0022-538X/80/05-0560/09$02.00/0 Vol.34, No.2
Effect of Acyclovir [9-(2-Hydroxyethoxymethyl)guanine]
onEpstein-Barr
Virus DNA Replication
BRENDA M.COLBY,`* JAMES E. SHAW,' GERTRUDE B. ELION,2 AND JOSEPH S. PAGANO'
CancerResearch Center, DepartmentofBacteriologyandImmunology,andDepartment ofMedicine, School of Medicine, UniversityofNorthCarolina, Chapel Hill,NorthCarolina27514,'andBurroughs
Wellcome Co., Research Triangle Park,NorthCarolina 277092
The effect of acyclovir [9-(2-hydroxyethoxymethyl)guanine] on Epstein-Barr
virus (EBV) DNA replication in thelymphoblastoid celllines P3HR-1 and Raji is reported. Acyclovirat a concentration of
100,uM
completely inhibited EBVDNA synthesis in superinfectedRajicells, butdid notinhibit DNA synthesis in mock-infected cells. The number of EBVgenomeequivalentspercell in the
virus-producing cell line P3HR-1 wassignificantly reduced by acyclovir, whereas the
number of latent EBVgenomesinRajicellswasnotaffected by the drug.Insitu cytohybridization performedonuntreatedP3HR-1cultures revealed thepresence
ofrelatively largeamountsof EBV DNA in 15to20% of the cells. Aftera 100JAM
drugtreatment,no P3HR-1cellscontainedlevels ofEBV DNAdetectableby in
situ cytohybridization. Indirect immunofluorescence studies demonstrated that duringtreatment with 100,uM acyclovir for 7 days, the percentage ofP3HR-1
cellsexpressing viral capsid antigen wasreduced. The EBV DNA remaining in
P3HR-1cellsaftertreatmentwith 100,uM acyclovir (approximately14genomes per cell) had the properties of covalently closed circular DNA with anaverage
molecular weight of 108 x 106, asdetermined bycontourlengthmeasurements.
Acyclovir [ACV; 9-(2-hydroxyethoxymeth-yl)guanine] isone of a new class of potent anti-viral compounds recently shown to be effective againstherpessimplexvirusreplication (4) and is oneofa seriesof synthetic acyclic nucleoside
compounds (28). ACV has a potent inhibitory activity against herpes simplex virus-infected cellsbut has low toxicity for normalcells. The
effect ofACVon another herpesvirus, Epstein-Barrvirus (EBV), has not beenreported
previ-ously.
EBVinfectsallhumanpopulationsand causes
infectious mononucleosis (5, 8, 13), a disease characterizedby infection and transformationof
B-lymphocytes (16, 17, 23) andby EBV-deter-mined nuclear antigen expression (27). EBV is
alsouniquelyassociated with Burkittlymphoma andnasopharyngeal carcinoma,twohuman ma-lignancies. Tissues from such tumor sources carry multiple latent copies of EBV DNA (21, 35-37). Because of its remarkable association with these clinical diseases and its transforma-tion abilities invitro,EBV isconsideredbymany to be a prime human tumor virus candidate. Until now there has been no drug available whichpossesses potent antiviralactivitywithout
havingseriousdeleterious effects on normal cell
replication.
Our studies were designed to determine the effects ofACV on EBV DNA replicationinthe
virus-producing cell line P3HR-1, in the
non-virus-producing line Raji, and in latently in-fectedRaji cells after superinfectionwith P3HR-1 virus. The resultspresented here are similar to those obtained when phosphonoacetic acid (PAA) was used. Our studies show that in the EBV system ACV is an effective inhibitor of viral DNA replication in productively infected cells but is essentially without effect on the
replication of viral DNA in latently infected cells, where cellularcontrol mechanisms appar-ently regulate EBV DNA synthesis.
MATERIALS AND METHODS
Cell cultures. Two Burkittlymphoma-derivedcell lines,Raji (6, 7, 25) and P3HR-1 (15),weremaintained
atbetween2x 105and 106cellspermlby dilution in RPMI 1640 medium containing 10% fetal bovine se-rum.
ACV. 9-(2-Hydroxyethoxymethyl)guanine,
ob-tained from the Burroughs Wellcome Co., was dis-solved inphosphate-bufferedsaline (130mMNaCl,5 mM KCI, 0.01 M sodium phosphate, pH 7.4) at a concentration of 10 mM with the aid of brief
sonica-tion, warmed to 37°C, filtered through a cellulose
acetate membrane (0.45,um; Millipore Corp.), and
storedat-20°C.
Viralantigenproduction.Indirect
immunofluo-rescenceassays for EBVearly antigen(EA) and viral capsidantigen(VCA) (12-14) wereperformed bythe methodof Henle andHenle (12); EA- VCA+ (Kam-pala) and EA'VCA+ (Ghana) sera wereused in the assays.
Preparationof P3HR-1 virus and viral DNA.
560
on November 10, 2019 by guest
http://jvi.asm.org/
Preparation ofvirus forsuperinfection, proceduresfor
superinfection,andpurificationof DNA from P3HR-1virus have been described previously (1,31).
