0022-538X/89/073016-10$02.00/0
Copyright© 1989, American Society for Microbiology
Functional Limits of
oriP,
the
Epstein-Barr Virus
Plasmid Origin of
Replication
THOMAS CHITTENDEN,' STEPHEN LUPTON,2 AND ARNOLD J.
LEVINE'*
Department ofBiology, Priniceton Univ,ersity, Princeton, Newv Jersey 08544-1014,1 andImmiunexCorporation,
Seattle, Washington 981012
Received 24 January 1989/Accepted 30March 1989
The Epstein-Barr virus (EBV) genome contains two cis-acting elements which are required for stable extrachromosomal plasmid maintenance in latently infected cells. The first consists of 20 30-base-pair (bp) repeats, eachof which contains a DNA-binding site for EBV nuclear antigen 1 (EBNA-1), the trans-acting
factorrequired for plasmid persistence. The second element is composed ofa65-bp dyadsymmetry,containing fourEBNA-1-binding sites. Deletionmutantswere constructed which reduce the number ofEBNA-1-binding
sites in the 30-bprepeats,alter the number of EBNA-1-binding sites in the dyad region, ortruncatethedyad element. The effect of the deletion mutations on plasmid maintenance was examined by transfecting
recombinant plasmids, containing both the mutated EBV sequences and a drug resistance marker, into D98-Raji cells. The plasmidsweretested for their abilityto generatedrug-resistant D98-Raji cell colonies and theircapacity tobe maintained in anextrachromosomal form without undergoing extensive rearrangements.
EBVplasmids with 12or 15 copies of the 30-bprepeatswerewildtypein bothassays.Plasmids with justtwo
orsixcopies of theserepeated elements failedto generate drug-resistant colonies atanormallevel, and normal
episomal plasmidswerenotdetected in theresulting colonies. Rare colonies of cells resulting fromtransfection
of these two- or six-copy mutants contained rearranged, episomal forms of the input plasmids. The
rearrangements mostoften produced head-to-tail oligomers containing a minimum of eight 30-bp repeated elements. Therearranged plasmidswere shown toberevertantforplasmid maintenance in that they yielded wild-typeor greater numbers ofdrug-resistant colonies and persisted atthe wild-typeor a greaterepisomal
copynumber. ByuseofanEBVplasmid that containedno30-bp elements,norevertantscould be isolated. One tofivecopies ofasynthetic linkercorrespondingtoaconsensus30-bp repeated element inserted intoaplasmid withno30-bp elementsnowpermitted thegenerationof oligomeric, episomal forms of themutant testplasmid. These experiments demonstratearequirement foraminimal number (sixtoeight copies) of the 30-bp repeated element. Deletions in the 65-bp dyad region had little or no effect upon the ability to generate enhanced numbers of drug-resistant D98-Raji colonies, indicating that the 30-bp repeated element is predominantly required for this phenotype. Plasmids bearing deletions in the 5' side of the dyadsequenceretainedthecapacity
toproduce stimulated numbers of drug-resistant colonies, but the extrachromosomal plasmids in these cells all contained structural rearrangements. In contrast, deletions impinging on the 3' side of the dyad element
completely abolished extrachromosomal plasmid maintenance. Simian virus 40 enhancer sequences placed adjacent (3' side) to the 65-bp dyad also abolished the ability of the EBV plasmids to be maintained in an extrachromosomal state. These studies identify the regions of the 65-bp dyad that are essential for extrachromosomal maintenance and point out a functional asymmetry which exists in the structurally
symmetrical element.
Epstein-Barr
virus(EBV)infection ofprimary Blympho-cytes results in the "immortalization" of these cells sothat
they can replicate and grow in cell culture indefinitely (17; reviewed in reference 10). In most cases infectious virus is not produced bythesecells and only asmall fractionof the viral genome is expressed in the form ofstable cytoplasmic
EBV-encoded mRNA(7, 26, 35). The 180-kilobase (kb) viral genomepersists as acircular,extrachromosomal,multicopy
plasmid inlatentlyinfected cells (13, 24). Such autonomous
plasmids
replicate only once during the S phase of the cellcycle
inacoordinate fashion with cellularDNAsynthesis (1,14). Latentlyinfected cells commonly express several EBV-encodedproteinslocalizedinthe nucleus (6, 9, 16, 19-21, 23,
31, 33) or the cell membrane (18). Of these proteins, EBV nuclear antigen 1 (EBNA-1) appears to be required for
replication
of the EBV plasmid (25, 37).Previous studies have identified a region of the EBV genome, oriP, as the putative viral replication origin
em-*Correspondingauthor.
ployed during latent infection (25, 36). Recombinant DNA plasmids containing oriP and a dominant selectable marker gene, such as G418 resistance or hygromycin resistance, wereshown to be maintained in an extrachromosomal form in cells expressing EBV proteins (25, 36, 37). Incorporation of the gene for EBNA-1 into such plasmids permitted episomal maintenance in cells not expressing other EBV-encoded proteins (25, 37), and mutations in the gene for EBNA-1 prevented extrachromosomal persistence of the plasmid (25). Deletion mapping localized two separable elements in oriP that were each required in cisfor extrachro-mosomal maintenance (25, 30). The first element consistsof 20copies of a tandemly repeated 30-base-pair (bp) sequence. Thesecond essential element, located about 1 kb away inthe viral chromosome, consists of a 65-bp sequence forming a dyad sequence symmetry. The dyad region harbors four related copies of the 30-bp repeat, with two of these copies embedded in the dyad structure itself (2). EBNA-1can bind to a12-bp palindromic core sequence which is found in each of the 30-bprepeats, as well as in the four related sequences
3016
on November 10, 2019 by guest
http://jvi.asm.org/
located in the dyad symmetry region (27), suggesting that EBNA-1 acts at these sites directly to facilitate plasmid maintenance. In some cases a segment of the EBV BamHI C fragment, including oriP, can functionally substitute forthe simian virus 40 (SV40) enhancer-promoter regulating a drug resistance gene (25), and the 30-bp repeat element can act as an EBNA-1-dependent enhancer when positioned adjacent to heterologous promoters (29). Other viruses, such as polyomavirus, have a similar organization at their origin of replication (34). A trans-acting viral protein,thelargetumor antigen, like EBNA-1, acts directly upon a replication origin
ofdyad symmetry to initiate DNA replication. Polyomavirus
replication also requires the presence of an enhancer ele-ment (in analogy to the 30-bp repeats) upstream from the dyad (8).
To study the sequence requirements within oriP, recom-binant plasmids were constructed containing deletion muta-tions in the 30-bp repeated sequences and in the 65-bp dyad
region. Between 6 and 12 of the 30-bp repeats appear to be required for extrachromosomal maintenance of these plas-mids in D98-Raji cells. Test plasplas-mids containing six or fewer
30-bprepeats weredefective for plasmid maintenance since theypersisted extrachromosomallyinrearranged form only. Isolated, rearranged plasmids were found to be revertant in thatthey had regained a higher number of 30-bp repeats and were fully wild type in maintaining the extrachromosomal copynumber. The structure of revertant plasmidsindicated
that the 30-bp repeats do not have to be organized in a tandemfashion in order to facilitate plasmid maintenance. The 65-bp dyad region alone was not sufficient forplasmid maintenance. Deletions extending into the 65-bp dyad ele-ment showed a significant asymmetry in this region.
Dele-tions removing the 5' side of the dyad did not abolish extrachromosomal maintenance of the plasmid DNA in
D98-Raji cells. In most cases the plasmids persisted after
undergoing structural alterations that may have compen-satedforthedeletions. In contrast,deletions truncating the 3' side of the dyad completely abolished plasmid mainte-nance, and rearranged episomal DNA was not observed.
Deletions that removed the EBNA-1 sites adjacent to the dyad structure alsodestroyedthecapacityfor
extrachromo-somal maintenance. The placement of heterologous se-quences immediately adjacent to the 3' side of the 65-bp dyad interfered with oriP function. The interference was alleviated bychangingthenatureoftheadjacentsequences.
