0022-538X/81/100011-09$02.00/0
Sequence
Complexity of Circular Epstein-Barr
Virus DNA
in
Transformed Cells
BEVERLY E. GRIFFIN,t EVA BJORCK, GUNNAR BJURSELL, AND TOMAS LINDAHL*
Department of MedicalBiochemistry, Gothenburg University, S-400 33 Gothenburg, Sweden
Received11February 1981/Accepted 3 June 1981
A simplified procedure, based on several methodspreviously used to isolate
circular DNAmolecules from bacteria,was devisedforthe preparationof
cova-lently closed circularviral DNA molecules from large quantities oflymphocytes
transformed by Epstein-Barr virus. The protocol can be applied both to virus
nonproducer lines and to lines containing cells activated to virus production.
Sufficient amounts of highly purified viral DNA of intracellular origin were
obtainedfrom B95-8 and Raji cellstoallow direct visual analysis of theirsequence
complexities after cleavage with EcoRI and separation of fragments by gel
electrophoresis. No major differences in complexity were observed between
circular DNA and linear virion DNA from B95-8 cells. The fragmentpatterns
observedinthis fashionagreewell withthosedetectedbyconventionalblotting
and hybridization methods. The procedure can also be used as an analytical
methodtoassayfor smallamountsof circular Epstein-Barr virusDNAmolecules
inothertransformedcells. In this connection,nocircular Epstein-Barr virus DNA
wasdetected inNamalva cells.
TheEpstein-Barr virus(EBV) transforms
hu-manBlymphocytes effectivelyto acellular form
thatgrowswell in tissueculture,andthe
trans-formed cellsexpress avirus-determined nuclear
antigen,EBNA (17, 27). Inmanysuchcells, the
viral genomes are present mainly as
non-inte-grated, covalently closed circular DNA
mole-cules (21, 23). Small amounts of EBV DNA
circles, suitable for characterizationby electron
microscopy, have been isolated previously in
more than 90%pureform(21).However, larger
quantities of circular EBV DNA formore
de-tailed studies ofsequenceorganization and
com-plexity havenotbeenavailable,because
stand-ardmethods usedtopreparesmall circular DNA
molecules from varioustypes ofcells have not
been applicable to EBV-transforned
lympho-cytes. Comparisons among viral DNA circles
presentin differenttypesof transformed cellsby
analysisof restrictionenzymecleavagepatterns
have therefore only been perforned with
unpu-rified (32)orpartly purified (16,29) EBVDNA
preparations, using selective hybridization
pro-cedures to detect the EBV DNAsequences. In
the present.study, wedescribe the purification
ofrelatively largeamountsofcircular EBV DNA
fromRajiand B95-8cells,free from host nuclear
DNA. The availabilityof such material has
al-t Presenal-t address: Imperial Cancer Research Fund,
Lin-coln's InnFields,London WC2A3PX,England.11
lowed an analysis of restriction enzyme digests
of circular intracellular EBV DNA by optical
methods.
MATERIALS AND METHODS
Cefls. The EBV-transformed lymphoid cell lines
Raji, Namalva, and B95-8 were propagated without
agitationassuspensioncultures in2-liter volumes in RPMI 1640 medium supplemented with 10% fetal bovine serum, 100U ofpenicillinperml,and100,ugof streptomycin per ml. Raji and Namalvaare Burkitt
lymphoma-derived lines (11, 27), whereas B95-8 isa
marmosetlymphocytelineimmortalized with human EBVfromamononucleosispatient (22). Best results wereobtained with actively growing cells, harvested
at adensityof106cells per ml andcontainingahigh
proportion ofliving cells (>85%) as judged by the
trypanblueexclusiontest.
Viral DNA and hybridization probes. Linear EBV DNAwasprepared from virus particles released by the B95-8 cell line (21). The EcoRI fragmentof circular EBV DNA from Raji cells, which contains sequencescorrespondingtothe endfragmentsof the linear DNA from virus particles, wascloned in the cosmid pHC79 (2a). These DNA preparations were radioactively labeledinvitrobynicktranslationinthe presenceof[a-32P]dCTP(28).Thespecific activityof thelabeled DNAwas1 x 108to2x108cpm
Lg-'.
