Require Specific Dimerization of the NS4A Protein Transmembrane
Domain
Andrew Kohlway,aNathan Pirakitikulr,bFrancisco N. Barrera,aOlga Potapova,bDonald M. Engelman,aAnna M. Pyle,b,c,d Brett D. Lindenbache
‹Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, Connecticut, USAa; Department of Molecular, Cellular, and Developmental Biology,
Yale University, New Haven, Connecticut, USAb; Department of Chemistry, Yale University, New Haven, Connecticut, USAc; Howard Hughes Medical Institute, Yale
University, New Haven, Connecticut, USAd; Department of Microbial Pathogenesis, Yale University, New Haven, Connecticut, USAe
Hepatitis C virus (HCV) NS4A is a single-pass transmembrane (TM) protein essential for viral replication and particle assembly. The sequence of the NS4A TM domain is highly conserved, suggesting that it may be important for protein-protein interactions. To test this hypothesis, we measured the potential dimerization of the NS4A TM domain in a well-characterized two-hybrid TM protein interaction system. The NS4A TM domain exhibited a strong homotypic interaction that was comparable in affinity to glycophorin A, a well-studied human blood group antigen that forms TM homodimers. Several mutations predicted to cluster on a common surface of the NS4A TM helix caused significant reductions in dimerization, suggesting that these residues form an interface for NS4A dimerization. Mutations in the NS4A TM domain were further examined in the JFH-1 genotype 2a replicon system; importantly, all mutations that destabilized NS4A dimers also caused defects in RNA replication and/or virus assembly. Computational modeling of NS4A TM interactions suggests a right-handed dimeric interaction of helices with an interface that is consistent with the mutational effects. Furthermore, defects in NS4A oligomerization and virus particle assembly of two mu-tants were rescued by NS4A A15S, a TM mutation recently identified through forward genetics as a cell culture-adaptive muta-tion. Together, these data provide the first example of a functionally important TM dimer interface within an HCV nonstruc-tural protein and reveal a fundamental role of the NS4A TM domain in coordinating HCV RNA replication and virus particle assembly.
H
epatitis C virus (HCV) is an enveloped, positive-sense RNA virus in theFlaviviridaefamily (1). HCV has been grouped into seven major genotypes and several subtypes (2,3). The 9.6-kb genome is translated into a single⬃3,000-amino-acid polypeptide by cap-independent translation through an internal ribosome en-try site (1). The polypeptide is cleaved into three structural teins— core, E1, and E2—as well as seven nonstructural (NS) pro-teins—p7, NS2, NS3, NS4A, NS4B, NS5A, and NS5B (4). The structural proteins and p7 are cleaved from the polypeptide by cellular signal peptidases, whereas cleavage of the NS proteins is accomplished by the NS2-NS3 (NS2-3) cysteine autoprotease and the NS3-NS4A (NS3-4A) serine protease (4).HCV NS4A is a 54-amino-acid polypeptide that anchors NS3 to the endoplasmic reticulum (ER) (1,5,6). NS4A encompasses an N-terminal, alpha-helical, single-pass transmembrane (TM) region required for association with the ER membrane (6–8), a central region that mediates interaction with the NS3 protease domain (5,6,9–11), and a C-terminal region implicated in both RNA replication and virus particle assembly (12–15). NS4A func-tions as a cofactor for both serine protease and RNA helicase ac-tivities of NS3 and exhibits genetic and physical interactions with several other NS proteins, including NS2, NS4B, NS5A, and NS5B
(15–19). Additional roles for NS4A include translation inhibition
(20, 21) through interaction with elongation factor 1A (22), regulation of the innate immune response through cleavage of the antiviral mitochondrial signaling adaptor MAVS as part of NS3-4A (23–25), and recruitment of creatine kinase B to facilitate viral genome replication (26).
To date, the molecular architecture of the HCV replication
complex and the process by which it is assembled from viral NS and host proteins remain obscure. Several studies have revealed interactions between HCV and host proteins (17,27,28). Nuclear magnetic resonance (NMR) and X-ray crystal structures have also been obtained for individual domains of HCV NS proteins (8,
29–33), and progress has been made in isolating crude replication
complexes from HCV-infected cells that retain endogenous repli-cation activity (34–36). Despite these gains, the molecular details of interactions between NS proteins and between NS proteins and host factors remain to be elucidated. Interestingly, TM regions appear to play a fundamental role in the incorporation of the NS proteins into the replication complex (7,8,37), motivating our efforts to understand their interactions.
To examine the propensity of the NS4A TM domain to ho-modimerize, we employed the GALLEX system, a well-character-ized two-hybrid assay capable of measuring the strength of TM domain interactions in biological membranes (38–40). In this
sys-Received23 July 2013 Accepted22 October 2013
Published ahead of print30 October 2013
Address correspondence to Anna M. Pyle, [email protected], or Brett D. Lindenbach, [email protected].
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JVI.02052-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02052-13
on November 7, 2019 by guest
http://jvi.asm.org/
tem, a TM domain is fused between an N-terminal, cytosolic, DNA binding domain of the LexA transcriptional repressor and a C-terminal, periplasmic domain of maltose binding protein (MBP). Dimerization of the heterologous TM domain results in a LexA fusion capable of repressing-galactosidase expression in
Escherichia coli(41,42). Similar bacterial two-hybrid systems have also been developed to examine homo- and heterotypic interac-tions between TM domains (43–48).
As a reference, we used the best-studied model system for the study of TM protein interactions, human glycophorin A (GpA), a single-pass TM protein that forms a homodimer and has been characterized in model membranes, detergent micelles, and two-hybrid assays (49–53). The “GxxxG” motif found in the GpA TM domain is one of the more prevalent and well-studied TM inter-action motifs, which also include “GxxxA,” “GxxxS,” and other variations (39,54–59). Within the GxxxG motif, glycines or other small residues, such as serine or alanine, allow the two alpha heli-ces to pack together in close proximity, leading to formation of hydrogen bonds between the carbonyl group and neighboring C␣ hydrogens (55,60). In addition,-branched residues such as iso-leucine and valine contribute to TM alpha-helical packing inter-actions by constraining side-chain rotamers and thereby limiting entropic penalties in helix interactions (55,58). We show here that the NS4A TM domain also contains variations of the GxxxG motif and that these motifs contribute to NS4A dimerization.
We find that dimerization of the NS4A TM domain is required for the HCV life cycle. The NS4A TM domain exhibits a strong propensity for dimerization in the GALLEX system, and several residues critical for oligomerization were identified through mu-tational analysis. Mutations in the NS4A TM domain were then examined in a genotype 2a replicon system, revealing strong cor-relations between NS4A dimer formation and RNA replication and/or virus particle assembly. All-atom molecular dynamics sim-ulationsin vacuoprovide a model for a right-handed dimer of NS4A alpha helices with a helix-helix interface that predicts the sensitive mutational sites. Together, these observations reveal the importance of the membrane as a scaffold in functional interac-tions of viral proteins.
MATERIALS AND METHODS
General computational tools.Graphpad Prism 6 was used for data anal-ysis and graphing. Models of the NS4A dimer were made in Pymol (PyMOL Molecular Graphics System, Version 1.5.0.4, Schrödinger, LLC, http://www.pymol.org). Adobe Photoshop and Illustrator CS6 were used to construct the figures. The NS4A TM alignment was created by using the UGENE toolkit (61).
Molecular biology.Standard molecular biology techniques were used for cloning; plasmids were sequenced at the W.M. Keck Foundation Cen-ter at Yale University. Forward and reverse primers for site-directed mu-tagenesis were ordered from Invitrogen. The pYSGR-JFH1/GLuc replicon was previously described by Phan et al. (15). The GALLEX backbone plasmid pBLM100 was previously described by Schneider and Engelman (38).
NS4A mutations were introduced into pYSGR-JFH1/GLuc by using the Quikchange protocol with 2⫻PFU DNA polymerase master mix (Stratagene). Forward primers used for mutagenesis are listed inTable 1. Amplification products were digested with DpnI for 6 h at 37°C and then transformed into MachI cells (Invitrogen). Plasmids were prepared (Qia-gen miniprep kit), sequences were verified, and correct mutants were subcloned back into pYSGR-JFH1/GLuc by using common NsiI and BsrGI restriction sites.
