0022-538X/88/051667-12$02.00/0
Copyright © 1988, AmericanSocietyforMicrobiology
The
Polyomavirus Enhancer
Comprises Multiple
Functional Elements
CHRISTOPHERR. MUELLER, WILLIAM J. MULLER, ANDJOHN A. HASSELL*
DepartmentofMicrobiology andImmunology, McGill University, 3775 University Street, Montreal, Quebec,
Canada H3A 2B4
Received 6 October 1987/Accepted 28 January 1988
The polyomavirus enhancer occupies244 basepairs within noncodingsequencesbetweentheearlyandlate
transcription units. To define more precisely the DNA sequences that make up the enhancer, we cloned it together with the viral early promoter upstream of a reporter gene, isolated mutants bearing deletions introduced in vitro intheenhancer, and measured the capacity ofthevariousmutantgenomestoexpressthe cat gene after transient transfection into mouse 3T3 cells. Analysis ofa large number of deletion mutants
revealed that the enhancer is between 102 and 172 base pairslong and can be divided into at least three functional elements. Relative to the entire enhancer, individual elements possessed little or no enhancer
activity. However, pairs ofelements enhanced transcriptiontolevels much higherthan thesumofindividual elements approximating the activity of the complete enhancer. These findings support the view that the polyomavirus enhanceriscomposedofmultiplesequenceelements thatfunctioncombinatorilyandimplythat ameasureofcooperationexists inthe interaction betweencellularprotein factorsboundtotheircognatesites in the enhancerand the transcriptional machinery ofthe cell.
Enhancerswere firstdiscovered in thegenome of simian
virus 40(SV40) (3, 12, 34) and polyomavirus (Py) (8). They
wereoperationallydefinedascis-actingsequencesthat could
enhance therateoftranscriptionfrom their naturalpromoter or from foreign promoters in an orientation- and
position-independent way (2). Enhancerswere subsequently
discov-ered in the noncoding region of other viral and cellular genomes(forareview, see reference 33). Some enhancers, especially those of cellulargenes, functionmost effectively in particular differentiated cells, whereas others, generally those ofviruses,act ina widevariety ofcelltypes.
Mostenhancersoccupyarelativelylargestretch ofDNA, and no nucleotide sequence has been identified that is commontoallenhancers.Manyof them shareoneofseveral short sequences which have been termed enhancer core sequences. One ofthese,
5'-G(C)G(C)C(T)CTGGA(T)A(T)-3', was first recognized to be important for function of the SV40 enhancer and is commonly referred to as the SV40
enhancer core (22, 53). Another sequence, 5'-A(C)GGAA GTGAA(C)-3',wasinitially recognizedwithin theborders of theadenovirustype5ElAenhancer and is also conserved in other enhancers(15). Commonly,eitherone orboth ofthese sequencesarerepeatedwithin enhancers. Forexample, both core consensussequencesarerepeatedwithin theborders of
the adenovirus type 2 ElA(16) and adenovirustype5 ElA (15) enhancers, and each is present at least once in the Py enhancer(35).
Geneticanalysesof theSV40enhancerhave revealed that
no one sequence, including the enhancer core motif, is
absolutely essential for its activity and have led to the realization that theSV40 enhancer is madeupoffunctionally
redundant sequence elements. Mutation ofone ofat least threeregions impairs but doesnotabolishenhanceractivity (53, 56). Similarly, revertants of SV40 mutants bearing deletions (47, 51) or point mutations (18, 19) within the enhancer contain duplications of one of the same three
elements identified by mutational analyses. Analysesof the
* Correspondingauthor.
Rous sarcoma virus enhancer (23), the bovine papilloma-virus enhancer (52), and the Py enhancer (17, 35, 49) have
led to similarconclusions.
Although the mechanism whereby enhancers function is notknown,it isbecomingclear thattheyactinconcertwith cellularfactors.This was demonstratedbyin vivo competi-tionexperimentsaftertransfection ofmonkeycells withtwo
recombinant plasmids, one containing only the SV40
en-hancer and the other carrying the SV40 enhancer and
promoter coupled to a reporter gene. Cotransfection of
increasing quantitiesof theenhancer-bearing plasmidled to decreased expression of the reporter gene (44). This
obser-vationisconsistent with thenotion that cells contain limiting
amounts of positively acting factors. Subsequently, direct physicalinteraction of nuclearproteinfactors with
enhanc-ers was demonstrated in vivo and in vitro (reviewed in reference 30). In SV40, the nuclear factors interactin vitro with thesame threeregions that were shown by mutational analysis tobe required forenhanceractivity (7, 55).
ThePyenhancerwasamongthefirsttobediscovered(8), butdespiteagrowing bodyof dataonthebindingofproteins to it (5, 13, 21, 38-41), relatively little is known about its precise locationandorganization.A244-base-pair (bp)DNA fragment, located within the viral noncoding region, has beenshowntopossessthecharacteristics ofanenhancer(8).
Herbomel et al. (17) have divided this segment into two regions (Aand B)that displaydifferentialactivity in mouse
fibroblasts and embryonal carcinoma cells, and several groupshaveidentifiedmultiplesequenceelementswithin the enhancer that activate DNA replication and transcription (36, 48, 49). However,the bordersof the enhancer and the functional sequences specifically required for transcription enhancement have not been identified. To this end, we
subjected the Py enhancer to deletion mutagenesis and tested the capacity of the resulting mutants to enhance expression of a reporter gene coupled to the Py early promoterortotheheterologous herpes simplexvirustype 1 thymidine kinasegene(tk)promoter. Herewereportonthe finestructureof thePyenhancerasrevealedbythisanalysis.
1667
on November 10, 2019 by guest
http://jvi.asm.org/
1668 MUELLER ET AL.
The enhancer is made up of at least three sequenceelements which we have numbered 1, 2, and 3. Individual elements function poorly or not at all to augment transcription, but pairwise combinations of elements enhance transcription nearly as well as all three elements together do. The three elements share few sequences, suggesting that multiple cellular factors interact with the Py enhancer. The latter notion is supported by recent findings that at least four different cellular proteinsbind to theenhancer invitro (5, 13, 21, 38-41).
MATERIALS AND METHODS
Cell culture andtransfections. Mouse3T3 cells were grown in Dulbecco modified Eagle medium containing 10% calf serum at 37°C in a humidified 7% CO2 atmosphere. To controlfor transfectionefficiencies, a large stock of 3T3 cells was kept frozen and portions were thawed before each experiment. Transfections were performed as described by Wigler et al. (54), withthe following modifications. The cells were trypsinized on the day before transfection and seeded in fresh medium at adensity of 1 x 105 to 3 x 105 cells per 100-mm (diameter) dish, and the medium was not changed on the day oftransfection. Duplicate plates of cells were generallytransfected with 10 pgofplasmid DNA per plate. After incubation overnight, the cells were washed with
phosphate-buffered saline containing 2 mMethylene
glycol-bis(P-aminoethyl
ether)-N,N,N',N'-tetraacetic acid,fol-lowed by phosphate-buffered saline alone, and then the medium was replenished. Cell lysates were prepared 48 h afteraddition of the DNA.
CAT assays. Chloramphenicol acetyl transferase (CAT)
assays were carried outas described by Gorman et al. (14), except that the cellswere lysed by three cycles of freezing
and thawing. Typically, lysates contained the contents of approximately2 x 106 cells. Thereaction mixture contained
0.2 ,Ci ofdichloroacetyl-1,2-[14C]chloramphenicol, 10 mM acetyl coenzyme A, 250 mM Tris hydrochloride (pH 7.5), and S to 50 ,lI of cell lysate in a final volume of 180 ,ul.
Reactions were incubated at 37°C for 1 h. The reaction mixture was extracted with cold ethyl acetate, and the
organic phase was removed and dried. The residue was suspended in 10
RId
of ethyl acetate and spotted on a thin-layer chromatography plate. The plate waschromato-graphed vertically with chloroform-ethanol (95:5). It was then autoradiographed, and the areas corresponding to the acetylated and nonacetylated forms were cut out and counted byusing liquid scintillation fluid. CATassays were performedfor eachlysate at three concentrations of protein
under conditions of substrate excess. This ensured the linearity ofthe assay and allowed accuratecomparison ofthe activity of the extracts. CAT activity was calculated as percent acetyled
['4C]chloramphenicol
per microgram oflysateprotein ina 1-h reaction. Generally,a20-,ul sample of
extract prepared from cells transfected with the positive
control plasmid (pdPyEcat) converted 30% ofthe chloram-phenicol to the acetylated form after a 1-h reaction. This corresponded to about 105 cpm at the 100% value for the positive control plasmid, pdPyEcat. The various mutant DNAs were tested in duplicate on three separate occasions by using atleasttwo independent DNA preparations. Criti-cal mutants, whose deletions defined boundaries, were tested five times with three different DNA preparations. To permitcomparisonsbetween experiments, the wild-type (wt) parentalplasmid, pdPyEcat, and an enhancerless construct,
pdPyB9Ocat, were always transfected in parallel with the
mutant DNAs to serve as controls.
