VIROLOGY, Oct. 1977,
Copyright ©1977 AmericanSociety forMicrobiology Printed in U.S.A.
Electron Microscopic Characterization of the
Defectiveness
of
a
Temperature-Sensitive
Mutant
of
Moloney
Murine
Leukemia
Virus
Restricted
in
Assembly
P. H. YUEN AND P. K. Y. WONG*
Department of Microbiology and School of Basic Medical Sciences, University of Illinois, Urbana, Illinois61801
Received for publication 6 May 1977
The effect of temperature shiftdown on the assembly of ts3 virions was
investigated by bothscanning (SEM)andtransmission (TEM) electron
micros-copy. Ts3 is a spontaneous temperature-sensitive mutant of Moloney murine leukemia virus (Mo-MuLV) which previous studiesindicatedtobe defectivein
assembly or release of the virions. Inthe present study, both SEM and TEM revealedthe following: (i) there were morecell-associated virionsints3-infected
cells grown at the nonpermissive temperature (390C) than either in cells grown
atthepermissivetemperature (340C) or in wild-type MuLV-infected cells grown
at
39QC;
(ii) there were morenormal single particles than multiploids (virionswith two or more pieces ofgenomic RNA) in ts3-infected cells grown at the
nonpermissive temperature; (iii) there were more multiploids in ts3-infected
cellsgrownatthenonpermissive temperature than either in cells grownatthe permissive temperatureor inwild-type MuLV-infectedcells grownatthe
non-permissivetemperature; (iv)upontemperature shift from 39to34WC,about90%
ofthecell-associatedvirionsdissociatedfromthe cell surface. TEM studiesalso
indicated that upon temperature shiftdown, virion assembly rapidly occurred. The above observations suggest that faulty assembly, which results in the production ofmultiploids, may not be the reason why ts3 virions accumulate on
the cell surfaceatthe nonpermissivetemperature.Therelativelyhigher
propor-tionofmultiploidsfoundints3-infected cells grownat39°Ccomparedwiththose
grown at34°C may be due tothe higher density ofbuddingvirions atthe cell
surfaceatthenonpermissivetemperature,which increases thepossibilityoftwo
or more particles assembling close to one another. The accumulation of ts3
virions in all stages ofassembly at the nonpermissive temperature, together
with the fact thatrapid assemblyand releaseofts3virionsoccurredon tempera-ture shiftdown, indicates that virion assembly is restricted after it has been initiated. Theprobablerole ofalteredglycoprotein(s)inrestrictingvirion
assem-blyisdiscussed.
One of the major eventsinthe replication of RNA tumor viruses is the assembly of the
viri-ons at the cell membrane. As yet the
mecha-nisminvolved ispoorly understood. It has been postulated (1, 4) that assembly of the virions is regulated by protein-protein interactions. One
approach toward understanding the processes
involvedinthe assembly of RNA tumor viruses is by defining the temperature-sensitive (ts) defects of viral mutants blocked in assembly. Recently, such mutants from both avian and mammalian RNAtumorviruses have been iso-latedand partially characterized (2, 3, 6-9, 11).
Ts3 is a spontaneous ts mutant of Moloney murineleukemia virus(Mo-MuLV) isolated by
Wong et al. (9). Previous studies (7, 8) have
indicated that the ts defect of ts3 may be dueto ablock either late intheassembly
proc-ess orintherelease ofthe virions. The present
investigation on the assembly of ts3 virions
further defines thets defect ofts3.
MATERIALS AND METHODS
Cells. Detailedproceduresfor thepropagationof TB cells, a fibroblastic cell line established from mixed cultures of bonemarrowandthymusCFW/D mouse cells, have beenpreviouslydescribed (7).
Virus. TB cellswere infectedin suspensionat a
multiplicity of infection of 5 with wild-type Mo-MuLV orts3,a ts mutantof MuLV. Infected cells(3 x 10') wereplated on each 60-mm plastic petri dish and incubatedatthe permissive (34°C) or nonper-missive (39°C) temperature for 48 h. Ts3-infected 222
on November 10, 2019 by guest
http://jvi.asm.org/
ts MUTANT OF MuLV RESTRICTED IN ASSEMBLY 223
cellswerealsoincubatedatthenonpermissive
tem-peraturefor48hand then shiftedtothepermissive
temperaturefor30or60min.
