JOURNAL OFVIROLOGY,Nov. 1993, P. 6797-6807 0022-538X/93/116797-11$02.00/0
Copyright © 1993, American Society for Microbiology
Mutagenesis of the
Yellow
Fever
Virus
NS2B Protein:
Effects
on
Proteolytic Processing,
NS2B-NS3 Complex
Formation,
and
Viral Replication
THOMAS J.CHAMBERS, ANN NESTOROWICZ, SEAN M. AMBERG, AND CHARLES M. RICE*
Department of Molecular Microbiology, Washington UniversitySchool of Medicine,
Box
8230,
660S.Euclid
Avenue,
St.Louis,
Missouri63110-1093Received4June 1993/Accepted 16 August 1993
To study the role of specific regions ofthe yellow fever virus NS2B protein in proteolyticprocessing and
associationwith the NS3 proteinasedomain,aseries of mutationswerecreated in the hydrophobic regionsand
ina centralconserved hydrophilic region proposed as a domain important for NS2B function.The effects of
these mutationsoncis cleavageatthe2B/3cleavage siteand onprocessingatotherconsensuscleavage sites
for the NS3 proteinase in the nonstructural region were then characterized by cell-free translation and
transient expression inBHK cells. Association between NS2Band the NS3 proteinase domainand theeffects
ofmutationsoncomplex formationwereinvestigated by nondenaturing immunoprecipitationof these proteins
expressed in infected cells, by cell-freetranslation,orby recombinant vaccinia viruses. Mutations within the
hydrophobic regions had subtle effects on proteolytic processing, whereas mutations within the conserved
domaindramatically reduced cleavage efficiencyorabolished all cleavages. The conserved domainofNS2Bis
alsoimplicated in formation ofan NS2B-NS3 complexon the basis of the ability of mutationsin this region
toeliminateboth association of thesetwo proteins and trans-cleavage activity. In addition, mutations which eithereliminatedproteolytic processingorhadnoapparenteffectonprocessingwerefoundtoabolishrecovery
ofinfectious virus following RNA transfection. These results suggestthat the conserved region of NS2B is a
domain essential for the function of the NS3 proteinase. Hydrophobic regions of NS2B whose structural integritymaynotbe essential for proteolytic processingmayhaveadditional functions during viral replication.
Yellow fevervirus (YF), theprototype member of the Flavi-virus genus, contains a single positive-stranded RNA genome
which encodes a single long open reading frame believed to
generate a polyprotein ofover 350 kDa (7, 29, 30). The gene
order for the polyprotein has been established as
5'-C-prM-E-NS1-NS2A-NS2B-NS3-NS4A-NS4B-NS5-3', where C, prM, and E denote structural protein precursors and NS1 through NS5
indicate the nonstructuralproteins. Recent investigations have established that several cleavages onthe flavivirus polyprotein
are mediated by a viral proteinase encoded within the N terminus of theNS3 protein (6, 13, 27, 33, 35). ForYF, these cleavage sites include the consensus sites
[G(A)RRIS]
which generate the N termini of NS2B, NS3, NS4A, and NS5 (see reference22, and foracompilation, seereference7),aswellasadditional cleavage sites within the NS2A and NS4A regions which have been proposedtooccur atQK{Tand
QRIS
sites, respectively (21, 26). In addition, the proteinase isbelieved tomediate a cleavage producing the C terminus of the virion capsid at a proposed consensus site (1). The flavivirus NS3
proteinase containsaserineproteinase domain with homology
tothesmallcellularproteinasesof thetrypsinsuperfamily (3, 4, 18). The importanceof residues in theproposedserine
protein-asecatalytictriad(YF,His-Asp-Ser) has been demonstratedby several studies employing site-directed mutagenesis of these
residues (2, 11, 33, 35). It has also been established that in
additiontoanintactproteinase domain,thesmall nonstructural
protein NS2Bisrequiredforsite-specific cleavage activityat the
consensusand additional sites(2, 6, 13, 21, 27, 35). Althoughthe
precise function of YF NS2B in processing of the flavivirus
polyprotein isnotknown,several linesof evidencesuggestthat
*Correspondingauthor.
this proteinmay associatewith NS3 such that the active form of the proteinase is aheterodimer. This evidence includes (i) the observation that cleavage of 2A,1 -3, polyproteins to generateNS2B and
3,,,
invitro isdilution insensitive(l1); (ii) the observation thatNS2B and theNS3proteinase domainareboth necessary fortrans cleavage at the 2A/2B, 2B/3, 4A/2K, 4B/5, and anchored capsid cleavage sites (1, 6, 21); and (iii) the
recent demonstration that the dengue virus type 2 (DEN2) NS2B and NS3 proteins can be coimmunoprecipitated (2). Without crystallographic data on the structure of the active NS2B-NS3 proteinase, the nature of the interaction between these two proteins remains unknown. However, sequence
comparison offlavivirus NS2B proteins reveals a highly
con-servedandchargeddomain flankedby regionsofhydrophobic amino acids (12). The highly conserved domain has been showntobeessential for NS3-mediatedcleavageatthedengue virus type 4 (DEN4) 2B/3 cleavage site (12). Because the predicted topology of the YF polyprotein suggests that
pro-cessing occurs in association with the membranes of the
endoplasmic reticulum,ithas beenhypothesizedthatthe small
hydrophobic proteins NS2A, NS2B, NS4A, and NS4B maybe
required for association of the large nonstructural proteins with membrane sites (30). Although most of the flavivirus
nonstructural proteins have been identified in membrane
fractions of infected cells (34), specific regions of these
pro-teins which participate in membrane-protein and protein-protein interactions have not been determined. In thisstudy, severalmutationswerecreated withinputativecytoplasmicand
membrane-associated domains of the YF NS2B protein, and
their effects on polyprotein processing, association between
NS2B andNS3, cleavage activityoftheproteinaseintrans,and
recovery of infectious virusweredetermined. 6797
Vol.67, No. I1
on November 9, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Cells andviruses. Infectionwith YF 17Dwas as described
previously (8). Growth of BHK-21, SW-13, BSC-40, and
BHK-21-15 cells has been described previously (6, 9). Prepa-ration of vTF7-3, the vaccinia virus expressing T7 RNA
polymerase (15),has been describedpreviously (6).CV-1 cells
were grown in Earle's minimal essential medium (MEM) containing5% fetal calfserum.
T7transcription plasmids.The useofpET plasmids (F.W.
Studier, Brookhaven National Laboratories) (32) and pTM3 plasmids (B. Moss,National InstituteofAllergyand Infectious
Diseases) (25)forcloningandexpressionof YFproteinsunder the control of the T7 promoterhas beenpreviously described
(6, 11).Plasmidswereconstructedwith standard methods and reagents (31), and the structures were verified by restriction
enzyme digests and nucleotide sequence analysis for all
con-structsengineered by oligonucleotide-directed mutagenesisor polymerase chain reactionamplification.
Construction of mutants and site-directed mutagenesis.
Mutations within theYFNS2Bproteinwerecreatedbytheuse
of natural restriction sites or by using restriction sites engi-neered by oligonucleotide-directed mutagenesis. CD38 was
constructed by creating two plasmids:
pET8c-NS2A,
10-3298D NcoI, containing a deletion of the coding sequence between the NcoI sites at 3834 and 4457 (YF nucleotide numbering),and
pET8c-NS2A,
10-3298D HindlIl, containing a deletion ofthe coding sequence between the HindlIl sites at 4343 (YF nucleotide) and 1883 (plasmid sequence). Following digestion with NcoI and HindIII, respectively, the linearized plasmids
were filled in with the DNA polymerase I large fragment (Klenow) (NcoI site) or digested with mung bean nuclease
(Hindlll site), and appropriate fragments were isolated and
ligated byblunt-end ligation. The NS2B fragment containing
the mutationwas subcloned into theplasmid pTM3-NS2B by
polymerase chain reaction amplification or introduced into
sig2A-5356 by exchange ofthe770-bp SstI fragment. Mutants CD3 and CD5were constructed by creation of intermediate
pET8c-2A,
10-3181
plasmids containing deletions between the SstI sites at4338 and 5112(YF nucleotides) and betweenthe HindlIl sites at 4343 (YF nucleotides) and 1833 (plasmid sequence), respectively. Fragments containingfilled-inSstI and HindIll sites were ligated with a three-piece ligation with avector-derived EcoRI-AvaI fragment. A few resulting clones unexpectedlycontainedanNheI siteatNS2B nucleotide 4330.