Recombinant DNA methods. The manipulations used to construct, isolate, and sequence the various recombinant
plasmidsweredescribedby Maniatisetal. (29).Weused the
nucleotide numbering scheme for Pyproposed by Soedaet
al. (46) and specifiedthe coordinates of DNA
fragments
in the clockwisedirectiononthePyphysicalmap.Theparental plasmid,pdPyEcat (seeFig. 1),wasconstructed intwostepsas follows. Plasmid pdPin7, which contains the BamHI
(nucleotide[nt] 4632)-to-EcoRI (nt 1560)
fragment
ofPyand aHindIIl linkerinplace ofsequencesbetweennt87 and nt 90, thelocation ofaformerBgiI site (36), wascleavedwithHphI, the ends of the
fragments
were rendered blunt withtheKlenowfragment of Escherichia coliDNA
polymerase I,
and the DNA was then cleaved with BamHI. The DNA
fragment
spanning
theBamHI site(nt4632)
totheHphI
site(nt 152) was
isolated,
and this DNA wasligated
topML-2
DNA (27) that had been first cleaved with
Hindlll,
bluntended,and thencleavedwith BamlIl.The
resulting plasmid,
pdPBHp, contains thePysequencesfromnt4632to nt
152,
with aHindIII linkerbetween nt87andnt90, between the
HindIll and BamHI sites of pML-2. The cat gene and
accompanying splicing
and3'processing signals
wereclonedat the ClaI site ofpdPBHp by
cleaving pSV2cat (14)
withHindIII and BamHI,
blunting
theends,
andadding
ClaIlinkers. A recombinant molecule
containing
thePy
early
promoter and cat gene in the proper orientation was
ob-tained. Because addition of ClaI linkers to bluntedBamHI
sitesregeneratestheBamHI
site,
itwasnecessaryto remove thesiteatthe end of thehybrid
transcription
unittoallowtheothersite at the
beginning
to be usedfor5' deletion muta-genesis. To thisend, permuted
linear molecules werepre-pared by partial
digestion
oftherecombinant withBamHI,
the endswerebackfilledwith the Klenow
fragment
ofDNApolymerase I, and the blunt-endedlinearDNAswere
circu-larized with T4
ligase.
Molecularcloning
of this DNAyielded pdPyEcat (see
Fig.
1). The useful features ofthisconstruct include
unique
Sall, BamHI,
HindIII,
andCIaI
sites that bracket orlie withinthePy sequences.
The 5' unidirectional deletionmutants were
generated
by
cleaving
pdPyEcat
withBamHI,
digesting
itwithBAL31forvarious periods, and
adding
BamHI orXhoI linkers to theends
(35).
The DNAwas thencleaved with HindIIIandan excess offamHI
orXhoI,
and thePy
enhancerfragments
were cloned between the BamHI and
HindIIl
sites ofpdPyEcatfor DNAs with BamHI linkersorbetween theSalI
andHindIll sites of
pdPyEcat
for DNAs with XhoI linkers.(Cleavage
of DNA with SalI and XhoIyielded
mutually
cohesive
ends.)
ConstructswithBamHI linkers containaBin theirnames(i.e.,
pdPyB5039cat;
seeFig. 2),
whereas the two deletion mutants with XhoI linkers carry an X in their names (i.e., pdPyX5097cat; seeFig.
2). Thelatter
wereoriginally
isolated for anotherreasonandwereincludedherebecause their
endpoints
are located between those definedbyBamHI linkers.
The 3' unidirectional deletion mutants were constructed
by cleaving pdPyEcat with
HindIII,
which cleaves near nt87,and digesting with BAL 31 nucleasefor various
periods.
HindIll
linkers were then added totheends,
and the DNA was cleaved withBamHI and anexcess ofHindIII. ThePy
enhancer sequences were cloned between the BamHI and HindIII sites ofpdPyEcat. Another set of 3' unidirectional deletion mutants was isolated
by
following
theprocedure
outlined
above,
butpdPyB5039cat
was used as a substrate formutagenesis,
aswellasthevectorforcloning
the deleted Py fragments.Mutants with deletions internal to the enhancer were J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
constructed by recombining appropriate DNA fragments
from5' and 3' unidirectional deletion mutants. The deletion mutants were those described previously that terminated in BamHI (5' deletion mutants) orHindlIl linkers (3' deletion mutants). In outline, the 5' deletion mutant DNAs were
cleavedwith BamHI. These DNAs served as the vector for
insertion of fragmentsbearing enhancer sequences from the 3' deletion mutants. The latter were obtained from 3'
dele-tion mutantsofthepdPyB5039catplasmid bycleavage with BamHI and HindIII. The enhancerfragmentswereligated to
the vector acrosstheir cohesive BamHI ends. These
mole-cules were then treated with the Klenowfragment ofDNA
polymerase I andcircularized byblunt-endligation.
Recom-binant plasmids carrying the appropriate orientation of en-hancer sequences were isolated. These deletion mutants
contain 16 bp of linker, replacing variable amounts of
enhancer DNA.
Plasmid pTE1was agift ofJ.Drouinand was constructed by T. Edlund (9). Py DNA fragments bearing the entire enhancer orits elements were clonedat theHindIll site of
the polylinker. This was accomplished by removing the
enhancer fragments from theappropriate construct, adding
HindIII linkers to its ends, and ligating to pTE1 that had
been cleaved with HindIII. Recombinant plasmids carrying
bothorientations ofthe enhancer were isolated and
charac-terized with various restriction endonucleases to confirm
theirstructure.
All mutant DNAs weresequenced by thechemical meth-ods of Maxam and Gilbert (31). Enzymes were used as
specified bythemanufacturer or as described by Maniatis et al. (29).
Si nuclease analysis. TotalcytoplasmicRNAfrom 10 to 20 100-mmplates (approximately4 x
106
cells) was isolated asdescribedby Favaloroetal.(10),and variousportions(10 to
100 ,ug) were hybridized to an excess ofa 5'-end-labeled,
single-strandedDNAprobe encompassingthepromoter and
transcription start sites (45). Hybridization was terminated
after 12 to 16 h, and the nucleic acids were treated with
nuclease
Si
(50). The protected DNAfragments wereelec-trophoresed through denaturing urea-acrylamide gels and
exposedto X-ray film.
RESULTS
Bordersof thePy enhancer. Tofacilitate mapping thePy
enhancer, we substitutedthe Py enhancer-promoter region
for that ofSV40 in plasmid pSV2cat (14), yielding a new
plasmid,namedpdPyEcat(Fig. 1). pdPyEcatcomprisesthe
Pyearly enhancer-promoterregion(nt 4632 to 152 or -810 to
+2), the cat gene and accompanying
SV40-p-globin
se-quencesthatprovidesplicingand3'-endprocessing signals, andpBR322sequences (nt 2066 to 23). Todefinetheborders ofthePyenhancer,weisolated a set of 5'(startingat nt4632)
and 3' (startingat nt 87) unidirectional deletion mutantsof
the enhancerregion, measuredtheircapacitytoexpress the catgeneafter transfection ofmouse3T3cells,andcompared this activity with that obtained after transfection ofthese samecells with the control plasmid pdPyEcat. Analysis of
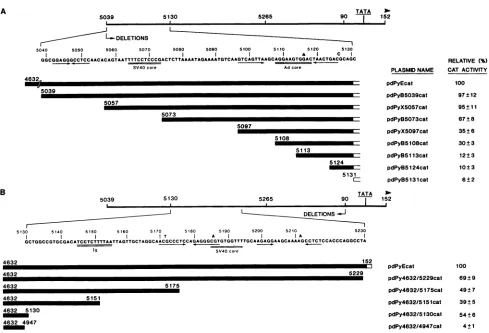
eight5'unidirectional deletion mutantsrevealed that 425bp
could be removed from the 5' region without affecting cat
expression (Fig. 2A). A small but reproducible drop in
activity occurred when sequences between nt 5057 (-385)
and nt 5073 (-369) were deleted. The mutant with an
endpoint at nt 5073 expressed the cat gene at 67% of the
control level. This deletion includes a sequence80% homol-ogous to the SV40 enhancer core. Removal of sequences
Ec oRIClaVBIarlHK
/B-GLOBI
N
cat
ENHANCER
4632 4693
BaoHIKpnl
PROMOTER
anm,",
5021 5128 5262 90 152
BclI PYUII Pvul KIndl ClaI4ind.'