Transmission electron microscopy(TEM).Viral
pelletsandwild-typeMuLV-orts3;infectedTBcells
scraped offwitharubberpolicemanwerefixedin 2%
glutaraldehyde in sodiumcacodylatebufferor
phos-phate-buffered salineandthen postfixedin 2%osmic
acid overnightor inafreshly prepared mixtureof
0.12gof potassiumferrocyanidein 6mlof1%osmic
acid. After dehydrationthroughgraded ethyl alco-hol, the cellswereembedded in Spurr'splastic.
Sec-tionswere mounted directlyon300-meshgrids and
stainedinuranylacetate at70°Cinawaterbathand
inReynoldslead citrate. Specimenswereviewed in
a Siemens 102 electron microscope at 80kV. For
quantitationofvisions inthin sections,cellsections
werechosenatrandom,and the typeofvirionand
thestageinassembly of thevisionswerescoredby
viewingatamagnificationof x80,000.
SEM. The details for preparing infectedcellsfor
scanning electronmicroscopy (SEM)and
quantita-tion of the cell-associated virions have been
de-scribed(8).
RESULTS
Relative proportion of normal C-type particlestovirionaggregatesints3-or
MuLV-infected cellsasdetermined by SEM.We
rea-soned thatif mostofthets3particlesproduced
at the nonpermissive temperature (39°C) are
"polymorphic" as reported by Wong and
Mc-Carter (7), in contrasttotheproduction of
nor-mal C-type particlesatthepermissive
tempera-ture (34°C), faulty assembly of the virion at
39°C may beindicated. Ts3-infected cellswere
therefore grown at 34 or 39°C for 48hor 39°C
for48 h and then shifted to 34°C for 15 or 60
min; MuLV-infected cells were grown at 39°C
for 48 h and preparedfor SEMasdescribed in
Materials andMethods.
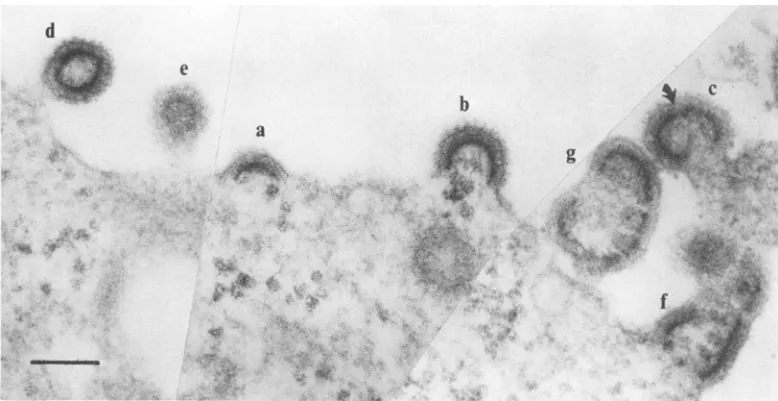
Figure 1A shows arepresentative portion of
the cell surface of a ts3-infected cell at the nonpermissive temperature 48hpostinfection. Most of the particles present were normal
C-type spherical forms. Scattered among them wereduplexes(black arrow) and short
microvil-lus-like structures (white arrow), which may
consist oftwoor morevirions alignedin
tan-dem(Fig. 1B). Triplexes (Fig. 1C and D), quad-ruplexes (Fig. 1E), and largeraggregates (Fig.
1F and G) were also observed in ts3-infected
cells grownat the nonpermissive temperature orgrownatthenonpermissivetemperaturefor 48hand thenshiftedtothepermissive temper-ature for 15min.
In ts3-infected cells grown at 34°C and in
MuLV-infected cellsgrownat39°C, the number
of particles observed on the cell surface was
greatly reduced as compared withts3-infected
cellsgrownatthe nonpermissivetemperature.
Duplexes and occasionally triplexes were also
observed in the former cultures; however,
larger virion aggregates were not found.
