,P. A. 1975.Temperature-sensitivemutants of herpes viruses. Curr. Top. Microbiol. Immunol. 70:52-100.
24. Schaller, H., C. Nusslein, F. J. Bonhoeffer, C.Kurz, and I.Neitzschmann. 1972.Affinitychromatography ofDNA-binding enzymes on single-stranded DNA-agarosecolumns. Eur. J. Biochem. 26:474-481. 25. Shanmugam, G., S. Bhaduri, M. Areaus, and M.
Green. 1975.DNA-binding proteins in thecytoplasm and in a nuclearmembrane complex isolated from uninfected and adenovirus 2 infected cells. Biochem-istry 14:332-337.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.504.58.257.59.212.2]26. Sigal, N., H. Delius, T. Kornberg, M. L. Gefter, and B. M. Alberts. 1972. ADNA-unwindingproteinisolated fromEscherichia coli: its interaction with DNA and with DNA polymerases. Proc. Natl.Acad. Sci. U. S. A. 69:3537-3541.
27. Stone, K. R., R. E. Smith, and W. K. Joklik. 1974.
Changes in membrane polypeptides thatoccurwhen chick embryo fibroblasts and NRK cellsare
trans-formed with aviansarcoma viruses.Virology
58:86-100.
28. Subak-Sharpe, J. H., S. M. Brown, D. A. Richie, M. C. Timbury, J. C. M. McNab, H. S. Marsden, and J. Hay. 1974. Cold Spring Harbor Symp. Quant. Biol. 39:717-730.
29. Tenen, D. G., P. Baygell, and D. Livingston. 1975.
Thermolabile T (tumor) antigen from cells trans-formed byatemperature-sensitive mutant of simian
virus 40.Proc. Natl. Acad. Sci. U. S. A. 72:4351-4355. 30. Van der Vliet, P. C., and A. J. Levin. 1973. DNA-binding proteins specific for cells infected by
adenovi-rus.Nature (London) 246:170-174.
31. Van der Vliet, P.C., A. J. Levine, M. J. Ensinger, and H. S. Ginsberg. 1975. Thermolabile DNA binding
pro-teinsfrom cells infected withatemperature-sensitive
mutantof adenovirus defective in viral DNA synthe-sis. J. Virol. 15:348-354.
32. Zimmerman,S. B., and C. J. Levin. 1975. A deoxyribo-nucleic acid ligase from nuclei ofratliver. J. Biol. Chem. 250:149-155.