0022-538X/84/090604-07$02.00/0
Copyright © 1984, American SocietyforMicrobiology
Proteins Encoded by
the
Long
Terminal
Repeat
Region
of
Mouse
Mammary
Tumor
Virus:
Identification
by
Hybrid-Selected
Translation
JANIS RACEVSKIS* ANDOM PRAKASH
MemorialSloan-Kettering Cancer Center, New York, New York 10021 Received14March 1984/Accepted 15 May 1984
The long terminal repeat(LTR) regionofmouse mammarytumor virus (MMTV) isknown tocontain an
open reading frame of sufficient length to code for a protein of 36,000 Mr. The coding capacity of the 3'
sequencesofMMTV genomicRNA hasbeen demonstrated by in vitro translation studies,which have reported
the synthesis of four related proteins: p36, p24, p21, and p18. These proteins are overlapping translation products of thesame openreading frame,with the smalleronesinitiatingatinternal methioninecodons. From thepredicted amino acidsequenceofthe LTR protein,wehave selectedaregion likelytobe antigenic, obtained a synthetic peptide ofthat region, and raised antiserum to the peptide. The antipeptide serum specifically
immunoprecipitated allfour proteins from in vitro translated genomic 3' MMTV RNA,plusanadditionalone
of32,000 Mr. PublishedsequencedataofMMRV LTRs showaninternal AUG codonataposition which could
initiate a protein of 32,000Mr. The three smaller in vitro translation products (p24, p21, and p18) were
consistently synthesizedinmuchgreater amountsthan the p36orp32protein. The relativeamountof each in vitrosynthesizedproteinfromgenomic MMTV RNA could be predicted andwasin goodagreementwiththe postulatedeffectof flanking nucleotidesontheefficiencyof the respective AUG initiation codon. Polyadenylated
RNAs,isolated from variousmousetissues, wereselected by hybridizationtoplasmid DNA containing MMTV
LTR sequencesimmobilized on nitrocellulose. In vitro translation ofhybrid-selected mRNAsisolated from
BALB/cmouse lactatingmammary glands andcarcinogen-induced mammary tumors, followed by immuno-precipitation with antipeptideserum,revealed that onlyonepolypeptidewassynthesizedby theMMTV
LTR-specific mRNA, the 36,000 Mr species.
Thelong terminalrepeat(LTR) ofmousemammarytumor virus (MMTV) is unique among retroviral LTRs in that it containsalongopenreading frameinitsU3 region(30).The
presenceofan open reading frame in the MMTV LTRwas
discovered byDNAsequencing of the proviralLTR(7) and byinvitro translation ofthe3' end ofgenomicMMTV RNA (4, 28).Theopenreadingframeinthe MMTVLTRhasbeen foundtobeconservedinall MMTV strainsanalyzed todate (6, 7, 10, 13, 17) and appears to have a coding capacity sufficienttocodefora36,000 Mr protein. Invitro translation studies (4, 22, 28) have reported that four polypeptides, unrelated to any MMTV structural proteins, could be syn-thesized from the 3' end ofgenomicMMTV RNA: polypep-tidesof36,000, 24,000, 21,000,and18,000 Mr. Thesamefour polypeptides were also observed after translation of RNA transcribed from cloned MMTV LTR sequences (5), thus firmly establishing their viral origin. The in vitro protein products of the LTR appear to be overlapping translation productsof thesameopenreading frame(4, 22, 28),with the smaller polypeptides probably initiating at internal AUG codons. In the in vitro translation systems, the three smaller polypeptides (24,000, 21,000, and 18,000 Mr) are always
synthesized in greater amounts than the 36,000 Mrprotein (4, 22, 28).
Search forexpressionof the MMTVLTRgeneinvivohas revealedthepresence, incertainmouse tissues, ofaspliced 1.6-kilobase (kb) mRNA species hybridizable with MMTV
LTRsequences (33).We have raised antiseratothe MMTV
LTR gene translation products, using a synthetic peptide
*Correspondingauthor.
604
predicted by the DNA sequence, and have found that a
36,000Mrprotein precipitablewith the antiserum is synthe-sized in vitro from hybrid-selected MMTV LTR-specific, 1.6-kb mRNA isolated from lactating mammary glands of BALB/c mice and 7,12-dimethylbenz(a)anthracene (DMBA)-induced mammary tumorsof BALB/c mice.
MATERIALS ANDMETHODS
Antipeptideserum.The 15-aminoacid-longsynthetic pep-tide was purchased from Peninsula Laboratories and was
coupled to the carrier protein keyhole limpet hemocyanin with the couplingreagent m-maleimidobenzoyl-N-hydroxy-succinimide esteraccording to the procedures of Lerneret al. (15). Rabbits were immunized with the peptide-keyhole limpet hemocyanin conjugate according to the following schedule (15): 500 p.g emulsified in complete Freund adju-vantsubcutaneouslyonday 0; 500 p.g inincomplete Freund
adjuvantondays 14,21,and 91. Animalswerebled 15 weeks after the firstinjectionandat2-monthintervals after booster
injections.
Activity of the antiserum wastested by immunoprecipita-tion of[35S]methionine-labeled in vitro translation products ofC3H MMTVgenomic RNA. MMTV C3H virus, isolated fromcell line Mm5mt/cl(20),wasusedasthesourceof viral RNA andwas provided by the Intramural Viral Resources Program, National Cancer Institute. Sucrose gradient-puri-fied35S viral RNAwas agiftfrom P. Etkind of thisinstitute.