For creation of the NS4A TM mutant library, forward and reverse
primers with SpeI and SacI restriction sites were used for PCR with the wild-type (WT) and mutant NS4A TM regions, which were then ligated into SacI/SpeI-digested pBLM100. One extra nucleotide was needed after the SacI restriction site to keep the TM domain and MBP in the correct reading frame. Ligations were transformed into MachI cells, plasmids purified with Qiagen miniprep kits, and sequences verified at the Yale Keck facility. The WT NS4A TM GALLEX constructs are listed in Table S1 in the supplemental material.
Cell culture replication and infectivity.pYSGR-JFH1/GLuc was di-gested with XbaI at 37°C overnight and mung bean nuclease at 30°C for 30 min. HCV RNA was made by runoff transcription with T7 RNA polymer-ase and purified by Qiagen RNeasy columns. HCV RNA was stored at ⫺20°C in ME buffer (10 mM MOPS [morpholinepropanesulfonic acid; pH 6.5], 1 mM EDTA).
Replication assays were performed in a 24-well plate format and seeded 1 day prior to the experiment with 5⫻104Huh-7.5[core-NS2] cells per well. Huh-7.5[core-NS2] cells were as described by Phan et al. (15) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) (In-vitrogen) containing 10% fetal calf serum (HyClone, Logan, UT) and 1 mM nonessential amino acids (Invitrogen). The Huh-7.5[core-NS2] cells were created by using the pLenti4 (Invitrogen) lentivirus transduction system to express the Jc1 core, E1, E2, p7, and NS2 proteins via the human cytomegalovirus immediate-early-1 promoter. HCV RNA was trans-fected with MIR-2250 mRNA lipid transfection reagent (Mirus) accord-ing to the manufacturer’s protocol. Media were collected at 8, 24, 48, 72, and 96 h posttransfection and clarified in a microcentrifuge at 16,000⫻g. Media were then mixed with 5⫻luciferase lysis buffer (NEB), andGaussia luciferase activity was measured in a Berthold Centro LB 960 luminescent plate reader with aGaussialuciferase kit (NEB). For luminescent mea-surements, 10 to 20l of media was assayed with 50l of luciferase reagent, and the signal was integrated over a span of 2 to 10 s.
For released infectivity assays, 100l of media collected from the 24-well replication assays was added to 96-well plates of naïve Huh-7.5[core-NS2] cells, previously seeded at 6⫻103cells per well. After 12 to
16 h of infection, each 96-well plate was washed with phosphate-buffered saline (PBS) and fresh DMEM was added to each well. Media were col-lected 48 h after the wash, andGaussialuciferase activity was measured as described above for replication.
For intracellular infectivity assays, Huh-7.5[core-NS2] cells were transfected with HCV RNA, washed with PBS after 8 h, and then harvested at 48 h posttransfection by trypsinization. Cells were centrifuged at 1,200⫻gfor 5 min, resuspended in DMEM, and subjected to three rounds of freezing with liquid nitrogen and thawing at 37°C. The cell lysates were then clarified at 16,000⫻gfor 5 min, and the lysates were tested for infectivity of naïve Huh-7.5[core-NS2] cells in 96-well plates as described above.
GALLEX -galactosidase assays. The pBLM100 expression con-struct, SU101 cells, and the GpA WT strain and GpA G83I GALLEX mu-tants were described previously (38). A modified protocol from Zhang and Bremer (62) was followed for the-galactosidase assays. Overnight cultures of SU101 cells were grown in LB media containing the appropri-ate IPTG (isopropyl--D-thiogalactopyranoside) concentration and 200
g/ml ampicillin. Cultures were diluted to anA600of 0.1 and grown at
37°C to anA600of 0.6. Then, 20l of cells was added to 80l of permea-bilization solution (100 mM Na2HPO4, 20 mM KCl, 2 mM MgSO4, 0.8
mg/ml hexadecyltrimethylammonium bromide, 0.4 mg/ml sodium de-oxycholate, 5.4l/ml-mercaptoethanol [BME]) and warmed to 30°C. A 600-l volume of prewarmed substrate solution (60 mM Na2HPO4, 40
mM NaH2PO4, 1 mg/ml o-nitrophenyl--D-galactoside, 2.7l/ml BME)
was added to the permeabilized cells, and the reaction mixture was then incubated at 30°C for various lengths of time until a yellow color appeared (longer for strong oligomerization). After approximately 60 to 120 min, 700l of 1 M Na2CO3was added to stop the reaction and debris was spun
down for 10 min in a microcentrifuge. TheA420was measured in a spec-trophotometer, and Miller units (MU) were calculated by using the
on November 7, 2019 by guest
http://jvi.asm.org/
lowing equation: MU⫽1,000⫻[A420/(A600⫻0.02 ml⫻substrate
incu-bation time)]. The interaction affinity data were fitted by using the hyperbolic function [monomer]⫽1⫺[[IPTG]/([IPTG]⫹Kd)], where Kd(dissociation constant)⫽[monomer]2/[dimer].
Test for insertion of GALLEX TM domain.To test if the chimeric NS4A TM GALLEX constructs localized to the inner membrane ofE. coli SU101 cells, membrane fractions were isolated by alkaline extraction fol-lowing the protocol of Russel and Model (63). Briefly, a 0.5-ml volume of cells was put on ice and an equal volume of ice-cold 0.2 M NaOH solution was added. Samples were then vortexed and centrifuged for 15 min at 4°C, and the pellet was resuspended in 1⫻SDS sample buffer. The soluble fraction was precipitated by adding 110l of 100% trichloroacetic acid (TCA) to the supernatant to achieve a final concentration of 10%. The mixture was kept at 4°C overnight, centrifuged, and washed with 0.5 ml of ice-cold acetone. The precipitant was then resuspended in 1⫻SDS buffer. Samples were separated on 12% Bio-Rad Tris-glycine gels, and Western blots were run in a Bio-Rad wet transfer apparatus onto polyvinylidene difluoride (PVDF) membranes. Maltose binding protein (MBP) poly-clonal (PA1-989) and anti-rabbit secondary horseradish peroxidase (HRP) (31460) antibodies were purchased from Thermo Scientific.
Test for orientation of GALLEX TM domain.To test for correct ori-entation of LexA-TM-MBP chimeric proteins, GALLEX constructs were transformed into NT326E. colicells lacking endogenous MBP (malE⫺) and cultured on M9-agar plates following a modified protocol from Schneider and Engelman (38). Transformants were plated onto LB plates containing 200g/ml carbenicillin and 0.01 mM IPTG. Single colonies were grown overnight in M9 minimal media (22 mM KH2PO4, 47.6 mM
Na2HPO4, 8.55 mM NaCl, 18.7 mM NH4Cl, 2 mM MgSO4, 0.1 mM
CaCl2) supplemented with trace elements (6.3M ZnSO4, 7M CuCl2,
7.1M MnSO4, 7.6M CoCl2, 60M FeCl3), 2g/ml thiamine, 0.4%
glucose, 200g/ml carbenicillin, and 0.02 mM IPTG. Overnight cultures were pelleted, washed with PBS, resuspended in 200l of PBS, and plated onto M9 minimal-medium plates with 1.5% maltose, 200g/ml carben-icillin, 1.5% noble agar, and 0.02 mM IPTG. Plates were incubated at 37°C for 3 to 4 days to assay for MBP complementation. We detected subtle variations in the levels of MBP complementation for a few of the NS4A TM mutants. A reduction in complementation could be explained by the structural constraints that the TM mutants impose on the MBP because of an altered oligomeric interface. MBP must be able to diffuse freely and bind to the MBP transporter in the proper orientation in order for com-plementation to occur (64).