Isolation of circular EBV DNA.Lymphoidcells
(2 x 109 to 4 x 109) were collected by low-speed
centrifugationandwashedoncewith0.14MNaCl-0.01
M sodium phosphate (pH 7.4). The cells were
sus-pendedinasmall amountof this saline solution by
on November 10, 2019 by guest
http://jvi.asm.org/
12 GRIFFIN ET AL.
brief blending inaVortex mixertogenerate athick but homogeneous cellsuspension of about109 cells per ml. Analkaline sodium dodecyl sulfate buffer was then immediately added to lyse thecells (1 ml per 107 cells). This buffer, which contained 50 mM NaCl-2 mM EDTA-1% sodiumdodecyl sulfate, broughttopH12.4 with 2 MNaOH, wasfreshly made and adjusted to thecorrectpH usinganalkali-resistant electrode,or by correcting for the reading error of conventional combination electrodes(by calibration withapH12.4 standard buffer that contains25ml of0.2MKCI,16.2 mlof 0.2 MNaOH,and58.8ml ofwater). A10- to 15-mlportionof the very viscous cell lysate was poured into a50-mlconical centrifuge tube (Falcon Plastics, Oxnard,Calif.),withsample separation being achieved by cutting through theviscouslysate with a pair of scissorsduring pouring.The tubewascapped, and the sample of the lysatewassheared for2min on aregular laboratory Vortex apparatus operating atmaximum speed (e.g.,aSupermixer, Lab-Line Instruments, Inc., Melrose Park, Ill.). The lysate should form a thin liquid film coveringmostof thewall of the tube during the Vortextreatment.The processwasrepeated until all of the DNA had been sheared. The combined, slightly viscous lysatewasthen incubatedat300Cfor 30mintoallowcomplete alkali-induced strand sepa-rationof linear DNAmolecules. The final lysatewas supplemented with 0.05 volume of 1 M Tris-hydro-chloride (pH7.1), toobtain apHvalue of 8.5to 9.0, and then with 0.2volume of3MNaCl. These additions weremadeslowly withgentle swirling. Proteinase K (E. Merck AG,Darmstadt, Germany), 0.02 volume of a0.5%solution,wasthenadded, and the lysate was incubatedat37°Cfor30mintoallowpartial degra-dation of cellular proteins. This protease digestion
appeared necessary forobtaining goodyieldsof EBV
DNAduring subsequentsteps.Thelysatewaschilled
to200Candmixed with one-third volume of redistilled phenol, previously saturated with0.2MNaCl-0.2M
Tris-hydrochloride (pH 8.0). The phenol extraction
was performed byslowly pouring the mixture back
and forth betweentwoglassbeakers. Afterchillingto
8to100C,thelysatewascentrifugedfor 20 min in
250-ml Corex bottlesat6,000 rpm inaSorvalllaboratory
centrifuge. A clear aqueous layer, containing RNA
fragments and covalently closed circular DNA, was observed aboveawhiteinterphaseofasingle-stranded
DNA andremainingprotein.The aqueous, nonviscous phasewasrecovered with awide-mouth pipette and
gentlyextracted with1volume ofchloroform-isoamyl
alcohol(24:1)toremove mostof thedissolved phenol. The phasesseparated quickly onstanding, and the bottom phase was removed and discarded. The aqueousphasewasmixed with2volumes of cold 95% ethanol and leftovernightat-20°C.Asmall precipi-tate developed, containing RNA and circular DNA. The RNA apparently servedascarrier and ensured effectiveprecipitation of the very dilute DNA in this step. The precipitate was recoveredby centrifuging portions of themixture for10mineachat6,000 rpm in thesame250-ml Corex tube. The tubewasdrained upside down for15 to 20min,and thenaCsCl(Merck, Suprapur) solution (6 g of CsCl in6ml of10mM Tris-hydrochloride-1 mM EDTA [pH 7.5]) was added to
J. VIROL.
thestill-moist precipitate. The latter dissolvedreadily,
and the solutionwastransferredto anultracentrifuge
tube. After the addition of 0.1 ml of 1% ethidium bromide andoverlayingwithparaffinoil,thesolution wascentrifuged for40hinaBeckman 50-Tirotor at 40,000 rpmand200C.At the endof the run, the tube wasexamined underlong-wave UVlight.Apelletof RNA and two DNA bands of approximately equal
intensitywere seenintypicalexperiments. The lower band, containingcovalently closed circular DNA,was collected through alargehole in the bottom of the tube with the aid ofaclosed-systemcollectiondevice,
andethidiumbromidewasremoved from this solution by fourextractions withanequalvolume of
isopropa-nol, presaturatedwithasimilarly concentratedCsCl
solution. The DNA solutionwasthensupplemented
withadditionalsolidCsCl (0.25 g/ml) and dilutedto
avolume of 6 mlbythe addition ofmoreCsClsolution.
The densityof this solutionwasadjustedto1.716g/
cm3 withthe aid ofarefractometer,and thesolution wascentrifuged for48h ina50-Tirotor at34,000 rpm
and200C.Atthe end of the run, 0.2-ml fractionswere
collected throughalarge hole in the bottom of the tube. Each fractionwasdiluted with3volumes of10
mM Tris-hydrochloride-1 mM EDTA (pH 8.0), and
theabsorbancy at260 nmwasdetermined.
Low-mo-lecular-weight RNA mainly resided in the bottom
fractions, and the peaks ofcircular EBV DNA and
lighter mitochondrial DNA wereseparated by2to 3 fractions. The EBV DNA (usually intwo fractions)
waspooledintoasiliconized Corex glass tube, mixed
with 2volumes of cold ethanol, and kept at -20°C
overnight. After centrifugation, the DNA was
dis-solved in50plof10mM Tris-hydrochloride-1 mM EDTA(pH8.0) and storedat40C.Long-term storage of thecovalentlyclosed circular EBV DNA in concen-tratedCsClsolution should beavoided, because inour
experiencenicksslowlyappear.
