0022-538X/85/070140-07$02.00/0
Copyright © 1985, American Society for Microbiology
Nucleotide
Sequence and
Structural
Features
of
a
Novel
Us-a
Junction
Present in
a
Defective
Herpes
Simplex Virus
Genome
EDWARD S. MOCARSKI,' LOUIS P. DEISS,2 ANDNIZA FRENKEL2*
Department ofMedicalMicrobiology, Stanford University School of Medicine, Stanford, California
94305,1
andDepartment
of
MolecularGenetics andCellBiology,
TheUniversity
of
Chicago,
Chicago,
Illinois 606372 Received 2 January1985/Accepted 8April 1985Defective genomes generated during serial propagationof herpes simplex virus type 1 (Justin) consist of tandem reiterations ofsequencesthat arecolinear with a portion of theS componentof the standard viral genome.Wedetermined thestructureof the novel
Us-a
junction,atwhich the Ussequencesofonerepeatunit join theasequencesof the adjacentrepeatunit.Comparison of the nucleotide sequenceatthisjunction with the nucleotide sequence of the correspondingUs
region of the standard virus genome indicated that the defectivegenomerepeatunitarosebyasinglerecombinationaleventbetweenanL-Sjunctionasequenceandthe Us region. Therecombinational process might have been mediated by limited sequence homology. The sequencesretained within theUs-ajunction further define the signal for cleavage and packaging of viral DNA. The herpes simplex virus (HSV) DNA genome is
com-posed of two components, L and S, each consisting of uniquesequences (UL and Us)bracketedbyinvertedrepeats inthesequencearrangement ab-UL-b'a'c'-Us-ca. The Land Scomponents invert relative toeach other,generating four equimolar isomeric forms of standard virusDNA(reviewed in reference 23). The inversions appear to involve site-specific recombination between inverted copies of the a sequence (17-20, 24).
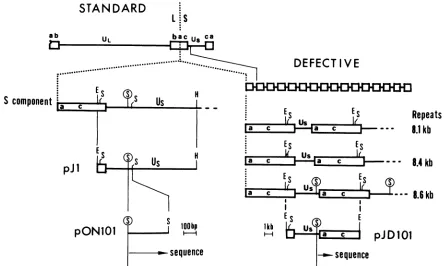
Previousanalysesof serially passaged stocks of HSVtype 1 (HSV-1) (Justin) revealed the presence of defective genomes composed of head-to-tail reiterations of simple subsets ofthe standard viral DNA sequences (5, 7, 8, 13). Three families ofrepeat units (see Fig. 1) differing in size were recognized in thesedefective viral DNA preparations (13). All three types of repeat units contained contiguous setsofsequencescorrespondingtotheDNAsequences that constitute the right terminus of the viral genome (when displayed in the prototype orientation). These sequences included the entire ca inverted repeat as well as various amountsoftheadjacentUs sequences. The threefamilies of repeatunits hadacommon structureinwhichthe asequence waslinked to aregion inUsthrough anovel
Us-a
junction. Defective virus genomes with a similar general structure (class I defective genomes) have been characterized in stocks of other HSV-1 and HSV-2 strains after undiluted propagation inculture(reviewed in reference 5).Inthe presence ofhelper virus DNA, monomeric repeat units of defectivegenomes as well as constructs derived by cloning ofthecorrespondingstandard virus DNAfragments canbepropagatedin virus stocks as concatemericdefective virusgenomes (1, 25, 27, 32). Two cis-acting functions were foundtobeessential forthis
propagation,
aDNAreplication
originand acleavage-packaging signal (5, 6, 25a, 28, 29, 32, 33). The HSV-1 (Justin) defectivegenomes were shown to containthereplication origin 1 (ori-1or
Oris)
located in the c inverted repeat sequence (19, 27, 28, 32). The cleavage-packaging signal in the Justin defective genome repeats recentlyhas beenshown to residewithin theUs-a
sequences constituting thejunctions betweenadjacent repeat units (6; L. P. Deiss and N. Frenkel, manuscript inpreparation).*Correspondingauthor.
Earlier studies on the structure of the a sequence in standardHSV-1 (F) DNAestablished that theL-Sjunction (with the structure bac) had an a sequence inthe arrange-ment
DR1-Ub-(DR2)l9-(DR4)3-Uc-DR1
(18,20). The DR1 (20 base pairs [bp]), DR2 (12 bp), and DR4 (37 bp) elements corresponded to stretches ofdirectly repeated sequences, whereas Ub (64bp) andUc
(58bp) corresponded to unique sequences located on the b and c sides within the a se-quence, respectively (seeFig. 3).Fromanalysis (20) ofthe termini of HSV-1 (F) DNA, it appeared that the 20-bp DR1 element contained the target sequenceforthecleavage of circularorconcatemericDNA. Thus, genomic termini could be generated by a single base-pair-staggered cleavage betweennucleotides 18and 19 of DR1 (seeFig. 4), leaving
11/2
bp oftheDR1 sequence(with
a 3' single base extension) at the S terminus and the remaining
181/2
bp oftheDR1 sequence at the Lterminus of thestandard viralgenome (20).We present here the detailed structureof thea sequence present inacloned HSV-1 (Justin) defective genome repeat unit. This work further definesthe structureofafunctional
cleavage-packaging signal
and the nature of the recombina-tional event leading to the formation of the defective virus genomes.MATERIALSAND METHODS
Cells and virus.Thederivation oftheHSV-1
(Justin)
series byserial undilutedviruspropagation in HEp-2 cellshas been describedpreviously (7).Plasmid cloning and nucleotide sequencing. The 8.6-kilobase (kb) repeat units ofthe HSV-1 (Justin) defective genomes werepurified fromanagarosegelafter electropho-resis ofEcoRI-digested, passage-15, HSV-1
(Justin)
DNA. TheEcoRI-cleaved
monomers (seeFig. 1)were clonedinto theEcoRI site ofpACYC184 (2),yielding
theJustin defec-tive(JD) clonedplasmid pJD101.The8.5-kbHindIII-EcoRI fragment from standard HSV-1 (Justin) DNAspanning
co-ordinates 0.910 to 0.965 was inserted between theHindIII and
EcoRI
sites ofpBR322 (seeFig. 1),yielding
theplasmid
pJl.