[image:3.585.42.542.78.427.2]Modeling of the NS4A TM dimer.A model for dimeric TM helices (see Table S2 in the supplemental material) was generated by using the CHI (CNS [Crystallography and NMR System] Helix Interactions) com-putational search strategy (65,66). Briefly, CHI was used to construct a pair of canonical alpha helices from the NS4A TM sequence with lengths of 16, 17, 18, and 19 amino acids with both left-handed and right-handed helical arrangements. Molecular dynamics (MD) simulations were per-formedin vacuoby using simulated annealing. Energy minimization was performed before and after the MD simulations, and the resulting struc-tures with backbone root mean square deviations (RMSD) of 1 Å or less were placed into clusters corresponding to the final model. The search was carried out over the entire two-body rotational interaction space (0 to 360°), and different interhelix distances (9 to 11 Å) and crossing angles (10
TABLE 1Site-directed mutagenesis primers for the HCV NS4A TM domaina
NS4A mutation JFH-1 codon H77 codon Replicon primer sequence Replicon GALLEX
T2A 1663 1659 CATGACCAGCGCGTGGGTCCTAG x x
W3A 1664 1660 GACCAGCACGGCGGTCCTAGCTG x x
W3F 1664 1660 GACCAGCACGTTTGTCCTAGCTG x
V4A 1665 1661 CAGCACGTGGGCCCTAGCTGGAG x x
V4G 1665 1661 CAGCACGTGGGGCCTAGCTGGAG x x
L5A 1666 1662 CACGTGGGTCGCAGCTGGAGGAG x x
L5G 1666 1662 CACGTGGGTCGGAGCTGGAGGAG x x
A6L 1667 1663 GTGGGTCCTACTTGGAGGAGTCC x x
G7A 1668 1664 GGTCCTAGCTGCAGGAGTCCTGG x x
G7L 1668 1664 GGTCCTAGCTCTAGGAGTCCTGG x x
G8A 1669 1665 CCTAGCTGGAGCAGTCCTGGCAG x x
G8L 1669 1665 CCTAGCTGGACTAGTCCTGGCAG x x
V9A 1670 1666 AGCTGGAGGAGCCCTGGCAGCCG x
V9G 1670 1666 AGCTGGAGGAGGCCTGGCAGCCG x x
L10A 1671 1667 TGGAGGAGTCGCGGCAGCCGTCG x x
L10G 1671 1667 TGGAGGAGTCGGGGCAGCCGTCG x x
A11L 1672 1668 AGGAGTCCTGCTAGCCGTCGCCG x x
A12L 1673 1669 AGTCCTGGCACTCGTCGCCGCAT x x
V13A 1674 1670 CCTGGCAGCCGCCGCCGCATATT x x
V13G 1674 1670 CCTGGCAGCCGGCGCCGCATATT x
A14L 1675 1671 GGCAGCCGTCCTCGCATATTGCC x x
A15L 1676 1672 AGCCGTCGCCCTATATTGCCTGG x x
A15S 1676 1672 AGCCGTCGCCTCATATTGCCTGG x x
Y16A 1677 1673 CGTCGCCGCAGCTTGCCTGGCGA x x
Y16S 1677 1673 CGTCGCCGCATCTTGCCTGGCGA x x
Y16F 1677 1673 CGTCGCCGCATTTTGCCTGGCGA x
Y16W 1677 1673 CGTCGCCGCATGGTGCCTGGCGA x
C17A 1678 1674 CGCCGCATATGCCCTGGCGACTG x x
C17S 1678 1674 CGCCGCATATAGCCTGGCGACTG x
L18A 1679 1675 CGCATATTGCGCGGCGACTGGAT x
L18G 1679 1675 CGCATATTGCGGGGCGACTGGAT x x
T20A 1681 1677 TTGCCTGGCGGCTGGATGCGTTT x x
G21V 1682 1678 CCTGGCGACTGTATGCGTTTCCA x
ax, the NS4A mutation was introduced into either the replicon or GALLEX context.
on November 7, 2019 by guest
http://jvi.asm.org/
to 40°) were explored. Clusters for both left-handed and right-handed dimers of NS4A alpha helices were obtained. However, only the right-handed dimer model was in agreement with the experimentally deter-mined interaction interface. The searching algorithm settled on a crossing angle of between approximately 25° and 30° in all of the clusters. The predicted interaction energy for NS4A residue W3 is high because stack-ing between aromatic rstack-ings of the TM helices, particularly in the 16-, 17-, and 18-amino-acid dimer models, can occurin vacuobut is unlikely to occur in a true lipid-water interface.
Modeling of NS3-4A in a membrane.The 19-amino-acid NS4A dimer was modeled into a membrane of 1-palmitoyl-2-oleoylphosphati-dylcholine (POPC) created with the VMD membrane builder (67). Two protease domains of NS3 (3O8B) were docked across from each other with the constraints imposed by the NS4A dimer and the␣0helix of the
NS3 protease domain (8). The cytosolic portions of the NS3protease-4A dimer were minimized with the Yasara Energy Minimization Server (68). For placement of the NS3 helicase, the domain was positioned and ori-ented relative to the protease domain through the alignment with the dengue virus NS3 structure (2WHX).
RESULTS
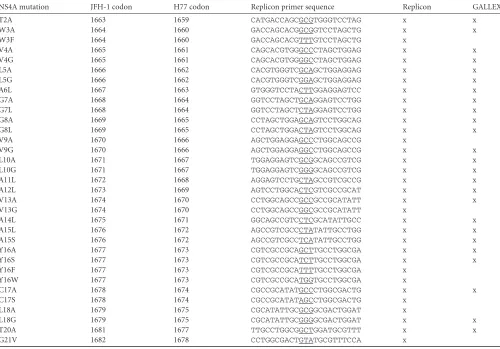
The NS4A TM domain dimerizes in biological membranes. Ap-proximately 80% of the NS4A TM region is identically conserved across all genotypes (Fig. 1A), suggesting that the sequence of this region has functional significance beyond acting as a TM anchor (69). Notably, the NS4A TM domain contains a central region with several invariant glycines and alanines (G7GxxAAxAA15), as
well as two valines, which could contribute to intramembrane oligomerization. Based on these observations, we hypothesized that the NS4A TM domain forms a homotypic protein-protein dimer within the plane of the membrane.
There are no generally applicable, direct biochemical methods to detect TM domain interactions within mammalian cells. There-fore, to test our hypothesis that the NS4A TM domain can self-interact, we utilized the GALLEX assay, a well-characterized two-hybrid system for measuring TM domain interactions, including human protein TM dimers, in biological membranes (38). Pep-tides corresponding to 16-, 17-, 18-, and 19-amino-acid segments of the NS4A TM domain (see Table S1 in the supplemental mate-rial) were fused between an N-terminal LexA transcriptional reg-ulatory domain and a C-terminal maltose binding protein (MBP). The chimeric NS4A TM constructs were expressed under the con-trol of thetacpromoter in SU101E. colicells containing a -ga-lactosidase reporter gene with an upstreamctxpromoter that is repressed by LexA dimers. Because the LexA regulatory domain and MBP do not dimerize on their own, oligomerization of the NS4A TM domain should drive LexA dimerization and repress
-galactosidase reporter gene expression (see Fig. S1 in the sup-plemental material).
While the length of a TM domain can influence the strength of oligomerization in the GALLEX system (38), all four of the 16- to 19-amino-acid segments of NS4A exhibited strong homotypic in-teraction (see Fig. S2 in the supplemental material). We therefore chose to further characterize the 19-amino-acid segment of the NS4A TM domain as a conservative choice for the region of inter-est. First, we compared the oligomerization affinity of the NS4A TM domain to that of the GpA TM domain previously character-ized in the GALLEX system, as well as to that of the GpA G83I mutant, which is defective in dimerization (38). pBR322, the backbone plasmid for expression of LexA-TM-MBP, served as a negative control. At 0.02 mM IPTG, which induced near-saturat-ing levels of chimeric protein, the wild-type (WT) NS4A and GpA
TM domains exhibited similar oligomerization affinities, and the measured-galactosidase activities (Miller units) were over 10-fold lower than those of the negative controls, GpA G83I and pBR322 (Fig. 1B).