This sitewasutilized tocreatemutation CD5 by linearization
withNheI anddigestion with mungbean nuclease and
religa-tion and also to create mutation CD3 by ligating fragments containingfilled-in NheI and HindIlI sites. Forcreation of 1D3
and1I3,twopET/BS(+)2B-3181 plasmids (11) containing StuI
sites at NS2B positions 4225 (1.1) and 4234 (1.2)were
engi-neered by site-directed mutagenesis. Constructs containing
eithera3-amino-acid insertion(113)or adeletion ofthesame
size(1D3) weregenerated by ligation of StuI-AvaI fragments
containingthe 5'StuI site of 1.2 andthe 3' StuI siteof 1.1or
the 5'StuI site of 1.1 andthe 3' StuIsiteof 1.2, respectively. Forcreation of 2D4, 214, and 2D13, twopET/BS(+) 2B-3181
plasmids containinganEcoRV siteat4273(2.1) andaStuI site at4285 (2.2)wereengineeredby site-directed mutagenesis. A
construct containing a 4-amino-acid insertion (214)was then
generated by ligation ofanAvaI-StuI fragmentcontaining the 3' StuIsite of2.2 withanAvaI-EcoRVfragmentcontaining the 5' EcoRV site of2.1. Aconstruct containing a 4-amino-acid
deletion (2D4) wascreated by ligating anAvaI-EcoRV frag-ment containing the 3' EcoRV site of 2.1 with anAvaI-StuI
fragment containing the 5' StuI site of 2.2. Construction of
2D13was
accomplished by
ligation
of theAvaI-StuIfragments
containing
the 3' StuIsiteof 1.2 and the 5' EcoRV site of 2.1.For creation of mutations 3D3 and
313,
twopET/BS(+)
2B-3181
plasmids
containing
Stul sitesat NS2Bpositions
4475(3.1)and 4485
(3.2)
wereengineered
by
site-directedmutagen-esis. Constructs
containing
either a 3-amino-acid insertion(313)oradeletion of thesamesize
(3D3)
werethengenerated
by
ligation
of StuI-AvaIfragments
containing
the 5'StuIsite of3.1 and the 3' Stulsite of 3.2orthe3'StuIsite of 3.1 and the 5' Stul site of
3.2,
respectively.
For creation of the CD4mutation,
twopET/BS(+)
2B-3181
plasmids
containing
anEcoRVsiteatNS2B 4376
(C4.1)
andaScaIsiteatNS2B 4388(C4.2)
wereengineered
by
site-directedmutagenesis.
Acon-struct
containing
a4-amino-acid deletionwas thengenerated
by
ligating
anAvaI-EcoRVfragment
with anAvaI-ScaI
frag-ment.Forcreation ofmutations
1D3D,
1D31,
113D,
and1131,
the
770-bp
SstIfragments
of2B-3181 constructscontaining
the3D3 and 313 mutations were each
ligated
into each of the2B-3181
plasmid
vectorscontaining
the 1D3 and 113mutations afterdigestion
withSstIanddephosphorylation.
Forconstruc-tionof
sig2A-5356
(6) containing
theCD38, CD4,
3D3,
and 313mutations,
the770-bp
SstIfragments
from themutant2B-31,81
constructs was
ligated
intosig2A-5356
afterdigestion
withSstI
and
dephosphorylation.
For construction ofsig2A-5356
plas-mids
containing
the1D3, 1I3,
2D4, 2D13,
213,
CD3,
andCD5mutations,
sig2A-3,81
constructscontaining
these mutationswere
generated
by
subcloning
PvuIIfragments
from themu-tant
2B-3181
constructs intosig2A-3181.
The mutations werethen reconstructed in
sig2A-5356 by
ligation
ofXbaI-HpaI
fragmentsfrom the
sig2A-3181
constructsintosig2A-5356
afterdigestion
withXbaI andHpaI
anddephosphorylation.
Con-struction of
sig2A-5356 plasmids
containing
the 1D3D and1131mutationswasachieved
by
ligation
of the SstIfragments
fromsig2A-5356
constructscontaining
the 3D3 and 313 mutationsinto
sig2A-5356
plasmids
containing
the 1D3 and 113muta-tions,
respectively,
after these weredigested
with SstI anddephosphorylated.
Vacciniavirus recombinants. Plasmids for
homologous
re-combinationwere
generated
by cloning
the2B,3181
or2B-3181
regionsintopTM3. ForpTM3-NS2B, pET8c-NS2B DNAwas
digested
with BamHI andpartially
withNcol, and the403-bp
fragment
wasligated
intopTM3which had beendigestedwithBamHIand NcoI. Constructionof
pTM3-3181
wasachievedbydigestion
ofpET8c-3181
DNA with MspI andfilling
in theoverhangingends withDNApolymerase
I,
followedbyfurtherdigestion
with NcoI. The622-bp fragment
wasthen ligatedtopTM3DNAafterdigestionwithSmaIandNcoI. Construction
of
pTM3-NS2B-3181
was accomplished by digestingpET8c-NS2B-318,
with NheI and partially with NcoI. The 1,236-bpfragment
wasligated
intopTM3 afterdigestionwith NcoI andSpeI.
pTM3-NS2B clones containing mutations CD3, CD5,andCD38were generatedfrom pET8c-NS2Bconstructs
con-taining
these mutations by the same strategy described forpTM3-NS2B. Vacciniavirus recombinantswere generated by
infecting
CV-1 monolayerswithwild-type (wt) vacciniavirusand transfecting with the calcium phosphate-precipitated pTM3-YFconstructs(24). Recombinant viruswasselectedby
passagingon humanTK- 143 monolayers in the presence of 25 ,ugof5-bromodeoxyuridineperml.Theresulting thymidine
kinase-negative
vacciniaviruswasplaque purifiedatleast twiceunder gptselection
(14)
onBSC-40 cells. Large-scale prepara-tions of recombinant virusweregrown andtiteredonBSC-40or BHK-21 cells in the absence of selection.
Invitrotranscription.5' capped transcriptswere produced
from linearized pET8c-YF plasmid DNA templates with T7
on November 9, 2019 by guest
http://jvi.asm.org/
YELLOW FEVER VIRUS PROTEINASE COMPLEX 6799
RNApolymerase(Epicenter) and conditions recommended by the manufacturer.
Cell-free translation. Cell-free translation was performed with rabbit reticulocyte lysates programmed with pET8c
tran-scriptsin accordancewith themanufacturer's (Promega)
spec-ifications. Proteinswere radiolabelled byinclusion of
[-3S]me-thionine (Amersham).
Transient expression assay. Expression of YFpolyproteins
with the transient expression assay has been described previ-ously (6). Briefly, confluent monolayers of BHK-21-15 cells
were infected with vTF7-3 (multiplicity of infection = 10); infection was followed by DNA transfection with lipofectin (Bethesda Research Laboratories) (15
pLg
of lipid per ,ug of DNA). Following an interval of2.5 h, cellswere labelled with75S]methionine as previously described (6).
Preparationofinfected cell extracts. ExtractsofYF-infected BHK-21 cells for preparation of YF-specific proteins were
prepared as previously described (9).
Immunoprecipitation. Lysates from transientexpression
ex-periments or proteins prepared by cell-free translation were
prepared with denaturing conditions as described previously
(9, 11). Lysates for analysis of proteinase complex formation
were prepared with nondenaturing conditions (1% Nonidet P-40, 0.1% sodium dodecyl sulfate [SDS], 1.0% deoxycholate,
150 mM NaCl, 50 mMTris-Cl [pH 7.5] [buffer A]). Following
solubilization of cell monolayers in thisdetergent mixture,the lysate wascentrifuged at30,000rpm in anSW50.1 rotorfor 30 min at 4°C. The supernatant was removed, and immunopre-cipitationwascarriedoutbyaddition of antiserum toNS2Bor
NS3. Immunoprecipitates were collected with Staphylococcus alureius Cowan strain I (Calbiochem) and then washed three times with buffer A and once with I M NaCl-50 mM Tris-Cl
(pH 7.5).
Gel electrophoresis. Washed immunoprecipitates were sol-ubilized in SDS sample buffer and
analyzed by
SDS-polyacryl-amide gel electrophoresis (PAGE)
by
methodspreviously
described (20).