A 7339
A:
aofm
.-9518
0 0
O-a :p
-AO
p20
EI}
p15
Ic
--pl2
-0p6
ED
p2
[B
-0 u0
04
0
x
0
X
pHm pSLneo
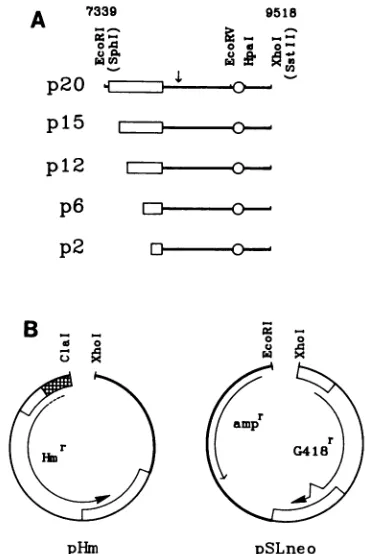
FIG. 1. Structure ofplasmids containing deletions in the 30-bp repeatdomain.(A) Mapofthe EBVplasmid maintenance region. nt numbering is according to Baer et al. (2). Symbols:
LII,
region containingthe 20copies ofa 30-bp repeat, which is progressively truncated inthe mutants to leave15,12,6, or 2intact30-bprepeats;0, region of 65-bp dyad symmetry;
4,
position ofan SstI linker insertion. (B) Structures ofplasmid vectors. pHm symbols: thick line, pML2sequencescontaining theColElreplication origin; ,transposon Tn5sequencesdirecting theexpression ofthe hygromy-cin B phosphotransferasegene in E. coli;
LII,
sequencesfromthe herpes simplex virus type 1thymidine kinasegene controllingthe expression ofhygromycin resistance in mammalian cells. pSLneo symbols: thick line, pML2 sequencesencoding ,-lactamase (amp) andcontaining theColEl replication origin;LII,
SV40 sequences controllingtheexpression oftheG418 resistancegene. Thedeletion mutations were cloned intheorientation showninto pHm(namedp20hm, pl5hm, pl2hm, p6hm, and p2hm) and pSLneo (named p20neo,pl5neo, pl2neo, p6neo,andp2neo).
MATERIALS ANDMETHODS
Cell culture.D98-Rajicellsare anadherent somatichybrid cell linecontaining multiplelatent EBVepisomes (11). Cells
were propagated in Dulbecco modified Eagle medium
con-taining 10% fetal bovine serum, 100 U ofpenicillin per ml, and100 ,ugofstreptomycinper ml.
Plasmid maintenance assay. D98-Raji cells were
trans-fected with 20 ,ug ofplasmid DNA (no carrier DNA) by a
modified calcium phosphate DNA precipitation procedure,
as described previously (25). Drug-resistant colonies arose
after 2to 3 weeks of selection in mediumcontaining either G418 (500 ,ug/ml) or hygromycin B (140 ,ug/ml). The cell culture medium was changed every 4 days. Numbers of drug-resistant colonies were estimated after staining plates with Giemsa. Colonieson remainingplates were pooled by
trypsinization and expanded in selective medium, and low-molecular-weight DNAwasprepared fromcell cultures by the method of Hirt (22). In some cases extrachromosomal plasmid DNA was recovered from Hirt preparations by
transforming competent Escherichia coli DH5
(Bethesda
ResearchLaboratories, Inc.) with thelow-molecular-weight
DNA extracted from 107 cells (15). Portions of the Hirt
extracts were digestedwith arestriction enzyme.
Digested
and undigested DNA preparations were fractionated on
0.9%
agarose gels and transferred to nitrocellulose as de-scribed previously (25, 32). Radioactive probes werepre-paredby nick translation ofvectorplasmid DNA,
resulting
in specific activities of about 108 dpm/,ug.
Hybridization,
washing of filters,andautoradiographywere
performed
also asdetailedpreviously (25).Plasmidconstructions. TheBamHI-CEBV DNAfrom the
SphI
siteatnucleotide(nt)7339 (ntnumbering
as inBaeretal.
[2])
to the SstII site at nt 9518(Fig. 1A)
contains thecis-acting genetic elements necessary for
plasmid
mainte-nance: the 30-bp repeat domain(LII)
and the65-bp
dyad
region
(0).
TheSphI-to-SstII
EBVfragmentwascloned into the ClaI-to-XhoI sites of thehygromycin
resistance-basedvectorpHm after the addition ofClaIand XhoIlinkers
(Fig.
1A). The 3.1-kb pHm plasmid containsthe gene for
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.344.527.77.354.2]mycin resistance (12), selectable both in E. coli (TnS
pro-moter) and inmammalian cells (herpes simplex virus type 1 thymidine kinase expression signals). A single SstIl linker
wasintroduced at the SmaI site (nt8193) in the EBV DNA (Fig. 1A,
I
) to facilitate subsequent manipulations. The resulting plasmid, p20hm, represents the wild-type plasmid used in these studies containing thefullcomplement of 30-bprepeatsandanunaltered dyadsymmetry element. Unidirec-tional deletions were introduced into the 30-bp repeat do-main by digestion with Bal 31 nuclease from the ClaI site in pHm. The fourdeletion mutations chosen for studyleave 15, 12,6, or2 intact copies of the 30-bprepeats, as determined
by gelelectrophoresis andDNAsequencing. These deletion mutations give rise to plasmids pl5hm, pl2hm, p6hm, and p2hm in the hygromycin vector. The ClaI-to XhoI region of EBV DNA from p2ohm and the deletion mutants was
subcloned into the EcoRI-to-XhoI sites of the G418 resis-tance vectorpSLneo (Fig. 1B), resulting inplasmids p20neo, pl5neo, pl2neo, p6neo, and p2neo. The construction of pSLneowasdescribed previously (25).
The pOneo vector, which lacks all 30-bp repeats, was
constructed by deletingthe entire 30-bprepeat domainfrom p20neo. The EBV DNA from theEcoRI site (nt 7339)tothe MluI (nt 8316) site was removed, and aBamHI linker was
inserted in its place. This results in aunique BamHI site700
bp upstreamfrom the dyad element. Multiple copies of the following synthetic double-stranded oligonucleotide,
corre-spondingtoa consensus 30-bp repeat, werecloned into the
BamHI site of pOneo:
GATCC AGATT AGGAT AGCAT ATGCT ACCCA GATAT A G TCTAA TCCTA TCGTA TACGA TGGGT CTATA TCTAG A head-to-tail orientation of tandem oligonucleotides was
ensured by digestion with BglII and BamHI after ligation, sinceonlyahead-to-tailarrangementofoligonucleotides will notrecreateaBamHI orBglII site. The number of inserted
oligonucleotides was determined by restriction fragment
sizingonhigh-percentage agarosegels. Plasmids withoneto
five tandem copies of the oligonucleotide were obtained
(plneothrough p5neo).
Deletions within the dyad symmetry region were made after first subcloning the EcoRV-to-XhoI region (for 5' deletions) or the SstI-to-HpaI region (for 3' deletions) of p20neo into Bluescript SK (Stratagene). Unidirectional de-letions from either the EcoRV or the HpaI side were
introduced using exonuclease III (Stratagene protocol) or
restrictionenzymes OxaNI, SmaI,and NdeI. Deletion
end-points were determined by the sequencing of
double-stranded plasmid DNA by a modified Sanger dideoxy
method (U.S. Biochemicals).Thedeletions chosenfor anal-ysiswere thenrecloned in p2oneo (replacingthe unaffected
dyad region of p2oneo). The resulting plasmids have struc-tures similar to that ofp20neo, except they contain either internal deletionsbetween the EcoRV site and endpointson
the 5' side of the dyad region (see Fig. 5) or deletions
extending between endpoints on the 3' side of the dyad and the XhoI site. The exonuclease-generated deletions are
namedaccordingtothe lasttwonumbers in thentposition of the deletion endpoint. For example, the 3' deletion 3.71A
removes EBV DNA between nt 9071 and the XhoI site. Similarly, the 5' deletion 5.49/removes EBV DNA between
nt9049 and the EcoRV site. Restriction enzyme-generated deletions Hpa3'A, Sma3'A, Nde3'A, andOxa5'Ahave dele-tion endpoints within the dyad region at nt positions 9136, 9108, 9062, and9008,respectively. The SstI-to-XhoI regions of 3' dyad deletion mutants were also cloned into p2Ohm
(replacing the nondeleted SstI-to-XhoI dyad region of
p20hm), forming plasmids pHpa3'Ahm, pSma3'LThm, p3.95Ahm, p3.71Ahm, and pNde3'zThm.