Purification of high-molecular-weight EBV
DNA from Rajiand P3HR-1 cells. Rajicells (8 x
107 cells) and8x 107P3HR-1 cellstreatedwith 100
,uMACVwereharvested, andhigh-molecular-weight
EBV DNAwas purified asdescribed previously (2), with the following modifications. Covalently closed circular EBV DNA was first localized by isopycnic centrifugation in ethidium bromide-cesium chloride;
the DNAwascentrifugedtoequilibriuminaSorvall TV-850 rotor at40,000rpm and 18°Cfor 18 h. The DNA banding at approximately 1.59 g/cm', which
included covalently closed circular EBV DNA (19), was centrifuged again on cesium chloride without ethidiumbromide. The viralDNAbandingat1.718 g/
cm3wasdialyzed against0.1xSSC (1x SSC is 0.15 M sodium chlorideplus0.015 Mtrisodiumcitrate).
Dur-ingstorage at 4°C, approximately 90% of the cova-lently closed circular DNA spontaneously converted
toanopencircular form. EBV DNAisolatedby this
procedurewasused forcontourlengthmeasurements. Preparationofcellular DNA for EBVgenome
number determination. Raji and P3HR-1 DNAs
from mock-treatedanddrug-treatedcultureswere
ex-tracted asfollows. Approximately 4 x 10' cellswere
pelleted, washedonce, andsuspended in
phosphate-buffered salineto5 x 106 cellsperml;0.5volume of Sarkosylsolution (3% Sarkosyl, 50mM Tris, pH 8.6, 15 mM neutralized EDTA) was added, andafter 10
minatroomtemperature,0.5%pronase wasaddedto afinalconcentration of 0.1%. After incubationat37°C
for 2 h, each sample was extracted twice at room
temperaturewithwater-saturated phenol equilibrated with 0.1 volume of 1 M Tris-hydrochloride (pH 8), followed by two extractions with equal volumes of
chloroform-isoamyl alcohol(24:1).DNAand RNA in the aqueousphasewereprecipitatedat -20°C over-nightwith 2.5volumes of salt-saturatedethanol. After
centrifugation at 10,000 x g, the pellet was washed oncewithcold 95%ethanol,dissolved in2ml of0.1x SSC,anddigested with50,ugof pancreatic RNaseper
mlfor 45 minat37°C. Sarkosylandpronase werethen
added to 1 and 0.1% (final concentrations), respec-tively;incubation at 37°Cwascontinued for45min.
Afterincubation, proteinwasremoved bytwophenol
extractions, followed by two chloroform-isoamyl al-coholextractions,andthe DNAwasprecipitated with salt-saturated ethanol overnight at -20°C. Precipi-tates weredissolved in2 mlof0.lxSSC; DNA
con-centrationswereestimated byabsorbanceat260nm. Hybridization.P3HR-1 andRajiculturestreated for 7days with100,uM ACVand mock-treated cultures were analyzed by in situ cytohybridization, as
de-scribed previously (24). Complementary RNA-DNA
hybridization, which was used to quantitate EBV DNA andtolocate EBV DNAonCsClgradients,was
conductedaspreviously described(21).
Electronmicroscopy andcontourlength
mea-surements.Opencircular EBV DNAwasspreadonto
Parlodian-coatedgrids (200-meshcopper; Pelco) bya
modification of the microdiffusion technique (18). In addition to EBV DNA, the spreading solution
con-tained 0.50Mammoniumacetate (pH7.5)and60,ug
of cytochromecper ml. Simian virus 40 (SV40) form II DNA wasincludedas aninternal size marker.
Contour lengths of open circular DNA molecules weredetermined by projectingelectron micrographs ontoaHewlett-Packard 986Adigitizertablet and
trac-ing theDNAimages witha cursorelectronically
cou-pled to aHewlett-Packard9825calculator. RESULTS
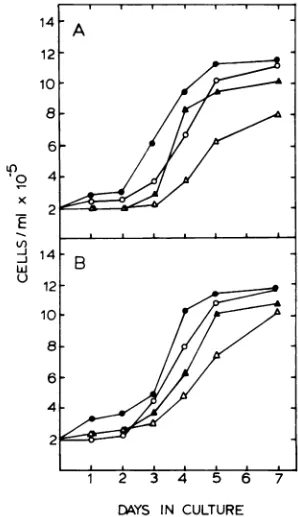
Effects of ACV on thegrowthof P3HR-1
andRajicells. The growthcurves of
lympho-blastoid cell lines cultured in the presence of variousconcentrationsofACV are shown inFig.
1. Cellswerepelletedandsuspendedto2 x 105 cells per ml in fresh media on day 0. A lag in
growthwas evident forallcultures after
resus-pension; however, for both celllines theperiod ofthe lag increased with increasing ACV con-centration. By day 7 the cell densities were
identical in mock-treated cultures and cultures
treatedwith 100
liM
ACV.Cultures treated with higherACV concentrationsdidnotattain mock-treated cell densities after7days.Superinfection ofRaji cells in the pres-enceof ACV. Superinfection of Raji cells with
14 A
12-10
8
6
-'0
4-LI)
J14-w B
U
12-10.