POLYOM
AVIRUS
FIG. 1. Structure of pdPyEcat. pdPyEcat is composed of pML-2 sequencesspanning the regionbetweenitsBamHIandEcoRIsites (thin line), Py sequences (darkened area) betweennt 4632 and nt 152, sequences encodingthe cat gene (open box), and sequences from SV40 and rabbit ,-globin (cross-hatched area) that provide splicing and 3'-end processing signals. The line below the circle depicts Py sequences and relevant restriction endonuclease cleav-agesites. The locations ofaDNAfragment withenhanceractivity (dark box above the line)andtheearly promoter (boxedareawith vertical lines) are shown. 5' unidirectional deletion mutagenesis originated from the BamHIsite, whereas 3' unidirectional mutagen-esis started from the Hindlll site. Thearrowindicates thestartsites anddirection ofearlytranscription. The superscriptrindicates that these restriction sites, which formerly existed, were abolished during cloning and are no longer susceptible to cleavage by the enzymesatthese sites.
between nt 5073 and nt 5097 (-345) further reduced the
activity of the plasmid to 35% of the control level. This
regionisrichin A andTresidues butcontainsnoconserved
enhancersequences. Deletionof 11 bpbetween nt 5097 and nt5108,correspondingto asequencealsopresent to theright ofanAdenhancer coremotifinaninvertedorientation, had noeffect. Anadditional threefolddrop inactivityto12% of
the control occurred when sequences between nt 5108
(-334) and nt 5113 (-329) were removed. These 5 bp
correspondto the first 5ntofthe adenovirusenhancer core sequence. The mutantwith thedeletionendpointat nt5113 wasable to express the cat gene at levels onlytwofold better than mutants thatlacktheentire enhancer(12 versus5%of the control). Deletion of sequences up to nt 5131 (-311)
yielded levels ofactivity identical to that obtained with the Py early promoter devoid of its enhancer (viz., 6% of the
control).These resultssuggestedthat the 5' borderof thePy
enhancer is located between nt 5057 (-385) and nt 5073
(-369),although5'deletionmutantsthatretainsequencesto nt 5108 possessed significant activity (30% of the control).
Moreover, sequences between nt 5131 and nt 5265, which make up the B enhancer(17), were unable toincrease cat
BamL
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.317.552.74.348.2]1670 MUELLER ET AL. A
5039 5130 5265 TATA
-9,0 152
DELETIONS
5040 5050 5060 5070 5080 5090 5100 5110 5120 5130
l
l l l A CGGCGGAGGGCCTCCAACACAGTAATTTTCCTCCCGACTCTTAAAATAGAAAATGTCAAGTCAGTTAAGCAGGAAGTGGACTAACTGACGCAGC
SV40core Ad core
4632
5039
I~~~~~~~~~~~~~~~~~M=
5057
5073
5131
5130 5265 90 TATA
RELATIVE (%)
PLASMID NAME CAT ACTIVITY
pdPyEcat pdPyBS039cat
pdPyXS057cat pdPyB5073cat pdPyX5097cat pdPyB5108cat
pdPyB5113cat
pdPyB5124cat
pdPyB5131cat
152
100
97±12
95± 11
67±8 35±6
30±3 12±3 10±3 6±2
I
~~~~~~~~~DELETIONS
5130 5140 5150 5160 5170 5180 5190 5200 5210 5230
T A I|A
GCGGCCGTGCGACATCCTCTTTTAcTTAGTTGCTAGGCAACGCCCTCCAGAGGGCGTGTGoreIIIcA^GAGGAAGcAAAAAGccTcTccACCCAGGCCTA
9g SV40core
4632
pdPyEcat 5229
pdPy4632/5229cat
pdPy4632/5 1 75cat
pdPy4632/5 151cat 5151
100
69±9
49±7 39±5
4632 5 130 pdPy4632/5 130cat 54±6
4632 4947
pdPy4632/4947cat 4±1
FIG. 2. Structureandactivityof 5' and3' unidirectionaldeletionmutantDNAs.The linediagramsatthetopsofpanelsAand Brepresent the noncoding region ofPy DNA. TATA represents the TATA box, and the arrowheads depict majorearly start sites fortranscription initiation. ThecatgeneisjoinedtoPysequencesat nt152(Fig. 1). Thesequencebelow the line has thesamepolarityasearlymRNA and
is numbered asdescribed by Soedaetal.(46). The deltaabove theGat nt5116indicates that this nucleotide wasnotpresentinourstrain ofPy.Similarly,the nucleotide letters above thesequencerepresentinsertions of bases found inourstrainof the virus. The double-underlined sequencesinclude theSV40enhancercore(SV40 core),the adenovirus ElA enhancercore (Ad core),andasequencefound in themouse
immunoglobulin heavy-chain enhancer (Ig). Thearrows underline perfect inverted repeats. 5' deletionmutagenesis began at nt4632 and proceededdownstream of the BamHl site(A),whereas 3'deletion mutagenesis originatedat nt87 andproceededupstreamof theHindIlI site atthis position (B). Relative catactivity ispresented as a percentage ofthe activityof the parentplasmid pdPyEcat. The standard deviationswerecalculatedfromatleast three independenttransfectionexperiments withaminimumoftwodifferent DNA preparations of eachplasmid.
expression beyondthatobtained withaconstructcontaining only the early promoter (nt 87 to nt 152). These data are reminiscent of those obtained earlier, which revealed a
gradual decline rather than a precipitous drop in gene
expression as measured by permanent-transformation as-sayswhensequenceswereremovedfrom the 5' extremityof
the early Pyenhancer-promoter region (20, 35).
Tomapthe 3' borderoftheenhancer, webegan mutagen-esisat nt 87(-57) and extended thedeletions unidirection-ally upstream of this position (Fig. 2B). All of thedeletion mutantsretainthe earlypromotercomposed ofanupstream element (5'-CCACCC-3') and the TATA box-cap site ele-ment'(4, 35). Deletionmutantsdefined byendpoints between nt87(-57)andnt5130 (-312)werereducedin their capacity toexpress thecatgene tolevels between 39 and 69% of the control. The largest 3' deletion mutantthat retained
signifi-cant levels ofactivity (54%of thecontrol)lackedsequences up to nt 5130 (-312). This mutant lacks the entire B enhancer and a number of conserved enhancer motifs, as
wellasthe origincoreforDNAreplication. Extension ofthe
deletion beyond nt 5130 to nt 4947 (-495) reduced the
activityof theresultingmutantDNAtoexpressthecatgene to levels 25-fold lower than that of the control, which is equivalent to that obtained with a construct containing an
enhancerless early promoter. These observations suggest thatthe 3' border of the enhancermapsbetweennt5130 and
nt4947. These resultsareinagreementwith those obtained earlier, which showed that the entire B enhancer ofPy (nt 5130 to nt 5265) could be removed from the viral genome
without markedly affecting early gene expression (35) or
virus viability(28).
Unlike the results obtained with the 5' deletion series, therewas nosmooth decline inactivityof theplasmidswith increasingdeletionsize,andonemutant(pdPy4632/SlSlcat)
wasless active than another(pdPy4632/5130cat) which had
sustained alargerdeletion. Thislikely reflects the fact that the activity ofeach 3' deletion mutant is due not only to deletion of essential enhancersequencesbutalsoto
reposi-tioningof the remainingenhancersequences relativetothe downstreampromoter elements.
To definethe 3'border of the enhancermoreprecisely,we
used anotherplasmid (pdPyBS039cat), containing the entire B
5039
40;jz
----
Itluor-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.68.557.64.397.2]enhancer butdevoid of those Py sequences between nt4632
and nt 5039, as a substrate for mutagenesis (Fig. 3). The
latter sequences correspond to coding sequences for the
structural proteins ofthevirus, whose deletion did notaffect expressionof the cat gene(Fig. 2A). The results are shown
in Fig. 3. As observed previously (Fig. 2B), deletions up-streamofnt87(-57)to nt5130(-312) slightlyreduced the
capacity oftheplasmidtoexpress the cat gene (63% of the
control).However, extension of thedeletion byonly 10 bp to
nt 5120 (-322) essentially abolished the capacity of the
resultingplasmidtoexpressthe cat geneproduct (7% of the
control). Extensionofthe deletion upstream of thisposition did not further reduce this activity. These results clearly establish the 3'border ofthe enhancer between nt 5130and
nt5120and implicate those sequences between these
posi-tions, which correspondalmostexactlyto aninverted repeat
motif, as an essential component of theenhancer.
The minimal enhancer. The5' unidirectional deletions to
nt 5073 (-369) and 3' unidirectional deletions to nt 5130
(-312) reduced the capacity of the resulting mutants to express the cat gene only marginally (67 and 63% of the
control, respectively). From these results, we inferredthat
the smallest DNA stretchcapable ofactingas anenhancer
was contained within a 57-bp DNA fragment defined by
these endpoints (5073 to5130). To determine whether this
was so, we cloned this fragment in its natural orientation upstream of the early promoterin the cat vector andtested
the capacity ofthe resulting plasmid,
pdPySO73/5130cat,
to express the cat gene after transfection of3T3 mouse cells.Unexpectedly low levels of CAT activity resulted
(17%
ofthe control
plasmid;
Fig. 4). This value is only fourfoldgreater thanthatobtained after transfection of3T3cellswith
arecombinantplasmid lacking the 57-bpDNAfragment but
retaining the early promoter (pdPyB90cat).Whenthis57-bp
fragment was cloned upstream of a slightly larger DNA
segment bearingthe Py early promoter and the replication origin core
(pdPySO73APcat),
thereby displacing it fartherfrom the promoter, the putative minimal enhancer hardly
increased cat expression at all in 3T3 cells (8% of the
control;Fig. 4). These results, contrary to our
expectations,
showed that the 57-bp fragment alone did not possess
significantenhanceractivity.