RESULTS
There are two EBV genetic elements, constituting oriP, that are required in cis for the extrachromosomal mainte-nanceof plasmids in latently infected cells:a30-bp sequence repeated 20 times and a65-bp sequencearranged in a dyad (25, 30). An extensive deletion analysis of these two ele-ments was undertaken in an attempt to understand the minimum sequence requirements for their function in la-tently infected cells.
Plasmid maintenance requirements for the 30-bp repeats. Therequirement for the 30-bp repeat region was first exam-ined by constructing plasmids bearing deletions which pro-gressively truncate the 30-bp repeatdomain. Bal 31 deletions were introduced into the 5' side of the repeat domain (Fig. 1),leaving 15, 12, 6, or just 2 of the original 20 repeats intact, while the dyad symmetry region was not affected in these plasmids. The deleted oriP sequences were cloned into two different vectors: pSLneo, encoding G418 resistance regu-lated by SV40 enhancer-promoter sequences, and pHm,
containingthe genefor hygromycin resistance selectable in both E. coli and animal cells. Deletion plasmids pl5hm, pl2hm, p6hm, and p2hm share the pHm vector backbone, whereas the deletion series pl5neo through p2neo contain the pSLneobackbone. Plasmids p20hm and p2Oneo contain the wild-type EBVplasmid maintenance sequences, with all 20 repeats intact, cloned into pHm and pSLneo, respec-tively. Previous studies have demonstrated that extrachro-mosomal plasmid maintenance is associated with a substan-tial (up to 200-fold) increase in the number of drug-resistant
coloniesrelative toaplasmid vector that must integrate into cellular DNA (25). Therefore, the ability of the deletion mutants topersist extrachromosomally was tested by trans-fecting the plasmids into D98-Raji cells (which supply EBNA-1 gene product in trans from the endogenous latent EBV genomes) and selecting either hygromycin- or G418-resistant colonies. Similar results were obtained with both the pHm-and thepSLneo-based test plasmids. This demon-strates that the conclusions of these experiments do not depend upon either the nature of the selectable marker employed (G418orhygromycin) or the nature of the plasmid
constructionsused.Typically, deletion mutants with 15 or 12 30-bp repeats gave rise to wild-type levels of drug-resistant colonies (Table 1). However, plasmids with just six or two copies of the 30-bp repeats were greatly impaired (10- to 40-fold) in their ability to generate drug-resistant colonies, yielding numbers comparable to a vector that must integrate into cellular DNA to persist in the cells (Table 1).
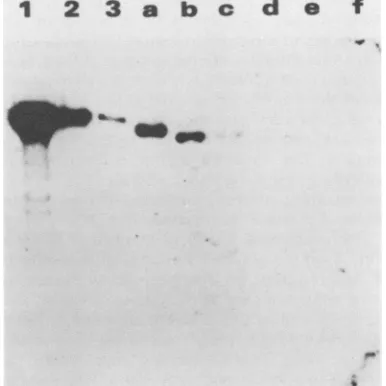
To determine whether these plasmids were maintained extrachromosomally, drug-resistant colonies were pooled and expanded in cell culture, and low-molecular-weight DNA was prepared by the method of Hirt (22). The low-molecular-weight DNA preparations were then analyzed by Southern blotting to determine whether the transfected plasmid persisted extrachromosomally in these cells. An analysis of these preparations digested withXhoI indicated that extrachromosomal DNA of the expected size was presentforp20neoand thepl5neo and pl2neo mutants (Fig. 2A, lanes a,b, and c, respectively). However, the extrachro-mosomal plasmid DNA detected in the low-molecu-lar-weight fractions of p6neo (Fig. 2A, lane d) and p2neo
transfections appeared to have undergone rearrangements,
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Transfection of mutant and revertant EBV plasmids Relative no. Estimated
Plasmid ofdrug-resistant episomal
b
D98-Rajicolonies" copyno.
pSLneo 1 0
p20neo 350 1
pl5neo 330 1
pl2neo 260 1
p6neo 35 5-10'
p2neo 10 20-50C
pHm 1 0
p20hm 340 1
pl5hm 340 1
pl2hm 280 1
p6hm 30 10-20'
p2hm 10 20-50'
p2Rlhm 550 10-20
p2R2hm 220 10-20
p2R3hm 200 10-20
p2R4hm 180 10-20
p6Rlhm 650 10-20
p6R2hm 330 10-20
a Relative number of drug-resistant colonies estimated 2 weeks after transfection,ascomparedwith the numberofcolonies produced by pSLneo
orpHm(set to1).
b Approximatecopy numberdetermined fromSouthernblots, by
compar-ing the hybridization intensity with plasmid DNAstandards (as inFig.2A). ' Indicatesthepresenceof rearranged extrachromosomal plasmidDNA.
as demonstrated by additional bands of anomalous size,
particularlyin thecaseofp2neo(Fig.2A, lane e). Appropri-ately sized form I (uncut)extrachromosomal plasmidDNA
(Fig.2B,arrow) was evidentforp20neo,pl5neo,andpl2neo
(Fig.2B, lanes a,b,and c). In contrast,formIplasmidDNA
ofthe correct size was not detectable for p6neo or p2neo, and the extrachromosomal DNA present was of much greatermolecularweightthanexpected(Fig.2B, lanes d and
e).Digestion ofthe HirtextractswithXhoI,whichcuts once in the parentplasmidstructure,reducedasubstantialportion
of the
higher-molecular-weight
extrachromosomal plasmidspecies, detected in the Southern blot ofundigested DNA
preparations, totheappropriate linearmonomersize(Fig. 2,
compare lane d in panel B and lane d in panel A). These results are consistent with the possibility that the high-molecular-weight plasmid species observedin these South-ernblots were
extrachromosomal,
multimeric forms ofthetransfectedplasmid.Thereisalwaysasmall levelofsheared
high-molecular-weight chromosomal DNA present in these Hirt extracts. This raises the possibility that the plasmid species detected in Fig. 2B represent tandem, integrated copies of thetransfected plasmidDNA. This appearstobe veryunlikelyfor tworeasons. First, Hirt DNApreparations
obtained from cell lines derived by transfection with
plas-mids thatareexpected tointegrate into cellular DNAnever
produced suchprominentbandsonSouthernblots. InaHirt
extractpreparedfrompSLneo,forexample, onlyfaint bands
ofless than one copy percell were detected
(i.e.,
seeFig.
6B,lane a). Second, many
independent
multimericorrear-rangedplasmidspecieswererescued inE. colifrom the Hirt extracts ofp6hm and p2hm transfections (see
below).
The large sizes of these rescuedplasmids
areconsistent with thehigh-molecular-weight plasmid
species
apparent in the Southern blots.One
interpretation
of thesefindings
isthat, whereas 12ormore copies of the 30-bp repeat are sufficient for
plasmid
A
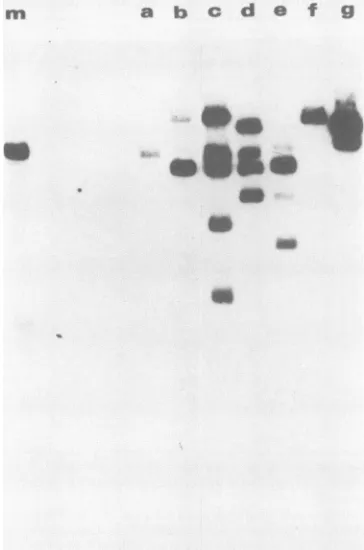
1 2 34 a b c d e_^.
a
Ba
b c d eBabo
do__
FIG. 2. Southern blot analysis of low-molecular-weight DNA extracts prepared from pooled G418-resistant D98-Raji colonies after transfection with plasmids containing deletions in the 30-bp repeatdomain. (A)Xhol-digestedHirtextracts. Lanes 1 through 4, Markers, containing 1,000, 500, 100, or 10 pg, respectively, of XhoI-digestedp2oneoDNAmixed withlow-molecular-weightDNA preparedfromD98-Rajicells(about5 x 106cellswere used forall Hirtpreparations). Low-molecular-weightDNAwasextracted from G418-resistant cells after transfection of p20neo (lane a), pl5neo
(laneb), pl2neo(lanec), p6neo(laned), andp2neo(lane e). DNA preparations were digested withXhoI, fractionatedbyagarosegel
electrophoresis, blotted to nitrocellulose, and probed with
32p_
labeledpSLneo. (B) UndigestedHirtextracts.Lanesathroughe are
thesame asinpanelA. Thearrowindicates theexpectedpositionof form Iextrachromosomalplasmid DNA.
maintenance,
plasmids
with 6 of fewer30-bp
repeats aredefectivefor
episomal
maintenance. TheextrachromosomalDNA present in rare
drug-resistant
coloniesresulting
fromtransfections of
plasmids
with six or fewer30-bp
repeatsmayrepresent
structurally
rearranged
formsofthedefectiveinput
plasmids
that haveregained
plasmid
maintenancefunction.