8 6
4-2
1 2 34 56 7
DAYS IN CULTURE
FIG. 1. Effect of ACVon thegrowthrate of cul-turedlymphoblastoidcelllines. The cell lines P3HR-1 (A)andRaji(B)weregrownat37°Cwithout ACV (-)orwith ACVatconcentrationsof100p.M(0),500
p,M (A), or 1,000 u.M (A). Cells were collected by centrifugationandresuspendedin thesamevolume
offreshmedium withorwithout ACV every48h.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.274.423.325.584.2]562 COLBY ET AL.
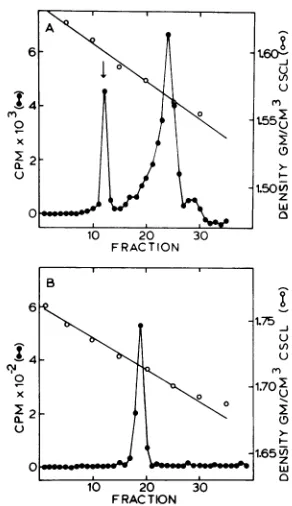
P3HR-1 virus leads to replication of EBV DNA, tosuppression of cellular DNA synthesis, and to fragmentation of cellular DNA (22, 31, 34). Such results are shown in Fig. 2A; superinfection of Raji cells with EBV inhibited host cell DNA synthesis, whereas EBV DNA was synthesized. DNA synthesis was measured as the number of counts incorporated into the DNA banding at 1.718g/cm3,the density ofviral DNA. Addition of 100 MMACV to the cultures 1 h after super-infection caused complete suppression of EBV DNA synthesis (Fig. 2A). It is of interest that hostcell DNA was not synthesized even in the presence of ACV. Mock-infected cells not treated with ACV and mock-infected cells
A
2-0
2 B
2
10 20 30
[image:3.503.62.246.76.487.2]FRACTION
FIG. 2. EffectofACVonEBV DNA synthesis in
superinfectedandmock-infected Rajicells. (A)
Neu-tral CsCl density profile of[32P]DNA synthesized during superinfection without ACV (0) or in the presenceof100pLMACV(0). (B)NeutralCsCldensity profileof[32PJDNA synthesized in untreated
mock-infectedcells (0) and inmock-infectedcells treated with 100p.MACV(0).Thearrowsindicate the posi-tion of1.718g/cm3 (density increasesfrom rightto
left). Eachprofilerepresents the label incorporated
into 106 cells. A totalof106Rajicells werepelleted andsuspendedin 0.3 mlofvirussuspensionatzero time.After1hat37°C,the cellswerepelleted,washed
twice,andsuspendedinphosphate-freemedium (min-imalessential mediumminusphosphateand
contain-ing2%dialyzedfetalcalf serum).At 9 hpostinfection 200ILCiof 32pwasadded,and incubationwas contin-uedfor24h.
treated with 100MMACVsynthesized only cell DNA (Fig. 2B). On Fig. 2B the position of 1.718
g/cm'
asdetermined by refractive index is indi-cated.The effects of various ACV concentrations on inhibition of EBV DNA synthesis were deter-mined. Rajicells were superinfected in the pres-enceofvarying concentrations of ACV, and the percent inhibition of EBV DNA synthesis at each drug concentration was measured. Inhibi-tionvalues obtained for 0, 0.1, 1, 10, and 100 yM ACV were 0, 20, 9, 70, and 100%, respectively. Percent inhibition values (Fig. 3) were calculated
from the following: [1 - (counts per minute incorporated into EBV DNA of 106
superin-fected Rajicellsat aparticular drug
concentra-tion/countsper minute incorporated into EBV DNA of 106 superinfected Raji cells)] x 100. Figure 3 shows that the dose required for 50%
inhibition
(ED5o)
of viral DNA synthesis was 7,uM.
Reduction of viral genomes in P3HR-1 cellsby ACV. Figure4shows theeffect of ACV onviral genomes in P3HR-1 and Raji cells
cul-tured for7days in the presence of varying drug concentrations. As determined by EBV comple-mentary RNA-DNA membrane hybridization
(Table 1), the numberofEBV genome
equiva-lents per cellin the producer cell line P3HR-1
decreased with increasing ACV concentration from 133 genomes per cell to 14 EBV genome equivalents percellin 72 to 96 h atthe maximum
drugconcentration (100
MuM).
Atthesamedrug concentrations, the numbers of EBV genomes ofRajicellswere notaffected.The
ED;o
calculatedfora50% reduction in the average genome num-ber for P3HR-1 cellsby ACVwas6
MuM.
The effect of drug removal on the number of EBVgenomeswasdetermined.After drug treat-mentfor 7 days, all cultures weresuspended in
fresh medium lacking ACV. At 14 days after
drug removal, the number of EBV genome
equivalentspercellinallcultures had returned
tothecontrol levels(Fig. 4).