Next, in order to calculate an approximate free energy of dimerization, we employed the method described for GpA by Fin-ger et al., which takes advantage of the linear relationship between IPTG concentration and induction of protein expression, at least within the approximate range of 0.001 to 0.1 mM IPTG (40). IPTG was titrated at concentrations ranging from 0 to 0.5 mM to vary the expression level of the NS4A and GpA TM domains (Fig. 1C), and the initial and saturating-galactosidase activities were used to normalize the monomeric fraction. Note that the GALLEX assay can register higher-order oligomers and that dimerization of the NS4A TM domain would be the simplest case. Based on hy-perbolic curves fitted to the data, the apparentKdvalues of the
NS4A and GpA TM domains were 2.9 and 2.1M, respectively; this was in close agreement with the 3.1M apparentKd
calcu-lated previously for GpA (40). These values correspond to appar-ent free energies of dimerization (⌬G) of 7.5 and 7.7 kcal/mol for NS4A and GpA, respectively. Furthermore, a fit to the Hill equa-tion yielded a cooperativity coefficient (“n”) of approximately 1 (data not shown). This apparent lack of cooperativity suggests that the NS4A TM domain is likely to form a dimer rather than a higher-order oligomer. Together, these data indicate that the NS4A and GpA TM domains have similarly strong tendencies to homodimerize in a biological membrane.
Mutations in the NS4A TM domain exhibit defects in dimerization.After establishing that the NS4A TM domain can interact with itself in the GALLEX system, we wanted to identify residues in NS4A that contribute to this interaction. We therefore created a library of mutants designed to disrupt TM helix packing interactions without affecting TM helix stability or membrane insertion. Based on these considerations, the mutations were lim-ited to substitutions of glycine, alanine, and leucine residues that are abundant in TM domains, with the one exception of a ty-rosine-to-serine mutation (58) (Table 1). In addition, we included double and triple mutants with mutations of G7, A11, and A15 residues, which could fall on the alpha-helical interface of a right-handed dimer, following thei,i⫹4 rule. These three residues form part of the conserved double-alanine and double-glycine patches and may be part of a GxxxG-like scaffold.
Generally speaking, for a hypothetical TM-TM interaction, the increase in volume from glycine to leucine is expected to disrupt tight packing of the TM helices. For example, the small glycine residues in the GxxxG-like motifs are important to allow close packing of helices and formation of backbone hydrogen bonds (see the introduction above). Therefore, larger residues at the gly-cine positions would cause steric clashes, preventing the two TM alpha helices from coming into close contact and inhibiting the formation of stabilizing hydrogen bonds. A decrease in volume from a leucine to a glycine could decrease van der Waals packing interactions between helices and introduce a destabilizing void or pocket. This point is highlighted by computational and experi-mental models predicting that a loss of buried surface area be-tween TM helices costs 26 to 39 cal · mol⫺1· Å⫺2of surface area, similar to the packing energetics of soluble proteins (70,71).
The ability of the WT NS4A TM domain to repress -galacto-sidase activity in the GALLEX assay implies that it was properly inserted and oriented within theE. colimembrane. However,
on November 7, 2019 by guest
http://jvi.asm.org/
fore measuring the-galactosidase activity of the NS4A TM do-main mutants, we wanted to test whether each mutation affected insertion and proper orientation into the membrane. To test for the correct orientation of the NS4A TM domain, with LexA in the
cytoplasm and MBP in the periplasm, we performed a maltose complementation assay with NT326 cells that lack endogenous MBP. We first tested each construct on minimal-medium plates supplemented with glucose and determined that the expression of
FIG 1NS4A TM domain dimerizes in the GALLEX system. (A) Alignment of the NS4A TM domain with representative sequences from the seven major HCV
genotypes. (B) A 19-amino-acid segment of the NS4A TM domain and a 16-amino-acid segment of the GpA TM domain dimerize in the GALLEX assay. The GpA G83I mutant is defective in dimerization; pBR322 served as a vector control.-Galactosidase activities were measured in Miller units and are displayed on a log2
scale to facilitate visualization of both large and small differences. (C) Estimation of dimer interaction affinity. The monomeric fraction was plotted against IPTG concentration and fitted to a hyperbolic curve. (D) NS4A TM mutant GALLEX plasmids and controls were transformed into NT326 cells lacking MBP (malE) and plated onto M9 minimal media with 1.5% maltose. Plates were incubated at 37°C for 3 to 4 days to test for complementation of MBP. (E) Membrane (M) and soluble (S) fractions from SU101 cells expressing 19-amino-acid NS4A mutant GALLEX plasmids were isolated by alkaline extraction. Fractions were run on 12% SDS-PAGE gels and subjected to Western blot analysis for MBP. The band runs at approximately 50 kDa. (F) The-galactosidase repression in SU101 cells of 19-amino-acid NS4A TM mutants in the GALLEX system. The pBR322 negative control was reproduced from the experiment described for panel B. Each -galactosidase experiment was performed in at least triplicate, and error bars represent the range.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.122.457.67.560.2]the WT and mutant NS4A TM domains did not affect cellular growth of the NT326 cells (see Fig. S3 in the supplemental mate-rial). Then, we established that the WT and mutant NS4A TM domains complemented the maltose auxotrophy of NT326 cells, indicating proper insertion and orientation of the TM domain
(Fig. 1D).
To further confirm the membrane insertion of the WT and mutant NS4A TM domains, we performed an alkaline extraction to isolate integral membrane proteins from SU101 cells. A West-ern blot analysis for MBP confirmed that the expression levels of the WT and mutant NS4A TM chimeras were similar and that the chimeras were predominately in the pellet (membrane) fraction of
E. coli, with little to no protein expressed in the supernatant (sol-uble) fraction (Fig. 1E). Therefore, we conclude that the expres-sion levels of correctly oriented and inserted NS4A TM chimeras were similar for all of the mutants tested in the GALLEX system.
After establishing that our library of mutations did not affect the insertion of the NS4A TM domain, we measured the -galac-tosidase activities of the single, double, and triple NS4A TM do-main mutants (Fig. 1F). The-galactosidase experiments were performed with a constant IPTG concentration of 0.02 mM, a concentration at which the WT NS4A TM domain repressed the maximum level of-galactosidase activity (Fig. 1C). This concen-tration of IPTG was chosen to maximize oligomer formation and thereby reveal dimer-disrupting mutations but was still in the pre-dicted linear range of protein induction based on previous work with GpA (40).
Single mutations in the double-glycine patch, G7 and G8, as well as in the second double-alanine patch, A14 and A15, demon-strated the greatest disruption in-galactosidase repression. As expected, the G7L and G8L mutations disrupted-galactosidase repression more than the G7A and G8A mutations. There were 2-to 5-fold defects in-galactosidase repression for leucine muta-tions in the first double-alanine patch, A11L and A12L, indicating that this patch appears to be less critical for oligomerization of the NS4A TM domain. However, a double- or triple-mutation com-bination of G7L, A11L, or A15L disrupted-galactosidase activity to nearly the level seen with the negative control, pBR322. This observation is particularly interesting because it suggests that the conserved alanine and glycine patches are instrumental in the packing of NS4A TM domain oligomers. Outside the double-gly-cine and -alanine patches, the only mutations that destabilized the NS4A TM dimer were L5G and V9G. Together, these data define residues in the NS4A TM domain that influence dimer stability.