Rearrangements
of this typemight
have been selected for in theseexperiments
because an increasedcapacity
for extrachromosomal maintenance results in asignificantly
enhancedability
toproduce
drug-resistant
col-onies.Totestthis
possibility, rearranged
plasmids
from Hirt extracts ofp6hm
andp2hm
transfections were isolatedby
transformation ofcompetent E. coli. Ofmore than 50plas-mids rescued from
p6hm
andp2hm
Hirt extracts,none werestructurally
equivalent
tothetransfectedparentplasmids,
asdetermined
by
restriction enzymefragment
mapping.
Ingeneral,
the rescuedplasmids
wereextremely
large
(>20
kb). Large
plasmids
of this size transformE.coli
with muchlower
efficiency
than do smaller-sizeplasmids (15);
there-fore,
theplasmid
rescuefindings
supporttheconclusionthatfew,
if any, of the smaller 6-kbparentplasmids
werepresentin the Hirt extracts.
Several
rescued,
rearranged
plasmids
were chosen forfurther
analysis.
Plasmidsp2Rlhm, p2R2hm,
p2R3hm,
andp2R4hm
were isolated fromlow-molecular-weight
DNAextracted from
hygromycin-resistant D98-Raji
cells that had been transfected withp2hm;
p6Rlhm
andp6R2hm
wererescued from
p6hm
Hirtpreparations.
Themajority
of these rescuedplasmids
wereextremely
large
(>20
kb)
and their structures were notanalyzed
in detail.However,
theon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.56.295.91.309.2]Clal
XhoI
Clal
Clal
a
Clal
FIG. 3. Structure of plasmid p2Rlhm. p2Rlhmwasisolated from the low-molecular-weight DNA fraction of hygromycin-resistant D98-Raji cells resulting from transfection of p2hm(adeletionmutant
with just two copies of the 30-bp repeats). The structure is a
head-to-tail tetramer of the parent plasmid p2hm. Symbols: thick lines, EBV DNA; _, truncated 30-bprepeatdomains (two 30-bp
repeats); dyad symmetry regions: --* hygromycin resistance
genes. The indentation shows the position of a small deletion
(approximately 300 bp) that resultsin theloss ofasingleXl?olsite.
tureofone plasmid, p2Rlhm, wasdetermined (Fig. 3).This
20-kbplasmid, isolated fromap2hmHirtextract, consists of fourcopiesof the transfected plasmid (p2hm) linked headto
tail as a perfect tetramer (Fig. 3), with the exception of a
small deletion of approximately 300 bp. The p2Rlhm struc-ture,therefore, hasatotal of eight30-bprepeats(foursetsof twocopies).
The possibility that these rearrangements had restored plasmidmaintenancewasthen testeddirectly by transfecting
therescued plasmids back intoD98-Rajicells. It isclearthat the rearrangedplasmids havearestored capacitytoproduce hygromycin-resistant colonies, compared with the parent plasmids from which they were derived (Table 1). In some cases the rescued plasmids yielded more colonies than did
the wild-type construct with 20 repeats (Table 1, compare
p2Rlhm andp20hm). Equimolar levels of plasmidswerenot
employed in this experiment (20 jig of plasmid DNA was
transfected in all cases, regardless of the plasmid size).
Therefore, the numbers of drug-resistant colonies reported inTable 1 may, in fact, underestimate the degree of
resto-ration ofplasmid maintenance function tothe rescued plas-mids.
The hygromycin-resistant D98-Raji colonies arising after transfection with the isolated, rearranged plasmids were
pooled and expanded in culture. The state of the plasmid DNA in each of these cultures was analyzed by Southern blots of low-molecular-weight DNA preparations digested with XhloI. The plasmids transfected into new cell cultures were maintained extrachromosomally (Fig. 4, lanes b through g) and at even higher copy numbers than was the
wild-type plasmid, p20hm (Fig. 4, lane a). The apparent
increase incopy number is due, in part, tothe large size of theplasmids. For example,onecopyofatetrameric plasmid
percell,whenlinearized with XlzolI, willproduce the hybrid-ization signal equivalent to four copies of a monomeric
plasmid per cell, even though they may be in equimolar
quantities. Significantly, the revertant plasmids do not
un-dergoanyfurther detectablestructural rearrangements. The bands detected on these Southern blots correspond to
re-FIG. 4. Extrachromosomal maintenance of rearranged plasmids isolated from p2hm and p6hm Hirt extractswhen transfected into D98-Raji cells. Lanem. MarkerDNA, containing 500pgofp2ohm DNA digested withX/ol and mixed with thelow-molecular-weight DNA extracted from 5x 10" D98-Rajicells(thenumberofcells uLsed for all Hirt preparaitions). Low-molecular-weight DNA was
pre-pared fromhygromycin-resistant colonies arising after transfection ofp20hm (lanea). p2Rlhm (lalneb), p2R2hm (lanec).p2R3hm(lane d). p2R4hm (lane e). p6Rlhm (lane f). and p6R2hm(lane g). Hirt
extractsweredigestedwithXl?olbefore fractionationon anagarose
gel and blotting to nitrocellulose. The filterwas probed with 32P-labeledpHm DNA.
striction fragment patterns of purified plasmid DNA visible
on ethidium bromide-stained agarose gels (not shown).
These experiments demonstrate that more than 6 (but fewerthan 12) 30-bprepeatsare required for plasmid
main-tenance inD98-Raji cells. Transfection of plasmids withsix
or fewercopies gave rise to rare drug-resistant colonies in
which rearranged formsof the transfected plasmid persisted extrachromosomally. The structural rearrangements
com-plemented orsuppressedthe plasmid maintenancedefect of the parent plasmid and restored efficientextrachromosomal
maintenance.
Insertionofthe30-bprepeatsequenceintoanEBVplasmid withonly the65-bp dyadelement.The experiments shown in Fig. 1 to4demonstrate thatprobablyat leasteight copies of
the30-bp repeat elementare requiredforboth the enhanced generation of drug-resistant colonies in culture and
extra-chromosomal maintenance. Interestingly, mutant plasmids-with as few as two such 30-bp repeated elements can
undergorearrangementsleadingtoarevertantEBV plasmid thatisasefficientasthe wildtypeinitsabilitytopersist.The questionremained whetheranEBV mutant plasmidwith no
copiesof the30-bp repeatedsequence,pOneo, could undergo
arearrangementtoproducearevertantepisome. Theentire 30-bp repeat domain was removed from p20neo (Fig. 1) to
form pOneo (the 65-bp dyad region remained intact). The
m a b c d e f g
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.95.270.81.240.2] [image:5.612.350.532.84.359.2]TABLE 2. Transfection ofEBV plasmids withincreasing numbersof 30-bp repeat sequences
No.otf d-ug--esistant Estimated
Plasmid D98-Raji colonies' episomal
copy no.'
p2Oneo 1,50() 1
pSLneo 3 0
pOneo 5
plneo 25 10-20'
p2neo 17 10-20'
p3neo 10 10-20"
p4neo 14 10-20'
p5neo 9 10-20'
Number ofdrug-resistant colonies estimated2weeksaftertransfection.
Approximatecopyntumberdetermined from Southern blots.