This result, when considered in the context ofthe data
obtained from the5' and 3' deletionanalyses, suggested to usthattheenhancer wascomposed ofacentral, 57-bpregion
5039 5130
andone oftwoflanking segments eitherupstream or
down-stream ofthe central segment. To testthis possibility and delineate the approximate borders of the flanking segments,
weeffectively restoredsequences alternately downstreamor upstreamofthe57-bpfragmentandmeasuredthecapacity of
theresultingplasmidstoexpress the cat gene in 3T3cells.In
practice, this was achieved by deleting sequences in the
3'-to-5'
direction from a cat vector that maintained the 5'borderofthe Pyenhancer(nt5073or -369) togenerateone set of mutants and by deleting sequences in the 5'-to-3'
direction from a similar vector that maintained the
previ-ously mapped3' borderofthe enhancer (nt 5130 or -312) to
obtainanother setofmutants. Analysis ofthephenotypes of
these two groups of mutants allowed us to identify the
locations of two partially overlapping enhancers (Fig. 4).
One enhancer maps between nt 5057 (-385) and nt 5130
(-312),whereas theother islocatedbetween nt5073(-369)
andnt5229(-213).Theseenhancers haveincommon those
57 bplocated between nt 5073 (-369) andnt 5130(-312).
The only common motif shared between the DNA
se-quences upstream and downstream of the central 57-bp
fragment, which augmented its activity, is the SV40
en-hancer core sequence (Fig. 2). Additionof essentially only
this sequence (nt 5057 to 5073) to the late border of the
centralelement increasedtheactivity ofthe 57-bpfragment fourfold(Fig.4). Addition ofsequences on theearly border ofthecentralelement that includedasequencemotif found
in theenhancers ofimmunoglobulin genes (2)increasedthe
activity ofthe DNA twofold. This activity was increased
approximately another twofold by sequences between nt
5175 and nt 5229, which contain the SV40 enhancer core
motif, as well as several inverted repeats (compare Fig. 2
and4).
Noteworthy is the observation that neither minimal
en-hancer was aseffectiveastheentireregion inaugmentingcat gene expression (cf. Fig. 2 and 4). For example, the 5057-to-5130 DNA segment was about 40% as effective as the entire enhancer in augmenting cat gene expression.
Simi-larly, the DNAregion between nt5073 and nt 5229 was70%
aseffectiveasthecontrolinactivatingcatgene
expression.
Although thedifference betweenthecapacity ofany oneof
the two minimal enhancer regions and that of the entire
region to potentiate cat gene expression was small, this
difference wasreproducible.
Structure of the Py enhancer. The
mapping
data weob-5265 90 TATA
I-
1|152
DELETIONS 2
5040 5050 5060 5070 5080 5090 5100 5110 5120 5130
A C
GGCGGAGGGCCTCCAACACAGTAATTTTCCTCCCGACTCTTAAAATAGAAAATGTCAAGTCAGTTAAGCAGGAAGTGGACTAACTGACGCAGC
SV40core Ad core
4632,, 4,_152
5039 .152
5130 5120
5109 5092
PLASMID NAME i pdPyEcat
pdPyB5039cat pdPy5O39/5130cat pdPy5O39/5120cat
pdPy5O39/5109cat
pdPy5O39/5092cat
RELATIVE(%) CAT ACTIVITY
100
98±10 63±9
7±2 4±1 4±1
5087
pdPy5039/5087cat 5± 2 FIG. 3. Structure andactivity of3' unidirectional deletionmutantsofpdPyB5039cat. Thefeatures andsymbolsrepresentedarethesame asthosedefinedin thelegendtoFig. 2.
2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.557.531.704.2]1672 MUELLER ET AL.
5265 90TATA1 152>
152
5229 5175
5151 5130
90 152
1
PLASMID NAME
pdPyEcat
pdPyBS073cat
pdPy5073/5229cat
pdPy5073/5175cat
pdPy5073/5151cat
pdPy5073/5130cat
152
5265 5073 5130
5057
5039 4632
11
pdPyB9Ocat
pdPyB5265cat
pdPy5O73APcat pdPyS5O7APcat pdPy5O39APcat pdPy4632APcat
4± 1 5±2 8± 2 36± 6 39± 5 65 ± 7
FIG. 4. Structure andactivity ofthecentralenhancerelement andrecombinants carrying flanking sequences. The sequences between nt 5073 and nt5130, representing the central enhancer element, are depicted by the black box. This element and flanking downstream or upstream sequenceswerecoupled to the Py early promoterlocatedbetween nt 87and nt152 ornt5265 andnt 152.The sequencesdeleted betweenthe enhancer and the earlypromoter are between the endpoints denoted. The symbols are the same as those defined in the legend toFig. 2.
tainedwereconsistent with the model that the Py noncoding
region contained two overlapping enhancers. However, it
was formally possible that the enhancer comprised three independent elements capable of functioning in pairwise combinations.To examine this, we deleted sequences in the
region ofoverlap between the two minimal enhancers
cor-responding to the central 57-bp element and tested the
capacity oftheresultingmutants to express the cat gene in 3T3 cells (Fig. 5). The mutant bearing the largest deletion,
which spanned the region between nt 5092 (-350) and nt
5130(-312),previously shownto beessential fortheactivity of eachenhancer, expressedthe catgene at levelsequivalent
to that of either one ofthe two minimal enhancer-bearing
5039 5130
plasmids, namely, 31%of the control (Fig. 5). Significantly, the mutant which lacks sequences between nt 5120 and nt 5131 was 47% as active as the control. Removal ofthese sequencescompletelyabolished theactivity of the enhancer
composed ofsequences between nt 5039 and nt 5130(Fig. 3). Other mutants with small deletions in this region affecting
conserved sequence motifs yielded relatively high levels of CATactivity,between 61and92% ofthecontrollevels. It is
likelythat theselesions did notcompletely remove essential
enhancersequences(i.e., pdPyA5092/5108catorpdPyA5087/
5108cat) oronly partially inactivated these sequences (i.e.,
pdPyA5109/5113cat). Theseobservations suggestedthat the
enhancer comprises at least three functionally equivalent
5265 90 TATA 152152
DELETIONS,
5040 5050 5060 5070 5080 5090 5100 5110 5120 5130
A c
GGCGGAGGGCCTCCAACACAGTAATTTTCCTCCCGACTCTTAAAATAGAAAATGTCAAGTCAGTTAAGCAGGAAGTGGACTAACTGACGCAGCTGGCC
.
SV40core Ad core
LINKER
5131
5092
5109 5113 5108 5087
PLASMID NAME
pdPy5O39Bcat pdPy5O39XBcat pdPyA5120/5131cat pdPyA5092/5131cat pdPyA5109/5113cat
pdPyA5092/5108cat pdPyA5087/5108cat
FIG. 5. Structureandactivityofmutantsbearingdeletions in the central element. Deletions withinthe enhanceroccurbetween theblack
boxes withnumberedendpoints. Control plasmid pdPySO39Bcat is identical instructuretopdPy5O39XBcat,exceptthatalinker interrupts the
continuityof the DNAat nt5130. Theinternaldeletion mutantscontain the samelinker in place of the deleted DNA.Thesymbols arethe
same asthosedefined in the legendtoFig. 2.
5039 5073 5130
I l
4632,
5073
RELATIVE (%) CAT ACTIVITY
100
76±9 69 ± 8 49±9 39± 7 17±5
RELATIVE (%) CATACTIVITY
100+ 90±12
47±8
31±7
61±10
92±11
105±13
M
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.62.553.75.293.2] [image:6.612.67.553.511.684.2]elements. Apparently, any pair of elements functions nearly as well as all three elements together do to enhance cat expression in 3T3 cells. In the remaining text, we refer to these DNA segments as enhancer elements and number them 1 (nt 5057 to nt 5073), 2 (nt 5073 to nt 5130), and 3 (nt 5131 to nt 5229).
To ensure that the deletions we had made affected en-hancer rather than upstream promoter elements, which could be components of the early or late promoter andto test theproposed structure for the Py enhancer more rigorously, we used another cat vectornamed pTE1 (9). pTE1 contains a hybrid transcription unit made up of the herpes simplex virus type 1 tk gene promoter (-109 to +50) fused to the cat
geneandaccompanyingdownstream signalsfrom SV40and
rabbit 3-globin to effect proper processing of its transcript (Fig. 6). pTE1 also possesses a polylinker 750 bp upstream from the start sites of transcription in tk sequences useful for
insertionof foreign DNA. This vector allowed us to test the
capacity of the Py sequences to enhance utilization of a
heterologous promoter(tk) when cloned at adistance from
thepromoter in bothpossible orientations.