The relative proportions of normal C-type
particlestoaggregatesinthe different cultures
examined are shown in Table 1. The figures presented in Table 1 may be only rough esti-mates,because onlyvirions inthe latestagesof
assembly could be identified with certainty.
The total number of aggregates may also be underestimated, since rod-shaped virions may be mistaken for microvilli. Atypically shaped or
unusually large spherical particles were not
included in the counts recorded in Table 1.
Since the same types of errors were
encoun-tered inall the samples examined, the figures
presented in Table 1 should be anindicationof
real differencesamongthe samples studied.
The resultspresented in Table 1 may be
sum-marizedasfollows: (i) there were more virions
onthe cell surface of ts3-infectedcells grown at
390C thanonts3-infected cellsgrown at34WC or
onMuLV-infected cells grown at39"C;(ii) more
than90%ofthe ts3 particles that had
accumu-lated on the cell surface at 390C disappeared
from the cell surfacewithin 1hafter
tempera-ture shiftdown; (iii) there were more normal
single particles than aggregates inthe
ts3-in-fected cellsgrown at390C; (iv)there were more
aggregates in the ts3-infected cells grown at
390C thanineither ts3-infectedcellsgrown at
34WC orMuLV-infected cells grown at390C. Theabovefindingssuggestthat formationof
morphologically aberrant particles may not be
thereasonwhyts3 virionsfail tocomplete
as-sembly and release from the cell surfaceatthe
nonpermissive temperature. The relatively
higher proportion of aggregates found in the ts3-infected cells grown at390C couldbe due to
thehigherdensity ofbuddingvirions atthe cell
surface, which increases theprobabilityoftwo
or more particles assembling close to one an-other. Furthermore, the accumulation of viri-ons atthe nonpermissive temperature may be attributed eithertoablockinsomeprocesses)
in virion assembly or to a ts defect in the release of the virion per se.This latter possibil-ity would be indicated ifmost of the particles werecompletelyassembled, but failed to
disso-ciatefrom the cell surfaceatthenonpermissive
temperature.
TEMstudies.Toinvestigatetheabove possi-bilities, TEM studies were conducted on
ts3-infected cellstodetermine thestageinassembly
at which the assembly of ts3 is blocked at the
nonpermissive temperature, and whether fur-ther assembly of these ts3 virions can take
place ontemperature shiftdown.
Ts3-infected TBcells grown at 34 or 39°C for
48h, and ts3-infectedTBcellsgrownat39°Cfor
VOL. 24, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
YUEN AND WONG
r,
-FIG. 1. (A) SEMofaportion ofa ts3-infected TB cell surface grown at the nonpermissive temperature showing normalspherical virionsaswellasduplexes (blackarrow) and short microvilli that may be short rod-shapedvirions(white arrow). Scale=500 nm.(B-G)Scale=250nm. (B) Shortrod-shaped virions; (C andD) triplexes;(E) quadruplexes; (FandG)largevirionaggregates.
no. -I
tk o' a
.T.
.,. 6 7,", , *.u
v, I
:1...
't -I
.0,'I"
I. .1'
I
-"
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.69.461.73.639.2]ts MUTANT OF MuLV RESTRICTED IN ASSEMBLY 225
48 hand thenshifted to34TC for 30 or60min,
wereprepared for TEMasdescribed in Materi-als and Methods.
Ts3 virons. Thin-section electron microscopy ofts3-infectedcells grownatthenonpermissive temperature revealed the presence of both
nor-mal C-type particles andmultiploids (particles
with more than one ribonucleoprotein compo-nent) in all stages ofassembly. The structure
and assembly of both types of particles have been reportedrecently (12).