Viral RNA was translated in vitro in a rabbit reticulocyte
lysate system (New England Nuclear) in the presence of
[35S]methionine (New England Nuclear). The immunoglob-ulin G (IgG) fraction of the highest-titer antiserum was
on November 10, 2019 by guest
http://jvi.asm.org/
MMTV LTR PROTEINS 605
isolated
by
affinity chromatography
on a column of thesynthetic peptide coupled
toCH-Sepharose
4B(Pharmacia
Fine
Chemicals, Inc.),
allasdescribedby
Walteretal.(32).
Immunoprecipitation. Initial
screening
of antisera wasperformed by adding
5 to10 ,ulofserum to analiquot of in vitro translatedMMTVproteins
inreticulocyte lysate, dilut-edinlysis
buffer(26):
20mMTris-hydrochloride (pH 7.5),
50 mMNaCl,
0.5% NonidetP-40,
0.5% sodiumdeoxycholate.
Immune
complexes
wereprecipitated
withprotein
A-Se-pharose (Pharmacia)
asdescribedpreviously
(26).Immuno-precipitates
wereanalyzed by electrophoresis
on5 to20%
exponential gradient polyacrylamide gels
followedby
fluo-rography
of thegels (24).
A more
quantitative
recovery of MMTVLTR-specific
proteins
wasobtainedby using
anindirectimmunoprecipita-tion
technique (2).
Antiserum- oraffinity-purified IgGs
wereinitially
preadsorbed
to a10%(vol/vol) Sepharose-protein
Asuspension
inphosphate-buffered
saline (pH7.2)-0.5%
Tween 20-0.1% bovine serumalbumin
(PTA)
for 2 hat4°C with rotation. TheSepharose-protein
A was then washed with PTA andaliquoted
intosamples
tobeanalyzed,
which were then allowedto reactovernight
at4°C
with continual rotation. TheSepharose-protein
Aantigen complexes
were then washedwith PTA andanalyzed
asdescribed above.RNA
purification.
RNAwas isolated fromlactating
mam-maryglands
(LMG)
and livers ofBALB/c mice(Jackson
Laboratories),
LMG of GR mice(Institute
for MedicalResearch), transplantable
DMBA-inducedmammarytumors of BALB/c mice(29)
(obtained
from N.Telang
of thisinstitute),
andtransplantable
leukemiccells
of DBA/2 mice(24).
RNA was extracted from tissuesby
theguanidinium
isothiocyanate-cesium
chloride method(18).
Tissues werefrozen in
liquid nitrogen, suspended
in 8 volumes of 6 Mguanidinium isothiocyanate-5
mMsodium
citrate(pH
7.0-0.1 M3-mercaptoethanol-0.5%
Sarkosyl,
and thenhomoge-nized inaSorvall Omnimixer. Addition of cesium chlorideto the
homogenates,
layering
on cesium chloridecushions,
centrifugation,
and RNA extraction were allperformed
asdescribed before
(18). Polyadenylated [poly(A)+]
RNAwas selectedby
passage over anoligodeoxythymidilic
acid-cellulose column aspreviously
described(28).
Analysis
ofpoly(A)+
RNAswasperformed
withformaldehyde-contain-ing
agarosegels (27),
with modificationsaccording
to Pra-kashetal.(23).
RNAswereelectrophoresed
in 1.4%agarosegels
containing
MAEbuffer(20
mMMOPS[morpholinepro-panesulfonic acid],
pH 7.0,
5 mM sodium acetate, 1 mMEDTA),
2.2 Mformaldehyde,
and0.5 ,ug of ethidium bro-mideper ml. RNAsamples,
10p.g,
weredenatured in 25pul
ofMAE buffer
containing
5%formamide,
2.2 Mformalde-hyde,
and0.02%
bromophenol
blueby heating
at60°C
for2 min beforeloading
on thegel.
Theelectrophoresis
was carried out for 3 h at 150V,
using
MAE buffer in the reservoirs. Thereservoir buffer wascirculated to eliminate concentrationgradients.
Thegel
was soaked for 30 min in 20x SSC(SSC
= 0.15 MNaClplus
0.015M sodiumcitrate)
before
photographing
underUVlight,
and then the RNAwas transferred to nitrocellulose(31).
Hybridizations
were car-ried out in 50%formamide-containing
solutions at 42°Caccording
to Thomas(31).
Labeled,
nick-translated probe was prepared fromthecloned 1.4-kb PstI DNAfragment of the 5' MMTV LTRregion
(16)(kindly provided
byJ. Majors and H.Varmus).
Hybrid selection.
Hybrid
selection was performedessen-tially
as describedby
Pachl et al. (21). The plasmid DNA used forpreparation
of filterswasthepBR322clonecontain-ing
the 1.4-kb PstIfragment
from the left end of the C3HMMTV DNA
(16).