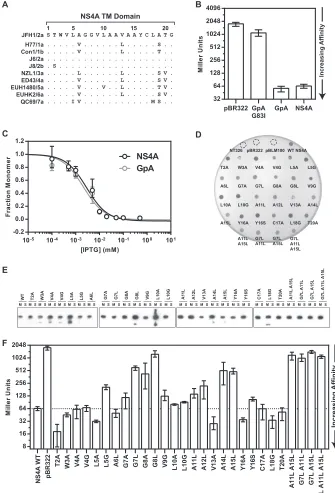
Several residues in the NS4A TM domain are required for HCV replication and infectivity.To study the role of the NS4A TM domain in viral replication and infectious virus production, we examined the phenotypes of NS4A TM mutants in the JFH-1 genotype 2a HCV replicon system (Table 1). In particular, we were interested in determining whether mutants with oligomer-ization defects in the GALLEX system also sustained defects in the viral life cycle. In addition, we expanded our mutant library to include G21V, a mutation previously identified to disrupt HCV replication (8), as well as additional substitutions (mainly to G, A, and L) at several positions: W3, V9, V13, Y16, C17, L18, and G21. All NS4A mutations were studied in the context of the bicistronic subgenomic replicon JFH1-GLuc, which constitutively expresses the secreted luciferase fromGaussia princeps(Fig. 2A) (15). To facilitate the production of pseudoinfectious virus particles, rep-licons were transfected into Huh-7.5 cells constitutively
express-ing core-NS2 (Huh-7.5 [core-NS2]) as described by Phan et al. (15). The experimental setup used to measure replication and re-leased infectivity is illustrated inFig. 2B. Media were collected at 8, 24, 48, 72, and 96 h posttransfection, and RNA replication was assayed by measuring the activity of luciferase released into the media of transfected cells. Virus production was determined by using the media or cell lysates to infect naive cells and measuring nascent luciferase production. A well-characterized defective NS5B polymerase mutant, GNN, was used as a negative control for the cell culture experiments.
Nearly half of the NS4A TM domain mutants—V4G, L5G, G7A, G7L, G8L, V9G, L10A, L10G, A11L, A15L, Y16A, Y16S, L18G, and G21V— had severe defects in RNA replication (Fig. 2C). Certain positions—V4G, L5G, G8L, V9G, Y16S, and L18G— required severe mutations (larger volume changes) in order to disrupt replication, whereas moderate mutations at the same po-sitions—V4A, L5A, G8A, V9A, Y16F/W, and L18A—retained WT or near-WT levels of replication. Two residues, G7 and L10, ex-hibited defects both with the moderate substitutions G7A and L10A and with the more severe substitutions G7L and L10G. The alanine residues were scanned with only one set of mutations— A6L, A11L, A12L, A14L, and A15L—and we observed replication defects in only two of those positions—A11 and A15.
Aromatic residues, particularly tyrosine and tryptophan, can foster aromatic-aromatic interactions between TM alpha helices or can contribute to stability at the lipid-water interface of a TM alpha helix (72,73). Because of the location of Y16 near the end of the NS4A alpha helix segment and the requirement for an aro-matic residue, Y16 may interact with the phospholipid head group region of the lipid bilayer. Similarly, W3 is invariant among HCV isolates but was surprisingly not required for HCV replication. Thus, W3 may contribute to the overall stability of the NS4A TM helix without being essential for the function of NS4A, at least in the context of our cell culture model.
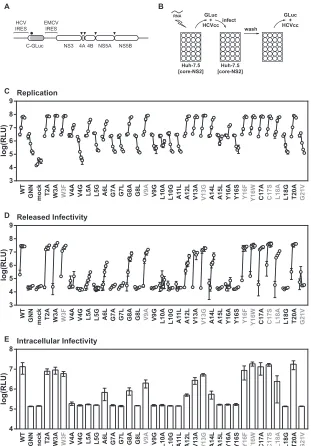
Next, we measured the level of infectious virus released into the media at each time point (Fig. 2D). As expected, mutants that were unable to replicate did not produce infectious virus. However, several mutants with near-WT levels of viral replication were im-paired in the secretion of infectious virus particles, including V4A, L5A, A6L, G8A, A12L, A14L, and L18A. Two mutants—V4A and L5A— had lags in replication that likely contributed to the de-crease in infectious virus production at earlier time points. In order to establish if the defects in virus production were due to reduced levels of virus assembly or virus release, we measured the levels of intracellular infectivity from cells lysed at 48 h postinfec-tion (Fig. 2E). The intracellular infectivity levels for each mutant correlated well with the levels of released infectivity, indicating that defects in virus production were attributable to decreased virus assembly, although we cannot quantitatively assess how sub-tle defects in replication contribute to defects in virus production. CHI all-atom simulation predicts a right-handed dimer of NS4A TM domains.To provide a detailed structural model of the NS4A TM homotypic interaction, we used the program CHI to perform a series of molecular dynamics simulations for TM helix interactions. CHI is a helix-interaction modeling package inte-grated within the CNS (Crystallography and NMR System) soft-ware suite (65,66). Two NS4A TM domain alpha helices were generated and then docked in locations adjacent to one another at the beginning of each simulation. After an initial energy minimi-zation, a simulation was initiated and the helices were allowed to
on November 7, 2019 by guest
http://jvi.asm.org/
explore the full rotational interaction space, interhelix distances, and crossing angles. Upon completion of the simulation, a final energy minimization was performed and the resulting structures were binned based on root-mean-square deviations (RMSD). Simulations were performed with NS4A TM helices of lengths of 16 to 19 amino acids and with two initial helix-helix crossing
angles. Clusters of resulting structures represent chemically favor-able interactions of the helices. Both right- and left-handed dimers of NS4A were generated; however, a right-handed parallel dimer, which aligns G7, A11, and A15, was more compatible with our experimental results.
The relative enthalpic interaction energies contributed by each
log(RLU)
3 4 5 6 7 8 9
A
Released Infectivity D
Intracellular Infectivity E
Replication C
B
NS3 4A 4B NS5A NS5B C-GLuc
log(RLU)
3 4 5 6 7 8 9
log(RLU)
4 5 6 7 8
HCV IRES EMCVIRES
Huh-7.5 [core-NS2]
Huh-7.5 [core-NS2] RNA
FIG 2Replication and infectivity of HCV subgenomic replicon in Huh-7.5 cells expressing core-NS2. (A) Diagram of the GLuc replicon construct for
monitoring HCV replication in Huh-7.5[core-NS2] cells. Triangles indicate sites cleaved by NS3-4A, and the circle indicates a signal peptidase cleavage site at the N terminus of the GLuc gene that leads to the secretion ofGaussialuciferase. IRES, internal ribosome entry site; EMCV, encephalomyocarditis virus. (B) Design of replication and released infectivity experiments. (C) Replication of NS4A TM mutants in the GLuc-JFH1 replicon context in Huh-7.5[core-NS2] cells. Media were collected at 8, 24, 48, 72, and 96 h and then assayed for secretedGaussialuciferase. The GNN mutant is the product of a mutation that disrupts the active site of the NS5B RNA-dependent RNA polymerase. The luciferase activity (yaxis) at each of the time points— 8, 24, 48, 72, and 96 h—is plotted in succession from left to right (xaxis) for each NS4A mutant. RLU, relative light units. (D) Released infectivity measurements of the media collected at 8, 24, 48, 72, and 96 h for each NS4A TM mutant. The media assayed for replication as described for panel C were used to infect naive cells, and then fresh media were assayed 48 h after infection for secreted luciferase. The time courses of luciferase expression are plotted as described for panel C. (E) Intracellular infectivity measurements of lysate from infected cells. Infected cells were trypsinized from plates, centrifuged, and subjected to three rounds of freezing and thawing. Cell lysates were then used to infect naive cells, and fresh media were assayed 48 h after infection for secreted luciferase. Each point or column in panels C, D, and E represents the average of the results of three experiments, and the error bars represent the standard deviations. Designations of mutants tested in cell culture but not in the GALLEX assay are colored gray in panels C, D, and E.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.137.448.66.512.2]residue were averaged from every simulation of 16- to 19-amino-acid helices and plotted inFig. 3A. A parallel right-handed 19-amino-acid NS4A TM dimer structure from our lowest energy bin is shown inFig. 3BandC. The predicted dimer model reveals that three residues—G7, A11, and A15— constitute the center of the dimer interface. Based on this model, it is clear that mutations with larger volumes at these positions would introduce steric clashes with the opposing helix and mutations with smaller vol-umes at other positions such as L18 would eliminate van der Waals packing interactions between helices (see Fig. S4 in the sup-plemental material).