If theyieldof circular EBV DNA is low, the position of thelighterband of mitochondrial DNA may be used as reference to localize the virus DNA. A sensitive fluorescence method (9)orhybridization of portions of eachgradientfraction with32P-labeledEBV cRNA has also beenemployedtolocalize the circular EBV DNAafterCsCl density gradient centrifugation. As an alternative tothe second CsCl fractionation step, in
someexperiments glycerol gradient centrifugationof
thecovalentlyclosed circular DNA from the ethidium
bromide-CsCl gradient was performed as described (21), and the materialsedimentingat 90 to115Swas recovered.
Inanalytical experimentstosearchfor the presence of circular EBV DNA in cells, fractions from the ethidiumbromide/CsCl gradientwereextractedwith isopropanol, boundtomembraneifiters,anddirectly hybridized withEBV32P-cRNA(21).
Electronmicroscopy. DNA preparationswere ex-amined by the formamide modification of the Kleinschmidt technique, and length measurements wereperformedasdescribed (21).BacteriophagePM2 DNAwasusedastheinternal size marker.
Gelelectrophoresisof EBV DNAfragments.A
2-pg amount of EBV DNA (circular or linear) was incubated with 20 U of EcoRI restriction enzyme
on November 10, 2019 by guest
http://jvi.asm.org/
CIRCULAR EBV DNA 13
(Boehringer, WestGermany) in 40 ,ul of 0.1 M NaCl-50mMTris-hydrochloride (pH7.5)-10mMMgC12-2
mMdithiothreitol at370Cfor 2 h. The reaction was stopped by the addition of 10
pd
of 40% Ficoll-0.1 M Tris-hydrochloride (pH 7.5)-50 mM EDTA-0.075% bromophenol blue, and 0.03- to0.6-jIgportions of the cleaved DNA wereapplied to an agarose slab gel (15 by 20 cm). Both 0.35% and 0.8% gels were used. Elec-trophoresis wasperformedfor 20 h and 1.5 V/cm in 40 mM Tris-acetate (pH 8.0)-5 mM sodium acetate-imMEDTA-0.01% ethidium bromide. Gels were pho-tographed under short-wave UV light.Southern blot-tingof the gels (31), hybridization with EBV DNA probes,and fluorography were performed as described
(29).
RESULTS
Isolationof circular EBV DNA. The
stand-ard methods usedfor the enrichment of circular
viralDNAmolecules frommammaliancells,e.g.,
by Hirt precipitation of the host DNA (18),
causeconsiderable losses whenappliedtoDNA
moleculesaslarge asthe EBV genome (2).
Re-cently, several methods involving alkali
treat-ment as an initial step to denature selectively
hostlinear DNA, butnotcovalently closed
cir-cular DNA,were described for the preparation
ofplasmidsfrombacteria, includingDNA
mol-ecules as large as Ti plasmids, F factors, and
cosmidscontaining recombinant DNA (3, 5, 8).
In ourhands,noneofthese methodswere
suc-cessful when directly applied to
EBV-trans-formed lymphocytes. The study of Currier and
Nester (8) on the preparation of Ti plasmids
contains several potentially useful purification
procedures,includinglimited shear treatment of
neutralcelllysatestoallow efficient strand
sep-aration of the host DNA onsubsequent alkali
denaturation, andphenolextractionathigh salt
concentrationto removethe denatured DNA. In
apparent contrast totheresults of thoseauthors,
weobserved that thenecessarysheartreatment
maybeperformedathigh pH withoutcausing
damagetoDNAcircles of the size of
10'
daltons,and this allows cell lysis and shearing to be
carried out directly in an alkaline buffer.
Fur-ther,after neutralization of such lysates,abrief
proteasedegradationstepappears toberequired
before phenol extraction. The procedure
adopted here avoidsasmuchaspossible lengthy
incubations under conditions that may cause
slowbutsignificantintroduction ofsingle-strand
breaks inlargeDNAcircles.
In ourpenultimate purificationstep, circular
DNA was isolated by banding in an ethidium
bromide-CsCl gradient.The band ofcovalently
closedcircularDNA,which isclearlydetectable
by illuminationwithlong-wave UVlightifmore
than109cellsareusedasstarting material, would
beexpectedtocontain all covalently closed
cir-cular DNAmoleculespresentin the cells,
inde-pendent of their size (upto atleast 105 daltons)
and base composition. Examination of such
DNA preparations from several lymphoid cell
linesby electronmicroscopyindicated thatthey
contain a mixture of mitochondrial DNA
(mtDNA) and EBV DNA, but no other circular
DNA molecules. In a typical experiment with
the Raji line, which carries multiple copies of
circular EBV DNA, the relevant ethidium
bro-mide-CsCl fraction contained about 30
mole-cules of mtDNA (average contour length 5.07
,pm)
permolecule of EBVDNA (average contourlength 51.5 ,um). Small amounts of catenated
circular DNA ofcontourlengths of10and15,um
were also observed, which may be ascribed to
thedimers and trimers of mtDNA knowntobe
present (6), and occasional linear DNA
mole-culeswerefound insomepreparations.As afinal
purification step, we employed either density
gradientcentrifugation in CsCl (without
ethid-ium bromide,toseparatethe circular molecules
accordingtobase composition) orglycerol
gra-dientcentrifugation (toseparate onthe basis of
size).Both methods reduced the level of
contam-ination ofthe circular EBV DNAwith mtDNA
toabout onemtDNA molecule perEBV DNA
molecules (Fig. 1). (With the glycerol gradient
centrifugation method, the EBV DNA fractions contain mtDNA enriched for the faster
sedi-menting dimer and trimerforms.) However,we
mainly used centrifugationin CsCl asthe final
purificationstep;since itavoidsanintermittent
dialysis step, the EBV DNA is obtained in a
smallervolume, and circular viral DNA
inad-vertently nicked during and after collection of
therelevant ethidiumbromide-CsClfraction is
recovered. Byacombination ofCsCl and
glyc-erolgradientpurifications, itshould be feasible
toreduce the level of contaminationtoless than
0.05mtDNAmoleculeperEBV DNAmolecule.