Theplasmid
DNA was linearizedby
cleavage with EcoRI, extracted with phenol-chloroform, and ethanolprecipitated. A 4-,ugamountoflinearized, denatured plasmid DNA was
spotted
per 6-mm nitrocellulose filter circle. Afterwashing
andbaking
thefilters, theywereplaced inhybridization
solution: 50%formamide,
0.4 MNaCl,
10 mMPIPES[piperazine-N,N'-bis(2-ethanesulfonic acid)],
pH 6.4, 0.2% sodiumdodecyl sulfate, 100 ,ugof calf liver tRNA per ml.Poly(A)+
RNA wasadded,
andhybridization
was carriedoutin avolume of 0.1 mlat42°C
for 18 h. The filters werewashed,
and the RNAwaseluted as describedprevi-ously (21).
The RNAwascoprecipitated
from ethanol with 5 ,ug of calf livertRNA, washed,
and translated in a rabbitreticulocyte
cell-freesystem. RESULTSAntipeptide serum. From the
predicted
amino acid se-quence of the C3H exogenous MMTV LTR openreading
frame(7), wechose a14-amino
acid-long region
ashaving
ahigh probability
ofbeing
antigenic
on the basis of itshigh
average amino acid
hydrophilicity
value. Inchoosing
thisregion
we followed the method ofHopp
and Woods(12)
in whichlikelyantigenic
sitesareidentifiedby locating regions
of
high
hydrophilicity
in the amino acidchain, by assigning
numerical
hydrophilicity
values to all constituent amino acids.Regions
containing
thehexapeptide
with thehighest
average
hydrophilicity
valuewerefoundtobeantigenic
in 12 of 12proteins
studied(12).
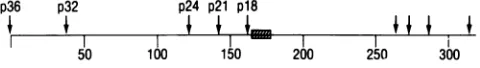
The 14-amino acid sequence isspecified by
codons 165through
178(numbering
the codonsstarting
with theAUG atthebeginning
of the openreading
frame
[17];
Fig. 1)
and has the sequence:Ile-Glu-Asn-Arg-Lys-Arg-Arg-Ser-Thr-Ser-Ile-Glu-Glu-Gln.
Thispeptide
is encodedby
thesequencebeginning
twocodons downstream fromthefourth internalAUG codon(6),
aregion
thatwecan deduce codes for the amino-terminalendofthe18,000 Mr in vitro translation product (Fig. 1). As it turns out, this sequence ishighly
conserved and thehydrophilic
hexapep-tide166-171 (Glu-Asn-Arg-Lys-Arg-Arg)
is identical in all fourknownMMTV sequences(6,7, 10, 13, 17).Only single
amino acid
differences,
all in thecarboxy-terminal
halfof the chosen 14-amino acid sequence, are present in the foursequenced
MMTVstrains. Anequallyhydrophilic
hexapep-tide is located atposition
267-272; however, the adjacent amino acidsarelesshydrophilic
thanthose flanking peptide 166-171.A 15-amino
acid-long
synthetic peptide containing the chosen 14-amino acidsequenceplusanoncodedcysteineat thecarboxy
terminus, for attachment to a carrier protein, wasobtained. The synthetic peptidewas coupled to keyholelimpet hemocyanin
via thecysteine residue with the bifunc-tional reagent m-maleimidobenzoyl-N-hydroxysuccinimide ester,following
the published procedures ofLerner et al. (15). Rabbits wereimmunized with the peptide carrier pro-teinconjugate according
tothe protocol of Lerneretal.(15),p36 p32 p24 p21 p18
-50 100 150 200 250 300
FIG. 1. Schematic representation of the 319-codon-long open reading frame ofthe C3H exogenous MMTV LTR, according to
Donehoweret al. (6). Numbers onbottom denote codon number, andarrowsindicatepositionof AUG codons. In vitro LTR transla-tion products are marked above their putative initiation codons. Shadedbox indicates location ofsynthetic peptide (codons 165 to
178). VOL.51, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.316.556.630.664.2]and sera were tested for activity byimmunoprecipitationof the in vitro translation products of exogenous C3H MMTV genomic RNA. After a 15-week immunization protocol, all three rabbits developed some precipitating antibodyactivity against the in vitro LTR proteins (pLTRs). One of the rabbits developed a considerably stronger response than the other two, and all subsequent experiments, including affinity puri-fication of IgGs, were performed with serum from this one animal. Further booster injections were also given, and higher-activity serum was obtained. In vitro translations were performed in a rabbitreticulocyte lysate system, with [35S]methioninelabeling and using a 35S fraction of genomic MMTV RNA as a template. The viral RNA used in these studies was isolated from exogenous C3H strain virus pro-duced by cell lineMm5mt/cl(20). Awhole range of proteins weresynthesized from this template (Fig. 2, lane A), includ-ing the high-molecular-weight gag-pol and gag precursors (4, 28) of 160,000, 105,000, and 75,000
Mr,
which were precipitated withanti-MMTVcoreprotein p28 serum (Fig. 2, laneD).Immunoprecipitationwith ananti-pLTR serum from an early bleeding showed that at least four proteins wereA
B
C
D
-160
--
-105
Z*-75
A
B
C
D
E
-MmiW
-
36
-
32
__ __
__ __
_b
4UNP-24
_,. 21
21
.
1
8
FIG. 3. [35S]methionine-labeled in vitro translationproducts of MMTV35Sgenomic RNA(lane A), immunoprecipitated with:(B) nonimmune serum; (C) anti-pLTR peptide serum; (D) anti-pLTR peptide serum in the presence of 5 p.g of synthetic peptide; (E) affinity-purifiedIgG fraction ofanti-pLTRpeptide serum.