Indicattesthe presenceof reairranged extrachromosomalpllsmid DNA.
pOneo vector failed to produce enhanced levels of G418-resistant colonies relative to pSLneo after transfection of D98-Raji cells (Table 2). Pooled colonies from this transfec-tion contained no detectable plasmid DNA in the low-molecular-weight fraction. Therefore, pOneo probably inte-grated into the host DNA and was unable to produce revertant episomal elements. One to five copies of a syn-thetic linker containing a consensus sequence of the 30-bp repeat element (25) were clonedintopOneo (plneo to p5neo). In all cases these plasmids failed to produce increasedlevels of G418-resistant colonies relative to pSLneo in D98-Raji cells(Table 2). When these colonies were expanded and the low-molecular-weight DNA fraction was analyzed, oligo-meric forms of the input plasmid DNA were found to persist extrachromosomally (Table 2). The inser-tion of as few as
7339
_.0 0 a
A: EC
-one30-bprepeat. therefore, permitted the selectionof high-molecular-weight. multimeric forms of the input plasmid and,concurrently. the selectionofthe property of extrachro-mosomal maintenance.
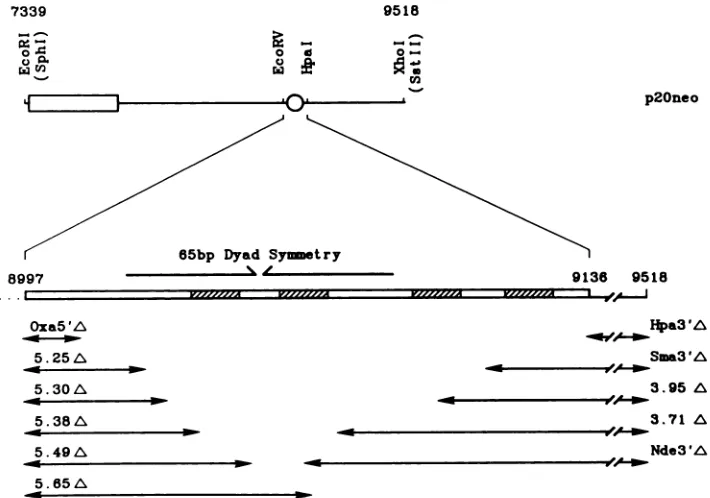
Plasmid maintenancerequirements within the dyad symme-try region. The results ofprevious studies (25. 30, 36) have identified a critical 140-bp region (Fig. 5, the 65-bp dyad) between the EcoRV and HpaI sites as necessary (in combi-nation with the 30-bp repeats) for EBV plasmid mainte-nance. To define the requirements within this region more
precisely, small deletions were introduced by limited diges-tion with exonuclease III or by use of restriction enzymes, resulting in the removal ofaDNA fragment (Fig. 5). The 5' (EcoRV side) deletions produced for this study share a common endpoint at the EcoRV site (upstream EBV se-quences were notaffected) and extend progressively intothe 65-bp dyad region (Fig. 5). Deletions OxaS'A, 5.25A, and 5.30A remove 0. 4.and9bp. respectively, ofthe 65-bpdyad structure, butdo not affect any of the four EBNA-1-binding sites (shaded regions of Fig. 5). The more extensive dele-tions 5.38A, 5.49A, and 5.65A remove 17, 28, and 44 bp, respectively, from the left side of the 65-bp dyad structure. Inaddition, 5.49A deletes the most 5' EBNA-1-bindingsite, and 5.65A deletes both EBNA-1 sitesembedded within the dyadstructure (Fig. 5). All 5' deletion mutants werecloned into a G418-based vector (Fig. 1), resulting in plasmid constructs identical to p2oneo, except containing the small deletions within the dyad region.
The effect of these mutations onthe ability of plasmidsto be maintained extrachromosomally was examined by trans-fectingtheplasmidsintoD98-Rajicells. All deletionmutants gave rise to high numbers ofG418-resistant colonies,
com-9518
p2Oneo
q5I
b dt-6 5 b~65p Dyad Sy tr
9136 9518 8997
Oxa5'A 5.25A 5.30A 5.38A
5.49A
5.65A
FIG. 5. Endpoints of deletions extending into the 5' and 3' sides of the 65-bp dyadsymmetry region. (Top) Malp of the EBV plasmid
maintenance regionpresentinthewild-type (nondeleted)plasmid p2Oneo. Batsenumbering is accordingtoBaeeretal.(2). Symbols: . 20 tandem30-bprepeats: 0.dyadregion.(Bottom)Detail of theEcoRV-to-Hpaldyadregionandendpointsofdeletion mutations.Crosshaitched boxes indicate thepositionof fourEBNA-1-bindingsiteswithin thisregion. Lineswitharrowsshow theregionsof EBV DNA that havebeen
deleted in the corresponding exonuclease- or restr-iction enzyme-generated mutations. nt positions ofdeletion endpoints are detailed in
Materials and Methods.
Hpa3'A
Sm&3'A
3.95 A
3.71 A
Nde3A
a --Vlrlrlv7m ---7,rV7,7,M I OA i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.55.294.103.211.2] [image:6.612.131.486.429.678.2]TABLE 3. Deletionanalysis of EBV plasmids
Estimated
Plasmid No. ofD98-Rajidrug-resistantcolonies" episomal
b
o.
copyno.'
pSLneo 5 0
p2Oneo 1,500 1
pOxa5'Aneo 1,400 1
p5.25Aneo 1,300 1
p5.3OAneo 1,400 5-10'
p5.38tAneo 1,100 5-10'
p5.49Aneo 1,400 5-10'
p5.65Aneo 1,000 5-10'
pHpa3'Aneo 1,300 0
pSma3'Aneo 1,100 0
p3.95Aneo 1,000 0
p3.71Aneo 1,000 0
pNde3'Aneo 1,100 0
pHm 4 0
p20hm 800 1
pHpa3'Ahm 1,000 1
pSma3'Ahm 900 <0.2
p3.95Ahm 800 0
p3.7lAhm 900 0
pNde3'hrhm 800 0
"Numberofdrug-resistant colonies estimated2weeksafter transfection.
bApproximatecopynumberdetermined from Southern blots. Indicates thepresenceofrearranged extrachromosomalplasmidDNA.
parable to a wild-type plasmid with an unaffected dyad
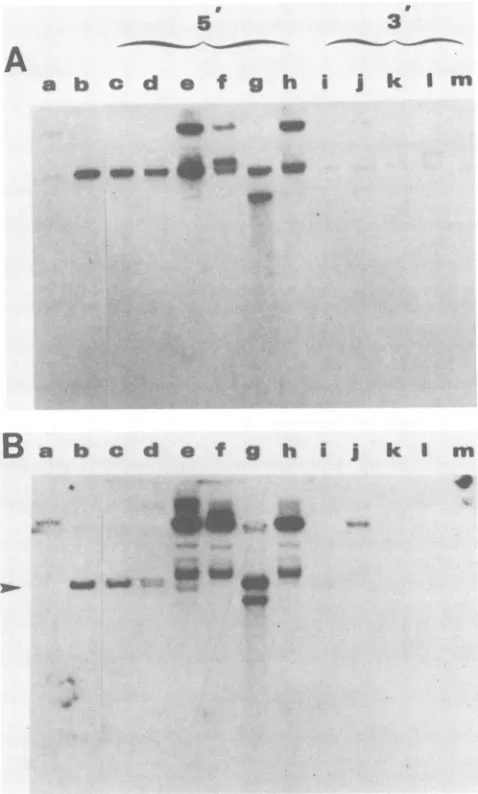
region (p2Oneo) (Table 3). This result was expected on the basis of previous findings that plasmids lacking the entire dyad region still give rise to stimulated numbers of drug-resistant colonies, despite being incapable of extrachromo-somal plasmid maintenance (25). Stimulation of the number ofdrug-resistant coloniesinthis situationmostlikely results from the EBNA-1-dependent enhanceractivity ofthe 30-bp repeats. The G418-resistant colonies arising after transfec-tion of the 5' deletransfec-tion plasmids were pooled and expanded in cell culture. Low-molecular-weight DNAwas prepared and analyzed by Southern blotting (Fig. 6). The blot of undi-gested small-molecular-weight preparations indicated that
appropriately sized form I plasmid DNA was present for
Oxa5'Aneo (Fig. 6B, lane c), which doesnotaffect the dyad structure, and p5.25Aneo, which removes 4 bp of the dyad structure(Fig. 6B, laned). Deletion ofanadditional 5 bp of the dyad structure (p5.3OAneo) resulted in the appearanceof predominantly higher-molecular-weight episomal plasmid
species (Fig. 6B, lane e). The more extensive deletions had a similar effect (Fig. 6B, lanes f, g, and h); a substantial
portion of the extrachromosomal DNA was of altered size. Extrachromosomal plasmid DNA was detected even in the case ofp5.65Aneo (Fig. 6,lane h), which deletes more than halfof the dyad structure and the two EBNA-1 sites con-tained within it. A small fraction of the signal detected in somelanes ofFig.6 may representintegrated plasmid DNA. Thefaint signal (less than one copy per cell) detected in the pSLneo vector Hirt preparation (Fig. 6, lane a) indicated some contamination of low-molecular-weight DNAs with high-molecular-weight cellular DNA. However, the
hybrid-ization signals from the 5' deletion mutants (Fig. 6, lanes e
through h) were substantially more intense than the low-level contaminants. The detailed nature of the altered plas-mid DNA structures present in the 5' deletion mutant Hirt extracts remain to becharacterized.