Based on the predicted role of residue A11 in stabilizing the dimeric interface and the observation that A11L caused profound defects in RNA replication, it was surprising that the A11L muta-tion had only a modest destabilizing effect on dimer formamuta-tion in the GALLEX assay. It is worth noting that in the context of the NS4A G7L and A15L mutations, the A11L mutation substantially exacerbated defects in GALLEX oligomerization (Fig. 1F). None-theless, to reconcile these results, we hypothesized that the NS4A A11L mutant may adopt an alternative dimer interface, a mecha-nism previously observed for mutations that unexpectedly stabi-lize GpA dimers (74). To test this hypothesis, we ran an additional CHI simulation on the NS4A A11L mutant to predict whether an alternate packing interface could exist for this mutant. We discov-ered that the NS4A A11L mutant displayed an interaction energy profile similar to that displayed by WT NS4A, suggesting a pro-pensity to dimerize, but the packing interface of the NS4A A11L dimer was shifted by one residue and the crossing angle increased from 29° to 36° (see Fig. S5 in the supplemental material). This prediction may explain why the A11L mutant showed oligomer-ization in the GALLEX system whereas the shifted packing inter-face could have disrupted replication in cell culture, further high-lighting the specificity and functional importance of dimer formation.
The NS4A TM dimer model complements the experimental data obtained from the GALLEX and cell culture mutational anal-yses. The highest interaction energies from the computational dimer model were observed for residues V4, G7, L10, A11, A14, A15, and L18. These predicted determinants for homodimeriza-tion overlap the set of empirically determined constraints for ho-modimerization detected via the GALLEX assay (G7, A11, A14, and A15) and overlap residues that, when mutated, caused defects in RNA replication (V4A, V4G, G7A, G7L, L10A, L10G, A11L, A15L, and L18G) or virus assembly (A14L and L18A). Although we did not detect GALLEX deficiencies at positions V4, L10, and L18, it is possible that subtle defects at these positions may have been missed because the NS4A TM domain was overexpressed at a high level under the control of thetacpromoter.
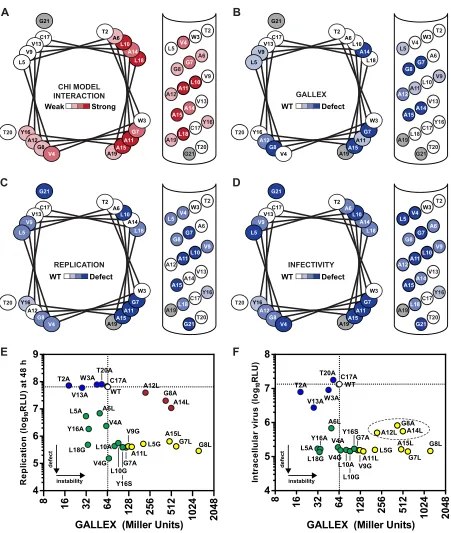
One surface of the NS4A TM alpha helix is responsible for homotypic interaction.A summary of the GALLEX, cell culture, and CHI experimental results, including helical wheel and helical net representations as well as graphs comparing the phenotypes in the cell culture assay versus the GALLEX assay, is shown inFig. 4. In the helix representations, residues were classified based on their sensitivity to mutation or contribution to calculated binding en-ergy. The typically observed crossing angles in right-handed (⫺40°) and left-handed (20°) TM dimers result in a unique inter-action interface that can be represented by modifying the number of residues per turn of a single alpha-helical wheel (75). This rep-resentation groups residues that fall together on a putative dimer
FIG 3CHI all-atom simulation predicts a right-handed parallel NS4A dimer.
(A) The CHI simulation was run with 16-, 17-, 18, and 19-amino-acid-length NS4A TM segments (see Table S1 in the supplemental material) at two differ-ent starting helix-helix crossing angles of 25° and 40°. The average interaction energy (kcal/mol) from the lowest energy cluster of each simulation was plot-ted for each residue. The NS4A T2 and W3 positions yielded interaction ener-gies of 1.0 and 8.0 kcal/mol, respectively, but are not shown in the graph because of the high interaction energies caused by the artifactual pi-pi stacking between the W3 positionsin vacuo(see Materials and Methods). (B) Top-down view of the predicted NS4A dimer with positions G7, A11, and A15 highlighted in red. (C) Side view of the predicted NS4A dimer with positions G7, A11, and A15 highlighted in red.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.80.245.62.568.2]FIG 4Helical wheel representations of the replication, infectivity, and oligomerization properties of mutants at each position of the NS4A TM domain. (A) Summary of the CHI simulation data. Amino acid positions are colored darker to represent a stronger interaction energy as predicted by the CHI simulation. (B) Summary of the GALLEX data on NS4A homo-oligomerization. (C) Summary of the HCV cell culture replication data. (D) Summary of the HCV cell culture infectivity data. For panels B, C, and D, amino acid positions are colored based on defects observed in the oligomerization, replication, or infectivity properties of mutants at each position. Amino acid positions are colored light (WT) to dark (defective) for each amino acid position according to whether a mutation or mutations at each position disrupted oligomerization, replication, or infectivity. Amino acid positions are colored a lighter shade if a mutation or mutations exhibited only small defects or if two mutations at the same position exhibited different phenotypes. (E) The replication phenotypes (indicated in RLU at 48 h) are plotted against the GALLEX interaction phenotypes. The white oval and dotted line represent the WT phenotype; blue, mutants with only minor defects in replication and GALLEX interaction; green, mutants with defects in replication but no defect in GALLEX interaction; yellow, mutants with defects in replication and GALLEX interaction; red, mutants with no defects in replication but defects in GALLEX interaction. (F) The intracellular infectivity phenotypes are plotted against the GALLEX interaction phenotypes. Mutants are colored as described for panel E. For panels E and F, unpaired Student’sttests determined that the L10A, L10G, Y16S, G7A, V9G, and A11L GALLEX results were statistically significant (P⬍ 0.05) compared to the WT NS4A results; however, mutants were labeled defective if their results were above the 2-fold threshold. Given the larger dynamic range of the HCV replication and infectivity assays, mutants were labeled defective if their results were above the 10-fold threshold.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.68.519.60.593.2]interface. Therefore, to more clearly represent the helix-helix con-tacts in a right-handed dimer, alpha-helical wheel representations were drawn with 3.9 residues per turn to align the interaction interface. For comparison, a left-handed alpha-helical wheel rep-resentation with 3.5 residues per turn was also included (see Fig. S6 in the supplemental material) (76). Note that the NS4A TM alpha helix still retained a canonical 3.6 residues per turn and that the helical wheel alterations represent only potential dimeric in-teraction interfaces. These data highlight strong correlations be-tween RNA replication, infectious virus production, and homo-typic interaction within the GALLEX assay. The predicted dimerization interface also aligns well with the surface important for RNA replication, although it is slightly shifted from the surface important for interaction within the GALLEX assay.
There is a remarkable concordance between the cell culture phenotypes and GALLEX results. Specifically, every point muta-tion that disrupts NS4A dimer formamuta-tion in the GALLEX sys-tem—L5G, G7L, G8A, G8L, V9G, A11L, A12L, A14L, and A15L— also disrupts either viral replication or infectious virus production (yellow points inFig. 4EandF). Three mutations—G8A, A12L, and A14L— destabilized dimer formation in the GALLEX assay but had minor effects on virus replication (red points inFig. 4E). However, these mutations caused strong defects in virus assembly (dotted oval inFig. 4F), suggesting that the mechanism by which they destabilize dimer formation is still compatible with RNA
rep-lication but may cause defects in virus assembly. Several muta-tions—V4A, V4G, L5A, A6L, G7A, L10A, L10G, Y16A, Y16S, and L18G— did not destabilize NS4A in the GALLEX assay but caused severe defects in cell culture (green points inFig. 4EandF). These data support a model in which the homotypic interaction of the NS4A TM domain is required for both viral replication and infec-tivity but suggest that other, heterotypic TM interactions may also be required.