A 370-bp Sall fragment spanning the region of Us involved in the generation of the defective genome wassubcloned from
pJl
into the Sall site ofpUC9
(31). This clonewasdesignated pON101.Allcloning
wasperformed by
140
on November 10, 2019 by guest
http://jvi.asm.org/
ab
0-S
component
:STAN DARD
L
.S
UL
DEFECTIVE
H
pc
pONIOI
US
HlOObp
sequence
us -
s
-Repeats
[a
cC-IZ-
-a
IC
...8.1kb
Usc
- -.8.4 kb
E
;~~~~~~~
--- 8.6kb
I I
I I
lkb hH
E
pJD1I1
sequence
FIG. 1. Structure andorigin of the defective HSV-1(Justin)genomesandthederivation of the standard and defective virus clones.Left, thepJ1 andpON101 cloneswerederivedbycloning segments of the S componentasdescribed in thetext.Right, the concatemericstructure of thedefective virusDNAmolecules.Repeat unit sequencescorrespondtothe standardvirusDNAsequencesfrom therightside of theS component whendisplayed inthe P orILarrangementsorthe left side of S whendisplayedin theIsorISL orientations (as shown in thisfigure).
Three major size families of repeat units (8.1, 8.4, and8.6kb)differing in theamountof Us sequences have beendescribedpreviously (13). pJD101wascloned as described in thetext.S,Sall; E,EcoRI;H,HindIll.Thecircled S denotes the Sall siteservingasthe referencepoint for nucleotide sequencing. The arrowspoint the5'-to-3' direction of the sequences showninFig. 2.
procedures described previously (21, 25). The nucleotide sequence of32P-end-labeledDNAfragmentswas determined bytheproceduresofMaxamandGilbert(16). The sequence of relevant regions of the plasmid pJD101 was determined starting from the Sall sitein Usand the Hinfl site in the c
sequence(18, 20).The sequenceof theplasmid pON101was determined from the BamHI and HindlIl sites adjacent to the insert inthepolylinker of pUC9 (31).
RESULTS
The presentstudywasundertakentodetermine the nucle-otide sequence of a functional cleavage-packaging signal within defective HSV-1 DNA and to define the structural organization of the novel Us-a junction found between adjacent repeat units ofthe defective virus genomes. We expected that the analysis of the structure of this novel junction would elucidate boththenatureof recombinational events which led to the formation of the defective HSV genomeandthe structure of thecleavage-packaging signal. By restriction enzyme analyses, we have previously (13) distinguished threemajorsizefamiliesofrepeat units in the HSV-1 (Justin)defectivegenomes (8.1, 8.4, and 8.6 kb; Fig. 1). In that study we suggested that the three classes of repeat units represented the products of recombinational events linking the a sequence to different subsets of the Us se-quences. Subsequent replication (most likely by a rolling circlemechanism) ofthe three types ofrepeat units yielded theconcatemericdefective genomes. Assummarized inFig. 1, the two smaller classes of repeat units present in the HSV-1 (Justin) defective genome did not contain the Sall
site located at position 0.944 of standard virus DNA, whereas the largest type of repeat unitsincluded this site.