Asimilaranalysis was undertaken with a series of deletion
mutations thatextendinto the 3' end(HpaI side) of the dyad
5
A
a b c
d
ef
gh
i
3
j
kl
I m--
4W,.N
e-qp
Ba
b c d e f 9h
i j k I mFIG. 6. Effectonplasmidmaintenance of deletionsinthe 5'and 3' sides of the dyadregionwhentested inthepSLneovector. (A) SouthernblotanalysisofXhoI-digestedlow-molecular-weightDNA preparations. Low-molecular-weight DNAwas extractedfrom 5 x
106 G418-resistant D98-Raji cells after transfection with pSLneo (lane a),p2oneo(lane b), the 5'deletionmutants(lanescthroughh: pOxa5'Aneo [c], p5.25Aneo [d], p5.3OAneo [e], p5.38Aneo [fl,
p5.49Aneo [g], and p5.65Aneo [h]), and the 3' deletion mutants
(lanesithroughm:pHpa3'/neo[i], pSma3'AneoU],p3.95Aneo [k], p3.71Aneo [1], and pNde3'Aneo [m]). XhoI-digested DNA was
electrophoresedon anagarosegel, transferredtonitrocellulose, and probed with 32P-labeled pSLneo. (B) Southern blot ofundigested low-molecular-weight DNApreparations. Lanesarethesame asin panel A; however, DNA was not digested with Xhol before frac-tionationon anagarosegel.
region.
pHpa3'zAneo
was constructedbydeletingthe400bpof EBV DNA between the HpaI and XhoI sites of p20neo
(Fig. 5). More extensive deletions from the 3' end were introduced by use of restriction sites (pSma3'Aneo and
pNde3'Aneo) to drop a DNA fragment or were made by
exonuclease III treatment
(p3.95Aneo
and p3.7lAneo) (Fig. 5). These deletions didnotaffect the upstream 30-bprepeat domain.The 3'-side deletions were tested in the plasmid mainte-nance assayby transfection ofD98-Raji cells. Again, all of
the deletion mutants gave rise to stimulated numbers of
OD
,1:11,*
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.64.301.93.322.2]G418-resistant colonies, equivalent to the wild-type plasmid p20neo (Table 3). Southern blots of low-molecular-weight DNApreparations from pooled, expanded cultures indicated that allof the 3' deletion mutants were defectiveforepisomal maintenance (Fig. 6, lanes i through m). The faint bands present in lanes corresponding to the 3' deletion mutants reflect plasmid DNA integrated into cellular DNA, since they are of extremely low intensity (less than one copyper cell) and comparable hybridization signals were obtained from the pSLneo vector alone (Fig. 6A and B;compare lanes i through m with lane a).
These results raise the possibility that DNA sequences between the HpaI and SstII sites might be involved in plasmid extrachromosomal maintenance because pHpa3'Aneo was defective for episomal maintenance in this assay (Table 3 and Fig. 6). This finding is in apparent conflict with the results of Reisman et al. (30), who showed that the HpoI-to-SstII region was dispensable for plasmid maintenance ina similar assay.
The Hpa3'A deletion, by removing the EBV DNA se-quencesbetween the HpaI and SstII sites of p20neo, brings the vector DNA sequences (SV40 sequences directing the expression of G418 resistance) immediately adjacent to the EBV dyad element. The possibility remained that the close proximity of the vector sequences, in particular the SV40 origin region, was interfering with the function ofthe EBV dyad element. To examine this possibility, the 3' deletion mutations (Fig. 5) were subcloned into a heterologous vec-tor, pHm (Fig. 1), which has a different selectable marker (hygromycin resistance) and contains no SV40 DNA se-quences. After transfection ofD98-Raji cells, hygromycin-resistant colonies were pooled and expanded in culture for
low-molecular-weight DNA preparations. Again, all plas-mids yielded high numbers of drug-resistant colonies (Table 3). Southern blots of these Hirt extracts showed that the
Hpa3'A deletion in the pHm vector background did not prevent plasmid maintenance (Fig. 7, lane b), as it did in the pSLneo background. A faint signal (less than one copy per cell) was visible for pSma3'Ahm (Fig. 7, lane c), indicating thatthis plasmid was, at best, inefficiently maintained extra-chromosomally. Deletion mutation p3.95Ahm, which re-moves two EBNA-1 sites but does not affect the dyad structure, completely abolished plasmid maintenance (Fig. 7, lane d). Similarly, the more extensive 3' deletion mutants
p3.71Ahm and pNde3'Ahm werealso incapable ofepisomal maintenance (Fig. 7, lanes e and f). These results suggest that the inability of the Hpa3'A mutation to support extra-chromosomal maintenance, when tested in the pSLneo vector (pHpa3'Aneo), was due to interference by adjacent vectorsequences, possibly the SV40enhancer-origin region. ThepHm vectorlacks theseinterfering sequences, since the
Hpa3'Adeletion facilitated extrachromosomal maintenance whentested in the pHm vector(pHpa3'Ahm, Fig. 7, lane b).
DISCUSSION
Todefinethe core orminimal EBVreplication originused inlatent infection,asetofdeletion mutants was constructed to map at higher resolution the requirements for EBV
plasmid maintenance withinthe two essential oriP regions:
the 30-bp repeat domain and the region of 65-bp dyad
symmetry. The genetic analysis ofthe repeat domain pre-sentedheredemonstrates that between6and12copiesofthe 30-bp repeat are required for plasmid maintenance. These
findings are consistent with theprevious results ofReisman et al., whofound 10 to 12 30-bprepeats to be sufficient for
2
3
a
b
c
d
e
f
FIG. 7. Plasmid maintenance capacity of deletion mutations in the3'endof thedyadregionwhen testedin thepHmvector. Latnes 1to3, MarkerDNA, containing 500, 50,and 5 pg,respectively, of
X/iol-digested p20hm, mixed with the low-molecular-weight DNA extractedfrom 5 x 10' D98-Raji cells (all extracts were prepared from this number ofcells). Low-molecular-weight DNA was pre-pared from hygromycin-resistant D98-Raji cells that arose after transfection with the 3' dyad deletion mutants(lanes a through f: p20hm
[a].
pHpa3'Ahm Ib], pSma3'Ahm [c], p3.95Ahm Id],p3.71Ahm le], and pNde3'Ahm
Ifl).
DNA preparations were di-gested with XlioI, fractionated on an agarosegel, blotted tonitro-cellulose, andprobed with 32 P-labeledpHm DNA.
plasmid maintenance (30). Deletions that left six or fewer
30-bprepeatsin an EBV plasmid orplasmids reconstructed
with uptofive copiesofa30-bp repeat oligonucleotidewere defective forplasmid maintenance in that they gave rise to
greatlyreduced numbersofdrug-resistant D98-Rajicolonies and persisted only in rearranged form. Southern blot
analy-sis and plasmid rescueexperiments indicatedthat a
substan-tial
proportion
of the extrachromosomal, rearranged DNAwas in the form ofhigh-molecular-weight, multimeric
plas-mids derived from the transfected DNA. Several rescued, rearranged plasmids were subsequently shown to have re-storedplasmidmaintenance capacity to awild-type level,as determined by their ability to produce
greatly
stimulated numbers ofdrug-resistant
colonies and to beefficiently
maintained extrachromosomally without further rearrange-ments. These
findings
indicate that, at a low frequency, plasmid rearrangements arise the restore plasmid mainte-nance capacity to transfectedepisomes
originally defective forextrachromosomal maintenance.Atpresent it is not known whenor howthese
rearranged
plasmid structures are
generated during
theplasmid
mainte-nance assay. It is possible that they aregenerated
predom-inantlyby recombinationduring
transfection of theplasmid
DNAintoD98-Rajicells. There is precedent for recombina-tion and rearrangement of transfected DNA
during
the calciumphosphate
transfectionprocedure
(4, 28).Presum-ably, similarrearrangementsoccur
during
thetransfection ofall
plasmids,
including
thewild-type
plasmid,
but theseevents are not detected because the parent
plasmid
iscapableofefficient
plasmid
maintenance inanunrearranged
form.