Toconfirmthat measures of cat activity reflected mRNA
levels, we compared the amount of cat mRNA and the
activity of itsproduct, CAT, aftertransfection of3T3 cells with pTE1 and its derivatives containing the Py enhancer.
We used nuclease Si analysis of RNA-DNA hybrids to
measure the abundance of correctly initiated steady-state mRNA. Figure 7 shows theresults obtained after
transfec-tion of mouse 3T3 cells with pTE1 alone or with pTE1
containingthe entire Py enhancer (nt 5039 to nt 5265) in the plus (the Py sequences facingthe tk promoter in the same
orientation as they do naturally before the Py early
pro-A
A
B
396-1+2+3 z..
x1):
1+2+3
z.
VECTOR
" X' ugRNA
344--o m
298-64X
96X
C
1x
1+2+3 1+2+3 VECTOR
Ac *
Ace
POLYOMAVIRUS
+
FIG. 6. Structure ofpTE1 and its derivatives bearing the Py enhancer. TheHindIII site is located withina56-bp polylinker. It
wasusedas asite for cloning Py DNA fragments bearing the entire enhancer, pairwise combinations of enhancer elements, or
individ-ual elements in either the plus or the minus orientation. The
blackened boxrepresentstheherpes simplex virus tkpromoter,the openbox represents thecatstructuralgene, and the hatched box
represents splicing and 3'-end processing signals from SV40 and
P-globin.
The remainder of the plasmid is composed of pBR322 sequences.CHLrnt
C L
8OX
IliX
1x
FIG. 7. Expression ofcatmRNAandenzymatic activity in 3T3 cellsafter transfection ofpTE1orpTE1 containing the Py enhancer. (A) Structure of the end-labeled DNA probe. The numbering is
relativeto thetranscriptioninitiation site (+ 1). Thearrowdepicts
the direction oftranscription and overlines thetwo principal start
sites of transcription. (B) Autoradiogram of a denaturing urea-polyacrylamide gelwhich contained theproductsofdigesting RNA-DNAhybrids withS1nuclease. The lanescontaining the products of Si analysis from 3T3 cells transfectedwithpTE1 bearing the entire Py enhancer in the plus orientation are marked by 1+2+3 (+).
Thosecontainingtheproductsobtained after transfection withpTE1 bearing the opposite orientation are marked by 1+2+3 (-). The lanes labeled VECTOR contained theproducts of Si digestion of
RNA-DNA hybrids usingRNA from cells transfected withpTE1.
The numbers onthe left indicate pBR322 DNA sizemarkers, and
thoseonthe bottom indicate the enhancement ratio. (C) Autoradio-gramofathin-layer chromatography plate containing
chloramphen-icol(CHL)and itsacetylated products (Ac).The lanesaremarked
asdescribed above. Equalamountsoflysate proteinwereassayed
for CATactivity.
moter) or the minus orientation. Figure 7A shows the structureof the EcoRI DNAfragmentusedas aprobeinthe
analysis of RNA obtained from transfected 3T3 cells. Cor-rectlyinitiated transcripts should protect 313 and 315 bases of the DNA probe. Two start sites for transcription are
locatedonthetemplate (32). Figure7B shows the results of theS1 analysis. Transfection of 3T3 cells withpTE1yielded
very low amounts ofcorrectly initiated RNA (the last two lanes in Fig. 7B).Theprotected fragmentsarenotapparent
+3'S
A
Ec
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.347.526.70.440.2] [image:7.612.60.298.405.633.2]1674 MUELLER ET AL.
here but are visible on longer exposures of the autoradio-gram. By contrast, transfection of these same cells with
pTE1bearing the Pyenhancer inthe plus (1 + 2 + 3 +) or
minus (1 + 2 + 3 -) orientation yielded much higher quantities of correctly initiated RNA. The magnitude of the enhancement of transcription was 64- and 96-fold,
respec-tively, for the plus and minus orientations of the Py
en-hancer. Assay of CAT activity in lysates from these same
cells transfected with pTE1 containing the Py enhancer in
the plus or minus orientation revealed anincreaseof 80- and
111-fold, respectively, bycomparison withthe control (Fig. 7C).These ratios correspondwell to the enhancement values obtained by analysis of cellular RNA and reinforce the
authenticity ofCATactivity measurements as indicative of steady-state levels of mRNA.
Itisnoteworthy thatcomplete deletion ofthe Py enhancer
from its early control region reduced cat expression by a
factor ofabout20-fold(Fig.2), whereasaddition of the same enhancer to the pTE1 vector increased cat expression
ap-proximately 100-fold (Fig. 7). This difference likely reflects
the overall strengths of the Py early promoter (upstream elements and TATA box) by comparison with the tk pro-moter. We have shown previously that the Py promoter is stronger than the tk promoter when assayed with the same reporter gene (11).
To confirm the proposed structure ofthe Py enhancer,
fragments of it corresponding to individual elements or
combinations ofelements were cloned in both orientations
750 bp upstream of the tk promoter in pTE1. Fourteen
different plasmids were constructed and assayed for their
capacity to express the cat gene protein product after
transfection of3T3 cells by comparison with the parental
plasmid, pTE1. The results of three independent
experi-ments are summarized in Table 1. In general, single en-hancerelements either didnotenhance catexpression atall
(element 1) or poorly enhanced cat gene
expression (i.e.,
element 3;average enhancement, threefold) by comparison
with the entire enhancer. One notable exception was ele-ment2,whichcould enhance cat geneexpression 12-foldin oneorientation(-) but only fourfoldin the other(+). Pairs
of enhancer elements potentiated catgene expression
sev-eralfoldbetter than the sumofeachelement,and this value
TABLE 1. Elevationofcatgeneexpression by Pyenhancer elements'
Enhancer Relative Enhancement PyDNA segment(nt) element(s) orientation factor"
5039-5092 1 + 1
5039-5092 1 - 1
5073-5130 2 + 4
5073-5130 2 - 12
5131-5264 3 + 3
5131-5264 3 - 3
5039-5130 1 + 2 + 7
5039-5130 1 + 2 - 83
5039-5092 and 5131-5264 1 + 3 + 31
5039-5092 and 5131-5264 1 + 3 - 26
5073-5229 2 + 3 + 43
5073-5229 2 + 3 - 61
5039-5264 1 + 2 + 3 + 83
5039-5264 1 + 2 + 3 - 101
"catgeneexpressionwasdeterminedby measuringtheactivityof the CAT enzymeincell lysates.
bThe enhancementfactor is the ratio oftheaverageactivity from cells
transfected with pTE1 carryingone or moreelements ofthe Pyenhancer dividedbythe averageactivity oflysatesfrom cells transfected withpTE1.
was nearly as high as that of all three elements together. There was a strongorientation effect ofcoupling element 2 to element 1,although such an effect was not manifested by a 2-plus-3 configuration of the enhancer. In this regard, it is noteworthy that when the SV40 enhancer trap vector was used to recover Py DNA fragments capable of restoring virus viability, only the 1-plus-2 configuration of the Py enhancer in the negative orientation relative to the SV40 early promoter was recovered (51).
To ensure that theenhanced levels of CAT activity were the result of
increased,
transcription of the cat gene, we measured by Si analysis the abundance and position of the 5' terminiof the catmRNAsthat resulted after transfection of 3T3 cells with the various plasmids in one experiment. This analysis showed that increased levels of correctlyinitiatedtranscripts resulted after inclusion of either all three orpairs ofPy enhancer elements in pTE1 (datanotshown). These results lend credence to the proposal that the Py enhancer comprises at least three enhancer elements that functionally substitute for one another in 3T3 cells. The three elements togetherfunction best, but any pair of ele-ments functions nearly as well as the entire enhancer. By contrast, individualelementsfunction poorlyornot atallto enhance catexpression in mouse fibroblasts.
DISCUSSION
The positions of the enhancer elements relative to other features in the noncoding region of the Py genome are illustrated inFig. 8. The entire enhancerregionoccupies no morethan 172bp (nt 5057 to nt 5229). It can bedividedinto
at least three apparently functionally redundant segments that we have named enhancer elements 1, 2, and 3. The borders of each element have not been rigorously
deter-mined,but theapproximate locationof each elementcanbe deduced fromtheanalyses of thevarious deletionmutants. Element 1 is likely located between nt 5057 and nt 5075,
element 2 is between nt5075 and nt 5130, andelement 3 is between nt5131 and nt5229.