Complementary RNA-DNA
cytohybridi-zationinsitu of P3HR-1 andRaji cells.The
cytohybridizationtechnique,asdescribedabove, wasusedtodetermine theproportionofcellsin mock-treated cultures and in cultures treated with 100
MuM
ACV whichwere harboring EBV DNA. After hybridization with EBV-specific complementary RNA, cells from mock- and ACV-treated Raji cultures demonstrateda lowbackgroundofgrainsdiffuselyscatteredoverall of thecells.Dense accumulations ofgrainswere
present over approximately 15 to 20% of the mock-treated P3HR-1 cells, which is in agree-ment with previously
published
data (24). In contrast,noACV-treatedP3HR-1 cellsdemon-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
IT
3-2 \ 40 Z
b2
u~~~~
60co .60 __0 z
1 0
0 0.1 1 10 100
[image:4.503.53.246.53.235.2]CONCENTRATION (pM)
FIG. 3. Inhibitionof EBVDNA synthesis in
super-infected Raji cells treated with varying
concentra-tions ofACV. The value obtainedfor each ACV concentration is thesumofthe32Pcountsperminute incorporated into EBV DNA (1.718 g/cm3) when
an-alyzedasdescribed in thelegendtoFig.2. Percent inhibition valuesweredefinedasfollows: [1-(counts
perminuteincorporatedintoEBVDNAof106
super-infected Raji cells at aparticular drug
concentra-tion/countsperminuteincorporatedinto EB VDNA of 106superinfectedRaji cells)]x100.
0 0.1 1 10
ACV CONCENTRATION(jiM) 100
FIG. 4. Effect of ACV removal on EBVgenome
equivalents in Raji and P3HR-1 cells. P3HR-1 (0) andRaji (0) celllinesweretreatedfor 7days with ACVatvarying concentrations. On day7the number
ofgenomes per cell was determined at each drug
concentration. Theremaining cells were harvested
andsuspended in drug-free medium, and numbers of genomes percellfor P3HR-1 (x) and Raji (T)cells were determined onday22(14 days afterACV
re-movalfrom thecultures).
strated dense accumulation ofgrains. The scat-ter ofgrains was uniform over all cells and the
background. Because the sensitivity of in situ
cytohybridization is rather low (approximately
60 genomes per cell), these results indicate a very highnumberof EBV genomes in15 to20% of themock-treated P3HR-1 cells and show that the number of genomes in the ACV-treated P3HR-1 cultures wasbelow the level of detect-ability.
Expression of viral antigens in P3HR-1
and Raji cells. Indirect immunofluorescence studies demonstrated that the percentage of P3HR-1 cells expressing VCA was reduced by
drug treatment (Fig. 5). Staining with serum which containedboth EA and VCAantibodies,
however, revealed no change after ACV treat-ment. This observation mustbe attributableto the continued expression ofEA' cellsin drug-treated cultures. Most cultures exhibited a de-crease inthe percentageofVCA+ cellsas early
as 24h; however,somecells continuedtoexpress
VCA after 7 days, a time when EBV DNA
synthesis in P3HR-1 cells was maximally in-hibited. The percentage ofVCA+ cells in most
drug-treatedcultures increased to thelevel pres-ent before drugtreatment within 15 days after drug removal. Raji cultures treated for 7 days
with ACV concentrations ranging from 1 to1,000
,uM
did not exhibit an inductionofeither EA or VCA.TABLE 1. EBV-specific[3HJcRNA hybridizedto
P3HR-IandRajicell DNA
cpmx10'hybridized
Celltype ACVconcn (,uM) per50pgofDNA" Day 7h Day226
P3HR-1 0 24.8 26.4
0.1 21.7 28.9
1.0 18.0 29.9
10.0 7.7 26.8
100.0 3.1 28.9
Raji 0 8.0 11.4
0.1 11.1 12.3
1.0 11.5 11.4
10.0 10.4 11.7
100.0 13.7 12.8
Average hybridization values of duplicate DNA
filters, each standardized to 50,tg of DNA perfilter.
Counts per minute bound by calf thymus (70 cpm) andHEp-2 DNA (100 cpm) were subtracted as back-ground.
hHybridization was conducted as described
previ-ously (21) with DNA from P3HR-1 and Raji cells maintained for 7 days in the presence of various ACV concentrations. On day 7 the remaining cells were
pelleted, washed, and placed in drug-free medium; genome levelsfor the same cultures were determined onday22.
;k
,)
140-z
w120
-j
.100
0
w
80.
-60 zw 40.
ED
20
LI
VOL.I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.255.448.391.538.2]564 COLBY ET AL.
[image:5.503.59.252.47.253.2]15 DAYS
FIG. 5. Effect ofACVonVCAexpressionin
P3HR-I cells. Cells weretreated for 7days with ACV at concentrations ofI uM (0), 10pM(A), 100pM(A), 500 pM (O1), and 1,000ptM (x). Mock-treated cells
(0)werealsoassayed forVCAexpression.Cellswere harvested and suspended in drug-free medium on day 7(arrow); the culturesweremonitoredforVCA untilday22.