Tostudy thearrangement of sequences at the novel
Us-a
junction, we digested the Justin defective genomes with EcoRI (this enzyme cleaves once within each repeat) and cloned the resultant monomeric repeat units into a plasmid vector. The clone pJD101 carried the largesttypeofrepeat units (8.6 kb), as defined by the presence of the Sall site adjacentto the
Us-a
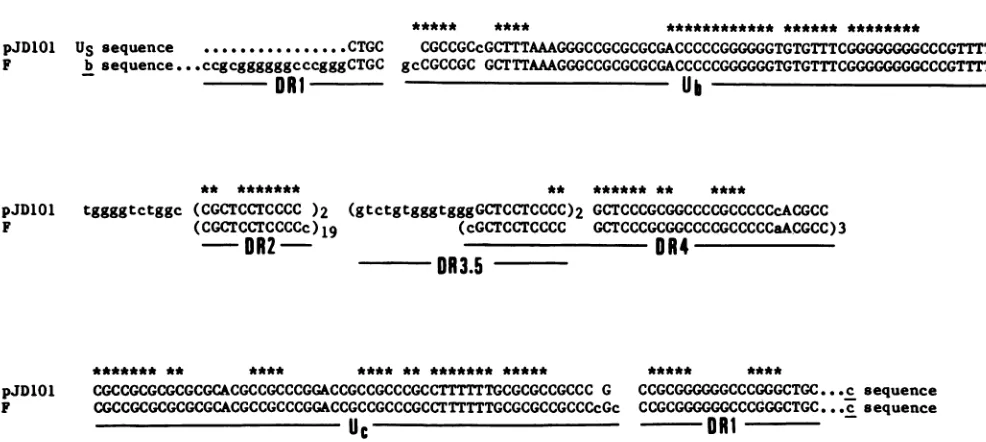
junction (Fig. 1). Inaddition, a 370-bp Sall fragment containing the relevant Us sequences of standard virus DNA was cloned into pUC9 plasmid to generate the clone pON101. The nucleotidesequence for the correspondingcloneddefective and standard virus DNA was determined by using the common Sall site (Fig. 1) as a reference point. All sequences were determined for both DNAstrands bytheproceduresofMaxamand Gilbert (16). Figure2displaysthenucleotidesequences ofpJD101 and pON101 alignedfrom the referenceSallsite,which occupies position 0.944 on thestandard virus DNA. Figure 3 shows the a sequence portion ofpJD101 displayed alongside the previouslydetermined nucleotidesequence of the aregionof standard HSV-1 (F) DNA (18, 20). The results of the nucleotide sequence analyses may be summarized as fol-lows. (i) TheUs
sequences in the pJD101 defective genome were homologous to the corresponding Us sequences of standard virus for 174 bp from the reference SailI site. (ii) The remaining sequences from nucleotide 175 to the end of thesequencedregion correspondedto an asequence resem-bling the a sequence of HSV-1 (F). (iii) The point of transition between theUsand the a sequence is denoted byanasterisk inFig.2and isdisplayedingreater detail in Fig.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.80.527.74.340.2]Sal I
pJD101 GTCGACGCCT TAATACCGAC TGTTGGCGGC CCATGCGTAC GAGGAAGTCG 50 pON101 GTCGACGCCT TAATACCGAC TGTTGGCGGC CCATGCGTAC GAGGAAGTCG pJD101 TTGGCCGCCT CGTCTTCGCT TTCCGAGTAG TAGGCTTCGG CCGAAACTGG 100 pON101 TTGGCCGCCT CGTCTTCGCT TTCCGAGTAG TAGGCTTCGG CCGAAACTGG pJD101 CGAGGCCGTG GGATAAAGCG GCACGGACAT GTCGGATCGC GCTGAGGAGT 150 pON101 CGAGGCCGTG GGATAAAGCG GCACGGACAT GTCGGATCGC GCTGAGGAGT pJD101 TGGGATCGGA GAGCCGGGAC GTCACTGCCG CCGCCGCTTT AAAGGGCCGC 200 pON101 TGGGATCGGA GAGCCGGGAC GTCATCGAGG CCGGAAGAAA GCTCCGGGTG pJD101 GCGCGACCCC CGGGGGGTGT GTTTCGGGGG GGGCCCGTTT TTGGGGTCTG 250 pON101 GGAAGTTGCG GTCGCAGTGA CCTCACGATT TTTAATTTGC TGCGGCTAGG pJD11 GCCGCTCCTC CCCCGCTCCT CCCCGTCTGT GGGTGGGGCT CCTCCCCGTC 300 pON101 CGGACCACCG GCCCTTTATG CGCCTCGGGC AATTGACGTC ACATACCACG
pJD1OI TGTGGGTGGG GCTCCTCCCC GCTCCCGCGG CCCCGCCCCC CACGCCCGCC 350
pON101 CAATCCCACA CAGGCGGCCC CCAAGCCGGA AGCCCCCCGG AGCCACCGAG
pJD11 GCGCGCGCGC ACGCCGCCCG GACCGCCGCC CGCCTTTTTT GCGCGCCGCC 400
pON101 CGGCCGAGG....
pJD101 CGCCGCGGGG GGCCCGGGCT GC.. .c sequence
FIG. 2. Alignment and nucleotidesequenceof thestandard and defective HSV-1 (Justin)DNAin theregion of the Us-ajunction.Thetwo
sequences arealignedatthecorresponding Sall siteatcoordinate0.944 onthestandard viralgenome.Thepoint of transition fromUsto a
sequencesis marked byanasterisk. The 174bp ofUssequences areshownin boldtype.