36
32
24
-_i-21
-28
18-FIG. 2. [35S]methionine-labeled in vitro translation products of
C3H MMTV 35SgenomicRNA(lane A), separatedona
polyacryl-amide gel, immunoprecipitated with: (B) nonimmune serum; (C) anti-pLTR peptideserum;(D)anti-MMTV coreproteinp28serum.
Molecularweights ofproteins (inthousands)aremarkedonsides.
specifically precipitated (Fig. 2, lane C). Immunoprecipita-tionof the in vitroproducts withalater, higher-titerserum as well as withaffinity-purified IgGof thisserum, conclusively demonstrated the specificprecipitation of the four polypep-tides, plus that ofafifthone of32,000 Mr (Fig. 3, lanes C and E). In general, it was observed that equally good results were obtained whether unfractionated serum or a purified IgG fraction was used for immunoprecipitation with the indirectimmunoadsorption technique (2). The precipitation of these fivepolypeptidescould becompletely abolishedby theaddition of a fewmicrogramsof the synthetic peptideto act as a competitive inhibitor (Fig. 3, lane D). Addition of synthetic peptide hadnoeffectonimmunoprecipitation with anti-coreprotein p28serum(datanotshown). Thesynthesis ofthe32,000 Mr proteinwasprobablynotpreviously report-ed (4, 22, 28) because it is synthesized in relatively low levels. Examination of thepublishedMMTV LTRsequence data (6, 10, 13) shows that an AUG codon is present in all four MMTV strains at a position (codon 38) which could initiate apolypeptideof32,000 Mr.The oneexceptionisthe
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.329.559.76.463.2] [image:3.612.80.292.296.680.2]MMTV LTR PROTEINS 607
sequence of the C3H viruspublishedby
Majors
andVarmus (17), which shows a valinecodon(GUG)atposition38 ofthe LTR openreadingframe. This result(17)isatvariance with the sequence published by Donehower et al. (6) since both studies were performed with exogenous C3H virusisolated from the same cell line: Mm5mt/cl (20). The C3H MMTV LTR open reading frame contains four additional AUG codons located near the 3' end of the openreading
frame. Any peptidesinitiatedatthese codonswouldnotbe detected with ourantipeptide serum since these codons are located downstreamfromthe chosenpeptidesequence. The relative amountsofeachofthefivepolypeptides synthesizedin vitro were observed to be consistentfrom experiment to experi-ment. The three smaller polypeptides (24,000, 21,000, and 18,000 Mr) were always synthesized in greater quantities than the36,000Mrprotein and the32,000Mrspecies,which wasalwaysthe leastabundant(Fig.4).Taking into consider-ation the factthat thedensitometerscan(Fig. 4) isameasure ofincorporated[35S]methionine,
the actualdisparity
in the relative amounts of synthesis of these proteins is even greater, since the smallerpolypeptides have fewer methio-nineresidues. Inaddition, sincetheproteinswere separated on apolyacrylamide gradient gel, there is a direct relation-ship between the molecular weight of a protein and the efficiencyofdetectingafluorographic signal fromthe labeled protein, because the smaller proteins are embedded in the thickerpartofthedried gel wheresignals arequenchedto a greater degree (11). The ratio ofthe five LTR proteins to each other is the same in the translation mix(Fig. 3, lane A) as it is in the immunoprecipitates (Fig. 3, lanes C and E), demonstrating that the antiserum has equal avidity for all five polypeptides. Anattractive explanation forthedifferent levelsofsynthesis ofthefiveLTRproteins is the postulated effect offlanking
nucleotides on theefficiency
ofan AUG codon to act as an initiation site (14). Analysis of the published sequence ofMMTVLTRs shows thattheputative initiative codon for the 32,000 Mr protein, which is least abundant, is in the least favored of all possible arrange-ments, with apyrimidine in the -3 position:YXXAUGX-Hybrid-selected translation. Various mouse tissues were analyzed for the presence of the reported MMTV LTR-specific, spliced, 1.6-kb mRNAspecies(33) byhybridization ofNorthern blots ofextracted poly(A)+ RNAs with cloned MMTV LTR probes. A specific band migrating faster than 18S rRNA could be detected in blots ofRNAs extracted from lactating BALB/c mousemammaryglands (Fig. 5, lane B) and DMBA-induced mammary tumors ofBALB/c mice (Fig. 5, laneC). NootherMMTV-specific mRNAscould be detectedinthesetissues, which isconsistentwith the report that carcinogen DMBA-induced mammary tumors of BALB/c mice do not express detectable levels of any MMTV structural proteins (9). No MMTV structural
pro-FIG. 4. Densitometertracingof laneC, Fig. 3.
Immunoprecipi-tate ofMMTV RNAtranslation products with anti-pLTR
peptide
serum.