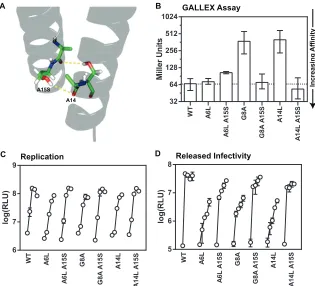
NS4A A15S suppresses defects in cell culture and GALLEX oligomerization.While our reverse genetic approach supports a model for NS4A dimerization, the discovery of sequence prefer-ences through forward genetic analysis could provide further ev-idence for this model. Interestingly, a mutation within the NS4A TM domain, A15S, was identified as a key adaptive change that, when combined with two additional mutations in NS3 and NS5B, conferred efficient replication and virus production to genotype 1a, 2a, and 2b viral genomes that were previously unable to repli-cate in cell culture (77, 78). Based on our model of NS4A dimerization, this serine substitution at position 15 could provide additional hydrogen bonds to help stabilize a TM dimer (Fig. 5A). Consistent with this, previous GALLEX studies showed that a ser-ine substitution was more stabilizing than an alanser-ine substitution for the second glycine within the GxxxG motif of GpA (39).
We therefore tested the effects of the NS4A A15S adaptive mu-tation in our assay systems. Because this adaptive mumu-tation was
Miller U n it s WT A6 L A6 L A 1 5 S G8 A G8 A A 1 5 S A1 4 L A1 4 L A1 5 S 32 64 128 256 512 1024 C GALLEX Assay Increasing Affinit y A WT A6 L A6 L A 1 5 S G8 A G8 A A 1 5 S A1 4 L A 14L A 15 S Replication log(RLU) 6 7 8 9 B log(RLU) 5 6 7 8 Released Infectivity WT A6 L A 6L A1 5 S G8 A G8 A A 1 5 S A1 4 L A1 4 L A 1 5 S D A15S A14
FIG 5The NS4A A15S mutation rescues replication and assembly defects of the NS4A A6L, G8A, and A14L mutants. (A) The A15S mutation in the context of
the proposed NS4A dimer model would create two additional hydrogen bonds with the backbone carbonyl of the preceding residue and in theory stabilize the NS4A dimer. (B) The-galactosidase repression in SU101 cells of A6L, G8A, and A14L with and without the A15S mutation in the GALLEX system. (C) Replication of A6L, G8A, and A14L with and without the A15S mutation in the GLuc-JFH1 replicon context in Huh-7.5[core-NS2] cells. Media were collected at 8, 24, 48, 72, and 96 h and then assayed for secretedGaussialuciferase. (D) Released infectivity measurements of the media collected at 8, 24, 48, 72, and 96 h for each NS4A TM mutant. The media assayed for replication in panel C were used to infect naive cells, and then fresh media were assayed 48 h after infection for secreted luciferase. Each cell culture and-galactosidase experiment was performed in at least triplicate, and error bars represent the standard deviation (cell culture) or range (-galactosidase).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.134.449.66.352.2]associated with increased levels of cell culture replication and vi-rus production in nonreplicative strains, we introduced the NS4A A15S mutation into the NS4A G8A and A14L mutants that exhibit defects in both virus production and homotypic interaction, as well as into the NS4A A6L mutant that has a defect in virus pro-duction but normal homotypic interaction. Not surprisingly, the NS4A A15S single mutant showed no cell culture or GALLEX defects and exhibited a slight increase in homotypic interaction (see Fig. S7 in the supplemental material). Interestingly, NS4A A15S suppressed the defects in homotypic interaction of the G8A and A14L mutants in the GALLEX assay (Fig. 5B; see also controls in Fig. S8 in the supplemental material). Furthermore, NS4A A15S suppressed the defects in infectious virus production of NS4A A6L, G8A, and A14L mutants and moderately improved cell cul-ture replication levels (Fig. 5CandD). Taken together, these for-ward genetic data further support the idea of a functional role for the homotypic interaction of the NS4A TM domain.
DISCUSSION
Our hypothesis, that NS4A TM domains have a propensity to oligomerize, is strongly supported by experimental evidence for dimerization in the GALLEX assay system (Fig. 1), by reverse ge-netic analyses indicating that NS4A TM oligomerization is impor-tant for HCV RNA replication and virus particle assembly (Fig. 2), by computational modeling that predicts a right-handed dimer structure with functionally important residues at the dimer inter-face (Fig. 3), and by identification of a cell culture-adaptive mu-tation that stabilizes NS4A TM dimer formation (Fig. 5). We pro-pose that NS4A interacts with itself, most likely as a dimer, through interaction motifs within the TM domain. While a dimer is the simplest case of homotypic interaction, is consistent with the lack of cooperativity seen in the GALLEX assay, and minimizes steric constraints that higher-order oligomers would likely en-counter, we cannot absolutely exclude the possible occurrence of higher-order homo-oligomeric interactions in biologically rele-vant membranes. Taken together, our results strongly support the idea of formation of functionally important NS4A homodimers through specific TM domain interactions.
Previously, Brass et al. used NMR to show that a peptide corre-sponding to the N-terminal 21 amino acids of NS4A can fold into an alpha-helical structure in the presence of several membrane mimetic compounds. Not unexpectedly, the NMR structure, which was deter-mined in 50% trifluoroethanol (TFE), aligns well with a single helix of the NS4A dimer predicted from the all-atom CHI simulation (8). However, interpeptide contacts were not reported in the NMR struc-ture, likely because TFE is an organic solvent that does not closely mimic the structure of a lipid bilayer. The association of TM alpha helices can be strongly affected by the chemical properties of sur-rounding lipids, solvents, and detergents (79–82). One advantage of the GALLEX system is that a biological membrane better approxi-mates the complex lipid bilayer environment of the ER than organic solvents or detergent micelles do.
Yang et al. recently detected small quantities of p14, a putative homodimer of NS4A, in genotype 1b replicon-bearing cells (83). The p14 form of NS4A was apparent only in the presence of ACH-806, an inhibitor of NS3-NS4A interaction (84), or when NS4A was overexpressed in the absence of NS3. These results suggest that p14 formation likely involves interactions with the central cofactor peptide rather than the NS4A TM domain. Furthermore, p14 was disrupted by 8 M urea but stable with respect to boiling in
Laemmli sample buffer, which is uncharacteristic of a TM inter-action (85,86). Thus, p14 may represent an aggregated form of NS4A. While the functional relevance of p14 remains unclear, the stability of the NS4A TM dimer characterized in this study strongly correlates with HCV replication and virus assembly.
An interesting ramification of an NS4A dimer is the effect that it might have on the NS3 protease/helicase it anchors into the membrane, which again differentiates it from p14. The NS3 heli-case domain has been crystallized as a monomer with and without RNA and other ATP analogs; the only exception is a cocrystal with a 16-mer single-stranded DNA long enough for two binding sites (29,33,87). Several biochemical studies on NS3 have hinted at either physical or functional oligomerization, although it is clear that the minimum functional unit is a monomer (88–91). The idea of functional oligomerization of NS3-4A units is appealing because NS3 exhibits a relatively low level of processivity for un-winding duplex RNA (92). NS4A dimerization could provide a way to increase the number of neighboring NS3 molecules and thereby enhance RNA unwinding efficiency. Indeed, NS3 may work in conjunction with NS4A to form a stable NS3-4A dimer and NS3-4A may have evolved the ability to pack favorably in a dimeric configuration in order to efficiently unwind HCV RNA.