The organization of the Py enhancer elements described
here is consistent with the structure of viable viruses with deleted or rearranged genomes (Fig. 8). All ofthe viable
deletion mutants isolated retain at least two enhancer ele-ments. For example, the virus dl2039L lacks sequences betweenpositions 5100 and5131 yet isviable (6). Thisvirus
does not contain enhancer element 2. Similarly, another virus
(d11024)
lacksenhancerelement 3 (28). There are also numerous strains and host range mutants of Py whose genomescontainduplicationsof enhancer element2 or3,as well as mutations within these elements (42). Duplicationsmay increase the strength of the elements, whereas
muta-tions may have the same effect or they may alter their
specificity, thereby allowingfor their interaction with pro-teins found only in embryonic cells orparticular differenti-ated cells of themouse. It is worth bearingin mind that the
organization of the enhancer and the relative strength of
each of the enhancer elements is likely to be different in different cell lines, presumably reflecting theavailabilityof
trans-acting factors in the cell (37, 43). The enhancement values for the various configurations ofthe enhancerare a measure of the activity of these factors. Therefore, the results reported here are valid formouse 3T3 cells andrat fibroblasts (data not shown) but may be different for other cell types.
Inspection of the enhancer region of viable deletion mu-tants,host range mutants, andnaturally occurringstrains of J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
POLYOMAVIRUS ENHANCER 1675
D 5100 5150 5200 5250 5292/1 20
a~~~~II a I I I I I I I|
[image:9.612.66.550.64.525.2]5100 5131
...~~~~~~...
...VIABLE VIRUSES
E.C.MUTANTS PCC4
5072
PyV STRAIN REPEATS
ENHANCER
=1=KA=KITQ
5114 5137
I soL
5057
5130
NU FA BIP
NU FA BIM
5040 5050 5060 5070 5080 5090 5100 5110 5120 5130
A c I
GGCGGAGGGCCTCCAACACAGTAATTTTCCTCCCGACTCTTAAAATAGAAAATGTCAAGTCAGTTAAGCAGGAAGTGGACTAACTGACGCAGC
, 4 .
SV40core Adcore
5120
)CLEAR 3T6 cells I I
CTOR 5114
t
t
5130NDING PEAl PEA2
(AP1)
5140 5150 5160 5170 5180 5190 5200 5210 5220 5230
l l IT A IA I I
TGGCCGTGCGACATCCTCTTTTAATTAGTTGCTAGGCAACGCCCTCCAGAGGGCGTGTGGTTTTGCAAGAGGAAGCAAAAGCCTCTCCACCCAGGCCTA
Ig SV40core
ICLEAR 3T6cells
2°
PEBI
,CTOR 2 1
JDING 5147 5162 5184 5194
5159 F9 cells
5155 5174
2° 10 2°
Rat Liver I_
5159 5175 5185 A 5201 52IS 5228
EBP20
FIG. 8. Schematicof features withinoradjacenttothePyenhancer.Thetoplinerepresentsnoncoding PyDNA numberedbythe method of Soedaetal. (49).Thesequences missingfrom the viableviruses arerepresented bythe thin lines which connect thestippledboxes. The regionsthatareduplicatedinhostrangemutantscapableofreplicatinginPCC4orF9cellsarerepresented byhatched boxes.One mutant able to replicate in F9cells differs from wt Py by a single A-to-G base substitution at nt5230. E.C., Embryonal carcinoma. Sequences duplicatedin severalwtstrains ofPyaremarkedbythe black box. Thenumbered, openboxesdepictthePyenhancer elements. The DNA sequence of the enhancerisdepicted below the boxes intwosegments. The sourceandbindingsitesofvarious nuclear factorsareshown below the sequence. The boxes represent the limits of regions protected from DNaseI digestion by the various nuclear factors. Ad, Adenovirus; Ig,immunoglobulin; 10 and20,bindingsites.
Pysuggestedthat the enhancer regioncontainstwo
enhanc-ers with different cell specificities. Atest ofthispossibility withDNAfragmentsfrom the enhancerregionofPyisolated from the wtgenome, several host range mutants, and awt
straincarryingaduplicationof enhancersequencescloned in avectorcarryingthechickenax-2 collagenpromoterlinked to the cat gene supported this model (17). These authors showed that a 134-bp fragment from the early half of the enhancer (the PvuII-4 fragmentofPy, nt 5131 to nt 5265), which was named element B, increased catexpression, on average, 3-fold inmouse3T6 fibroblastsorPCC3embryonal
carcinoma cells, whereas the late half of the enhancer region, the A element (the BcII-to-PvuII fragment from nt 5021 to nt 5130), increased cat expression, on average, 13-fold in 3T6 cells but only 4-fold in PCC3 cells. The B enhancer from host rangemutantsofPy capableof replicat-ing in embryonal carcinoma cell lines functioned with in-creased efficiency relativeto thewtB enhancertoaugment
cat expression to the same extent in both 3T6 and PCC3
cells, confirming previous results (26). We did not address
thecell typespecificity of the various enhancerelementsas we have defined them, but our results are qualitatively SOSC
5265
5135
5230
A
F-9G
5215 5233
5229
A I I a I I I I I I I I I
VOL.62, 1988
5075U c-LMImr-N I0
on November 10, 2019 by guest
http://jvi.asm.org/
1676 MUELLER ET AL.
similar to those of Herbomel et al. (17) and define in greater detail the locations of the various elements. The A enhancer is essentially equivalent to enhancer elements 1 and 2,
whereasthe B enhancer is equivalent to enhancer element 3.
Nucleotide sequence comparisons between the Py
en-hancer elements reveal little homology, but each element
contains short sequence motifs that have been identified in
otherenhancers (Fig. 8). The SV40 enhancer core sequence,
5'-G(C)G(C)C(T)CTGGA(T)A(T)A(T)-3', is found in
en-hancerelement 1 (an 8-of-10-bp match:
5'-CGGGAGGAAA-3') and in element 3 (a 9-of-10-bp match;
5'-AGTGTGGTTT-3'). There is a perfect match to the human adenovirus
enhancer core, 5'-A(C)GGAAGTGAA(C)-3', in enhancer
element2, and a sequence found in the mouse
immunoglob-ulin heavy-chain gene enhancer, 5'-TCCTTTTTAA-3', is
also perfectly conserved in enhancer element 3 (2). Two of the enhancer elements (2 and 3) also contain inverted repeats near the enhancer core sequences. Each of these sequence
motifs is likely to be functionally important because their
deletion or mutation impairs the activity of the elements.
Moreover, many of these sequence elements are protected
from DNaseI digestion by cellular proteins (Fig. 8).
Deletion of the SV40 enhancer core motif in element 1
slightly reduced the activity of the enhancer to 67% of the
control, and addition of sequences (nt 5057 to nt 5073) that
containthis motif to element 2 (nt 5073 to nt 5130) increased
the activity of element 2 fourfold (from 8 to 36% of the
control). Moreover, the inherent weakness of element
1-element1 by itself in pTE1 did not increase cat expression in 3T3 cells-correlates with the relatively poor match of the
SV40enhancercore sequence in this element to the consen-sus. In this regard, it may be noteworthy that DNA-binding
assayshave failed to detect the interaction of cellular factors withthis element in vitro. These results are suggestive ofa role for the SV40 core sequence in enhancer element 1
functionbut are clearly not conclusive.
Enhancer element2 contains an adenovirus enhancer core
sequence flankedby an inverted repeat (Fig. 8). Deletion of the lateregionproximal inverted repeat sequence (nt 5096 to
5104)from an element 2 plus element 3 configuration of the
enhancerhad little or no effect on its activity, but deletion of theearly-sideinverted repeat from an element 1 plus element 2construct abolishedenhancer activity. These results imply
that the hairpin structure that could be generated between
these two sequences is not important for the functioning of
the elements and that the two sequences, despite their
common primary structure, are not functionally equivalent.
Perhaps it is the orientation or position of the conserved
inverted repeatsequence relative to the adenovirus enhancer
corethat is important for the proper functioning of element 2. Deletion ofthe first 5 bp of the adenovirus enhancer core
sequence caused a threefold reduction in the activity of a
2-plus-3
configuration
of the enhancer relative to a plasmidthat retained these sequences. It may be noteworthy that
only part of this 10-bp sequence, the last 5 bp
(5'-TGACT-3'),iscommonly duplicatedin various wt strains of Py. The
commonlyduplicated sequence spans nt 5114 to nt 5137 (42).