4.Thedefectivegenome a sequence retainedonly 4 bp of the sequences had a complete 20-bp DR1 element that was
DR1 elementadjacent to the Ub sequences, and these 4bp identical to that found in standard HSV-1 (F) DNA (20). werejoinedto
Us
sequences at asinglepoint, generatingthe Thus, the novelUs-a
junction could be described as theUs-a
junction. In contrast, the boundary between a and c product ofasingle
recombinational eventlinking
theUs
***** **** ************ ****** ********
pJDIO1 US sequence...CTGC CGCCGCcGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTT F b sequence...ccgcggggggcccgggCTGC gcCGCCGC GCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTmT
DRI
Ub
** ******* ** ****** ** ****
pJD101 tggggtctggc (CGCTCCTCCCC )2
(gtctgtgggtgggGCTCCTCCCC)2
GCTCCCGCGGCCCCGCCCCCcACGCCF (CGCTCCTCCCCc)19 (cGCTCCTCCCC GCTCCCGCGGCCCCGCCCCCaACGCC)3
DR2
0R4
OR3.5
******* ** **** **** ** ******* ***** ***** **
pJDO1I CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCCGCCC G CCGCGGGGGGCCCGGGCTGC...c sequence
F CGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCCGCCCcGc CCGCGGGGGGCCCGGGCTGC...c sequence
Uc
ODRi
FIG. 3. Alignmentof the HSV-1(Justin)defective genomeasequencewith that ofstandardHSV-1(F)DNA. Thenucleotide sequence for the HSV-1 (F)asequenceis taken frompreviousstudies byMocarski and Roizman(18,20). Nonhomologous sequencesareshownin smallletters,andhomologous sequencesareshown incapitalletters.Thedesignationsfordirectrepeats(DR)andunique(U)elementsare
those usedpreviously (18, 20)with theadditionofDR3.5,arepeat thatis absentfromtheHSV-1(F)asequence.Asterisks denote sequences conservedinallHSV-1and HSV-2 strainssequencedtodate thatareavailable, i.e.,theHSV-2strainHG52(3),theHSV-1strainsF(18,20), USA-8 and 17(3), andJustin(thispaper).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.138.485.75.344.2] [image:3.612.64.557.454.675.2]Us-a ,.. gagagccgggacgtc C ...
JUNCTION ... ctctcggccctgcagt GAC ... L-S ... ccgcggggggcccgggFTGCj... JUNCTION *.. ggcgccccccgggcc
L COMPONENT ... ccgcggggggcccggg TGT C ... S COMPONENT
TERMINUS ... ggcgccccccgggccc A 3CG ... TERMINUS
FIG. 4. Comparisonof the Us-ajunction with theDR1 element
atthe L-Sjunction,and the sequences locatedatthe L andS termini of standard HSV-1 (Justin) DNA. Purified virion DNA was end labeled with terminal deoxynucleotidyl transferase and 32P-labeled
cordycepin triphosphate. The BamHI terminal fragments were purifiedafterelectrophoresis inanagarosegel, digested withAvaI,
and subjected to electrophoresis on a DNA sequencing gel as previouslydescribed for HSV-1(F)DNA(20).Theboxed sequences denote the 4 bpof the DR1 element retained in the Us-apJD101
junction.
sequenceto an asequence,with thecrossover
point residing
within the left (b-side) DR1 element. (iv) Although closely
related,
the a sequences ofHSV-1(Justin)
and (F) DNAs differinseveralrespects.Asshowninaprevious
report(14), thesize oftheHSV-1(Justin)
asequencewasapproximately half(264bp) that ofthe HSV-1 (F) sequence (501 bp). The presenceofonly
twocopies
ofDR2 in theJustinasequence(compared
with 19copies
in theasequenceofF)
andonly
asingle
copy of aportion
of DR4 (versus 3copies
in F) accountedforthesize differences.Inaddition,
thepJD101a sequencehadtwocopies
ofarepeatwhichwetermedDR3.5 because itoccupied
theequivalent position
betweentheDR3 andDR4elements described forthea sequenceofF DNA.Although
10 of the 23bp constituting
DR3.5closely
re-sembled the DR2 repeat, the
remaining
13bp
had nocounterpartin the HSV-1(F) a sequence.
Interestingly,
the DR3.5 element was present in sixcopies
within the asequenceofthe HSV-1 strainUSA-8 studied
by
Davisonand Wilkie(3)
butwasabsentfromthea sequenceofthe HSV-1 strain 17(3).
The presence ofthese sequences in theJustin and USA-8 strainsmight
reflectacloserelationship
between these two strains of HSV-1.DISCUSSION
Theresults of the studies
presented
inthispaperaddress the nature ofeventsleading
to thegeneration
of defective HSV genomes and thestructureof theasequenceinrelation to its function inthecleavage-packaging
process.Three
points
can be maderegarding
thegeneration
of defective virusgenomes.First,
it appears that the defective HSV genomes evolved as a consequence ofa recombina-tionalprocessinvolving
an asequence.Thisconclusionrests on theobservationthat thepoint
oftransitionfrom Usto a sequence in the defective genome repeatunit
occurred within thea sequenceitself.Second,
our studies have revealed that the a sequence whichbecameincorporatedinto the Justindefective genome repeatunitwasoriginally derived fromanL-Sjunction (Fig. 1)rather thanfromanS end. This conclusionrests onresults of studies whichweredesigned
todetermine thestructureof the L and S termini of standard HSV-1 (Justin) DNA.Specifically,
these studies (E. S.Mocarski, unpublished
data)
were done with32P-labeled
cordycepin triphosphate
and terminal deoxynucleotidyl transferase to label the termini of standard HSV-1 (Justin) DNA. The labeled end
fragments
were then analyzed by using protocolssimilartothose previously used to analyze the termini of standard HSV-1(F)DNA(20). Thededucedstructureof the L and S termini is displayed in Fig. 4. The Justin S end, like its F counterpart(20),containedan asequenceterminatingwith 1 bp of the DR1 element and an additional 3' protrudingbase, whereas the L endterminated with theremaining
181/2
bpof the DR1 sequence. The novel Us-a junctionwhich retained 4 bp of the DR1 sequence could not have been formed by recombination involving the S terminus because that termi-nus retained only 11/2 bp of DR1. Therefore, the original recombinational eventleading to the joining of theUsandasequences within the pJD101 repeat unit most likely in-volved an a sequence situated within an L-Sjunction.