Construction offull-length YF cDNA templates and RNA
transcription and transfection.
pET-8c-sig2A-535(,
plasmids
containing NS2B mutations were used to reconstruct
full-length YF cDNA templates
by
methodspreviously
described (28) with slight modifications. TheAvrII-Nhelfragment
fromsig2A-535,6
clonescontaining
the relevantNS2B mutationswas transferred to the YFM 5.2plasmid,
and the AatII-NsiIfragments of the YFM 5.2 clones andYF 5'3'IVwere
ligated
after recovery ofappropriate
fragments
fromlow-melting-temperature agarose
gels.
By thismethod,
approximately
100 ng of full-length template wasgenerated
on the basis ofanalysis by agarose
gel
electrophoresis.
Full-length
templates
were linearized with Xhol, and SP6
transcripts
were synthe-sized aspreviously
described(28).
Integrity
of YFtranscripts
was determined by
nondenaturing
RNAgel
electrophoresis.
Transfection of
subconfluent
SW-13 cells was carried outwithconditionsdescribed for BHK-21 cells
(28). Briefly,
mono-layerswere washed with
phosphate-buffered
saline(PBS)
andtransfected with a mixture of
PBS,
lipofectin (Bethesda
Re-search Laboratories),and RNAtranscripts
for 10 minat roomtemperature.Thetransfection mixture wasthen removed, cells
werewashed with MEM, and thenthe
monolayer
wasoverlaidwith 1%
agarose-MEM
containing
2% fetalcalf serum-25 mMHEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid),
pH 7.0. Plaques were visualized after 96 h
by staining
withneutral red or
crystal
violet. For recovery of virus forplaque
assay, monolayers transfected with 250 ng oftranscripts
wereincubatedin MEM
containing
2%fetal calfserumfor 60 to72h, following which the medium was removed, clarified
by
low-speed
centrifugation,
andstoredat -70°C
until theplaque
assay was
performed.
Virus was titeredby plaque
assay on SW-13cellswithoverlay
conditionsasdescribed above.RESULTS
Structures ofNS2B mutations.
Figure
I illustrates the YF NS2Bprotein
with the conserved central 40-amino-acid do-mainflankedby
threeregions
ofhigh
hydrophobicity.
Since the2A/2B
and2B/3
cleavages
arepresumed
to occur in thecytoplasm,
thepredicted topology
of thepolyprotein
suggests
that these
hydrophobic regions
may be membrane-associated segments and the conservedregion
may be acytoplasmic
domain,
although
this has notbeen proven. The threehydro-phobic regions
and the conserved domain weretargeted by
mutations
consisting
of in-frame deletions orinsertions,
insome cases
accompanied by
conservative or nonconservativeamino acid substitutions. Mutation ID3 contains a
3-amino-acid deletion of residues 18 to 20
(relative
to the NS2B Nterminus)
associated withthesubstitution of G forVatresidue16. The
113
mutation contains a repeat of residues 18 to 20(AGL)
inserted between residues 17 and 18. The 2D4muta-tion contains a 4-amino-acid deletion of residues 33 to 36
(AVGG).
The214
mutation consists ofa 4-amino-acidinser-tion
(VGGS)
between residues33and34,
associated withthesubstitution of S for A at residue 33. The 2D13 deletion removes residues from
positions
21 to33,
associated with thesubstitution of S forLatresidue21. Mutations CD3 and CD5
in the conserved domain consist of 3- and 5-amino-acid deletions
(LEL
andLELKK)
from residues53to55 and 51 to55,
respectively.
CD38consists ofadeletion of 38 amino acidsfrom residues56to
93,
associated with the substitution ofTforP at residue 93. CD4 consists of a 4-amino-acid deletion
(ISGS)
fromresidues67to70,
associated with theconservative substitutions ofDforEatresidue66andTfor Satresidue 71. The 3D3 mutation consists of a deletion of three residues(TSL)
from residues 100 to102,
associated with thenoncon-servative substitution ofKforMatresidue99.313containsan
insertion of 3 amino acids
(SQL)
between residues 100 and101. The 1D3D and
1131
double mutations consist of combi-nations of 1D3 with 3D3 and113
with313,
respectively.
Cell-free translationof
2B-3181
transcripts containing
NS2Bmutations.
Figure
2illustrates thecleavage
patterns
of2B-3,8,
polyproteins
containing
the mutationsinthehydrophobic
and conserved domains of NS2B.Following
1 hoftranslation,
thewt
2B-3181
polyprotein
is cleaved almostcompletely
to the21-kDa
318,
and 14-kDa NS2Bproteins (lane
b).
2B-318,
polyproteins
containing
mutations in any of the threehydro-phobic
regions
exhibited levels ofcleavage
similar to that ofthewt
(lanes
ctogand1toq), although
precise
quantitation
of thesecleavage
reactionswas notperformed.
In theseexperi-ments,
cleavage
efficiency
is based on the amount of the 21-kDa3181
protein
formed,
because of differences amongmutantforms of NS2B
(see
below).
ThemutantNS2Bproteins
exhibitedelectrophoretic
mobilities consistent with their pre-dicted molecularweights (Fig. 1).
Mutant forms of NS2Bcontaining
deletions withinthe firsthydrophobic region
(1D3,
2D13, 1D3D,
andID31,
Fig.
2,
lanes c, e, n, ando)
werepresent
in diminished amountscompared
with the wt form.Such deletions mayrendertheNS2B
protein
moresusceptible
to
degradation
in thereticulocyte lysate.
However, thesemutations may alter the
reactivity
ofNS2B
with the antisera usedforimmunoprecipitation.
In the absence ofimmunopre-cipitation,
comigration
ofNS2B withendogenously
produced
globin
obscures anyability
to differentiate thesepossibilities
(data
notshown).
Incontrasttomutationsinthehydrophobic
V()L.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
5'NC
5' ORF
Cap-STRUcTURALI
NONSTRUCTURAL,**
v
tUlJ
il
4
J
1
I2AWB
3
A
4B
5
sig2A-5356
LIi
1111
NS2B
WT
1D3
113
2D4
2I4
2D13
CD3
CD5
CD38
CD4
(13.9)
(13.7)
(14.2)
(13.7)
(14.3)
(12.5)
(13.6)
(13.4)
(9.9)
(13.6)
3D3 (13.7)
313
(14.3)
1D3D
(13.4)
1131 (14.5)
CONSERVED
.
I
I
o1
a
FIG. 1. Schematic of theYFgenome,nonstructuralpolyprotein,thesig2A-5356construct, andthe NS2Bproteinwith the mutationsengineered within this protein.Thesig2A-5356construct(6)contains thesignalsequence(solid region) precedingNS1 fusedtoNS2Aandextendsthrough
the first 356 amino acids ofNS5.Asterisks indicatepotentialN-linkedglycosylationsites. The shadedregionindicates theNS3 proteinasedomain.
Putativesignalase cleavagesarerepresentedbysolid diamonds.OpenarrowsindicatecleavagesmediatedbyNS3 proteinase.Theopeninverted
triangle indicates NS1-2Acleavage byanundefined proteinase.Within the NS2Bprotein, the central conserved domain is indicated. Vertical
striped regionsindicate thepositionsof thehydrophobic regions.Mutationsareindicatedbyopenboxes(deletions)orsolidboxes(insertions).
Thepredictedmolecularweights (inthousands) of NS2Bproteinscontainingthese mutationsareindicated inparentheses. ORF,openreading frame.