Whatever the role of the adenovirus enhancer core
se-quence,it is clearthat it cannot function by itself to endow
enhancer element 2 with activity. If the enhancer core
sequence is important, then it must be coupled to the
inverted-repeat
motifon its right to be active. Similarly, theearly-side inverted repeat alone does not represent the
activity of enhancer element 2, because it alone, when
coupledtoenhancerelement3, does not increase the activity
ofthat element. Therefore, both the adenovirus enhancer
core motif and the inverted repeat toits right arerequired for enhancer element 2function. This inference is substantiated by the observation that reiteration of a 26-bp fragment that includes both of these sequences (nt 5108 to nt 5130) forms an active enhancer when tested in HeLacells (49). Recently, Piette and Yaniv (41) have provided evidence that two separate cellular factors (PEAl and PEA2) interact with enhancer element 2 (Fig. 8). The sequences that are pro-tected from DNase I digestion by these two factors
corre-spond almost exactly to the minimal sequences required to constitute an enhancer upon reiteration. PEAl may be the same asAP1,
which binds to elements of the SV40 and human metallothionein enhancers by recognizing the se-quence 5'-TGANTA(C)A-3' (24, 25). The latter sese-quence overlaps with the last 4 bp of the adenovirus enhancer core sequence, 5'-A(C)GGAAGTGAA(C)-3'. PEA2 interacts with inverted-repeat sequence to the right of thePEAl-binding site.
Enhancer element 3 contains several notable sequence features, including a sequence homologous to one found in the immunoglobulin enhancer, an SV40 enhancer core, and several G-C-rich inverted repeats (Fig. 8). The relative contribution that each of these makes to the activityof the enhancer is not clear from our analysis, because few deletion mutants were isolated and characterized that independently affect them. However, our results suggest that the major activity of element 3 resides between
nt
5175 and nt 5229, which contains all of these features. This was indicated by analysis of 3' deletion mutants of a 2-plus-3 configuration of the enhancer. A threefold drop in activity resulted when sequences between nt 5229 and nt 5175 were deleted, but no further decline was observed when additional sequences between nt 5175 and nt 5130 were removed. These results imply that the latter sequences (nt 5130 to nt 5775) by themselves contribute little to the activity of the element. This is consistent with the observation that the reiteration of sequences between nt 5130 and nt 5167 did not form an active enhancer when assayed in HeLa cells (49).Nuclear factors have been characterized from a variety of sources that bind to enhancer element 3 (Fig. 8). Two groups have demonstrated the binding of a factor(s) from F9 embry-onal carcinoma cells to sequences between nt 5155 and nt 5174 (13, 38), and one of them has shown that sequences between nt 5150 and nt 5178 are sufficient for binding of the F9 factor (38). These sequences overlap partially with an inverted repeat but not with the
SV40
enhancer core se-quence. A factor from 3T6 cells has also been characterized which binds primarily to sequences between nt 5184 and nt 5194, corresponding to theSV40
enhancer core motif, and secondarily to sequences between nt 5147 and nt 5162, which include part of the immunoglobulin sequence (40). The primary binding site is sufficient to allow specific factor interaction with the DNA, whereas binding of the same factor to the secondary site is dependent on the initial interaction of the factor with the primary site and is appar-ently sequence independent (40). Finally, a purified factor (EBP20) from rat liver binds primarily to sequences between nt 5185 and nt 5201, corresponding almost precisely to the SV40 enhancer core in element 3. At higher concentrations, EBP20 binds to two secondary sites between nt 5159 and nt 5175 and between nt 5215 and nt 5228 (21). It is not known whether binding to the secondary sites requires prior binding to the primary site or not, but much higher concentrations of the factor are required to achieve binding at the secondary sites relative to the primary site. The relationship among J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
these variousfactors is not known, nor is itknown whether
they act as activators or repressors ofenhancer function.
We did not rigorously examine whether each enhancer
element has intrinsic activity. However, it may be significant that both enhancer elements 2 and 3, but not 1, indepen-dently increased cat expression in 3T3 cells in the context of the pTE1 vector (Table 1). Element 3 increased expression 3-fold, whereas element 2 increased expression 4- to 12-fold,
depending on its orientation relative to the promoter in
pTE1. As noted previously, a 26-bp fragment that includes sequences mapping entirely within element 2 (nt 5108 to nt 5130) is able to enhance transcription in HeLa cells if reiterated five times (49). Enhancer elements 2 and 3 have been previously identified asreplication activators,aand 3,
respectively(36; W. J. Muller, D. Dufort, and J. A. Hassell,
manuscript in preparation), and each of these elements
independently activatesPy DNA replication in 3T3 cells. By contrast, enhancer element 1 does not activate replication nor does it independently enhance transcription in these
cells (Mulleretal., in preparation). We are currently testing whetherreiteration of each of theelements results in
forma-tion of an enhancer. Our expectation, based on previous results with Py (49) and SV40 (37, 43), is that each element willconstitute afunctional unit, but more experiments will have to be performed to define the minimal functional
domain of each element and the best conditions to assay
their activity.
The mechanism whereby enhancers act to increase
tran-scription is not known. It is very likely that components of the transcriptional machinery interact with the various se-quences in the enhancer elements. The fact that pairs of enhancer elements function much better than the additive
effect of each element and that reiterations of the same
elementfunctionbetterthan a single element argue that the factors that bind to their cognate sites in the elements
interact withoneanother to stabilize their binding to DNA or thatcombinationsof factors interact much better with other
components of the transcriptional apparatus (i.e., RNA
polymeraseII)thanasinglefactor does. Although we did not
directly test which of these alternatives is correct, it is
noteworthy that the spacing between enhancer elements
does not seem to be critical for their cooperative function.
For example,deletion of3, 10, 15, 20, or 37 bp fromelement
2,combined withacorresponding insertionof 16 bp of linker DNA, yielded element 1 plus three configurations of the enhancer which were all about equally active, and various
strains and host range mutants of Py exist which carry
deletions, duplications, or both within the enhancer which
disrupt the spacing between pairs of enhancer elements,
apparently without deleterious consequences (reviewed in
reference 1). These observationsargue against the idea that
protein-proteincontacts between factors aligned on adjacent
enhancer elements are critical for enhancer function.
In-stead, theyfavor thenotion that the activity of the enhancer resultsfromtheabilityof factors bound to it to interact with
othercomponents ofthetranscriptional machinery, such as
factors boundtopromoter elements or RNA polymerase II.
Resolution ofthis, however, will require moreexperiments
with mutants containing lesions that deliberately interrupt the distance between the various enhancer elements or between multiples of the same element.
ACKNOWLEDGMENTS
We thankMonicaNaujokas for excellent technical assistance and Paula Bauersfeld for typingthemanuscript.
This research was supported by theMedical Research Councilof Canada andthe National Cancer Institute of Canada. C.R.M. was supported by a graduatefellowship from the Natural Sciences and Engineering Research Council ofCanada, W.J.M. was supported by a graduate fellowship from the Medical Research Council of Can-ada, and J.A.H. is a Terry Fox Cancer Research Scientist of the National Cancer Institute of Canada.
LITERATURE CITED
1. Amati,P. 1985. Polyoma regulatory region: a potential probe for mouse cell differentiation. Cell 43:561-562.
2. Banerji, J., L. Olson, and W. Schaffner. 1983. A lymphocyte-specific cellular enhancer is located downstream of the joining region in immunoglobulin heavy chain genes. Cell 33:729-740. 3. Banerji, J., S. Rusconi, and W. Schaffner. 1981. Expression of a
,3-globin gene is enhanced by remote SV40 DNA sequences. Cell27:299-308.
4. Bohnlein, E., K. Chowdhury, and P. Gruss. 1985. Functional analysisof theregulatoryregionofpolyomamutant F9-1 DNA. Nucleic Acids Res. 13:4789-4809.
5. Bohnlein,E., and P. Gruss. 1986. Interactionof distinct nuclear proteins with sequences controlling the expression of polyoma-virus early genes. Mol. Cell. Biol. 6:1401-1411.
6. Cowie, A., C. Tyndall, and R. Kamen. 1981. Sequences at the capped 5'-ends of polyoma virus late region mRNA's: an example ofextreme terminal heterogeneity. Nature (London) 307:241-245.
7. Davidson, I., C. Fromental, P. Augereau, A. Wildeman, M. Zenke, and P. Chambon. 1986. Cell-type specific protein binding to the enhancer of simian virus 40 in nuclear extracts. Nature (London) 323:544-548.
8. deVilliers, J., and W. Schaffner. 1981. A small segment of polyomavirus DNA enhances the expression of a cloned
P-globin gene over a distance of 1400 base pairs. NucleicAcids Res.9:6251-6264.
9. Edlund, T., M. Walker, P. Barr, and W. Rutter. 1985. Cell-specific expression of the rat insulin gene: evidence for role of two distinct 5' flanking elements. Science 230:912-916. 10. Favaloro, J., R. Treisman, and R. Kamen. 1980. Transcription
maps of polyoma virus-specific RNA: analysis by two-dimen-sional nucleaseS1 gel mapping. Methods Enzymol. 65:718-749. 11. Featherstone, M. S., M. A. Naujokas, B. J. Pomerantz, and J. A. Hassell. 1984. A plasmid vehicle suitable for the molecular cloning and characterization of mammalian promoters. Nucleic Acids Res. 12:7235-7249.
12. Fromm, M., and P. Berg. 1982. Deletion mapping of DNA regions required for SV40 early region promoter functions in vivo. J. Mol. Appl. Genet. 1:457-481.