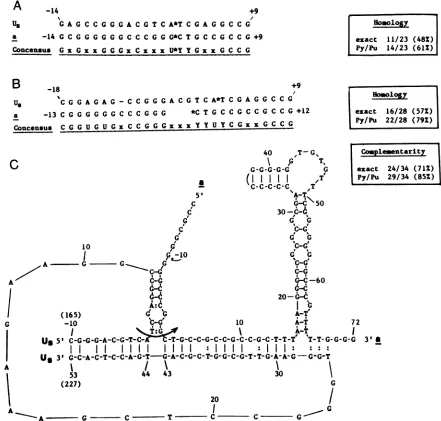
The final ppint regarding the evolution of the defective HSV genomes relates to the mechanism of the Us x a sequencerecombination. The structure of the
Us-a
junction in the pJD101defectivegenome repeatcould bemost simply explained bya single recombinational eventlinking the Us and asequences. It is intriguing, however, that the crossover site occurred close to the cleavage target within the a sequence, suggesting that the original repeat unit evolved as an aberrant product of an a sequence-mediated activity, most likely cleavage-packaging or inversion. Indeed, as depicted in Table 1 and Fig. 5, the corresponding Us region in a standard virus DNA contained three types of homolo-giestothe a sequence thatmighthavefacilitated theUs x a recombination. First,severalstretchesof sequences closely resembling elements of the a region could be identified within the Usregion at a relatively distant location(50to 200 bp) from theUs-a
junction point. Thesepartlyhomologous sequences are listed in Table 1. One or more of these sequences mighthaveplayedarole inthe aberrant recogni-tionofthecorresponding Us region bythe cleavage-packag-ing or the inversion machinery, thereby facilitating subse-quentinteraction with a.A second type of sequence similarity existed in close proximity to the
Us-a
junction and included the partly homologoussequencesdepictedinFig.SB. These sequences might have played a role in stabilizing recombinational intermediates, although little homology couldbefoundwhen the Us and a sequences were aligned precisely at the crossover site (Fig. SA). Finally,astretchof Ussequences, occurring inan invertedpolarity25 to 53 bpawayfrom theUs-a
junctionsite exhibitedcomplementaritytoboththeUs andasequencesimmediatelyflanking
thejunction (Fig.5C). This sequence might have thus served tojuxtapose the Us and a sequences before an excision-ligation event. The juxtaposed sequence might have been stabilized by the base-paired stem structures depicted in Fig. SC. Similar models for recombinations promoted by short sequence homology or by alignment through a third sequence have been previously proposed (see, for example, references 9 and 22).We cannot predict whether the data and hypotheses presentedabove will beconsistent with the structureof the smaller families of the Justin repeat units or with the structure of defective genomes generated from other HSV strains (4,5, 10, 11, 15,25). However, itisnoteworthy that from restriction mapping data, many but not all, ofthese defective genomes appear to have evolved from an a
se-quence "invasion" of internalgenomic sequences (11, 15), an event that most likely occurred in the pJD101 Justin repeat. Thecombinedpressure toacquirethe DNA replica-tionoriginaswell as thecleavage-packaging signaland at the
same time remain within the size constraints imposed on defective genome repeat units (12) undoubtedly plays a
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Ussequencesdistaltothecrossoverpoint showing partial homologytosequencesin a
Homologye
Regiona Location' Strand' Sequenced Exact Py/Pu
(%) (%)
Us -46/-61 2 129TGTCCGTGCCGCTTTA"14 12/16 14/16
a(Ub) +2/+17 1 176TGCCGCCGCCGCTTTA'91 (75) (87)
Us +96/+82 2 268TAAAGGGCCG-GTG256 12/14 13/14
a(Ub) +16/+29 1 19TAAAGGGCCGCGCG203 (86) (93)
Us +131/+ 147 1 305CCCACACAGGCGGCCCC321 13/17 14/17
a(DR4) +144/+160 1 318CCCGCTCCCGCGGCCCC334 (76) (82)
Us +168/+ 150 2 342CTCCGGGGGGCTTCCGGCT324 13/19 16/19
a(DR1) -16/+2 1 CCGCGGGGGGCCCG-GGCT226 (68) (84)
Us +181/+ 131 2 355GGCCGCTCGGTGGCTCCGGGGGGCTTCCGGCTTGGGGGCCGCCTGTGTGGG305 32/51 36/51 a(Ub) +21/+71 1 195GGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTTTGGG245 (63) (71)
Us ++172/+145 2 346GTGGCTCCGGGGGGCTTCCGGCTT-GGGG319 20/29 23/29
a(Ub) +45/+72 1 219GTGTTTCGGGGGGGG-CCCGTTTTTGGGG246 (69) (79)
a Thesubregion withinacontainingthe sequenceshown isgiven inparentheses.
bThe numbers referto theposition of the sequence relativetotheUs-acrossoverpoint.
cStrand1has beendesignatedasthestrand shown inFig.2.
dThe numbersrefertotheposition in thesequence shown inFig.2.
e Exacthomology is scored foridentical bases; Py/Pu homology is scoredforpairsofpyrimidinesandpurines. controlling role in theevolutionofHSV defective genomes.