The size distributions of circular EBV DNA
from B95-8 andRajicells, isolated onthe basis
of its covalently closedstructure and
guanine-cytosinecontent,weredetermined. Inagreement
withprevious results onglycerol
gradient-frac-tionated material (21), the EBV DNA circles
fromRajicellsappearedtobe ofuniforn length.
Ten molecules were measured, with contour
lengths of 50.5 to 52.2 ym, andanaveragelength
of 51.4,tmwith a standard deviationof1.2%was
observed. In contrast,theB95-8 EBV DNA
cir-cles showed afinite but distinct size
heteroge-neity,althoughtheiraveragelengthwassimilar
to that of Raji EBV DNA. In 11 molecules
measured, contourlengths variedbetween47.6
and 57.7,tm, andan average lengthof52.4,um
VOL. 40,1981
on November 10, 2019 by guest
http://jvi.asm.org/
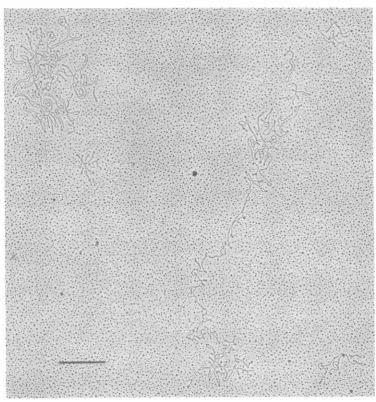
FIG. 1. Electronmicroscopiccharacterization ofpreparations of circular EBV DNA. In a typical field of
apreparation fromRajicells,employingamodified Kleinschmidt technique forspreading,only covalently
closedcircular DNA molecules were observed. Two large circles that correspond to EBV DNA and three small circles that correspondtohuman mtDNA could be seen. The bar in the figure represents 1,um.
with a standard deviation of 5.6% was found. (Raji),sincelinear viral DNA molecules present
Although the Raji and B95-8 circles are of the incells activated to virus production in the
for-samelengthwithinexperimental error,the size mer lines areremoved together with the linear
variation in the B95-8material is significant (P host DNA during the initial stages of
purifica-<0.01) incomparisonwith Raji. This variation tion. Theexistence of circular EBV DNA
mole-is due presumably to the presence of differing cules in the cells of producer lines has been
amounts ofrepeatedsequences atthe site cor- demonstrated previously by treatment of
cul-responding to the ends of the linear molecule tures with inhibitors of viral DNA replication
(seebelow, Fig.2c) and/or at the internal repeat before preparation of DNA circles (7, 14). No
region (4, 12, 29). It is noteworthy that the such pretreatment with inhibitors is necessary
procedure for isolation of circular EBV DNA in thepresentprocedure.
devised here appears to work aswell for virus TheEBV DNA in Rajicells, which is present
producer lines(B95-8) asfor nonproducerlines as 50 to 60copies ofcircular DNA molecules and
14 GRIFFIN ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.69.446.61.464.2]CIRCULAR EBV DNA 15
amuchsmaller amountofintegrated viral DNA
sequences, amounts to about 0.1% of the total
DNAis eachcell(21). Weobtained6to
9,ug
ofcircularE5BVDNAfrom2x 109to3 x109Raji
cells; since each cell contains about6pgofDNA,
thisrepresents ayield of circular EBV DNA of
about 50%.
EcoRI cleavage patterns of DNA circles.
TheintracellularcircularEBVDNApresentin
virus-transforned cells is of similar lengthtothe
linear virion DNA (21), and reassociation
kinet-icsdata (19) andanalysis of restrictionenzyme
fragments by Southernblotting followed by
hy-bridization with radioactively labeled virion
DNA (16, 29, 32) have suggested that the
se-quencecomplexity of the circular DNA is similar
tothat ofvirion DNA.Nevertheless, it hasnot
1 2 3 4
been established whether some of the EBV DNA
circles in a cell are nonidentical, or whether
defectivemoleculescontainingcellular DNA
se-quences(notdetectedby hybridizationwith an
EBV DNA probe) do not also occur. These
possibilities havenowbeenrendered much less
likely by the direct visualization ofthe DNA
fragments obtained by cleavage of circularDNA
preparationswith theEcoRIenzymeand
sepa-ration byagarosegelelectrophoresis. The
frag-ment pattern obtained by EcoRI digestion of
twodifferent preparationsofcircular EBVDNA
from the marmoset B95-8 lymphocyte line is
shown inFig. 2a, together with fragments
ob-tained from linear B95-8-derived virion DNA
and from hostmarmosetmtDNA.Although
cir-cularEBV DNA preparations used stillcontain
1 2 3 4 1 2 3 4
A8- uni
C
-DE=
G
-H
-_m ...
a
J
-m
K-M m
...