13. Fujimura, F. 1986. Nuclearactivity fromF9 embryonal carci-noma cells binding specifically to the enhancers of wild-type polyoma virus and PyEC mutant DNAs. Nucleic Acids Res. 14:2845-2861.
14. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 15. Hearing, P., and T. Shenk. 1983. The adenovirus type 5ElA
transcriptional control region contains a duplicated enhancer element. Cell 33:695-703.
16. Hen, R., E. Borrelli, P.Sassone-Corsi, and P. Chambon. 1983. Anenhancer element is located 340 base pairs upstream from the adenovirus-2 ElA cap site. Nucleic Acids Res. 11:8747-8760.
17. Herbomel, P., B. Bourachot, and M. Yaniv. 1984. Two distinct enhancers with different cell specificities coexist in the regula-toryregion of polyoma. Cell39:653-662.
18. Herr, W., and J. Clarke. 1986. TheSV40enhancer is composed of multiple functional elements that can compensate for one another. Cell 45:461-470.
19. Herr, W., and Y. Gluzman. 1985. Duplications of a mutated simian virus 40 enhancer restore its activity. Nature (London) 313:711-714.
20. Jat, P., J. Roberts,A. Cowie, and R. Kamen. 1982. Comparison ofpolyomavirus early and late promoters by transcription in
on November 10, 2019 by guest
http://jvi.asm.org/
1678 MUELLER ET AL.
vitro. Nucleic AcidsRes. 10:871-889.
21. Johnson, P., W. Landschulz, B. Graves, and S. McKnight. 1987. Identification of rat liver nuclear protein that binds to the enhancer core element of three animal viruses. Genes Dev. 1: 133-146.
22. Laimins, L. A., G. Khoury, C. Gorman, B. Howard, and P. Gruss. 1982.Host-specific activation of transcription by tandem repeatsfrom SV40 and MSV. Proc. Natl. Acad. Sci. USA 79: 6453-6457.
23. Laimins, L. A., P. Tsichlis, and G. Khoury. 1984. Multiple enhancer domains in the 3' terminus of the Prague strains of Roussarcomavirus. Nucleic AcidsRes. 12:6427-6442. 24. Lee, W., A. Haslinger, M. Karin,and R.Tjian. 1987.Activation
of transcription by two factors that bind promoter and enhancer sequencesof the human metallothionein gene and SV40. Nature (London) 325:368-372.
25. Lee, W., P. Mitchell,and R. Tjian. 1987. Purified transcription factor AP-1 interacts with TPA-inducible enhancer elements. Cell 49:741-752.
26. Linney, E., and S.Donerly. 1983. DNA fragments from F9 PyEC mutantsincreaseexpression of heterologous genes in transfect-ed F9 cells. Cell35:693-699.
27. Lusky, M., and M.Botchan. 1981.Inhibition of simian virus 40 replication in simian cells by specific pBR322DNAsequences. Nature(London) 293:79-81.
28. Luthman, H., M. G. Nilsson, and G. Magnusson. 1982. Noncon-tiguous segments of the polyoma genome required in cis for DNAreplication. J. Mol. Biol. 161:533-550.
29. Maniatis, T., E.F. Fritsch, and J. Sambrook. 1982. Molecular cloning:alaboratory manual. ColdSpring HarborLaboratory, Cold Spring Harbor, N.Y.
30. Maniatis, T.,S.Goodbourn, and J.A.Fischer. 1987.Regulation of inducible and tissue-specific gene expression. Science 236: 1237-1245.
31. Maxam, A. M., and W.Gilbert. 1980. Sequencing end-labeled DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
32. McKnight, S., E. Gavis, R. Kingsbury, and R. Axel. 1981. Analysis oftranscriptional regulatory signals of the HSV thy-midine kinase gene: identification ofanupstreamcontrolregion. Cell 25:385-398.
33. McKnight,S.,and R.Tjian. 1986.Transcriptionalselectivity of viral genes inmammalian cells. Cell 46:795-805.
34. Moreau, P.,R.Hen,B.Wasylyk,R.Everett,M. P.Garb,and P. Chambon. 1981. The SV4072 base pair repeat has a striking effect on gene expression both in SV40 and other chimeric recombinants. Nucleic Acids Res.9:6047-6068.
35. Mueller, C. R., A. M. Mes-Masson, M. Bouvier, and J. A. Hassell.1984. Locationof sequences inpolyomavirusDNAthat arerequiredforearly geneexpressionin vivo andin vitro. Mol. Cell. Biol. 4:2594-2609.
36. Muller, W.J., C. R. Mueller, A.-M. Mes, andJ. A. Hassell. 1983. Polyomavirus origin forDNAreplication comprises mul-tiple genetic elements. J. Virol. 47:586-599.
37. Ondek, B.,A.Shepard, and W. Herr. 1987. Discrete elements within the SV40 enhancerregion display different cell-specific enhancer activities. EMBOJ. 6:1017-1025.
38. Ostapchuk, P., J. Diffley, J. Bruder,B.Stillman,A.Levine,and P. Hearing. 1986. Interaction of a nuclear factor with the polyomavirus enhancer region.Proc. Natl. Acad.Sci. USA83:
8550-8554.
39. Piette, J., M. H. Kryszke, and M. Yaniv.1985. Specific interac-tion of cellularfactors with theB enhancer ofpolyoma virus. EMBO J.4:2675-2685.
40. Piette, J., and M. Yaniv. 1986. Molecular analysis of the interaction between an enhancerbinding factor and its target DNA. Nucleic Acids Res. 14:9595-9611.
41. Piette, J.,and M. Yaniv.1987. Twodifferent factors bindtothe a-domain of the polyoma virus enhancer, one of which also interacts with the SV40 and c-fos enhancers. EMBO J. 6:1331-1337.
42. Ruley, H. E., and M. Fried. 1983. Sequence repeats in a polyomavirus DNA region important for gene expression. J. Virol. 47:233-237.
43. Schirm, S., J. Jiricny, and W. Schaffner. 1987. The SV40 enhancercanbe dissected intomultiple segments, each witha different cell typespecificity. Genes Dev. 1:65-74.
44. Scholer, H., andP. Gruss. 1984. Specific interaction between enhancer-containing molecules and cellular components. Cell 36:403-411.
45. Sharp,P.A.,A.J. Berk,and S.M.Berget. 1980.Transcription mapsof adenovirus. MethodsEnzymol.65:750-768.
46. Soeda, E., J. Arrand,N.Smolar, J. Walsh,and B.Griffin.1980. Coding potential and regulatory signals of the polyoma virus genome. Nature(London) 283:445-453.
47. Swimmer, C., and T. Shenk. 1984. A viable simian virus 40 variant that carriesa newlygenerated sequence reiteration in place of the normal duplicated enhancer element. Proc. Natl. Acad. Sci. USA81:6652-6656.
48. Tang, W.J., S. L.Berger,S. J. Triezenberg, and W. R. Folk. 1987. Nucleotides in the polyomavirus enhancer that control viral transcription and DNA replication. Mol. Cell. Biol. 7: 1681-1690.
49. Veldman, G.M., S.Lupton,and R.Kamen.1985.Polyomavirus enhancercontains multiple redundant sequence elements that activate bothDNAreplication and geneexpression. Mol. Cell. Biol.5:649-658.
50. Weaver,R.F.,and C.Weissmann. 1979.Mapping ofRNAbya modification of theBerk-Sharp procedure: the5'termini of 15S
p-globinmRNAprecursorand mature10S,-globinmRNAhave identical mapcoordinates. Nucleic AcidsRes. 7:1175-1193. 51. Weber, F., J. deVilliers, and W. Schaffner. 1984. An SV40
"enhancer trap" incorporates exogenous enhancers or gener-atesenhancersfrom itsownsequences. Cell36:983-992. 52. Weiher, H.,and M. Botchan.1984.Anenhancer sequence from
bovinepapilloma virusDNAconsists oftwoessentialregions. Nucleic AcidsRes. 12:2901-2916.
53. Weiher, H., M. Konig, and P. Gruss. 1983. Multiple point mutations affecting the simian virus40enhancer. Science219: 626-631.
54. Wigler, M., A. Pellicer, S. Silverstein, and R. Axel. 1978. Biochemical transfer ofsingle-copyeukaryotic genesusingtotal cellularDNA asdonor. Cell 14:725-731.
55. Wildeman, A. G., M. Zenke, C. Schatz, M. Wintzerith, T. Grunstrom,H.Matthes, K.Takahashi,and P. Chambon. 1986. Specificprotein bindingtothe simian virus40enhancerinvitro. Mol. Cell. Biol.6:2098-2105.
56. Zenke, M., F. Grundstrom, H. Matthes, M. Wintzerith, C. Schatz,A.Wildeman,and P. Chambon.1986.Multiple sequence motifs are involvedin SV40 enhancerfunction. EMBOJ. 5:387-397.
J. VIROL.