A
B
C
D
E
3524
S-O-P* so" -
7.8 Kb
_."a
% -3.8 Kb
~
~~~~
-1.6 Kb
I
BALB/c
DDBA/2
GRFIG. 5. Northern blot analysis of poly(A)+ RNAs extracted from: (A) BALB/c mouse liver; (B) BALB/c LMG; (C) DMBA-induced BALB/cmammary tumor; (D) DBA/2 T-cell lymphoma;(E) GR LMG. A10-,ug portionofpoly(A)+ RNA (passed oncethrough an oligodeoxythymidylate column) was applied to each lane. Hy-bridization was done with the 1.4-kbPstI fragment containing the MMTV LTRsequence. The film was exposed for 48 h, except for laneD, whichwasexposed for 12 h [the RNA in lane D was twice poly(A) selected].
teins could be detected in the DMBA-induced mammary tumors or in LMG of BALB/c mice by the technique of protein blotting and reaction with anti-MMTV serum
(data
not
shown). Analysis
ofGRmouseLMG RNA (Fig. 5,lane E) revealed the presence of the two RNA species usually found inMMTV-producing
cells (25): thegenome-length,
35S, 7.8-kb-long RNA,
and the 24S, 3.8-kb-longenvelope
geneproduct.Noadditional, smaller, MMTV-specific RNAs could be detected in the GR LMG, even after
prolonged
exposures. A number of MMTV-specific RNAs were ob-served in the sample extracted from the MMTV
antigen-expressing(24) T-cell lymphomaofDBA/2mice(Fig.5, lane D).Inadditiontothe35SRNA andabandmigratingslightly faster than
24S,
there appear to be two lower-molecular-weight species:oneofabout 1.6 kb and another still smaller (Fig. 5,lane D).MMTV-specificmRNAs were selectedbyhybridizationto cloned MMTV LTR sequences immobilized on nitrocellu-losefilters asdescribed in Materials and Methods. Hybrid-ized mRNAswereelutedby boilingand thenethanol precip-itated, washed, andtranslatedin arabbitreticulocyte
lysate
in vitro protein translation system. The [35S]methionine-labeled products of in vitro translated mRNAs were immu-noprecipitatedwithantipeptideserum aswellas apolyclonal
24K
18K
36K 21K
32K VOL. 51, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.314.559.77.394.2] [image:4.612.67.274.612.703.2]608 RACEVSKIS AND PRAKASH
anti-MMTV serum, and the precipitates were analyzed on polyacrylamide gels (Fig. 6). In addition to the DMBA-induced mammary tumors and LMG of BALB/c mice, we also analyzed LMG of GR mice and T-cell lymphomas of DBA/2 mice. LMG of the high mammary tumor incidence strain GR produce large quantitiesofvirus(1), and theT-cell lymphomas of DBA/2 mice express high levels ofMMTV antigens, although they do not secrete mature virions (24). The most striking result was that only one LTR-specific protein, the 36,000
Mr
species, was synthesized from the mRNAsof the BALB/c LMG and mammary tumors (Fig. 6, lanes C and G). Precipitation of the 36,000 Mr translation product was completely abolished by the addition ofa few micrograms of synthetic peptide (Fig. 7, lanes D and F). Synthesis of other MMTV-specific proteins could not be detectedin thetranslation mixtures of mRNAs isolated from BALB/c tissues (Fig. 6, lanes DandH). The pLTR pattern obtainedfrom the translation of mRNAs of GR LMG (Fig. 6, lane A; Fig. 7, lane A) was very similar to patterns obtained from translation of genomic RNA, probably reflecting the fact that most if not all of the MMTV LTR translation occurred from genomic RNA. The "p36" LTR protein of GRvirus appearedtohave a slightly higher molecular weight than theanalogous protein ofBALB/corC3H. An interest-ing resultwas obtained fromtranslation of mRNAs isolated from DBA/2 T-cell lymphomas: the predominant LTR-spe-cificprotein
was the 32,000 Mr polypeptide (Fig. 6, lane E; Fig. 7, lane G). Immunoprecipitation with polyclonal anti-MMTV serum showed that awhole range of MMTV struc-turalproteins
and precursors were synthesized by the po-ly(A)+ RNAs selected from GR LMG and DBA/2 lymphomas(Fig.
6, lanes B andF).
A B
C
D
E
F
G
F
*...
{.
}hss tjUi .... ;|-sW*. =, .i..*
EMs...
'
._B*.
.. _E
L
..
., s
. ::
H I
i.
-36
-32
,- 24 F 21
- 18
GR BE/c DBA/2 B/c
FIG. 6. Analysis of in vitro translation products encoded by hybrid-selectedRNAsofGRLMG(lanesAandB);DMBA-induced mammarytumorof BALB/cmouse(lanesC andD);transplantable
T-celllymphomaofDBA/2mice(lanesEandF); LMG ofBALB/c mice(lanesGandH). Translationproductswere immunoprecipitat-ed with anti-pLTR peptide serum (lanes A, C, E, and G) and
polyclonal anti-MMTVserum (lanes B, D, F,and H). Lane I isan
aliquotof in vitrotranslationproducts.
A
B
C
D
E
F
G
H
36--
-
32-? 4 4
_8
rGR B u
Blc
DBAff2
FIG. 7. Analysis of in vitro translation products encoded by hybrid-selected RNAs of: GR LMG (lanes A and B); DMBA-inducedmammary tumor ofBALB/cmouse(lanes C andD);LMG ofBALB/c mice (lanes Eand F);T-celllymphomaof DBA/2 mice (lanes Gand H). Thetranslation productswereimmunoprecipitated with anti-pLTRpeptide serum(lanesA,C,E,andG)andanti-pLTR peptideserumin thepresenceof 5
pg
ofthesynthetic peptide(lanes B, D, F, andH).Immunoprecipitation
analysis, with anti-p-LTRserum, of extracts of in vitro labeled primary cultures of LMG and mammary tumors, orfreshly excised minced tissues,yielded
equivocal results because of high background levels and cross-reactions with unrelated
proteins.