_ __
_
_ __
__
_o
*:'
_ _h.
__
__ __
_r
e
em
_b
0
eD
a
b
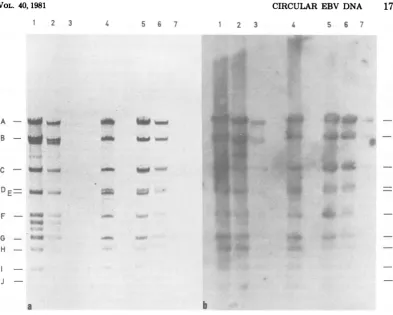
cFIG. 2. EcoRIcleavagepatternof circular EBVDNAfromB95-8cells,asdeterminedbyelectrophoresis in a0.8% agarosegeL Thecapital letters in theleftmarginrefertotheEcoRIfragments of B95-8 linear EB virion DNA(12). Lane1containsmarmosetmtDNAfromB95-8cells; lanes2and3contain two preparations
ofcircular EBV DNAfrom different batchesofB95-8 cells, carriedoutatdifferent times; lane4contains linear EBV DNAfrom virusparticles produced byB95-8 cells. (a) Direct visualization of fragments by
ethidium bromidefluorescence;(b) detectionofEBV DNA sequencesbySouthernblottingandhybridization
withnick-translated32P-labeledvirionDNA; (c)same as(b), but thenick-translated32P-labeledprobe used was acloned EBV DNAfragmentthat containedthe terminal repeat sequences rather thanwhole virion DNA.
VOL. 40,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.50.443.246.565.2]16 GRIFFIN ET AL.
about 1 mtDNA per EBV DNA molecule, in
other respectsthe fragments from circular
ver-sus linear B95-8 EBV DNAs appear virtually
indistinguishable. Possibly, theD,Efragment is
broader and ofslightly higher molecular weight
in thecircular material.Itisclear, however, that
extra bandscorresponding todefective viralor
other DNAsare notseen, evenwhen the method
ofmeasurementdoes notrelyonhybridization
with aselected probe. Moreover, it canbe
as-sumed thatfragments observed in the
prepara-tionsof circular EBVDNA are notdue to
con-taminating virionDNA, sincemorethan98% of
thestarting materialwascircularasdetermined
byelectron microscopy. Inaddition,
hybridiza-tionexperimentswithradioactively labeled EBV
virion DNA after Southern blotting ofthe gel
showed similar patterns for both circular and
linear B95-8 EBVDNAs(Fig. 2b).Asexpected,
thelow-molecular-weight bands in thecircular
DNAswhose positionscoincided withfragments
ofmtDNAdidnothybridize with the probe.
After EcoRI cleavage of linear virion DNA,
oneendof the molecule is knowntobecontained
inthe D fragment. AnEcoRI cleavage site
oc-cursclosetothe otherend, and it is observedas
aseries of smallfragmentsof different sizes since
the terminalsequenceiscomprised ofavariable
numberofcopies ofa 500base-pairrepeat(12).
Ourresults with linear virus DNA (Fig. 2c)are
inagreementwith those data. However,aclear
difference between linear and circular B95-8
DNAs isrevealed byhybridization withacloned
EBV DNAfragment that contains the terminal
sequencesof thelinearmolecules.Inthecircular
EBV DNAmolecules,sequencesrepresentedby
thisprobe occur at theposition of the D
frag-ment, which hybridizes strongly, whereas the
small fragments are not found (Fig. 2c). This
would be theexpected result if thetermini of a
linear molecule werejoined during
circulariza-tionby recombination betweenrepeat sequences
present atboth ends (13, 20,21).Inaddition,a
bandslightly largerthan theDband isobserved
inthecleavedDNAcirclesby hybridization with
the sameprobe (Fig. 2c). Thisrepresents a join
fragment ofthe D and Ibands, present in
sub-molar amounts due to partial resistance to
EcoRIat acleavagesite closetothe leftendof
the linear molecule (L. Rymo, T. Lindahl, S.
Povey,andG.Klein, Virology,inpress).
Similarexperiments with circular EBV DNA
fromRajicellsareshown inFig.3togetherwith
data for the circular andlinear forms ofB95-8
EBV DNA. To obtainbetter resolutionoflarge
fragments,a0.35% agarosegelwasusedinstead
of a 0.8%gel.The data showthat,inthe case of
Raji EBV DNAalso, thefragment patternsas
observedbyethidium bromidefluorescenceare
closelysimilar to thoserevealed byhybridization
withnick-translated 32P-labeledEBVDNAfrom
B95-8 virus particles. In contrast to a recent
report (16), several types ofdefective or
rear-rangedEBV DNA moleculeswere not seen in
thepurifiedcircular viral DNA fromRajicells.