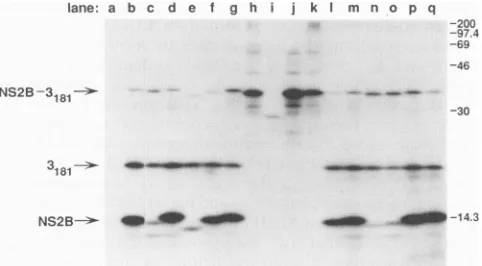
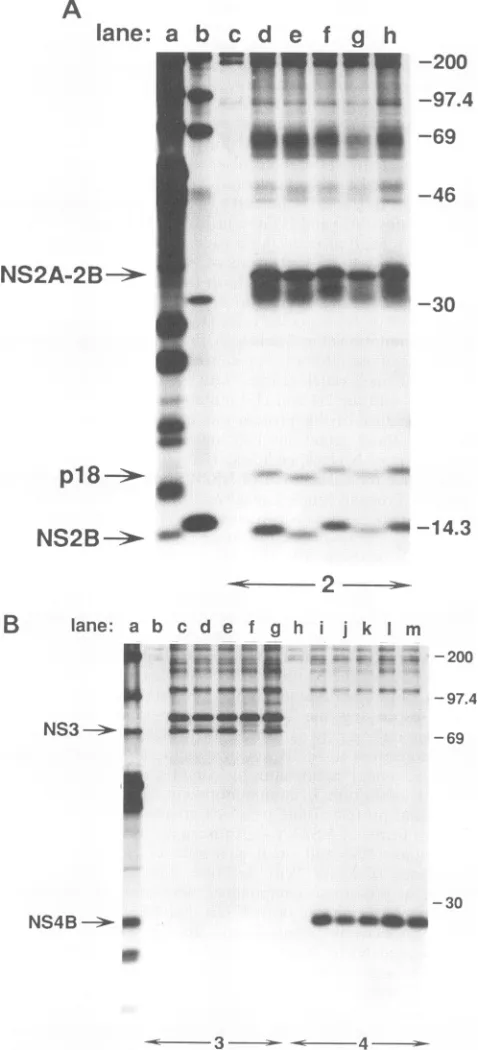
domains, 2B-3181 polyproteins containing deletions in the
conserved domain (CD3, CD38, CD4, andCD5, lanes htok)
exhibited nocleavage productsconsistent in sizewith 3181 or
NS2B. AminoramountofNS2B and3181wasdetectablefrom
the CD4 mutant afterprolonged exposure (data notshown).
Therelativeamountofthe 2B-3181 polyproteincontaining the CD38 deletionwasdiminished.Although this deletionreduces
immunoreactivity withNS2B antisera (datanotshown),
reac-tivity with antisera to NS3 should not be altered in the
presenceofSDS. Thissuggeststhat theCD38deletionrenders the 2B-3181 polyprotein unstable in this expression system.
Taken together, these results suggest that the conserved do-mainwithinNS2B isnecessaryforsite-specific cleavageatthe
2B/3cleavage site.
Effect ofmutations in NS2B on processing at the 2A/2B, 3/4A, 4A/4B, and 4B/5 cleavage sites. In order to determine whether mutations within NS2B which permit or abolish
cleavageat the 2B1/3cleavage site incell-freetranslation alter
theNS3proteinase-mediatedcleavagesatadditional
nonstruc-tural cleavage sites, processing of
sig2A-5356
polyproteins containing selected NS2B mutations wasanalyzed with tran-sient expression in BHK-21-15 cells (6). Figure 3A illustrates the patternsofcleavagesfrommutants1D3, 113, 3D3,and 313analyzed by antisera to NS2B. Compared with processing of
thewt polyprotein (lane d), all ofthese mutantpolyproteins
exhibited essentiallysimilarpatternsofprocessing(laneseto
h),althoughtheapparentmolecularweightsof the2B-specific proteins are altered because of the presence ofdeletions or
3'NC 3'
H2
]
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.118.475.73.489.2]YELLOW FEVER VIRUS PROTEINASE COMPLEX 6801
lane: a b c d e f g h i j k m n o p q
NS2B-3181-*
-__
3
181 - -
-,=4_4 1.
FIG. 2. Cell-free cleavage activity of 2B-3181 polyproteins
contain-ing mutations in NS2B. T7 RNA transcripts were used to program
rabbit reticulocyte lysate,and incubationswerecarriedoutfor 60min
at
30°C.
Following RNasetreatment, samplesweretreated with SDS(0.5%), heated to70°Cfor 5 min,and then diluted and
immunopre-cipitated as described in Materials and Methods with a mixture of
antisera specific for the YF NS2B and NS3proteins.
Immunoprecipi-tated proteins were analyzed by electrophoresis on SDS-15%
poly-acrylamide gels, and proteins were visualized by fluorography with sodium salicylate. Lanes:a, noRNA; b,wt;c,1D3; d,113; e,2D13;f,
2D4;g,214;h, CD3; i, CD38; j, CD4; k,CD5;1,3D3;m,3I3;n, 1D3D;
o, 1D31; p, 113D; q, 1131. Molecular weights, in thousands, are
indicated atright.
insertions.Figure 3B illustrates the processing of thesemutant
polyproteins as analyzed with antiserato the NS3 and NS4B proteins. The
sig2A-5356
polyproteins containing NS2Bmuta-tions 1D3, 113, and313 (lanes d andj,eand k, andg andm)
exhibited patternsof NS3- and NS4B-specific cleavage
prod-ucts similartothose generated bythewtpolyprotein (lanesc
and i). The NS3-specific proteins include the 75-kDa NS3 protein andamoreslowlymigrating form (80kDa) consistent
withan uncleaved NS3-4Aprotein. The NS3-4A species has
beenvariably observed in this assay and in YF-infected cells
and is believed to result from inefficient cleavage atthe 3/4A cleavage site (6).In contrast, the 3D3mutation (lanesf and1) generated a reduced amount of the mature NS3 protein (75 kDa) relative to the wt, suggesting that this deletion may
further impair cleavage efficiency at the 3/4Acleavage site. Figure 4 illustrates the effects ofconserved domain
muta-tions CD3, CD4, CD5, and CD38 on processing of the
sig2A-5356
polyprotein, as analyzed with antisera to NS2B,NS3,orNS4B. In contrasttothe cleavage productsgenerated
by thewtpolyprotein (Fig. 4A, lane d,and4B, lanesdandk),
expression of
sig2A-5356
polyproteins containing mutationsCD5, CD3, andCD38 yielded no detectable processing (Fig. 4A, lanes f to h), but only a high-molecular-weight protein which comigratedwiththe unprocessedpolyproteingenerated
by replacing the putative catalytic NS3 serine residue with
alanine (NS3 amino acid position 138) (11) (Fig. 4A, lane 3,
and B, lanes e and 1). The
sig2A-5356
polyprotein containing the CD4deletiongenerated NS2B-, NS3-, andNS4B-specificproteinswhichwere consistent withprocessing at the2A/2B,
2B/3, 4A/4B, and 4B/5cleavagesites,althoughatlowefficiency
(Fig. 4A, lanei, and4B, lanesi and p). The 3/4Acleavage is
apparently very inefficient in the presence of this mutation. TheNS2B-specificproteinsincludedaspecies with an
electro-phoretic mobilityslightly fasterthan that of thewt NS2A-2B
cleavageproduct,consistentwith the4-amino-aciddeletion in
NS2B.SmallamountsofNS2B-specificproteinsmigratingwith
molecularmassesslightlysmallerthan 18and 14kDawerealso
[image:5.612.69.310.81.214.2]observed afterprolonged exposure. These results suggest that the CD4 mutation reduces cleavage efficiency rather than abolishing NS2B-3 proteinase activity as is the case with the otherconserved domain mutations.
Figure 5 illustrates the processing patterns of sig2A-5356
polyproteins containing the NS2B mutations 2D4, 214,
2D13,
1D3D, and 1131 as analyzed with NS2B-, NS3-, or NS4B-specific antisera. The mutant polyproteins generated NS2B-specificproteins whosemolecular weights corresponded to the sizespredicted by the engineered insertions or deletions (Fig. 5A, lanes e to i, and 5B, lanes d to h and k too). The presence ofmutations2DI3and1D3D consistently led to reduced levels ofNS2B-related proteins in transient expression, and proteins corresponding to mutant forms of NS2B were observed only after long exposures. Results with the NS3 antisera revealed that only the 1D3D mutation had any detectable effect on
production of NS3-specific cleavage products, and similar to
the3D3mutation (Fig. 3B, lane f), the 3/4A cleavageefficiency
wasreduced. An additional protein ofapproximately 27 kDa in molecular mass which reacted with NS3-specific antiserawas
observed with the 214 and 1131 mutations (Fig.SB, laneseand h). The nature of this protein was not further characterized. None of these mutations had any detectable effect on the production ofNS4B-specific cleavage products.