Characterization of EBV DNAremaining
inP3HR-1 cells afterdrugtreatment.After 10 days, some EBV DNA remained in ACV-treatedP3HR-1cells.The EBV DNAremaining
was characterized by isopycnic centrifugation and electron microscopy. Figure6Ashowsthat asignificantquantityoftheACV-resistant viral
DNAbandedattheposition expectedfor
cova-lently closed circular molecules (1.59 g/cm3) when analyzed by ethidium bromide-cesium chloride density gradient centrifugation. Some EBVDNA banded between 1.56 and 1.57g/cm:,
theposition ofopen circular and linear DNAs. The EBV DNA from thepeakrepresenting
co-valently closed circular DNA was isolated and fractionatedonasecond cesium chloridedensity gradient without ethidium bromide (Fig. 6B). This DNA rebanded as a single peak at the
bouyant density expected for EBV DNA; the
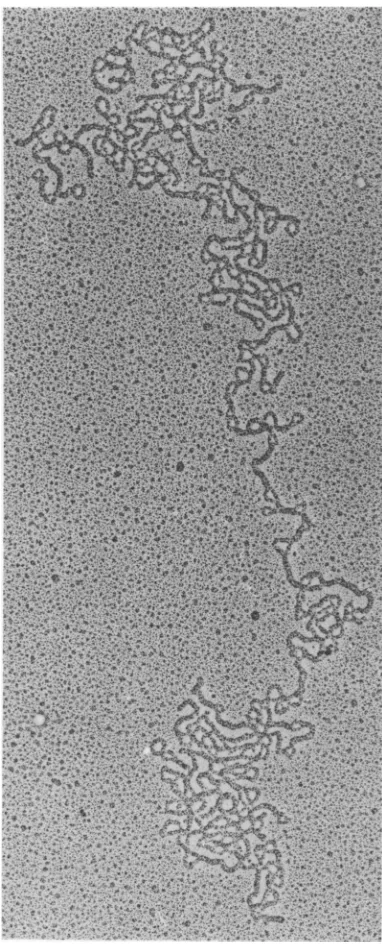
DNAfrom thispeakwasisolatedandvisualized by electronmicroscopy (Fig. 7and8). Figure7
shows a covalently closed circular supercoiled
DNA molecule recovered from drug-treated P3HR-1cells,andFig.8 showsanicked circular
DNAmolecule isolated from thesame prepara-tion.
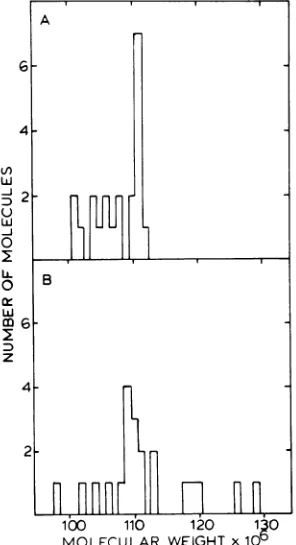
Analysis of EBV DNA by electron mi-croscopy. Contour lengths and corresponding
molecular weights were determined for 22
cir-cular DNA molecules isolated from Raji cells
and 21 circular molecules isolated from
drug-treated P3HR-1 cells. The contour length of EBV DNA was determined relative to that of SV40 form IIDNA. The mean lengths of 10 to 15 SV40 DNA molecules (standard deviation, 1.5%) which were measured within the same frameasthe opencircular EBV DNA molecules wereused to calculate the molecular weight of EBV DNA. Themolecular weight ofSV40DNA
(3.4x 106) was calculated for these studies from a precise nucleotide sequence analysis ofSV40 DNA (9, 26). The mean values obtained for the molecularweights of EBV DNAs from Raji and ACV-treated P3HR-1 cells were 111 x 106 ± 1.8 x
106
and 108 x 106 ± 2.1 x 106, respectively. Thedistribution of molecular weights is shown inFig. 9.DISCUSSION
ACV has littlecytotoxicity forlymphoblastoid cells atthe concentration(100,uM) necessary for complete inhibition of EBV DNA synthesis. A slight delay in cellreplication is noted for Raji and P3HR-1cellsimmediatelyaftersuspension in medium containing 100 yM ACV; however,
6 1.60-:
jo~~~~~~~~~~_ Ul
02t5503
10 20 3
1.50W z w O
10 2 30
FRACTION
B 6
-1.75 LI)
0
-~~~~~~~~1.70
2z
0_0 0% w
10 20 30
FRACTION
FIG. 6. Ethidiumbromide-cesiumchloride(A)and
cesium chloride(B)densitygradientcentrifugations ofcovalentlyclosed circular EBVDNAremainingin P3HR-Icellsaftertreatmentwith 100 LMACV. EBV
DNAwaslocalizedbyhybridizationacrossthe
gra-dientswithEBV-specific complementaryRNA. The
arrow in (A) indicates the position of covalently
closed circular DNAwhich rebandsin neutralCsCl (B)with abouyant densityof1.718g/cm3.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.285.431.309.562.2]FIG. 7. Electronmicrograph ofacovalently closed
circularEBVDNA molecule isolated fromP3HR-1 cellstreatedwith100,M ACV.x54,000.
the cell densitiesreturn tocontrol levelswithin 7 days in the continued presence of the drug
(Fig. 1). Furthermore, in ourlaboratory P3HR-1 cells have been grown in medium containing
100,iMACVformorethan1year.
ACVcompletely inhibits EBVDNA synthesis in superinfected Rajicells andreduces the
av-erage number of EBV genomes per cell in P3HR-1cultures to a maximum level of 14 in 72 to96 h. Incontrast, EBV DNA synthesis in Raji cells is not affectedby ACV; the number of genomes percell remainsunchanged foratleast 2 weeks in its presence. It has been reported previously that the number of latent EBV genome equiva-lents in P3HR-1 cells isapproximately 11 after cells are treated withcycloheximide (33) or PAA (34).The close agreement between these results and ours implies that ACV inhibits only the productive replication of EBV.