The fragment patterns obtained here are in
agreement with recent mapping data and
hy-bridization studies (4, 16, 26; Rymo et al., in
press), which show that the EcoRI C and D
fragments ofRaji EBV DNA are larger than
those of B95-8 EBV DNA. Thus, the Raji C
band occurs immediately below the B band,
whereas the D band is foundat aposition similar
tothat of the B95-8C band.Hybridization with
the cloned EBV DNA probe containing
se-quences present atthe termini of linear B95-8
DNA yielded hybridization with the Raji D
band, butnot with the A, B, C, or any of the
smaller bands. Asdiscussed elsewhere(Rymoet
al., inpress), partial resistance toEcoRI
cleav-age at two sites generated two submolar
frag-ments. One ofthese, which iscomprised of the
G1 and Lfragments, comigrated with the largest
mtDNA fragment and was only detected by
hybridization (Fig. 3b). The other, which
mi-grated between bands B and C (Fig. 3b), was
comprised of the D and I fragments, since it
hybridized both with a proberepresenting
ter-minalsequences anda cloned EcoRI fragment
Iprobe. Additional bands visually observed in
Raji circular DNA correspond to EcoRI
frag-ments from human mitochondrial DNA. The
latter yields three fragments on cleavage with
EcoRI (10); two of these are present between
the EBV DNA F and G fragments (Fig. 3),
whereas the third is smaller thanthe EBV DNA
Jfragment.
Search for smallamountsofcircular EBV
DNA. The procedurewe employed for
prepa-ration ofcircularEBVDNAdependsonefficient
removal oflinear DNA beforedensity gradient
centrifugation,sothe material used for ethidium bromide-CsCl fractionation is about 200-fold
purifiedwithrespect to circular EBV DNAas
comparedtowholecelllysates.Hybridization of
suchmaterial,orofgradientfractionscontaining
covalently closed circularDNAmolecules,with
an EBVprobeprovides asensitive method for
detecting smallamounts ofcircular viral DNA
molecules in varioustypes ofcells. Control
ex-perimentswithmixturesofRajicells anda
100-foldexcessof Africangreenmonkeykidneycells
indicatedthat onecovalently closedEBV DNA
circle per 5 to 10 cellscouldbe detected (data
not shown). We have used this approach to
search forcircular EBV DNA in the Namalva
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CIRCULAR EBV DNA 17
1 2 3
A - __
4 5 6 7
o k
1 2 3
.: *: ... ...
... ...
:: .. .. .^.:.
...';'.-:';,
:: t.,.sSS :"::...
||
i,C
=l
w ET_....
I
_:i.!
.:t...* | S ::
£' |
*,', W ffi
4 5 6 7
:..
e1zi ..ii
WE~~~~~~~~~i
C - SW wW
DE
F -
--
H-
I-J
[image:7.496.47.441.50.365.2]-a
FIG. 3. EcoRIcleavagepatternsofcircular EBV DNAfromRajicellsand B95-8 cells,asdetermined by
electrophoresisina0.35%agarosegel.Letterscorrespondtobandsobtained withreference B95-8 virionDNA (lane 4).Lanes1, 2, and3 contain cleavedRajicircular EBVDNA, 0.6, 0.3,and0.03pg,respectively. Lane4 contains cleavedB95-8 virionDNA,0.4 pg.Lanes5, 6,and 7contain cleavedcircularEB VDNAfrom B95-8
cells, 0.6, 0.3,and 0.03 pg,respectively. (a)Directvisualization offragmentsby ethidium bromide fluorescence; (b)oneofseveralexposurestodetect EBV DNAsequencesbySouthernblotting and hybridization,asin Fig.
2b. Thephotographshown isalongexposuretoreveal thesmall DNA fragments.
line, an EBV-transformed lymphoid cell line
that contains much less EBV DNA thanmost
other such lines, i.e., only 2 to 3 viral genome
equivalents per cell (1, 25). No sequences
hy-bridizingtoEBV DNAweredetected in circular
DNAfrom Namalvacells, indicatingthat EBV
DNA is not present as covalentlyclosed DNA
circles in this line. Similarly, nocircular EBV
DNAhas beendetected inAW-Ramos, another
EBNA-positive lymphoid cell line that contains
only small amounts of EBV DNA (E. Bjorck,
unpublishedobservations). AW-Ramos has been
shown previously to carry its EBV DNA in
integrated form (2). It now appearslikelythat
thisis also the case forthe Namalva cell line.
Moreover, these data indicate that thepresence
ofcircular EBV DNAmolecules,although
usu-ally found in Burkitt lymphoma lines, is not
obligatory for cellular transformationtooccur.
DISCUSSION
The circular EBVDNAmoleculespresentin
virus-transformed humancells occurclosely
as-sociated with the host DNA. Thus, such EBV
DNAisorganized in nucleosomes (30) andonly
replicates during the S phase (15). In these as well asotherregards, it has properties similarto
anotherplasmid found in eucaryotic cells, the
2-ttmDNAfrom
yeast (24).