Cross-reactions with otherwise unrelatedproteins, especiallywithabundant ones,havebeen observedwhenantipeptide
seraareused toanalyze whole-cellextracts(19).Thisphenomenonis
proba-blybecause antibodies to a singlelinear sequence will also recognize with some affinity a closely related linear se-quence which might be present by chance on
proteins
that are otherwise unrelated (19). Antiseraprepared
against
synthetic peptides have weak
precipitating activity
and do noteffectquantitative
recoveryofantigens,
eveninantibody
excess; thus, resolution of this problem will have to await the production of hyperimmune anti-pLTR
polyclonal
se-rum.Hybrid-selected translation analysis was also
performed
on RNAs extractedfrom tissue culture cellsof cell lines of normal BALB/c mammary
glands
as well as mammary tumors; no LTRproteins could be detected.DISCUSSION
To search for expression ofthe MMTV LTR gene
prod-uct,weraised antiserumagainsta
synthetic
peptide
predict-ed by the DNA sequence of the C3H exogenous MMTV LTR open reading frame (6, 7, 17). The chosen 14-aminoJ. VIROL.
4'W
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.323.561.74.374.2] [image:5.612.67.303.405.658.2]MMTV LTR PROTEINS 609
acid sequence lies near the presumed amino terminus of the 18,000 Mr in vitro translation product (4, 22, 28). The antiserum was found toimmunoprecipitateallfour previous-ly reported (4, 22, 28) LTR translation products, plus low levels of a fifth one of 32,000 Mr.
Invitro translation ofgenomic MMTV 35S RNA always produced the five LTR proteins in the same quantitative ratio,withthe three smallerpolypeptides(p24,p21, andp18) being the most abundant. In terms of total amount of synthesis, as measured by
[35S]methionine
incorporation, the five LTR polypeptides were always synthesized in the following relative order: p24 > p18 > p21 > p36>p32.The difference between themost abundantprotein (p24)and the leastabundant one (p32) was at least 20-fold (Fig. 4). Ifthe observedquantitative ratios ofthe invitroLTRproductsare caused by RNA degradation, then one would expect to observe alinear inverse relationship betweenthe molecular weight of each LTR protein and the quantity synthesized; this isnot the case. RNA analysis andin vitro synthesis of Pr759'l andPrlY"g-P"/
indicate thatdegradation ofRNAis minimal. Furthermore, the same pattern was also previously observed by Peters et al. (22) upon in vitro translation of threedifferent viral RNAs as well asfourdifferent plasmid-derived cRNAs. One of the most attractiveexplanationsfor theobserved variationin rates of translation isthe postulat-ed effectof flanking nucleotides on theefficiency ofagiven AUG codon to act as an initiation signal of eucaryotic translation, as put forth by Kozak (14). Kozak (14) has shown thatnucleotidesin the-3 and +4 positions relative to an AUG(3 xx codonexert apronounced effect on the efficiency of that codon and the frequency with which that codon is used for eucaryotic initiation of translation. In Table 1, the seven arrangements of initiation codons and nucleotides -3 and +4 are listed in decreasing frequency (topto bottom) of utilization as initiation signals in eucaryotic mRNAs (14). Listed alongside the appropriate codon ar-rangement is the MMTV LTR translation product which is assumed to beinitiatedon thatparticularcodonarrangement (6, 7, 17). Comparison ofTable 1 and Fig. 4 demonstrates that there is a very good correlation between the relative amount of in vitro synthesis of each LTR protein from a genomic RNA template and the relative efficiency of its respective initiation codon. The proposed initiation codon hierarchyisespecially successful atpredictingthe very low level of synthesis ofp32, whose initiation codon is in the least favored ofall arrangements. The selective importance of the initiation codon, however, holds onlyforthe in vitro translation ofgenomic MMTV RNA. If the natural mRNA is used astemplate, however, then only the 36,000 Mr polypep-tide is synthesized, asdemonstratedbyusing hybrid-select-ed LTR-specific mRNAs of BALB/c LMG and DMBA-induced mammary tumors. This result confirms previousTABLE 1. Initiation codonarrangementsindescendingorderof efficiency"andcorresponding MMTV LTRproteinsh
Protein(s) Codonarrangement'
AXXAUGG AXXAUG A
p21, p24 AXX AUG Y
p18 GXX AUG G
GXX AUG A
p36 GXXAUG Y
p32 YXXAUG X
a Kozak(14).
bDonehoweretal.(6).