Insitucytohybridization performedon P3HR-1 cells reveals the presence ofsignificant levels of EBV DNA in 15 to 20% of the population. After drug treatment, no cells contained levels of EBV DNA detectableby thetechniqueof in situ cytohybridization. This is consistent with ourdata, which demonstrate that approximately 14 EBV genomes per cell remain in P3HR-1
cultures treatedwith 100lOMACV, a level well below the limits ofdetectability of
cytohybridi-zation. EAsynthesiscontinuesnormally despite
the fact thatmostcultures exhibitadecrease in
the percentageofVCA+ cellswithin 24 h after drugaddition.
These results are comparable to those ob-tained by treating Raji, P3HR-1, and
superin-fectedRaji cells with PAA (29, 32-34), although the effective dose of PAAwassevenfoldgreater than the concentration of ACV used in these studies.
AsACV and PAA inhibitonlytheproductive
replicationofEBV, they may have similar mech-anisms ofaction,eventhough thesetwoantiviral compounds arestructurally dissimilar. It will be
interesting to determine whether ACV or its
phosphorylated derivatives inhibit EBV DNA
replication by bindingto a virus-specific DNA
polymerase,ashasbeensuggestedfor the mech-anism of action of PAA in the herpessimplex
virus (HSV) system (20), orwhether it inhibits EBV DNA synthesis by chain termination. ACV,anacyclic nucleoside analog of guanosine, is phosphorylated in HSV-infected cells to mono-, di-, and triphosphates (4). ACV is also
phosphorylatedin superinfected Raji cells, but the level of phosphorylation is considerably lower than that observed inHSV-infectedcells
(unpublished data). The formation of ACV monophosphateinHSV-infected cells isaresult of virus-induced thymidine kinase activity in thesecells (4). A similar enzyme which
phospho-rylates ACV in the EBV system has not been found (unpublished data).
Different DNA polymerases may replicate EBVDNAin productivelyinfected and latently infected cells. ACV has no inhibitory effect on theincorporation of label intoDNAinRajicells
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.55.246.67.539.2]566 COLBY ET AL.
ip!~~~~~~~~~~~~~~~~~~~~~~
: 8. Electronmicrographof an opencircular EBV DNA molecule after spontaneous conversion from entlyclosedcircularform. x15,000. Theopen circularform ofSV40 was included as an internal size
nce.
(Fig.2B).Perhaps EBV DNA synthesis in Raji
A
cells
is resistant to ACV because DNAreplica-tion is dependent
on host ratherthan viral DNAG polymerases.
After superinfection, host DNA synthesis in
ACV-treated Raji cells is inhibited. Inhibition
4 occurs eventhoughACV blocks the replication
ofEBV DNA(Fig. 2A). This result suggests that
en the inhibition of host DNA synthesis during
Lii superinfection is an early event which precedes
D 2 l l n r thesynthesis of viral DNA. This hypothesis is
lll % UU 1 consistent with previous findings which show
o that bothevents occur at about the same time
E
.(22).
Anotherpossibility
is that ACV inhibitsO B hostDNAsynthesis in superinfectedRajicells.
a: TheED50for the replication of EBV DNA in
w
superinfected Raji cells was 7
tiM.
This valuev agrees well with 6 ,uM, the concentration
re-z quired for a 50% reduction in the average
ge-4 nome number of P3HR-1 cells. ED50 values for
HSV type 1 (based on a 50% reduction in infec-tious
titer,
asdeterminedby
plaque assay)
are2 0.1
,lM
in Verocells
(4)
and0.7MuM
inHeLa cells(3). The variation indose response observed for
r ACV in the HSV and EBV
systems
could beattributed to the different cell systems and
00
b10
H 20I30
methodsusedtomeasure theED50.
Insupport
MOLECULAR WEIGHT x
100
of this idea is the recentreport that the ED50 for 9. Size distribution of circular EBV DNA HSV DNA replicationidi
infected Vero cells is ulesisolatedfromP3HR-1 (A) and Raji (B) cell approximately 1,uM (10),avalueslightlylower than that observed byusfor EBV DNArepli-FIG
covalE
refere
FIG molec lines.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.62.453.67.303.2] [image:7.503.83.231.352.625.2]cation (determined bythesamemethod).
A significant quantity of the EBV DNA
re-maining in P3HR-1 cells after drugtreatmentis covalently closed circular DNA. This possibly
representsthenucleosomal EBV DNA recently foundinP3HR-1 cells (30). Theviral episomes haveanaveragemolecular weight of 108 x 106,
asdetermined bycontourlengthmeasurements.
This isclosetothe molecular weight( 111 x 106)
whichweobtained for theepisomal DNA of the
non-virus-producing cell line Raji. A previous estimatefor themolecular weight of EBV DNA fromRaji cellswas106x10' (19). However, this
measurementwasdetermined by priorselection
of DNA from glycerol gradients. We mayhave
detected the larger forms of molecules in Raji cells because our isolation procedure did not
preselect covalently closedEBVDNAmolecules
onthebasis of size. Thepresence ofcovalently
closedcircularEBVDNA withan average
mo-lecular weight of 100 x 106 in PAA-treated P3HR-1 cells has been reported recently (11). Our molecularweightvaluesarebasedonSV40
DNA astheinternal size reference. The values
reported previouslyforRajiand P3HR-1 circu-lar viral DNAswere determinedby usingPM2
DNAasthe sizereference.