Rearranged
extrachromosomal DNAwas notdetected in a Hirt extract derived from cells transfected with aplasmid containing
adyad
element butlacking
all of theon November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.336.529.84.277.2]30-bp repeats (Table 2). Thus, the presence ofatleastoneor two 30-bp repeats appears to be required in order for rearrangementsof this type to restore plasmid maintenance (Table 2). Alternatively, the presence ofat least one or two repeatsis required tofacilitatethe rearrangement or recom-bination of the transfected plasmidDNA.
It is not clear what structural features of the
rearranged
plasmids are responsible for the restoration of plasmid maintenance. The structure of one rescued
plasmid
with restoredepisomal maintenance,p2Rlhm(Fig. 3),wasfound to be a tetramer of the transfected plasmid p2hm. The restoration ofplasmid maintenance in p2Rlhm may bedue to the regeneration of a critical number of 30-bp repeats linked in cis on the plasmid (four sets oftwo repeats,fora total ofeightrepeats). The30-bprepeats may,therefore,
not need to be contiguous in the plasmid tofunction, sincetheyare widely spaced in the p2Rlhm structure. p2Rlhm also
contains four dyad symmetry
regions,
and it remainspossi-ble that the core EBNA-1-binding sites within the dyad
elements act to partially substitutefora
deficiency
of30-bp
repeats. Inany casethe EBVplasmidmaintenance elements can function with considerable versatility with respect to their relative organization. Reisman et al. showed that the
spacing and orientation of the 30-bp repeats relative tothe dyad element were not critical for their function (30). The revertantplasmids isolated in this studyattest tothe remark-ableflexibilitywithregardtothe typesofstructuresthatcan
constitute afunctional
plasmid
maintenance system.Deletion mutationswithin the
dyad
symmetryregion
were not associated with substantial reductions indrug-resistant
colony number, even in the absence of
episomal
mainte-nance. The sequences between the EcoRV site and the left side of the65-bp dyad structure aredispensable
forplasmid
maintenance inD98-Rajicells. A mutation thatremoves4bp
of the 5' end ofthe65-bp dyad structurealso
permits
normalplasmidmaintenance. Deletion ofanadditional 5
bp (total,
9bp) of the left side of the dyad structure, or even more extensive 5' deletions, resulted in the appearance of
episo-mal DNA of altered size relative to the transfected test
plasmid. The rearranged DNA detected in transfections of the 5' dyad deletions differs from the rearranged DNA presentin the 30-bprepeat deletion mutants in at least two respects. First,asubstantial fraction ofthe
episomal
5'dyad deletion mutant plasmid DNA is not of extremely highmolecularweight (Fig. 6). Second, the appearance of rear-ranged 5' dyad deletion plasmid species is not associated with a large drop in the numberof drug-resistant D98-Raji
colonies.Itisdifficult, therefore,tointerpretthe presenceof rearranged DNA in the 5' dyad deletion plasmid Hirt ex-tracts as an infrequent, spontaneous rearrangement that restores plasmid maintenance capacity (as appears to hap-penin thecaseof extensive deletions within the30-bprepeat domain). A possible explanation for these results could be that the 5' dyad deletions do, in fact, destroy plasmid maintenance and give rise to integrated forms of DNA. Episomal, oligomeric plasmidDNA species could be gener-ated by recombination between multiple, tandemly inte-grated plasmids. Alternatively, small-molecular-weight het-erogeneousDNAfragments could result from replication out of the chromosome in an onionskin fashion (3). Another
explanation forthese results may be that the moreextensive 5'dyaddeletionsdestroy plasmidmaintenance,yetgiverise to numeroustransientlyresistantD98-Raji colonies. Indeed, Reisman et al. (30) have shown that some plasmids with altered numbers of the 30-bp repeats, introduced into
D98-Raji cells, yield abortively resistant colonies that score
positive
in a2-weekdrug-resistant
assay, but not a4-weekassay (see Tables 1 to
3,
which were scored at 2weeks).
During
thesubsequent
process ofpooling
the colonies andexpanding
theculture forHirtpreparations,
thetransiently
resistantcolonies
might
then die andrarecolonies,
contain-ing
rearranged plasmid
DNA with restoredplasmid
mainte-nance
activity,
would dominate the culture. In eithercase itis
particularly
surprising
thatextrachromosomalDNA,
evenif
rearranged,
is present forpS.65Aneo,
since thisplasmid
has lost more than halfof the
dyad
structure and the two EBNA-1 siteswithin it.Previous deletion
mapping
localizedacriticalrequirement
forplasmid
maintenance within thedyad region,
but it remainedpossible
that theepisomal
maintenancerequire-mentsin this
region
wereduesolely
to thecontributionsof the fourEBNA-1-binding
sites within it. Deletion mutant5.30A
provides
evidence that thedyad
structureitselfplays
a role in
plasmid
maintenance. The 5.30Amutation,
which removes 9bp
ofthe 5' endofthe65-bp
dyad
symmetrybutdoes not
directly
affect any of the fourEBNA-1-binding
sites,
results in thepersistence
ofextrachromosomal DNApredominantly
of altered size.The initial attemptstodefine the 3' limitforEBV
plasmid
maintenance werecomplicated by negative
interferencefrom the
pSLneo
vector sequences. There are several rea-sonstosuspect that theinterference ofEBVorigin
functionmaybe duetoSV40
enhancer-origin
sequences.Interferenceby
SV40enhancer sequences onDNAreplication
has beenreported
previously
(5).
Deletion of the 400bp
ofEBVDNAbetween the
HpaI
and SstIl sitesdestroys
plasmid
mainte-nance when tested in the
pSLneo
vector, but not in thehygromycin
vectorpHm.
This deletionplaces
theSV40
origin region
immediately adjacent
to the65-bp
dyad
in theSLneo vector.
Furthermore,
anegative
effect of theSV40
origin
upon EBV-derivedplasmid
copy numberwasdemon-strated in a
previous study
(25).
Aplasmid
containing
the BamHI-C elements in thepSLneo
backbonep404
was main-tainedat 1to 10copies
percell. Aplasmid
identicaltop404,
exceptharboring
a deletion of the SV40origin region
(p404A),
was maintained atreproducibly higher
copynum-bers,
20 to 40copies
percell(25).
Thesefindings
mayhaverelevancefor
previous
studies that haveattempted
tofunc-tionally
substitute theplasmid
maintenancerequirement
forthe
30-bp
repeat domain with the SV40enhancer-origin
region (29).
From the
analysis
of thehygromycin-based
3' deletion mutants, the 3' end of thedyad
region
has been showntobequite
sensitive to mutation. Deletionsextending
past theSmaIsite
completely
eliminatedepisomal
maintenanceand,
in contrast to the 5'
deletions,
did not result in thepersis-tenceof extrachromosomal DNAofaltered size. Theresults
reported
here confirm that EBV sequences 3' to theHpaI
site are
dispensable
forplasmid
maintenance inD98-Raji
cells
(30).
Deletion to the SmaI site resulted in a loss ofplasmid
maintenance or, atbest,
very inefficientplasmid
maintenance
(less
than one copy percell).
The failure ofp3.lAhm
to be maintainedextrachromosomally
demon-strates that the
65-bp dyad
structure alone is not sufficientfor
plasmid
maintenanceand,
furthermore,
suggests acriti-cal role for the
EBNA-1-binding
site sequences adjacent tothe
dyad
structure. This is reminiscent of the SV40origin,
where sequences
adjacent
to, butnotwithin,
theSV40dyad
structureare animportant
partof thecorereplication
origin
(34).
on November 10, 2019 by guest
http://jvi.asm.org/
LITERATURE CITED
1. Adams, A. 1987. Replication oflatent Epstein-Barr virus ge-nomesin Rajicells. J. Virol. 61:1743-1746.
2. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P.J. Farrell,T.J. Gibson,G.Hatful,G. S. Hudson,S. D.Satchwell, C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA se-quence andexpression ofthe B95-8Epstein-Barrvirus genome. Nature(London)310:207-211.