' Y=pyrimidine;X =any oneofthe four ribonucleotides.
observations
(14)
thattheprimary
determinantofa function-alinitiationsiteofanmRNA is its proximitytothe 5' end of the message.Although
we cannot discount thepossibility
that the activated LTRin BALB/c tissues
only
contains the AUG forp36,
this seemsratherremote since the other four AUG codons have been conserved in all foursequenced
MMTV
strains;
furthermore, only
oneLTRproduct
appears to besynthesized
by
theDBA/2 LTR mRNAaswell.The
transcription
of the MMTV LTR gene into aspliced
mRNA(33),
whichwehave demonstratedtobeafunctional messagecoding
for a36,000 Mr
protein,
in two types of BALB/cmouse tissues indicates the probable activation of an endogenous provirus.The expression of MMTV LTR sequences in BALB/c mouse LMG wasforetold by earlier hybridizationstudies which detectedanelevated level of 3'-end MMTV RNA expression in these tissues (8). Since BALB/cmice donotcarryamilk-borneorexogenousvirus,
a possible source of the LTRproducts might be the incom-plete endogenous provirus unit I (3). Invitro translation of hybrid-selected mRNAs from MMTV-producing tissues such as GR mouse LMG does not give an answer as to whetherfunctional LTR mRNAs are present, since whatwe observe isprobably primarilytranslation ofgenomicMMTV RNA. In addition, no LTR-specific, small mRNA
species
could be detected in the GR tissues by Northern blot
analysis.
A similar analysis of RNAs isolated from aT-cell lymphoma of DBA/2 mice indicates the presence of two small, LTR-specific RNAs and yields the interesting obser-vation that only the 32,000 Mr LTR translation product is produced. The synthesis of a single product suggests the presence ofa functional LTRmRNA. This lymphoma cell lineproduces high levels of both MMTV gag and env gene products (24); it does not, however, produce mature virions. Thesynthesisof the32,000 Mr polypeptide perhapsindicates thatitsinitiationcodonin theDBA/2provirus differs or that the pLTR gene in theactivatedDBA/2provirus is shorteror lacks the first AUG codon.The possible function ofthe MMTV LTR gene
product
canonly be the subject of speculation at thistime;however, a tantalizing clue is our failure to detect LTR proteins in tissue culture cellsofBALB/c mouse mammary glands.One of the alterations in cells of mouse mammarygland tissue culturecell lines as compared to cells in the original tissue is loss of the ability to respond to certainhormones,including prolactin (34).
ACKNOWLEDGMENTS
We aregratefultoP.Etkindfor hergiftof thepurified viralRNA and to N. Telangfor the transplantable mouse mammary tumors. We thankR. Kopelman forexpert technicalassistance.
This work was supported by Public Health Service grant CA-16599 from the National Cancer Institute.
LITERATURE CITED
1. Bentvelzen, P., and J. Hilgers. 1980. The murine mammary tumor virus, p. 311-355. In G. Klein (ed.), Viral oncology. RavenPress, New York.
2. Bunol, T. F., and R. A. Reisfeld. 1982. Unique glycoprotein-proteoglycancomplex defined by monoclonalantibody on hu-manmelanoma cells. Proc. Natl. Acad. Sci. U.S.A. 79:1245-1249.
3. Cohen,J. C., J. E. Majors, and H. E. Varmus. 1979. Organiza-tion of mousemammary tumorvirus-specific DNA endogenous to BALB/cmice. J. Virol. 32:483-496.
4. Dickson, C., and G. Peters. 1981. Protein-coding potential of mouse mammary tumorvirus genome RNA as examined by in vitrotranslation. J. Virol. 37:36-47.
VOL.51,1984
on November 10, 2019 by guest
http://jvi.asm.org/
5. Dickson,C., R.Smith,and G. Peters.1981.In vitrosynthesis of
polypeptides encoded by the long terminal repeat region of
mouse mammarytumorvirus DNA. Nature (London) 291:511-513.
6. Donehower, L. A., B. Fleurdelys, and G. L. Hager. 1983. Fur-ther evidence for the protein coding potential of the mouse mammary tumor virus long terminal repeat: nucleotide
se-quenceofanendogenousproviral long terminalrepeat.J.Virol. 45:941-949.
7. Donehower, L. A., A. L. Huang, and G. L. Hager. 1981. Regulatory and coding potential of themouse mammarytumor virus long terminal redundancy. J. Virol. 37:226-238.
8. Dudley, J. P., J. M. Rosen, and J. S. Butel. 1978. Differential expression of poly(A)-adjacent sequences ofmammary tumor virus RNA in murine mammary cells. Proc. Natl. Acad. Sci. U.S.A. 75:5797-5801.
9. Dusing-Swartz, D., D. Medina, J. S. Butel, and S. H. Socher. 1979. Mouse mammary tumor virus genome expression in
chemical carcinogen induced mammary tumors in low- and high-tumor-incidence mouse strains. Proc. Natl. Acad. Sci. U.S.A. 76:5360-5364.
10. Fasel, N., K. Pearson, E. Buetti, andH.Diggelmann. 1982.The region ofmouse mammarytumorvirusDNAcontainingthelong terminal repeatincludesalong codingsequenceandsignalsfor
hormonally regulated transcription. EMBOJ. 1:3-7.
11. Harding, C. R., and J. R. Scott. 1983. Fluorographylimitations on its use for quantitative detection of 3H and '4C-labeled proteins in polyacrylamide gels. Anal. Biochem. 129:371-376.
12. Hopp, T. P., and K. R. Woods. 1981. Prediction of protein antigenic determinants from aminoacidsequences. Proc.Natl. Acad. Sci. U.S.A. 78:3824-3828.