In conclusion, ACV inhibits the productive replication of EBV DNA but has no apparent
effect on the latent EBV genomes in cultured lymphoblastoid cell lines. Consistent with this
fact, it has been possible to demonstrate the
presenceofACV-resistantcovalentlyclosed
cir-cular EBV DNA in the P3HR-1 cell line.
Be-causeof its lowcytotoxicity,ACV isauseful tool
for theelucidation of cellular and viralprocesses
related to EBV replication and to viral DNA persistenceinlatentlyinfectedcells. Itspotential
as a clinically useful antiherpetic agent is
cur-rently underassessment.
ACKNOWLEDGMENTS
Wethank Marie Harris and Claire Moore forexpert tech-nicalassistance, Werner Henle forVCA+serum,JackGriffith
andClaire Moore forSV40DNA andadviceregarding electron
microscopy, Phil Furmanfor valuablediscussion, and Fumi Bostic forsecretarialassistance.
Thisinvestigationwassupported by Public Health Service grant5-POI-CA19014fromtheNational Cancer Instituteand
byagrantfrom theBurroughs Wellcome Co.
LITERATURE CITED
1. Adams,A. 1975.Preparation of Epstein-Barr virus from P3HR-1 cells andisolation of virusDNA,p.129-146.In
D. V.Ablashi, H. G. Aaslestad, and G. de The (ed.),
Epstein-Barr virus, production, concentration, and
pu-rification.InternationalAgency for ResearchonCancer, Lyon, France.
2. Andersson-Anvret, M., and T. Lindahl. 1978. Inte-grated viral DNAsequencesinEpstein-Barr virus-con-verted humanlymphoma lines. J. Virol. 25:710-718.
3. Collins,P.,and D.J. Bauer.1979. Theactivityinvitro
against herpes virus of
9-(2-hydroxyethoxymeth-yl)guanine(acycloguanosine),a newantiviral agent. J. Antimicrob. Chemother.5:431-436.
4. Elion, G.B.,P. A.Furman,J. A.Fyfe,P.deMiranda, L.Beauchamp, andH. J.Schaeffer.1977.Selectivity of action of anantiherpeticagent, 9-(2-hydroxyethoxy-methyl)guanine. Proc. Natl. Acad. Sci. U.S.A. 74:5716-5720.
5. Epstein,M.A.,and B.G.Achong.1977.Recent progress in Epstein-Barr virus research. Annu. Rev. Microbiol. 31:42 1-445.
6. Epstein,M.A.,B.G. Achong,and Y. M. Barr. 1964. Virusparticles in cultured lymphoblasts from Burkitt's lymphoma. Lanceti:702-703.
7. Epstein,M.A., B. G.Achong,Y. M.Barr, B.Zajac,G. Henle,andW.Henle. 1966.Morphologicaland viro-logicalinvestigations on cultured Burkitt tumor lym-phoblasts (strain Raji). J. Natl. Cancer Inst. 37:547-559.
8. Evans,A.S.,and J. C.Niederman.1976.Epstein-Barr virus, p. 209-223. In A. S.Evans (ed.), Viral infections in humans. PlenumMedicalBookCo.,New York. 9. Fiers, W.,R.Contreras, G.Haegeman,R.Rogiers,A.
VandeVoorde, H.Han Heuverswyn,J. Van Her-reweghe, G. Volckaert, and M. Ysebaert. 1978. Complete nucleotidesequence ofSV40DNA. Nature (London) 273:113-120.
10. Furman, P. A., M. H. St. Clair, J. A. Fyfe, J. L. Rideout,P. M.Keller,and G. B. Elion. 1979. Inhi-bition ofherpessimplexvirus-induced DNApolymerase activity and viral DNA replication by 9-(2-hydroxy-ethoxymethyl)guanine and its triphosphate. J. Virol. 32:72-77.
11. Gussander, E.,and A.Adams.1979.Intracellular state ofEpstein-Barr virus DNA inproducer cell lines. J. Gen. Virol. 45:331-340.
12. Henle, G.,and W.Henle. 1966.Immunofluorescencein cells derivedfromBurkitt's lymphoma.J.Bacteriol.91: 1248-1256.
13. Henle,G., andW.Henle. 1970.Observationson child-hood infections with theEpstein-Barr virus. J. Infect. Dis. 121:303-310.
14. Henle, W., G. Henle, B. Z. Zajac, G. Pearson, R. Waubke,and M. Scriba. 1970.Differentialreactivity ofhuman serums with early antigensinducedby Ep-stein-Barrvirus. Science 169:188-190.
15. Hinuma, Y., and J. T. Grace. 1967.Cloningof immu-noglobulin-producinghuman leukemicandlymphoma cells inlong-termcultures. Proc. Soc. Exp. Biol. Med. 124:107-111.