Thelarge
size of EBVDNA and its association with host DNAhave
made itdifficulttopreparecircular EBVDNA
exceptby laborious procedures which in the end
yield only minute amounts of material (21).
Thus,manystudiesoncircular DNAhave used
crudepreparations of this material (7, 14, 16, 29,
32). The procedure described here, which is
adapted with important modifications from
sev-eral methodspreviouslyemployed in other
sys-b
40,1981
on November 10, 2019 by guest
http://jvi.asm.org/
18 GRIFFIN ET AL.
temsfor isolation of circularDNA, allows
con-venient isolation of EBV DNA circles in
amountsreadily detected byconventional
opti-cal methods. The only contaminating material
appears to be host mtDNA. Such DNA
prepa-rationsmaybe used forgenetic mapping studies
(Rymo et al., in press) and other experiments
that require access torelatively large amounts
of circular EBV DNA. The procedure should
alsobe well suitedtosearch for smallamounts
ofcircular herpesvirus DNA molecules in
var-iouscell lines and tissue biopsies.
In our experiments, we have used circular
EBV DNAtostudy itscomplexity andcompare
sequenceswith thosefound in virion DNA. The
sequence complexityofsuchintracellular EBV
DNA, as estimated by direct visualization of
restriction enzyme fragments after gel
electro-phoresis,appears tobesimilartothat previously
determined by nucleic acid hybridization (29).
Ouranalysis of the EcoRIfragmentsof circular
EBV DNA indicatesthe virtual absence ofany
marked heterogeneity or defectiveness in the
viral DNA preparationsinvestigated. In this
re-gard, it isinteresting to notethatcircular EBV
DNA fromRaji cells kept in acell bank since
1967 and recently thawed shows an identical
EcoRIcleavagepattem toEBVDNA prepared
from Raji cells grown continuously for many
yearsintissueculture,asestimatedbySouthern
blotting and hybridization (Rymo et al., in
press).Althoughasurveyof theEcoRI fragment
patterns of circular EBV DNA from several
differentsourcesrevealed detectabledifferences
between almost all natural isolates (29), their
overall similarity and the clean fragment
pat-terns observed here (Fig. 2and 3) indicate that
the sequences of intracellular circular EBV
DNAdonotrearrange to amarkedextentduring
continuedproliferation of the hostcells. These
resultsareinapparentconflict withareportby
Helleretal. (16) ofrearrangementand
hetero-geneity within EBV DNA from Rajicells.Those
authors,however, employedcrudepreparations
of viralDNAcontaininganexcessof host DNA,
and the occurrence ofpartialrestrictionenzyme
cleavage productswasnotassessed.
The relationship between the linear EBV
DNA in virus particles and the intracellular
circularandintegratedviral DNA formsremains
unclear. It seems likely, however, that the
cir-cular form is generated by recombination
be-tweenthe terminiof incomingvirion DNA (13,
20, 21). It is noteworthy that the size
heteroge-neity present in B95-8virionDNA (12) also is
observed in B95-8circular DNA. Theabilityof
the hostcelltoreproducefaithfullythe circular
EBV DNA,without generation of shortened or
defective molecules, may be an obligatory
re-quirement forthepreservation of this commonly
occurringlatent form of the virus.
ACKNOWLEDGMENTS
This work was supported by research grants from the Swedish and Danish Cancer Societies andby Public Health Service contract1CP8-1020 within the Virus CancerProgram
of the National Cancer Institute. LITERATURE CITED
1. Andersson,M.1975.Amounts of EBV DNA in somatic cell hybrids between Burkitt lymphoma-derived cell lines. J. Virol. 16:1345-1347.
2. Andersson-Anvret, M., and T. Lindahl. 1978. Inte-grated viral DNA sequences in EBV-converted human lymphoma lines. J. Virol. 25:710-718.
2a.Arrand, J.R.,L. Rymo,J. E.Walsh,E.Bjorck,T.
Lindahl,and B. E. Griffin. 1981. Molecularcloningof thecomplete Epstein-Barr virus genome as a setof
overlapping restriction endonuclease fragments. Nu-cleicAcids Res. 9:2999-3014.
3. Birnboim,H.C., and J.Doly. 1979.Arapid alkaline extractionprocedureforscreening recombinant plasmid DNA.Nucleic Acids Res. 7:1513-1523.
4. Bornkamm, G.W.,H.Delius,U.Zimber,J. Hudew-entz,and M.A.Epstein.1980.Comparisonof
Epstein-Barr virus strains of differentorigin by analysis of the viral DNAs. J. Virol. 35:603-618.
5. Casse, F.,C.Boucher,J.S.Julliot,M.Michel,and J. Denarie. 1979. Identification and characterization of large plasmids of Rhizobium meliloti using agarose gel
electrophoresis.J.Gen. Microbiol. 113:229-242.
6. Clayton,D.A.,R.W.Davis,and J.Vinograd.1970.
Homology and structuralrelationshipsbetween the di-meric and monodi-meric forms of mitochondrial DNA from human leukemicleukocytes.J.Mol. Biol. 47:137-153.
7. Colby,B.J.,J. E.Shaw, G.B.Elion,and J. S.Pagano. 1980. Effect of acyclovir [9-(2-hydroxyalkoxy-methyl)guanine]onEBV DNAreplication.J. Virol. 34: 560-568.