Evidence for association of NS2B and the NS3 proteinase domain. Previous studies suggested that YF NS2B and NS3
may be associated as a complex (11). That mutations in the conserved domain of NS2Bdramaticallyreduced orabolished processing of the nonstructural polyprotein suggests that this region may be involved in the formation of an NS2B-NS3 complex. A physical association between these two proteins
was studied by coimmunoprecipitation experiments using dif-ferent sources of proteinase components and either NS2B-specific or NS3-NS2B-specific antisera.Figure6 illustrates the results of coimmunoprecipitation using samples from YF-infected cells, cell-free translated
2B-3181,
or vaccinia virus recombi-nants expressing the2B-3181
polyprotein or 2B and31x8
proteins individually as sources of proteinase. With infected cells, antiserum to NS2Bweakly
coimmunoprecipitates
NS2B and NS3 under nondenaturing conditions (Fig. 6A, lanef),
whereas antiserum
3c
coimmunoprecipitates NS2B,NS3, and additional proteins (lane g) which comigrate with NS4B and the two forms of NS2A (8). Antiserum 3rweakly
coimmuno-precipitates NS3 and small quantities of NS2B under these conditions (lane h). With cell-free translated2B-31,8
as asource of proteinase components, antiserum
2B,
but not3r,
coimmunoprecipitated both NS2B and NS3
(Fig.
6B,
lanes d and f, respectively), whereas antiserum3c
weakly
coimmuno-precipitated NS2B (lane e). Similar to results with infected cells, antisera to NS2B and3c coimmunoprecipitated
NS2B and 3181 when expressed from a vaccinia virus recombinant expressing2B-318,
(Fig. 6C,lanes candd).
Antiserum3c,
but not 2B,coimmunoprecipitatedboth NS2B and3181 whenthese proteins were expressedwith separate vaccinia virus recombi-nants (lanes e andf). These results suggest that differences may exist between the interaction of NS2B and NS3 or3181
in cell-free and cellular systems (seeDiscussion).
Complex formation between 3181 and NS2B
proteins
con-taining mutations in the conserved domain. Because resultswith nondenaturing conditions indicated that NS2B and 3181 could be
coimmunoprecipitated,
theimportance
of the con-served domain in NS2B for this association wasanalyzed by
coimmunoprecipitationexperiments
using
vaccinia virusre-combinants expressing
318I
and NS2Bproteins containing
mutation CD3, CD5,or CD38. Thisapproach
was undertaken because lack of cleavage of2B-3181
polyproteins
containing
VOL.67, 1993
NS2B -> 40 .40 asom
on November 9, 2019 by guest
http://jvi.asm.org/
A
lane: a b
c
Ir
NS2A-2B
->-3
p18
-*
.NS2B
>
mB
lane: a b c d e,
_'-
_NS3 > mm a
NS4B > M
-r, -
3-FIG. 3. Transient
expression
of si mutations inNS2B.
BHK-21-15
celltransfected with
pET-8c-YF
DNAs Methods. Cells were labelled for 4SDS-solubilized
lysates
werenrenart
these mutations precluded direct analysis of the role of the
d
ef
g h conserveddomain incomplex
formation. Inaddition,complex
formation could not be demonstrated bycotranslation of the -200 NS2B and
381,
proteins in a cell-free system. Figure 7 illus-trates that undernondenaturing
conditions, antiserum to -97.4 NS2B immunoprecipitates the wt NS2B protein but not31,8
(lane
c)
when theseproteins
arecoexpressed
(also Fig. 6C).
S 11S
^ * Z 69 Antiserum toNS2B
alsoimmunoprecipitates
theNS2B
con-served domain mutants CD3, CD5, and CD38, but does notcoimmunoprecipitate
318I
(lanese,g,andi).Antiserum to NS3-46
(3(c)
coimmunoprecipitates
3I8I
and thewtNS2Bprotein
(lane
d),but notthe mutantNS2Bproteins(lanes f, h,andj).These results are consistent with the hypothesis that the conserved domain constitutes a structural feature of the NS2Bprotein
which is important for the association betweenNS2B and the
-30
NS3proteinase
domain(318,).
Thesecoimmunoprecipitation
experiments included a 1 M NaCl wash for which the associ-ation of
31
1with atleast some of the wtNS2B protein appearsto be stable.It is possible that3181canassociate withtheNS2B
mutants under less stringent salt conditions such that the deletion ofcharged residues may reduce but not
necessarily
eliminate association. In such acase, somefunctional
activity
could remain (seebelow).
Cleavage assayofwtand mutantNS2B proteins
expressed
in trans. In order to determine whetherthe conserveddomain
mutations CD3, CD5, and CD38 might still allow functional
-14.3 proteinase
activity
despite
their effectsonreducing
association between NS2Band 3I81 underhighsaltconditions,the abilityof these mutantsto allow trans cleavage in conjunction with --2 -... -~
318,
(6) was tested. Vaccinia virus recombinants expressing NS2B-3181, NS2B and3,I8,
or 318I and each of the NS2BmutantsCD3, CD5,and CD38 wereused aspotentialsources f g h i j k I m of proteinase for cleavage of the substrate
3540-5356
(6)which*
-, -, *.;4- -200 contains intact 3/4A, 4A/2K, 4A/4B, and 4B/5cleavage
sites,but no NS3 proteinase domain. Appearance of the NS4B -97.4 proteinisusedasevidence oftranscleavageatthe 4A/2K (21)
and
4B/5
sites.Expression
of theNS2B-31,81
recombinant or-69 coexpression of the NS2B and
318,
recombinants generatedproteinase activity
which cleaved the3540-5356
substrate togenerate NS4B(Fig. 8, lanes 1 and m). (Cleavageatthe3/4A sitewas not examined.) In contrast, coexpression of the
3.1I
recombinantwith anyofthe three recombinant NS2Bmutantsfailedtogeneratean activeproteinase,onthe basisofthe lack ofcleavageof the
3540-5356
substrate(lanesntop). AntiseratoNS2B immunoprecipitated proteins corresponding to the wt
-30 and mutant NS2B proteins (data not shown). These results suggest that mutationswhich abolishassociationofNS2Band
the
3118
proteinase domain alsoeliminate trans-cleavageactiv-ity.
EffectofmutationsinNS2Bonrecovery ofinfectious virus. Inaprevious study,itwasdemonstratedthatmutations which
reduce or abolish NS3 proteinase activity are deleteriousfor viral
replication
(11). Because the NS2B mutations studieda--..r,- 4 ;0. here had variable effectson
proteolytic
processing,
their effectsig2A-5356,
polyproteins containing on viral replication were therefore determined. The pheno-lswere infected with vTF7-3 and types of five of these mutations (1D3, 2D4, 3D3, CD3, and ,as described in Materials and CD4) weredetermined byconstructing full-lengthYF cDNA 4.5 h with[35S]methionine,
and templates and assaying the infectivity of RNA transcripts ed and immunonrecinitated withantisera to YFnonstructural
protein
NS2B, -3, or -4B(indicated
atbottom). Immunoprecipitates
wereanalyzed
by
SDS-14%(A)
or-12%
(B)
PAGE, followedby
fluorography.
Thepositions
of YFproteins
from transfectionsorfromYF-infectedBHK-21 cellsimmu-noprecipitated
withmousehyperimmune
ascitic fluidare indicatedatthe left. Sizes of molecular
weight
standards(in
thousands),
asdetermined with
'4C-labelled high-molecular-weight
proteins,
arein-dicated on the
right.
(A) NS2B-specific
cleavage
products.
Lanes:Lanes: a, YF proteins; b, high-molecular-weight markers; c, mock-transfected cells (vTF7-3 infectiononly); d, wt sig2A-5356; e, 1D3; f, 113; g, 3D3; h, 313. (B) NS3-and NS4B-specific cleavageproducts. Lanes: a, YF proteins; b and h, mock-transfected cells; c and i, wt
sig2A-5356;d andj, 1D3;eandk, 113; fand l,3D3;g and m, 313.
.VttIf
*MP
0
-aw-:
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.55.294.83.608.2]YELLOW FEVER VIRUS PROTEINASE COMPLEX 6803
lane:
ab
c
d
ef
g hi
w WW -200
_- ~~~~~-97.4
-69
-46
A
lane:
a
b
c
d
e
f
g h i
_~~~~~~~~~~:,... i lT
..-mo
Wm..