16. Jondal, M., and G. Klein. 1973. Surface markerson
human B and Tlymphocytes.II. Presence of Epstein-Barr virus receptors on Blymphocytes.J. Exp. Med. 138:1365-1378.
17. Kieff, E.,and J. Levine. 1974.Homologybetween Burk-ittherpesviral DNA and DNA in continuous lympho-blastoid cells frompatientswith infectious mononucle-osis. Proc.Natl. Acad. Sci. U.S.A. 71:355-358. 18. Lang, D.,and M. Mitani. 1970.Simplified quantitative
electron microscopy ofbiopolymers. Biopolymers 9: 373-379.
19. Lindahl, T.,A.Adams, G. Bjursell, G.W.Bornkamm, D.Kaschka-Dierich, and U.Jehn. 1976. Covalently closed circularduplexDNA ofEpstein-Barrvirus in a humanlymphoidcell line. J. Mol. Biol. 102:511-530. 20. Mao, J. C.-H., E. E. Robishaw, and L. R. Overby.
1975. Inhibition of DNApolNmerasefrom herpes sim-plex virus-infected Wi-38 cells by phosphonoacetic acid. .J.Virol. 15:1281-1283.
21. Nonoyama, M., and J. S.Pagano. 1971. Complemen-tary RNA specific to the DNA of the Epstein-Barr virus: detection ofEpstein-Barr viral genome in non-productivecells. Nature (London)New Biol. 233:103-106.
22. Nonoyama, M.,and J.S.Pagano. 1972.Replicationof
on November 10, 2019 by guest
http://jvi.asm.org/
viral deoxyribonucleic acid and breakdown of cellular deoxyribonucleic acid in Epstein-Barr virus infection. J. Virol. 9:714-716.
23. Pagano, J. S. 1974. The Epstein-Barr viralgenomeand
its interactions with human lymphoblastoid cells and chromosomes, p.79-116. InE. Kurstak and K. Mara-morosch (ed.), Viruses, evolution andcancer.Academic PressInc., New York.
24. Pagano, J.S., and E.-S. Huang. 1974.The application of RNA-DNA cytohybridizationtoviral diagnostics,p.
279-299. In E. Kurstak and R. Morrisset (ed.), Viral immunodiagnosis. Academic Press Inc., New York. 25. Pulvertaft, R. J. V. 1965. A study of malignanttumors
in Nigeria by shorttermtissue culture. J.Clin. Pathol. 18:261-273.
26. Reddy, V. B., B. Thimmappaya, R. Dhar, K. N. Sub-ramanian, B. S. Zain, J. Pan, P. K. Ghosh, M. L. Celma, and S. M. Weissman. 1978. Thegenomeof simian virus 40. Science 200:494-502.
27. Reedman, B. M., and G. Klein. 1973. Cellular localiza-tion ofanEpstein-Barr virus (EBV)-associated comple-ment-fixing antigen in producer and non-producer lym-phoblastoid cell lines. Int. J. Cancer 11:499-520. 28. Schaeffer, H. J., L. Beauchamp, P. de Miranda,G.B.
Elion, D. J. Bauer, and P. Collins. 1978. 9-(2-Hy-droxyethoxymethyl)guanine activity against viruses of theherpesgroup.Nature(London) 272:583-585.
29. Seebeck, T., J. E. Shaw, and J. S. Pagano. 1977. Synthesis of Epstein-Barr virus DNA in vitro: effects of phosphonoacetic acid, N-ethylmaleimide, and ATP. J.
Virol. 21:435-438.
30. Shaw,J.E., L. F. Levinger, and C.W.Carter, Jr. 1979.Nucleosomalstructure of Epstein-Barr virus DNA in transformed cell lines. J. Virol. 29:657-665. 31. Shaw, J. E., T. Seebeck, J.-L.H.Li, and J. S. Pagano.
1977. Epstein-Barr virus DNAsynthesizedin superin-fectedRajicells.Virology77:762-771.
32. Summers, W. C., and G. Klein. 1976. Inhibition of Epstein-Barr virus DNA synthesis and late gene expres-sionbyphosphonoacetic acid. J. Virol.18:151-155. 33. Tanaka, A., M. Nonoyama, and B. Hampar. 1976.
Partial elimination of latent Epstein-Barr virus ge-nomes from virus-producing cells by cycloheximide. Virology70:164-170.
34. Yajima, Y., A. Tanaka, andM.Nonoyama. 1976. In-hibition of productive replicationofEpstein-Barrvirus DNAby phosphonoaceticacid.Virology71:352-354. 35. zur Hausen, H., V. Diehl, H. Wolf, H.
Schulte-Holthausen, and U.Schneider. 1972.Occurrence of Epstein-Barrvirus genomes in humanlymphoblastoid cell lines.Nature(London)New Biol. 237:189-190. 36. zur Hausen, H., and H. Schulte-Holthausen. 1970.
Presence of EBvirus nucleic acid homologyin a "virus-free" lineofBurkitttumourcells. Nature(London)227: 245-248.
37. zurHausen, H., H. Schulte-Holthausen, G. Klein, W. Henle, G. Henle, P. Clifford, and L. Santessen. 1970. EBV DNA in biopsiesof Burkitt tumours and anaplastic carcinomas of the nasopharynx. Nature (London)228:1056-1058.