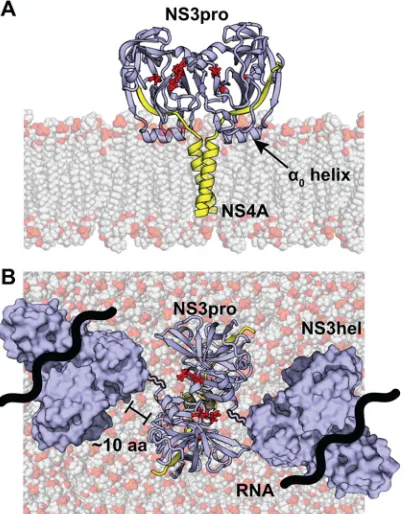
An NS4A dimer would bring NS3 molecules into very close contact because there is only a small two- or three-amino-acid linker between the TM domain and the NS3 cofactor region in NS4A. Moreover, the␣0helix of the NS3 protease domain fastens
NS3 to the membrane in a constrained configuration (8). To pro-vide epro-vidence that the NS3 protein could exist in a stable and sterically favorable configuration upon NS4A TM dimerization, we modeled the putative NS3 protease-4A dimer with the con-straints from the␣0helix of the protease domain and our
compu-tationally predicted NS4A TM domain dimer (Fig. 6A). This model predicts that the protease domain is anchored tightly against the membrane and is free to rotate only in the plane par-allel to the membrane surface. Previous structural and biochemi-cal work on NS3 from HCV, dengue virus, and Murray Valley encephalitis virus indicates that the⬃10-amino-acid linker region between the protease and helicase domains can accommodate a rotation of up to 180° between domains (93–95). This flexibility is sufficient for placement of adjacent NS3 helicase domains and would allow both domains to orient themselves above the mem-brane (Fig. 6B). Since NS4A stabilizes NS3 and activates its en-zyme activities (6), we predict that NS4A dimerization may lead to dimerization of NS3-4A. However, we cannot rule out the possi-bility that NS4A dimerization occurs independently of an interac-tion with NS3 (i.e., between one molecule of NS3-4A and one molecule of NS4A or between two NS4A molecules).
In addition to NS3-4A, oligomerization has been reported for all other HCV NS proteins, including p7 (96,97), NS2 (27,98), NS4B (99,100), NS5A (31,32,101), and NS5B (102,103). NS4A dimer formation might be required in order to allow NS3-4A to make contacts with other oligomeric viral protein complexes dur-ing replication or virus particle assembly. In fact, our cell culture replication and infectivity data highlight a few positions—L5, A6, and V9 —that do not appear to be part of the NS4A dimer inter-face and that may be important for additional packing interac-tions with TM domains of the other NS proteins. One future goal is to establish whether the NS4A monomer-dimer equilibrium shifts between the stages of replication and viral particle assembly or if NS4A is a stable dimer throughout the viral life cycle.
on November 7, 2019 by guest
http://jvi.asm.org/
Several systems have been developed to study the complete life cycle of HCV in cell culture, including functional cDNA clones for HCV genotypes 1a (strains H77 and TN), 1b (strain NC1), 2a (strains JFH-1, JFH-2, and J6), and 2b (strain J8) (77,78,104– 109). With the exception of strain JFH-1, most HCV isolates re-quire adaptive mutations for efficient replication in cell culture. Li et al. (77) recently identified three key replication-enhancing mu-tations during cell culture adaptation of genotype 2a strain J6. Notably, these adaptive mutations led to high levels of replication and infectious virus production and could be used to adapt genet-ically distinct genotype 2b (strain J8) and genotype 1a (strain TN) isolates (77,78). Of particular interest for our study was the iden-tification of the adaptive mutation NS4A A15S (residue A1672 in HCV strain H77), one of the three amino acid positions we define as the core of the NS4A dimer. Previous work on the GxxxG motif revealed that glycine-to-serine substitutions have a higher pro-pensity to form dimers than glycine-to-alanine substitutions in the context of the GpA TM domain (39). In addition, serines have been shown to help tightly pack two helical TM domains as evi-denced by the serine zipper motif (110). Based on these consider-ations, we hypothesized that NS4A A15S increases the dimeriza-tion affinity of the NS4A TM domain, leading to higher levels of cell culture replication and assembly. To that end, we discovered that NS4A A15S caused a subtle increase in GALLEX dimerization
relative to WT NS4A (see Fig. S7 in the supplemental material), rescued GALLEX defects of two NS4A mutants—G8A and A14L
(Fig. 5B)—and rescued cell culture defects of three NS4A
mu-tants—A6L, G8A, and A14L (Fig. 5CandD). Although our NS4A dimer model predicts that positions 6, 8, and 14 reside outside the dimer core, each position is still expected to provide an energetic contribution to the overall stability of the NS4A dimer (Fig. 3A). Thus, NS4A A15S may rescue virus replication and assembly by strengthening the NS4A TM dimer.
While the consequences of stronger NS4A dimer formation need further study, it is important to note that WT J6 and TN genomes are infectious in chimpanzee liver but unable to replicate in the Huh-7 cultured human hepatoma cell line (111,112). Thus, the three cell culture-adaptive mutations identified by Li et al. (77,
78), including NS4A A15S, may influence critical virus-host inter-actions. Given that NS3-4A cleaves the MAVS antiviral signaling molecule at the ER-mitochondrion interface (113), one intriguing possibility is that the stability of the NS4A dimer may influence trafficking of NS3-4A to this compartment.
Interestingly, a new computational tool was successfully used to design peptides that target and disrupt GxxxG-mediated inter-actions between␣IIb-/␣v- integrins and the3- integrin (114). The
sequence of the NS4A TM domain is very well conserved across genotypes and could potentially be targeted by these “peptide in-terceptors” for future therapies with broad effect (115). In partic-ular, our results suggest that the conserved “GGVLA” sequence in the NS4A TM domain may be an optimal target for this strategy. Our report presents the first evidence of homodimerization be-tween NS4A TM domains, which we show to be essential for HCV replication and virus particle assembly. Conceptual understanding of the quaternary structure within the HCV replication complex has been somewhat overlooked since the advent of an infectious HCV cell culture system. This inattention is unfortunate because so little is known about how each piece of the replication complex interacts, and, to date, attempts at reconstitution of the HCV replication com-plex from its individual components or exogenous RNA replication from isolated crude replicase extracts have been unsuccessful. The evidence for an NS4A TM dimer strongly suggests that more atten-tion should be given to understanding how the TM domains interact during the HCV virus life cycle.
ACKNOWLEDGMENTS
We thank S. Somarowthu for help with the computational dimer model and F. Yang for help with the cell culture experiments.
This work was supported by U.S. Public Health Service grants AI089826 (to B.D.L. and A.M.P.) from the National Institute of Allergy and Infectious Diseases and GM073857 (to D.M.E.) from the National Institute of General Medicine. A.M.P. is an Investigator of the Howard Hughes Medical Institute.
REFERENCES
1.Lindenbach BD, Murray CL, Thiel HJ, Rice CM.2013.Flaviviridae, p
712–746.InKnipe DM, Howley PM (ed), Fields virology, 6th ed, vol 1. Lippincott Williams & Wilkins, Philadelphia, PA.
2.Simmonds P, Bukh J, Combet C, Deleage G, Enomoto N, Feinstone S,
Halfon P, Inchauspe G, Kuiken C, Maertens G, Mizokami M, Murphy DG, Okamoto H, Pawlotsky JM, Penin F, Sablon E, Shin IT, Stuyver
LJ, Thiel HJ, Viazov S, Weiner AJ, Widell A.2005. Consensus
propos-als for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology42:962–973.http://dx.doi.org/10.1002/hep.20819.
3.Nakano T, Lau GM, Lau GM, Sugiyama M, Mizokami M.2012. An
updated analysis of hepatitis C virus genotypes and subtypes based on the
FIG 6Model of the putative NS3-4A dimer in the membrane. The NS4A
dimer generated by CHI was modeled in a homogenous membrane of POPC. The NS3 protease domain (blue) was attached to each NS4A TM domain (yellow) and modeled onto the membrane with the structural constraints im-posed by the␣0helix. Due to structural constraints from the POPC membrane,
NS4A dimer, and␣0helix, the NS3 protease domains would have only one
plane of rotation parallel to the surface of the lipid membrane. (A) Side view of the NS3-4A model in a POPC membrane. Serine protease active site residues are shown in red. (B) Top view of the NS3-4A model in a POPC membrane. A surface representation of the NS3 helicase domain is shown with a flexible linker that is approximately 8 to 10 amino acids in length. The approximate path of RNA through the helicase is shown.