NS2A-2B
>
QmbIW
-30
p18
->b w~~~~-Fw-w,I.
mm
NS2B
>2
-NS2B
>0C
2
2B
lane:a b c d ef
g h ij
k I m n o pNS3-*-S
-14.3
B lane: a b c d e f g h
7t
_t-
* *.:4 .U0-*Ia
NS3 -- ms -=
i j k I m n o
' -200
-97.4 -69
-200 -97.4
-69 NS4B -4
-46
-30
-14.3 -30
NS4B >
[image:7.612.67.305.81.577.2]ri 3---1 '* 4
FIG. 4. Transientexpression of
sig2A-5356
polyproteins containing mutations in the NS2Bconserved domain. Experimental procedures,YFand molecularweight markers,andfigurelabelsareasdescribedin thelegendtoFig.3.(A)NS2B-specific cleavage products.Lanes:a,YF
proteins; b, high-molecular-weight markers;c,mock-transfectedcells; d,wt;e,transfection withsig2A-5356,containingsubstitution of alanine for serine atNS3 amino acid 138 (sig2A-5351*); f, CD5; g, CD3; h,
CD38; i,CD4.(B) NS3- andNS4B-specific cleavage products.Lanes: a, YF proteins; b, high-molecular-weight markers; c and j, mock-transfectedcells;dandk,wtsig2A-5356; eand1,sig2A-5351*;f andm,
CD5; gandn, CD3;h ando,CD38; iandp,CD4.
derived from eitherwtormutant constructs. Incontrast to wt
transcripts, which yielded more than 200 plaques after
trans-fection of250ngofRNA,noneof themutanttranscriptswere
[image:7.612.327.562.89.497.2]capable of generating infectious virusby plaqueassay.Medium
FIG. 5. Transientexpression ofsig2A-5356 polyproteins containing mutations in NS2B. Experimental procedures, YF and molecular
weight markers, and figure labelsare asdescribedin the legendtoFig. 3. (A) NS2B-specific cleavage products. Lanes: a, YF proteins; b,
high-molecular-weight markers;c,mock-transfected cells; d,wt
sig2A-5356;
e, 2D4; f, 214; g, 2D13; h, 1D3D; i, 1131. (B) NS3- andNS4B-specific cleavage products. Lanes: a, YF proteins; b and i,
mock-transfected cells; candj,wtsig2A-535,,; d and k,2D4;e and
214;fandm,2D13;gandn, 1D3D; hando, 1131.
collected at 72 h after transfection with 250 ngoftranscripts
was analyzed for infectious virus by plaque assay on SW-13 cells.wttranscripts yielded 107PFU/ml, whereasnoinfectious
viruswasrecovered after transfection with mutant transcripts (datanot shown). These resultssuggest that mutationsin the hydrophobic domains as well as in the conserved domain of NS2B are deleterious for viral replication.
DISCUSSION
In previous studies, the absoluterequirement forNS2B for cleavages mediated by the YF NS3 proteinase within the nonstructural region of the polyprotein wasdemonstrated (6, 21). Thesetwoproteinshave been shown togenerate
protein-A
p18
->
-200 -97.4 -69
-46
-30
-14.3 VOL. 67? 1993
NS2A-2B
> 40 4w &at
on November 9, 2019 by guest
http://jvi.asm.org/
6804 CHAMBERS ET AL.
A
lane: a b c d e f g h
4
NS3->
-In
B
C
lane: a b c d e f lane: a
.,
b c d e f,,WIF
';--'...
..._
_l'
_NS2B
-31
>3181
NS2B > NS2B >
3 -_ l
181
NS2B >
2 3c 3r 2 3c 3r 2 3c 3r 2 3c 3r 2 3c 2 3c 2 3c
FIG. 6. Analysisofassociationbetween NS2B and the NS3proteinase domainbynondenaturingimmunoprecipitation. (A)BHK-21 cellswere
infected or mock infected with YF17D and labelled from 18 to 24 h postinfection. Monolayerswere solubilized in 1% Nonidet P-40-1%
deoxycholate-0.1%SDSin 50 mM Tris(pH 7.5)-150mMNaCl,andthelysatewasreacted with antiserumtoNS2Bortwodifferent antisera to NS3(3,and3r).Immunoprecipitateswerewashedasdescribedin MaterialsandMethods,solubilized in Laemmlisample buffer,andanalyzedby SDS-PAGE witha 14% gel. Lanes:a, YFproteins; b,high-molecular-weightproteins; c toe, mock-infectedcells;ftoh, YF17D infected. (B)
Analysis of NS2B and3lx, association in cell-freetranslation.Reticulocyte lysatesweresolubilized innondenaturing detergentsand reacted with NS2BorNS3 antiseraasdescribed above.Immunoprecipitated proteinswereanalyzedon a 14%gel.Lanes:atoc,noRNA;dtof,2B-3181 RNA. (C) AnalysisofNS2B and311association using vaccinia virus recombinants. BHK-21 cellswereinfected with vTF7-3 and eitherarecombinant expressing
NS2B-318,
or tworecombinantsexpressing NS2BandNS31,1.At3 hpostinfection,cellswerelabelled with[35S]methionineasdescribed inMaterials and Methods,for4h.Followinglabelling, immunoprecipitation andanalysisofNS2B-andNS3-specific proteinswerecarriedoutasdescribed forpanel A. Lanes:a andb,vTF7-3 only;candd,vNS2B-3181;e andf,vNS2Bplus
v31,,.
Linesto the rightofeachpanelindicate molecularmasses(in kilodaltons) indecreasing order:200, 97.4, 69, 46,30(except inpanel B),and 14.3.ase activity when expressed as a
2B-3181
polyprotein or as separate NS2B and3I8I
proteins and to cleave substrates in trans under both circumstances. It has been hypothesized, therefore, that the active YF proteinase is a heterodimer,composed of NS2B and the NS3proteinase domain.Although
it has been reported that the DEN2 NS2B and NS3 proteins
can form a stable complex (2), the structural basis for this interaction has not beendefined. These previousfindings are now extended toindicate thataconserved domainwithin the
YF NS2B protein is essential for several properties of the proteinase. These includeciscleavageatthe2B/3 cleavagesite;
transcleavage,atleastatthe4B/5 site; and association of NS2B with the NS3 proteinase domain. In addition, the conserved domain and other regions within NS2B are required for YF
replication.
The mutations studied in these experiments had variable effects on processing of the YF nonstructural polyprotein
precursor. Whereas no major alteration of the pattern of
processingwasobservedwith mutations inthefirst two
hydro-phobic regions (1D3, 113, 2D4, 2D13, 214, 313, and 1131),
reducedcleavageefficiencyat the 3/4A site was observed when
mutationswere introduced into the third hydrophobic region
(3D3 and I1D3D). Mutations within the conserved domain either reduced cleavage efficiency at all sites (CD4) or
abol-ishedallcleavage activity(CD3, CD5, and CD38). Interpreting the role ofspecific regions ofNS2B targeted by these
muta-tions is limited by the fact that certain mutations included
either conservative or nonconservative amino acid
substitu-tions. However, a similar approach for analyzing the role of
hydrophobic residues in the processing of the poliovirus
polyprotein has yielded useful information (16), and in some cases, even nonconservative substitutions have been tolerated
in membrane-associated regionsof proteins (5). The failure of
mutationsin thefirstand second hydrophobic regionsofNS2B
toinhibitprocessing of the
sig2A-5356
polyproteinsuggests thattheseregionsmaynotbe critical for either the topologyof the
polyprotein necessary for processing or the participation of
NS2B inproteinaseactivity. Theeffect of the mutation in the
third hydrophobic region on 3/4A cleavage suggests that the
NS2B-NS3proteinase maybe sensitive to different structural
constraints forcleavageatthissite, which has been observedto occur inefficiently relative to cleavage at other sites (6, 23).