The predictions of the model above, namely that various types of repeat units of defective virus genomes might containone or moresequenceshomologous to a sequences, arecurrentlyunder investigation.
In a recent set of studies (6; Diess and Frenkel, in preparation)wehave fused theJustinUs-ajunction(derived by subcloningfrom the pJD101 plasmid) to a functional HSV DNAreplication origin. Analysesoftheability ofthe
result-ant clone to become propagated into packaged defective genomes in the presence of helper virus DNA clearly showed that the
Us-a
junction fragment contained a func-tional cleavage-packaging signal. Several points can there-fore be madefromthe resultsofthe presentstudy regarding thecleavage-packaging process. First, ourdataextend ear-lierreports regarding variations in the a sequences of dif-ferent HSV-1 strains. Specifically, thefindingof arelatively small numberofthe DR2 and DR4 elements in theJustin asequenceindicatedthatahigh degreeofreiteration ofthese sequenceswasnotessential for any of the knownasequence functions, aconclusion consistent with the previousresults ofDavisonand Wilkie(3). Inaddition,theJustinasequence containedtworeiterations ofthe23-bpDR3.5 sequence and in that respect resembled theasequenceoftheHSV-1 strain USA-8 which contained six copies of this sequence (3). These results clearly indicate significant flexibility in the structuralorganization of thea sequence.
Anadditional point which can be made from ourstudies relates to the finding that the HSV-1 (Justin) defective genome repeatretainedonly4bp of the DR1 sequenceatthe
Us-a
junction. Therefore, the presence of two complete copies ofthe DR1 sequence, oneateachboundaryof a,was notrequired for cleavage-packaging. Furthermore, as seenin Fig. 4 and 5, the
Us
sequences at the novel junction differed considerably from the DR1 sequences which they replaced. There are currently two possible explanations for thesefindings. First, the DR1 portionwhichwasretained in the defective genome repeat (4 bp) could contain thees-sential sequences recognized duringthecleavage-packaging
process. Alternatively, it is possible that the cleavage-packaging signaldoes not include anyof the DR1 sequences. Rather, the cleavage-packaging process could involve the recognition of sequence elements present at internal location(s) within the a sequence, followed by cleavage within sequences located at a constant distance from the recognition signalitself. Although further workis requiredto
test these hypotheses, it is noteworthy that there is a considerable lack of sequence homology between the DR1 elementof HSV-2(strainHG52[3])and the DR1elementsof HSV-1 strains (3, 18, 20; this paper). At the same time, results fromouradditional studies(5, 26) have revealed that HSV-1 a sequences (fromstrains Patton, F, and the Justin defective genomes) couldbeefficientlyrecognized for cleav-age and packaging by trans-acting products specified by an heterologous HSV-2 helper virus. These observations strongly suggestthat the DR1 sequenceitself doesnotform thecleavage-packaging signal.Ourfindingthattwocomplete copies ofDR1 werenotessentialforthe
cleavage-packaging
of theJustindefective genomes lends further supportfor this hypothesis.
In contrasttothedivergence oftheDR1elements, certain internal subsetsoftheasequencehavebeenconservedin all the HSV-1 and HSV-2 strains sequenced to date. These conserved sequences have been marked with asterisks in Fig. 3. Of these various shared sequences, the oneswithin the Ub and Uc regions occupyin all HSV strains
(including
pJD101) a similar position relative to the L and S cleaved ends. Homologyto the Ub conserved nucleotide sequences
canbefound in otherherpesvirusesaswell,includinghuman cytomeglovirus where it resides within the
putative
a se-quence (30;R. R. Spaeteand E. S.Mocarski, submitted for publication). It is noteworthy that theregion
ofhomology between thesedifferentherpesvirusasequencesiscontained within thelargehairpinloop shown inFig.5C.Therefore,wesuggest that these sequences could serve as
recognition
signals forcleavage-packagingand that thecleavageprocess itselfcould involveameasuringfunctiontodefined distances away from these signals. The recent
finding
(26) that theon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.54.557.90.283.2]A
-14 +9U8 G A G C C G G G A C G T C A*T C G A G G C C G
a -14 GC G GG GGGC C C G GG*C T G C C G C C G+9 Concensus G x G xx G G G x C x x x U*Y Y G x x G C C G
ts -18 +9
Us C G G A G A G - C C G G G A C G T C A*T C G A G G C C G a -13 C G G G G G G C C C G G G *C T G C C G C C G C C G
Concensus C G G U G U Gx C C G G Gx x x Y Y U Y C Gx x G C C G +12
Homology
exact 11/23 (48%)
Py/Pu 14/23 (61%)
Hosology
exact 16/28 (57%) Py/Pu 22/28 (79%)