13. Kennedy, N., G. Knedlitschek, B. Groner, H. E. Hynes, P.
Herrlick, R. Michalides, and A. J. J. van Ooyen. 1982. Long terminal repeats ofendogenous mouse mammary tumorvirus
containalongopenreadingframewhich extends intoadjacent
sequences. Nature(London)295:622-624.
14. Kozak, M. 1981. Possible rolesofflankingnucleotides in recog-nition of the AUG initiator codon by eukaryotic ribosomes.
Nucleic Acids Res.9:5233-5252.
15. Lerner, R. A., N. Green,H.Alexander,F.T.Liu, J. G.Sutcliffe, and T. M. Shinnick. 1981. Chemically synthesized peptides predicted from thenucleotidesequenceof thehepatitisB virus genome elicit antibodies reactive with the native envelope protein of Dane particles. Proc. Natl. Acad. Sci. U.S.A.
78:3403-3407.
16. Majors, J. E., andH. E. Varmus. 1981. Nucleotidesequencesat
host-proviral junctions for mouse mammary tumor virus. Na-ture(London) 289:253-258.
17. Majors, J. E., andH. E. Varmus. 1983. Nucleotide sequencing ofanapparentproviralcopyofenvmRNA definesdeterminants ofexpressionof the mousemammarytumor virus envgene. J. Virol. 47:495-504.
18. Maniatis, T.,E. F. Fritsch, andJ. Sambrook. 1982. Molecular
cloning: alaboratory manual. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
19. Nigg, E. A., G. Walter, and S. J. Singer. 1982. On the natureof crossreactions observed with antibodies directed to defined epitopes. Proc. Natl. Acad. Sci. U.S.A. 79:5939-5943. 20. Owens, R. B., and A.J. Hackett. 1972. Tissue culture studiesof
mouse mammary tumor cells and associated viruses. J. Natl. CancerInst. 49:1321-1332.
21. Pachl, C., B. Biegalke, and M. Linial. 1983. RNA andprotein encoded byMH2virus: evidence for subgenomic expressionof v-myc. J.Virol. 45:133-139.
22. Peters, G., R. Smith, S. Brookes, and C. Dickson. 1982. Conser-vation ofprotein coding potential in the long terminal repeats of exogenous and endogenous mouse mammary tumor viruses. J. Virol. 42:880-888.
23. Prakash, O., R. V. Guntaka, and N. H. Sarkar. 1983. Evidence foraprokaryoticpromoterin the murine mammary tumor virus long terminalrepeat.Gene 23:117-130.
24. Racevskis, J., and N. H. Sarkar. 1982. ML antigen ofDBA/2 mouseleukemias: expressionof an endogenous murine mamma-ry tumorvirus.J. Virol. 42:804-813.
25. Robertson, D. L., and H. E. Varmus. 1981. Dexamethasone induction ofthe intracellular RNAs of mouse mammary tumor virus.J. Virol. 40:673-682.
26. Schultz, A. M., E. H. Rabin, and S. Oroszlan. 1979. Post-translational modification of Rauscher leukemia virus precursor polyproteinsencoded by the gag gene. J. Virol. 30:255-266. 27. Schwinghamer, M. W., and R. J. Shepherd. 1980.
Formalde-hyde-containing slab gels for analysis of denatured tritium-labeledRNA. Anal. Biochem. 103:426-434.
28. Sen, G. C., J. Racevskis, and H. N. Sarkar. 1981. Synthesis of murinemammary tumorviralproteinsinvitro. J. Virol. 37:963-975.
29. Telang, N. T., M. R. Banerjee, A. P. Iyer, and A. B. Kundu. 1979. Neoplastic transformation of epithelial cells in whole mammary gland in vitro. Proc. Natl. Acad. Sci. U.S.A. 76:5886-5890.
30. Temin, H. M. 1981. Structure, variation and synthesis of retrovirus long terminalrepeat. Cell27:1-3.
31. Thomas, P.S. 1983. Hybridization of denatured RNA trans-ferred or dotted to nitrocellulose paper. Methods Enzymol. 100B:255-266.
32. Walter, G., M. A. Hutchinson, T. Hunter, and W. Eckhart. 1981. Antibodies specific for the polyoma virus middle-sized tumor antigen. Proc. Natl. Acad. Sci. U.S.A. 78:4882-4886. 33. Wheeler, D. A., J. S. Butel, D. Medina, R. D.Cardiff,andG. L.
Hager. 1983. Transcription ofmouse mammary tumor virus: identification ofacandidatemRNAforthelong terminalrepeat geneproduct. J. Virol. 46:42-49.
34. Yang, J., J.Enami, and S. Nandi. 1977.Regulation ofmammary tumor virus production by prolactin in BALB/cfC3H mouse
normalmammaryepithelial cells invitro.CancerRes. 37:3644-3647.
![FIG. 3.nonimmuneaffinity-purifiedMMTVpeptide [35S]methionine-labeled in vitro translation products of 35S genomic RNA (lane A), immunoprecipitated with: (B) serum; (C) anti-pLTR peptide serum; (D) anti-pLTR serum in the presence of 5 p.g of synthetic peptide; (E) IgG fraction of anti-pLTR peptide serum.](https://thumb-us.123doks.com/thumbv2/123dok_us/1421782.94896/3.612.329.559.76.463/nonimmuneaffinity-purifiedmmtvpeptide-methionine-translation-products-immunoprecipitated-presence-synthetic.webp)