This observation must be interpreted cautiously because the
introduction of acharged residue in this hydrophobic region
may reduce the stability or activity of the proteinase, which might haveamoredramatic effectatinefficiently cleaved sites such as 3/4A. Within the conserved domain, elimination of only three amino acid residues (CD3 mutation)wassufficient
to abolish allcleavage activity. The pronounced effect of this andother mutationsin this region of NS2Bsuggests that the conserved domain has a critical role in proteinase function. The conserved domain of DEN4 has been shown to be required forcis cleavage of NS2B-3 polyproteins and specifi-cally to encompass a 40-amino-acid region whose N and C
termini have beenprecisely determined (12). The N-terminal region of theDEN4NS2B conserved domain wasfoundtobe especially important for cleavage activity,and the results with theCD3andCD5mutationsreported here areconsistent with this finding. In fact, comparison of the homology among
flaviviruses reveals a higher level of conservation among
residues in the N-terminal portionof the domain (12). Results
reported here with the CD4 mutation indicate that certain
residues, particularly toward the C-terminal end of the
40-amino-aciddomain,maybe less criticalfor proteinase activity. It is unlikely that the effects of mutations in the conserved
region result merely from distortionof theconfiguration ofthe
2B/3 cleavage site.
sig2A-5356
polyproteins containing theCD38mutationcanbecleaved intrans atthe 2B/3 cleavage site
J.VlIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.71.541.85.261.2]YELLOW FEVER VIRUS PROTEINASE COMPLEX 6805
lane: a b c d e
f
g
h
ij
In7
w_2004
.''..js 69
w w * .:-~-46
*#_
-30
4m am - 4 -21.5
lane: a b c d e f g h i j k I m n o p -200
- - __ a
3540-5356 -97.4
-69
-46
-30
3181
NS2B > - _ -14.3
-12.5
[image:9.612.68.308.80.327.2]antisera: 2
3c
23c
23c
23c
2 3cFIG. 7. Analysis of NS2B and NS3 association using vacciniavirus
recombinants expressing NS2B deletions in the conserved domain. Experimentalprocedureswereperformedasdescribed in thelegendto Fig. 6C. Lanes:aand b. vTF7-3 only;cand d, vNS2B plusv3,8,;eand
f, vNS2BCD3 plus
v3,1,;
g and h, vNS2BCD5 plus v3151 i and j,vNS2BCD38 plusv3,x
(10), indicating that the cleavage site is still accessible in the
context of this mutation. Similar findings have been observed with the DEN4 2B/3 cleavage site (12). Furthermore,
muta-tions which reduce orabolish YF 2B/3 cleavage do not affect processing at other cleavage sites (10), suggesting that
inter-action ofthe NS3 proteinase domain with NS2Bmayprecede
cleavage at the 2B/3 site and other proteinase-dependent cleavage sites.
The results reported here also suggest that the conserved domain may be an important determinant of the interaction
between NS2B and the NS3proteinase domain. With different systems forexpression of NS2B and NS3, theseproteins have been showntoremain associated duringimmunoprecipitation undernondenaturing conditions, although the results depend
on the source of the proteins and the antibody used. Differ-ences were observed in the association of NS2B and NS3 amongcellularextractsand cell-freetranslationsystems.These findings may result from differences in the folding of the proteins in thesetwosystems,resulting in eitheradecrease in
availability of antibodytospecific binding sitesor analteration in the proportions of complexed versus uncomplexed NS2B and NS3.Onlyalimited fraction of NS2Bappearstoassociate in trans with the proteinase domain under nondenaturing conditions. This could reflect the fact that association of the
two proteins is inefficient when they are produced in trans
relative to cis because of concentration effects or structural
differences which exist when these proteins fold separately rather thanas a2B-31,8 polyprotein. The failure of YFNS2B proteins containing deletions in the conserved domain to
associate with the NS3 proteinase domain in coimmunopre-cipitation experiments is consistent with the hypothesis that either this domain forms a binding site for the proteinase
domainorthe domaindirectlyaffects asiteonNS2B which is
-14.3
3 4
FIG. 8. trans-cleavage analysis using
3,81
and NS2B conserveddomain mutants. BHK-21-15 cells were transfected as described in
Materials and Methods and in the legend to Fig. 3. Following
coinfection with vTF7-3, v3181, and vaccinia virus recombinants
ex-pressing wt or mutant NS2B proteins, cells were transfected with
3540-5356'Cellswere labelled with [35S]methionine for 5 h, following
which immunoprecipitation and analysis of proteinswerecarriedout as described in the legend to Fig. 3. Lanes: a, YF proteins; b, high-molecular-weight markers;candj, vTF7-3 alone; d and k,vTF7-3
plus 3540-5356 DNA; e and 1, vNS2B-3151; f and m v3181 plus vNS2BCD3; g and n, v3181 plus vNS2BCD3; h and o, v3181 plus vNS2BCD5; i andp,v3l plus vNS2BCD38.
important for interaction with NS3.Althougha morecomplete
understanding of complex formation could be obtained by testing complex formation in trans with vaccinia virus NS2B recombinants containing mutations in the hydrophobic
re-gions, analysis of in vitro-translated
2B-31.1
polyproteinscon-taining these mutations revealed that, under nondenaturing conditions, coimmunoprecipitation of NS2B and
3.1x
occurs(data not shown). The clustering of charged residues in the conserved domain is consistent with its function as asite for interaction between proteins (17). The predicted cytoplasmic orientation of this region, based on its hydrophilic character
and thetopology of the surroundinghydrophobic regions (30), is consistent with the hypothesis that NS2B may be a
mem-brane-associated protein which interacts with the cytoplasmic NS3 and localizes it to membranes. Although it has been reported that onlyafraction of NS3 islocalized tomembranes in thepresenceof NS2B(13), it is possible that morethanone
protein, possiblyevenhostproteins, mayparticipate in
associ-ation of NS3 with membranes. Localization of NS3 to the membrane may be necessary for processing of the viral
polyproteinorfor functionof the replication complex,both of which arebelieved tobe membrane-associated processes(see reference 7 forareview). On the basis ofthe currentdata,we
favor amodel in which association of the proteinase domain with the NS2B conserved domain occurs prior to and is
necessaryfor2B/3 cleavageand is followed byformation ofa
stable complex. A conformational change may occur in
con-junction with 2B/3 cleavagewhich enhances proteinaseactivity, since 2B-318, polyproteins which are blocked for cleavage at
the2B/3 site have reducedcleavage activity inti-atisrelative to acis-processedwt2B-3,81 proteinase (10). Apossiblerole for
NS2B may be to stabilize NS3 in an active conformation.
Factors which disrupt this interaction, by association with
either NS2B or NS3, could be important for regulating
pro-teinase activity during virus infection. However, we cannot
3181
>VOL. 67, 1993
NS4B
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.330.566.82.251.2]exclude the
possibility
thatthe conserved domain functionstostabilize thestructureofNS2Band that the site for interaction withthe NS3
proteinase
domain does notdirectly
involve thisregion.
Morecomprehensive genetic
and biochemical andultimately high-resolution
structural studies will berequired
toprecisely
define thesite(s)
of interaction.In contrast to the variable effects that mutations in NS2B had on
polyprotein processing,
all mutations tested had lethal effects on viralreplication
as testedby
the recovery of infec-tiousplaque-forming
virus. The deleterious effects ofmuta-tions which abolishor
significantly
reduceproteolytic
process-ing (CD3
andCD4)
are similarto resultspreviously
obtained with mutations at Ser-138 of the NS3proteinase
domain.Although processing
appeared
normalwith the 1D3 and 2D4mutations,
andonly
slightly
altered with the 3D3mutation,
thesewereall lethal for
replication.
Theprecise
stageatwhich the mutations blockreplication
has not been defined. It ispossible
that these mutationsdisrupt
either theassembly
orthe function of thereplication complex,
which has beenproposed
to consist of the NS3 and NS5
proteins
together
with theproteins
derived from the NS2 and NS4subregions (reviewed
in reference
7).
Theprecise
interactions between NS2B and these additional viralproteins
remaintobedefined, although
coimmunoprecipitation experiments
with YF-infected cellex-tracts with antisera to NS4B and NS5 suggest that stable interactions mayoccur
(10).
Ithas been observed forpoliovirusthat some mutations in the
hydrophobic
domain of the 3Aregion
interfere with RNAsynthesis
in some cases withoutabolishing polyprotein
processing
(16).
Inaddition,
mutations within theproposed
FGloop
of thepoliovirus
3Cproteinase
also exhibit
disparate
effectsonproteolytic processing
andviralreplication
(19).
Itwould appearfromexamples
such asthese thatpolyprotein processing
andassembly
ofprocessed proteins
into functional
replication complexes
are discrete steps, andproteins
which are involved inprocessing
andreplication
may have several functions. Whether the NS2B mutations studied here actsolely
through altering
the association of NS2B andNS3,
orby interfering
with otherfunctions ofNS2B,
remainstobedetermined.