3. Botchan, M., W. C. Topp, andJ. Sambrook. 1979. Studieson
SV40 excision fromcellularchromosomes. ColdSpringHarbor Symp. Quant. Biol. 43:709-719.
4. Calos, M. P., J. S. Lebkowski,and M. R. Botchan. 1983. High mutation frequencyin DNA transfected into mammalian cells. Proc. Natl. Acad.Sci. USA 80:3015-3019.
5. Campbell,B.C.,and L.P.Villareal. 1986. Lymphoidandother tissue-specific phenotypes ofpolyomavirus enhancer
recombi-nants: positiveandnegative combinationaleffectsonenhancer specificityandactivity. Mol. Cell. Biol. 6:2068-2079.
6. Dambaugh,T.,K.Hennessy,L.Chamnankit,and E. Kieff.1984. The U2regionofEBV DNA may encode EBNA2. Proc. Natl. Acad. Sci. USA81:7632-7636.
7. Dambaugh, T.,K. Hennessy,S. Fennewald,and E. Kieff. 1986. The genomeand itsexpressionin latentinfection,p. 105-140.In M. Epsteinand B.Achong(ed.),TheEpstein-Barrvirus:recent advances. Heineman, London.
8. de Villiers, J., W. Schaffner, C. Tyndall, S. Lupton, and R. Kamen. 1984. Polyoma virus DNA replication requires an enhancer. Nature (London)312:242-246.
9. Dillner, J.,B.Kallin,H.Alexander,I.Ernberg,M.Uno,Y.Ono, G. Klein, and R. Lerner. 1986. An Epstein-Barr virus deter-minednuclearantigen(EBNA5) partlyencodedbythe transfor-mation associatedBam W-Y-H regionofEBV DNA: preferen-tial expression inlymphoblastoid cell lines. Proc. Natl. Acad. Sci. USA83:6641-6645.
10. Epstein,M.A.,and B.G.Achong (ed.). 1979.TheEpstein-Barr virus. Springer-Verlag, Berlin.
11. Glaser, R., and M. Nonoyama. 1974. Host cell regulation of inductionofEpstein-Barrvirus.J. Virol. 14:174-176.
12. Gritz,L., andJ. Davies. 1983. Plasmid-encoded hygromycin B
resistance: the sequence of hygromycin B phosphotransferase gene andexpression inE. coliand Saccharnoiycescerelisiae. Gene 25:179-188.
13. Gussander, E., and A. Adams. 1984. Electron microscopic evidence forreplicationof circularEpstein-Barrvirusgenomes inlatently infectedRajicells. J. Virol. 52:549-556.
14. Hampar, B., A. Tanaka, M. Nonoyama, and J. Derge. 1974.
Replication of the resident repressed Epstein-Barr virus
ge-nomes during the early S phase
(S-I
period) ofnon-producer Raji cells. Proc. Natl. Acad. Sci. USA 71:631-633.15. Hanahan,D.1983.Studiesontransformation ofEscherichiacoli with plasmids. J. Mol. Biol. 166:557-580.
16. Hearing,J.C.,J.-C.Nicolas,and A.J.Levine.1984. Identifica-tion of Epstein-Barr virus sequences that encode a nuclear antigen expressedin latentlyinfectedlymphocytes. Proc. Natl. Acad. Sci. USA 81:4374-4377.
17. Henle, W., V. Diehl, G. Kohn, H. zur Hausen,and G. Henle. 1967. Herpes-type virus and chromosome marker in normal leukocytes after growth with irradiated Burkitt cells. Science 157:1064-1065.
18. Hennessy, K.,S. Fennewald,M.Hummel,T.Cole,and E.Kieff. 1984. A membrane protein encoded by Epstein-Barr virus in latent growth transforming infection. Proc. Natl. Acad. Sci. USA 81:7207-7211.
19. Hennessy, K.,S.Fennewald,andE. Kieff. 1985. Athird nuclear
protein in lymphoblasts immortalized by Epstein-Barr virus. Proc. NatI.Acad. Sci. USA 82:5944-5948.
20. Hennessy, K., M. Heller, V. Van Santen, and E. Kieff. 1983. Simple repeat array in Epstein-Barr virus DNA encodes part of the Epstein-Barr nuclear antigen. Science 220:1396-1398. 21. Hennessy, K., and E. Kieff. 1985. A second nuclear protein is
encoded by Epstein-Barr virus in latent infection. Science 227: 1238-1240.
22. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369. 23. Kallin, B., J. Dillner, I. Ernberg, B. Henrikson, A. Rosen, W.
Henle, G. Henle, and G. Klein. 1986. Four virallydetermined nuclear antigens are expressed in Epstein-Barr virus trans-formed cells. Proc.Natl. Acad. Sci. USA 83:1499-1503. 24. Lindahl, T., A. Adams, G. Bjursell, G. W. Bornkamm, C.
Kascha-Dierich, and U. Jehn. 1976. Covalently closed circular duplex DNA ofEpstein-Barr virus in a human lymphoid cell line. J. Mol. Biol. 102:511-530.
25. Lupton,S., and A. J. Levine. 1985.Mapping genetic elements of Epstein-Barr virus that facilitate extrachromosomal persistence of Epstein-Barr virus-derived plasmids in human cells. Mol. Cell. Biol. 5:2533-2542.
26. Orellana, T., and E. Kieff. 1977. Epstein-Barr virus-specific RNA. 11. Analysis of polyadenylated viral RNA in restringent. abortive,and productiveinfection. J. Virol. 22:321-330. 27. Rawlins,D.R., G. Milman, S.D.Hayward, and G.S.Hayward.
1985.Sequence-specific DNA binding of the Epstein-Barr virus nuclear antigen (EBNA-1) to clustered sites in the plasmid maintenance region. Cell 42:859-868.
28. Razzaque, A.,H.Mizusawa, and M. Seidman. 1983. Rearrange-mentandmutagenesis of a shuttle vector plasmid after passage in mammalian cells. Proc. Nati. Acad. Sci. USA 80:3010-3014. 29. Reisman, D., and B. Sugden. 1986. trantls activation of an Epstein-Barr viral transcriptional enhancer by the Epstein-Barr viral nuclear antigen 1. Mol. Cell. Biol. 6:3838-3846.
30. Reisman, D., J. Yates, and B. Sugden. 1985. Aputativeorigin of replication of plasmids derived from Epstein-Barr virus is composed oftwo cis-acting components. Mol. Cell. Biol. 5: 1822-1832.
31. Ricksten, A., B.Kallin, H. Alexander, J. Dillner, R. Fahraeus, G.Klein,R.Lerner,and L.Rymo. 1988.BamHI Eregionofthe Epstein-Barrvirusgenome encodes three transformation asso-ciated nuclearproteins.Proc. Natl. Acad.Sci. USA 85:995-999. 32. Southern, E. M. 1975. Detection ofspecific sequences among DNAfragmentsseparated by gel electrophoresis. J. Mol. Biol. 98:503-517.
33. Summers,W.P.,E. A.Gragan,D.Shedd,M.Robert, C.-R. Liu, andG. Miller.1982. Stableexpressionin mousecells ofnuclear neoantigenafter transfer ofa3.4-megadalton clonedfragmentof Epstein-Barr virus DNA. Proc. Natl. Acad. Sci. USA 79: 5688-5692.
34. Tooze, J. (ed.). 1981. Molecular biology of the tumorviruses, part 2. DNA tumor viruses, 2nd ed. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
35. Van Santen, V., A. Cheung, and E. Kieff. 1981. Epstein-Barr virus (EBV) RNA. Vil. Viral cytoplasmic RNA in a restrin-gently infected cell line transformed in vitro by EBV. Proc. Natl. Acad. Sci. USA 77:5453-5457.
36. Yates,J. L., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA 81:3806-3810. 37. Yates,J. L.,N.Warren, andB.Sugden. 1985. Stablereplication
ofplasmidsderived fromEpstein-Barrvirus in various mamma-lian cells. Nature(London)313:812-815.