40 ,T-G%
Couplementarity
C\
,~~~~~~~~~~~~~~~~~~G
T,G-G-G-G-G G exact 24/34 (71%)
O
( | | | | { T Py/Pu 29/34 (85%)
- ~~~C-C-C-C-C ,T
5 ' A-T
,C G,-C 50
C 30-C-G
G G G
C C-G
10GG
GI
I~~~~~~
I 1A G G
A G G-C-60
20-G-C
(165) G G
A-I
-10 C- 10 A-T 72
Ug
5' --C-C-G-C-C-G-C-C-G-C-T-T-T T-T-G-G-G-G3'1
US 3' G-C-A-C-T-C-C-A-G-T-G-A-C-G-C-T-G-G-C-G-T-T-G-A-A-G G-G-T
1 53 44 43 30
(227) G
20 /
A C G
A G C T C C G
FIG. 5. Thealignment of the Usanda sequences surrounding the Us-a crossover point. (A) When aligned precisely at the junction (denoted by asterisk), the Usanda sequences show relatively poor homology. The numbers next to the sequences denote the sequence position relativetothecrossoverpoint. Thesequence to theleft oftheasterisk intheUssequence and to the right of theasterisk in the a sequence are fused in theUs-a junction.Exacthomologyreferstoidenticalbasepairs,whereasPy/Pudenote pyrimidines orpurines present atanequivalent position. (B) Moreextensivehomology isobserved with theUs and asequences proximal to the Us-ajunction point are slightly misaligned relative to the Us-ajunction site tomaximize homology. (C) A significantly more stable structure can be formed by juxtaposing theUsand ajunction points through theircomplementarity to a Us sequence located 26 to 53 bp away from the crossover point. Thearrow denotes theproposed recombinationaleventinvolving theexcision-repairprocess. The level of complementarity (precise base pairs and Py-Pu pair)wascalculated forthehorizontal"backbone" sequence (the base-paired region positions 28 to 53 of Us juxtaposing the Usandasequences) and the leftstemformed by annealingof DR1 with the Us sequence.
human cytomegalovirus a sequence can substitute for an HSV-1 a sequence as a cleavage-packaging signal in a constructed HSV-1 defective genome amplicon (25) re-inforcesthis notion. Studies with constructed HSV defective genomes arecurrently in progress to further investigate the involvement of different portions of the HSV-1 a sequence in thecleavage-packaging process.
ACKNOWLEDGMENTS
WethankBernardRoizman for support and S. Rudofsky and A. Liuforexcellent technical assistance.
These studies were supported by Public Health Service grant AI-15488 from the National Institutes of Health and by National ScienceFoundation grant PCM 8118303(N.F.) and by Public Health ServiceResearch grantAI-20211and a Leukemia Research Founda-tion Grant (E.S.M.). Portions of this work were done in the laboratoryof B. Roizman where E. S.M. was aSpecialFellowofthe LeukemiaSociety ofAmerica.
ADDENDUM IN PROOF
After the acceptance of this paper we learned that S. L. Varmuza and J. R. Smiley (Cell, in press) have
independ-r'l
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.84.525.68.489.2]ently shown that cleavageoccurs at a constantdistancefrom internal a sequences and that DR1 is not required for this
process.
LITERATURE CITED
1. Barnett, J.W.,D. A. Eppstein, and H. W. Chan. 1983.ClassI defective herpes simplex virus DNA as a molecular cloning vehicle in eukaryotic cells. J. Virol. 48:384-395.
2. Chang, A. C. Y., and S. N. Cohen. 1978. Construction and characterization of amplifiable multiple DNA cloning vehicles derived from the P1SA cryptic plasmid. J. Bacteriol. 134: 1141-1156.
3. Davison, A.J.,and N. M.Wilkie.1981.Nucleotide sequences of the jointbetween the L and S segments of herpes simplex virus type 1 and 2. J. Gen. Virol. 55:315-331.
4.
Denniston,
K.J., M. J. Madden, L. W. Enquist, and G. Vande Woude. 1981. Characterization of coliphage lambda hybrids carrying DNA fragments from herpes simplex virus type 1 defective interfering particles. Gene 15:365-378.5. Frenkel, N.1981.Defectiveinterfering herpesviruses, p.91-120. In A.J. Nahmias, W. R. Dowdle, and R. S.Schinazi (ed.), The human herpesviruses: an interdisciplinary prospective. Elsevier-North Holland Inc., New York.
6. Frenkel, N.,L. P.Deiss, and R.R.Spaete. 1984.Studies of HSV DNApropagation using HSV amplicons. UCLA Symp. Mol. Cell. Biol. New Ser. 21:289-299.
7. Frenkel, N., R. J. Jacob, R. W. Honess, G. S. Hayward, H. Locker,and B.Roizman.1975.Anatomyofherpessimplex virus DNA. III. Characterization ofdefective DNA molecules and biological properties of virus populations containing them. J. Virol. 16:153-167.
8. Frenkel, N., H. Locker, W.Batterson, G.S. Hayward, andB. Roizknan. 1976. Anatomy ofherpes simplex virus DNA. VI. Defective DNA originates from the S component. J. Virol. 20:527-531.
9. Glickman,B.W.,and L.S.Ripley. 1984. Structural intermedi-atesof deletion mutagenesis:arole forpalindromicDNA.Proc. Natl. Acad. Sci. U.S.A.81:512-516.
10. Kaerner, H.C.,I. B.Maichle,A.Ott, and C.H.Schroder.1979. Origin oftwo different classes of defective HSV-1 Angelotti DNA.Nucleic Acids Res. 6:1467-1478.