ACKNOWLEDGMENTS
The contributions of Marc S. Collett, whoprovided antisera, and
ChangS. Hahn andBarryFalgout,for discussionsrelatingtothiswork,
aregratefully acknowledged.
T.J.C.wassupported byPublic HealthServiceawardA100973.A.N.
wassupported bytheNational Health and Medical Research Council of Australia. C.M.R. acknowledges support from the Pew Memorial
Trust, the Public Health Service (AI31501),andU.S. Armycontract DAMD17-87-C-7154.
REFERENCES
1. Amberg,S.M.,andC. M. Rice.Unpublisheddata.
2. Arias, C. F., F. Preugschat, andJ. H. Strauss. 1993. Dengue 2 virus NS2B andNS3form astablecomplex that cancleaveNS3 within the helicase domain.Virology 193:888-899.
3. Bazan,J.F.,andR.J. Fletterick. 1989.Detection ofatrypsin-like serine protease domain in flavivirusesand pestiviruses.Virology 171:637-639.
4. Bazan, J. F., and R. J. Fletterick. 1990. Structural and catalytic models oftrypsin-likeviral proteases. Semin. Virol. 1:311-322. 5. Boyd,D.,andJ.Beckwith. 1990. Therole ofcharged amino acids
in the localization of secreted and membrane proteins. Cell 62:1031-1033.
6. Chambers,T.J.,A.Grakoui,andC. M. Rice. 1991. Processing of the yellow fever virus nonstructural polyprotein: a catalytically activeNS3proteinasedomain and NS2B arerequiredfor cleav-agesatdibasic sites. J. Virol. 65:6042-6050.
7.
Chambers,
T.J., C. S. Hahn, R.Galler, and C. M. Rice. 1990.Flavivirus genome organization, expression, and replication.
Annu. Rev. Microbiol. 44:649-688.
8. Chambers,T.J., D. W. McCourt,and C. M. Rice. 1989. Yellow fever virus proteins NS2A, NS2B,and NS4B: identification and partial N-terminal amino acid sequence analysis. Virology 169: 100-109.
9. Chambers,T.J.,D. W.McCourt,andC. M. Rice. 1990. Production ofyellow fever virus proteins ininfected cells: identification of discretepolyproteinspeciesandanalysisofcleavagekineticsusing
region-specific polyclonalantisera.Virology177:159-174. 10. Chambers, T.J., and C. M. Rice.Unpublished data.
11. Chambers, T.J., R. C.Weir, A. Grakoui, D. W. McCourt, J.F. Bazan, R.J. Fletterick,andC. M. Rice. 1990. Evidence that the N-terminal domain of nonstructuralproteinNS3 fromyellowfever virus isaserine proteaseresponsibleforsite-specificcleavagesin the viralpolyprotein. Proc. Natl. Acad. Sci. USA 87:8898-8902. 12. Falgout,B., R. H.Miller, andC.-J.Lai.1993. Deletionanalysisof
dengue virus type 4 nonstructuralprotein NS2B:identification of
a domain required for NS2B-NS3 proteinase activity. J. Virol. 67:2034-2042.
13. Falgout, B., M. Pethel, Y. Zhang, and C.-J. Lai. 1991. Both nonstructural proteins NS2B and NS3 are required forthe pro-teolyticprocessing ofdengue virus nonstructuralproteins.J.Virol. 65:2467-2475.
14. Falkner, F., and B. Moss. 1988. Escherichia coli gpt geneprovides
dominant selectionfor vaccinia virus openreading frame expres-sion vectors. J. Virol.62:1849-1854.
15. Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss. 1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that synthesizesbacteriophage T7 RNApolymerase. Proc. Natl. Acad. Sci. USA83:8122-8126.
16. Giachetti, C., S.-S. Hwang, and B. L. Semler. 1992. cis-acting lesionstargeted tothe hydrophobic domainof a poliovirus
mem-braneprotein involved in RNAreplication.J.Virol. 66:6045-6057. 17. Gibbs, C. S., and M. J. Zoller. 1991. Identification of functional residues in proteins by charged-to-alanine scanningmutagenesis. Companion to Methods Enzymol. 3:165-174.
18. Gorbalenya, A. E., A. P. Donchenko, E. V. Koonin, and V. M. Blinov. 1989. N-terminal domainsofputative helicases of flavi-and pestiviruses may be serine proteases. Nucleic Acids Res. 17:3889-3897.
19. Hammerle, T., A. Molla, and E. Wimmer. 1992. Mutational analysis of the proposed FG loop of poliovirus proteinase 3C identifies amino acids that are necessary for 3CD cleavage and might be determinants of a function distinct from proteolytic activity. J. Virol. 66:6028-6034.
20. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
21. Lin, C., S. M. Amberg, T. J. Chambers, and C. M. Rice. 1993. Cleavageat a novel site in the NS4A region by the yellow fever virus NS2B-3 proteinase is a prerequisite for processing at the downstream4A/4B signalase site. J. Virol. 67:2327-2335. 22. Lin, C., T. J. Chambers, and C. M. Rice. 1993. Mutagenesis of
conservedresidues at the yellow fever virus 3/4A and 4B/5 dibasic cleavage sites: effects on cleavage efficiency and polyprotein processing.Virology192:596-604.
23. Lobigs, M. 1992. Proteolytic processing of a Murray Valley encephalitisvirus non-structural polyprotein segment containing the viral proteinase: accumulation of aNS3-4Aprecursor which requires mature NS3 for efficient processing. J. Gen. Virol. 73:2305-2312.
24. Mackett, M., and G. L. Smith. 1986. Vaccinia virus expression vectors.J. Gen. Virol. 67:2067-2082.
25. Moss, B., 0. Elroy-Stein, T. Mizukami, W. A. Alexander, and T. R. Fuerst. 1990. New mammalian expression vectors. Nature (Lon-don)348:91-92.
26. Nestorowicz, A., T. J. Chambers, and C. M. Rice. Unpublished data.
27. Preugschat, F., C.-W. Yao, and J. H. Strauss. 1990. In vitro processing of denguevirus type 2 nonstructural proteins NS2A, NS2B,and NS3. J.Virol. 64:4364-4374.
28. Rice, C. M., A. Grakoui, R. Galler, and T. J. Chambers. 1989.
on November 9, 2019 by guest
http://jvi.asm.org/
YELLOW FEVER VIRUS PROTEINASE COMPLEX 6807
Transcription of infectious yellow fever virus RNA from full-length cDNA templates produced by in vitro ligation. New Biol. 1:285-296.
29. Rice, C. M., E. M. Lenches, S. R. Eddy, S. J. Shin, R. L. Sheets, andJ. H.Strauss. 1985.Nucleotidesequenceof yellow fever virus: implicationsforflavivirusgene expressionand evolution. Science 229:726-733.
30. Rice,C. M., E. G. Strauss, and J. H. Strauss. 1986. Structure of the
flavivirus genome, p. 279-326. In S. Schlesinger and M. J.
Schlesinger(ed.),TheTogaviridaeand Flaviviridae.PlenumPress, New York.
31. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning:alaboratorymanual, 2nd ed. Cold Spring Harbor
Labo-ratory, Cold SpringHarbor, N.Y.
32. Studier, F. W., A. H. Rosenberg, J. J. Dunn, and J. W.Dubendorff.
1990. Use of T7 RNA polymerasetodirect expression ofcloned
genes.Methods Enzymol. 185:60-89.
33. Wengler,G.,G.Czaya, P. M. Farder, and J. H. Hegemann. 1991. In vitro synthesis ofWest Nile virus proteins indicates that the
amino terminal segment ofthe NS3 protein contains the active
centre of the protease which cleaves the viral polyprotein after multiple basic amino acids.J. Gen.Virol. 72:851-858.
34. Wengler,G.,G. Wengler, T. Nowak, and E. Castle. 1990. Descrip-tion ofa procedure which allows isolation of viral nonstructural
proteins from BHKvertebrate cells infected with the West Nile
flavivirus in astatewhich allows their direct chemical
character-ization. Virology 177:795-801.
35. Zhang, L.,P. M.Mohan,and R. Padmanabhan. 1992. Processing
and localization of dengue virus type 2 polyprotein precursor
NS3-NS4A-NS4B-NS5. J.Virol.66:7549-7554. VOL.67, 1993