8. Currier,T. C., and E. W.Nester. 1976. Isolation of covalently closed circular DNA of high molecular weight from bacteria. Anal. Biochem. 76:431-441. 9.Davis, R.W.,D. Botstein, and J. R. Roth.1980. A
manual forgenetic engineering. Advanced bacterial ge-netics, p. 184.Cold Spring Harbor Laboratory, Cold
SpringHarbor, N. Y.
10.Drouin,J.1980.Cloningofhuman mitochondrial DNA inE.coli J. Mol. Biol. 140:15-34.
11. Epstein,M.A.,Y. M.Achong,Y.Barr,B. Zajac,G.
Henle,and W. Henle.1966.Morphologicaland
viro-logical investigationsoncultured Burkitt tumor
lym-phoblasts (strainRaji).J. Natl.Cancer Inst.
37:547-559.
12.Given, D.,and E.Kieff.1978.Linkage map of restriction enzyme fragments of the B95-8 andW91 strains of Epstein-Barr virus. J. Virol. 28:524-542.
13. Given, D.,D.Yee,K.Griem,and E. Kieff.1979.Direct repeatsatthe endsofEpstein-Barr virus DNA. J. Virol. 30:852-862.
14. Gussander, E.,and A. Adams.1979.Intracellularstate
ofEBV DNA inproducer cell lines. J. Gen. Virol. 45:
331-340.
15.Hampar, B., A. Tanaka, M. Nonoyama, and J. G. Derge.1974.Replicationoftheresident repressed EBV genomeduring the early S phase of nonproducer Raji celLs. Proc. Natl. Acad. Sci. U.S.A. 71:631-633.
16. Heller, M.,T.Dambaugh,andE.Kieff.1981. Epstein-BarrvirusDNA. IX.VariationamongviralDNAsfrom J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
producer and nonproducer infectedcells.J. Virol.38:
632-648.
17. Henderson,E.,G.Miller,J.Robinson,and L. Heston.
1977.Efficiency of transformation of lymphocytes by
EBV. Virology76:152-163.
18. Hirt,B. 1967.Selective extraction of polyomaDNA from
infectedmouse cellcultures. J. Mol.Biol. 26:365-369. 19.Kawai, Y.,M.Nonoyama, and J. Pagano.1973.
Reas-sociation kinetics forEBVDNA:nonhomologyto mam-malian DNA and homology of viral DNA in various diseases. J. Virol.12:1006-1012.
20. Kintner, C.,and B.Sugden.1979.Thestructureof the termini of the DNA of EBV. Cell17:661-671.
21. Lindahl, T.,A.Adams, G.Bjursell, G. Bornkamm, C. Kaschka-Dierich, and U. Jehn. 1976. Covalently closedcircularduplex DNA of EBV inahuman
lymph-oid cellline.J.Mol. Biol. 102:511-530.
22. Miller, G.,and M.Lipman.1973. Releaseofinfectious EBV by transformedmarmosetleukocytes. Proc.Natl. Acad. Sci. U.S.A. 70:190-194.
23. Nonoyama, M., and J. Pagano. 1972. Separationof
EBV DNAfrom large chromosomalDNA in non-virus
producingcells.Nature (London) NewBiol. 238:169-171.
24. Petes,T. D.1980.Moleculargenetics ofyeast.Annu.Rev. Biochem.49:845-876.
25. Pritchett, R.,M.Pedersen, andE.Kieff. 1976. Com-plexity ofEBVhomologous DNA in continuous lym-phoblastoidcelllines.Virology 74:227-231.
26. Raab-Traub, N.,T. Dambaugh, and E.Kieff. 1980.
DNA of EBV. B95-8, the previous prototype, is an
unusualdeletion derivative. Cell 22:257-267. 27.Reedcman,B.M. and G. Klein. 1973.Cellular localization
ofanEBV-associated complement-fixing antigenin
pro-ducerandnonproducer lymphoblastoidcelllines.Int. J. Cancer11:499-520.
28.Rigby, P.,M. Dieckmann,C. Rhodes,and P. Berg. 1977. LabelingDNAtohigh specific activityin vitroby nick translation withDNApolymeraseI. J. Mol.Biol. 113:237-252.
29. Rymo, L.,T. Lindahl,and A. Adams. 1979. Sites of
sequence variability in EBV DNA from different
sources.Proc. Natl. Acad. Sci. U.S.A. 76:2794-2798.
30. Shaw,J.E., L F.Levinger, and C. W.Carter,Jr.
1979.NucleosomalstructureofEpstein-BarrvirusDNA
intransformedcelllines. J.Virol.29:657-665. 31. Southern, E. M. 1975. Detection ofspecific sequences
amongDNAfragmentsseparated by gel
electrophore-sis. J.Mol. Biol.98:503-517.
32. Sugden,B. 1977.Comparisonof EBV DNAs in Burkitt
lymphoma biopsy cellsand incellstransformed in vitro.
Proc.Natl.Acad. Sci.U.S.A.74:4651-4655.
on November 10, 2019 by guest
http://jvi.asm.org/