11. Knopf, C., G.Strauss,A.Ott-Hartmann, R. Schatten, andH.C. Kaerner.1983. Herpessimplex virus defective genomes: struc-tureof HSV-1 ANG defective DNAof class II and encoded polypeptides. J.Gen. Virol. 64:2455-2470.
12. Kwong, A. D., and N. Frenkel. 1984. Herpes simplex virus amplicon: effect of sizeonreplication of contructed defective genomes containing eukaryotic DNA sequences. J. Virol. 51:595-603.
13. Locker, H., and N. Frenkel. 1979. Structure and origin of defective genomes contained in serially passaged herpes simplex type 1 (Justin). J. Virol. 29:1065-1077.
14. Locker, H., and N. Frenkel. 1979. TheBamI, KpnI, andSall restriction enzyme maps oftheDNAs ofherpes simplex virus strains Justin and F: occurrence ofheterogeneities in defined regions of the viralDNA.J. Virol. 32:429-441.
15. Locker, H.,N. Frenkel,andI.Halliburton. 1982. Structure and expression of class II defective herpes simplex virus genomes encoding infected cell polypeptide number 8. J. Virol. 43:574-593.
16. Maxam, A.M., andW.Gilbert. 1979. Sequencing end-labeled DNAwith basespecificchemicalcleavage. MethodsEnzymol. 65:499-560.
17. Mocarski,E.S., L. E. Post,andB. Roizman. 1980. Molecular engineering of the herpes simplex virus genome: insertion ofa second L-Sjunction into the genome causes additional genome inversions. Cell 22:243-255.
18. Mocarski,E. S.,and B. Roizman. 1981. Site-specific inversion sequence of the herpes simplex virus genome: domain and structural features. Proc. Natl. Acad. Sci. U.S.A.78:7047-7051. 19. Mocarski, E. S., and B.Roizman. 1982. Herpesvirus-dependent amplification and inversion of cell-associated viral thymidine kinase gene flankedby viralasequences andlinkedto anorigin of viral DNA replication. Proc. Natl. Acad. Sci. U.S.A. 79:5626-5630.
20. Mocarski,E.S.,andB.Roizman.1982. Structure and role of the herpes simplex virus DNAtermini ininversion, circularization and generation of virion DNA. Cell31:89-97.
21. Post, L. E., A. J. Conley,E.S.Mocarski, and B. Roizman. 1980. Cloning of reiterated and non-reiterated herpessimplex virus 1 sequencesas BamHIfragments. Proc. Natl. Acad. Sci. U.S.A. 77:4201-4205.
22. Ripley, L. S., and B. W. Glickman. 1982. Unique self complementarity of palindromic sequences provides DNA structural intermediates for mutation. Cold Spring Harbor Symp. Quant. Biol. 47:851-861.
23. Roizman, B. 1979. The structure and isomerization of herpes simplex virus genomes. Cell 16:481-494.
24. Smiley, J. R.,B. S.Fong, and W.C.Leung. 1981. Construction of a double-jointed herpes simplex virus DNA mnolecule: in-verted repeats are required for segment inversion, and direct repeats promote deletions. Virology 113:345-362.
25. Spaete,R. R., and N. Frenkel. 1982. Theherpes simplex virus amplicon: a neweucaryotic defective-virus cloning-amplifying vector. Cell 30:295-304.
25a.Spaete,R.R., and N. Frenkel. 1985. Theherpes simplex virus amplicon: analyses of cis-acting replication functions. Proc. Natl.Acad. Sci. U.S.A. 82:694-698.
26. Spaete,R.R.,and E.S.Mocarski. 1984.Trans-acting functions encoded by herpes simplex virus-1 recognize cis cleavage-packaging signals on cytomegalovirus DNA. UCLA Symp. Mol.Cell. Biol. New Ser. 21:359-372.
27. Stow,N. D. 1982.Localization ofanorigin ofDNAreplication within theTRs/IRsrepeatedregion of the herpes simplex virus type1genome EMBOJ. 1:863-867.
28. Stow,N. D., and E.C. McMonagle. 1983. Characterization of theTRsIIRsorigin ofDNAreplicationofherpes simplex virus type1.Virology 130:427-438.
29. Stow, N. D., E. D. McMonagle, and A. J. Davison. 1983. Fragmentsfrom both termini oftheherpessimplex virus type1 genome contain signals required fortheencapsidationof viral DNA. NucleicAcids Res. 11:8205-8220.
30. Tamashiro, J. C.,D.Filpula, T.Friedman, and D. H.Spector. 1984.Structure ofheterogeneous L-Sjunctionregion of human cytomegalovirus strainAD169 DNA. J.Virol. 52:541-548. 31. Viera, J., andJ. Messing. 1982. The pUC plasmids, an M13
mp7-derived system for insertion mutagenesis and sequencing withsynthetic universalprimers. Gene 19:259-268.
32. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes
simplex virus DNA: localization ofreplicationrecognition sig-nals within defective virus genomes. Proc. Nati. Acad. Sci. U.S.A. 78:742-746.
33. Vlazny,D.A.,A. D.Kwong,and N. Frenkel.1982.Sitespecific cleavage-packagingofherpessimplexvirusDNAandthe selec-tive maturation of nucleocapsids contianing full length viral DNA. Proc. Natl. Acad. Sci. U.S.A.79:1423-1427.