Effect of temperature shiftdownon the
as-sembly of ts3 virions. A composite view of
ts3-TABLE 1. Relative proportionsofnormalC-type particles to aggregatesa ints3-orMuLV-infected
cultures determinedby SEM
Total cell Total no. % of ag-surfaeof art'- gregates Type ofculture srfaex of 1 tototal
area exam- cles/100- no. of par-ined (Am') Am2area
tiles
ts3,390C(48h) 705 250 18
ts3, 39 -- 34"C 326 228 18
(15min)
ts3,
39-34"C 285 11 3(60min)
ts3,340C (48 h) 367 38 8
MuLV, 39"C 825 29 8
(48h)
aDuplexes, triplexes, etc.
infected cells grown atthe nonpermissive
tem-perature for 48 h is shown in Fig. 2. TEM indicates that although both normal particles andmultiploidsindifferent stagesinassembly were present ints3-infected cells grown at the nonpermissive temperature, less than 20% of these particles werecompletely assembled (Fig. 2d) and on the verge of dissociating from the cellsurface. This finding is consistent with the infrequent observation of extracellular
imma-tureC-type particles and the virtual absence of mature C-type particles in the intercellular spacesof ts3-infected cells grown at the nonper-missive temperature (Fig. 3A). The above ob-servations suggest that at the nonpermissive temperature, virion assembly is initiated but probably restricted. It was further reasoned that if ts3 is temperature sensitive in some
factor (function) restricting assembly, on
tem-peratureshiftdown,oneshould be able to detect rapidvirion assembly andrelease. This would be reflected ina change in the composition of virions in the different stagesofassembly. In anattemptto verify the abovehypothesis, the relative proportions of virions in the various stagesofassembly were comparedinthe follow-ing cultures: ts3 grown at 390C for 48 h, ts3 grownat39°Cfor48 h and thenshiftedto 34°C for 30 or 60min, andts3grown at34°Cfor48h. The normal budding C-type particles were
arbi-d
4.
e
b
-!1
I
.l, '.'-r
a
[image:4.504.56.445.398.599.2]4...0 g
FIG. 2. Composite electron micrograph ofa ts3-infected TB cell grown at the nonpermissive temperature for 48h.Scale =100nm.(a)Normal virion at an early stage in assembly with anarc-shapedribonucleoprotein
(RNP).(b)Normal virionat anintermediate stageinassembly with a semicircular RNP. (c) Spherical duplex
atalate stage in assemblyshowing a break in the RNP (arrow) and a larger size and broader linkage to the cellcompared with (d). (d) Normal virion at a very late stage in assembly showing complete assembly of the
RNP,narrowlinkagetothecell, and resealing of the cellular membrane. (e) Normal virion at a very late stage inassembly sectioned through the core shell. (p) Multiploid virion at an early stage inassembly showing breaksinthe RNP and broadlinkage to the cell. (g) Multiploid during the final stages in assembly showing similarcharacteristics as pointed out for the normal virion shown in (d).
VOL. 24, 1977
... l.
on November 10, 2019 by guest
http://jvi.asm.org/
A
c.i
C.'I
.. .S
A?
A.
..;R-A...
COOLA,
I'
hii -5Fs
'..
su3b N ,
o'aid
'it
Se or,
i:
e
as
all".}
B
V.
s
a.-1i
b'
a'X
aX
4$~~~~~~~
...
.i~~~~~~~~~~~~~A
w. .a.
*. i
! 4 i
AWL 4.
.F
L4NY;,
ALto
^j!
[image:5.504.71.457.39.618.2]zio
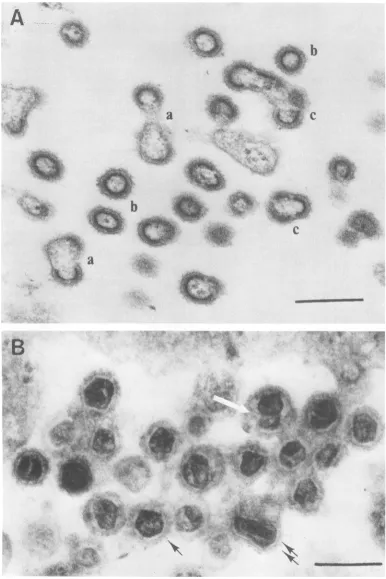
FIG. 3. (A)Uncommon viewshowinganaccumulationofbuddingvirions(a), completeassembled normal
virions (b), andmultiploids (c) in the intercellular spaceofts3-infected cellsgrownatthe nonpermissive
temperature.Scale=200nm.Not all thecompletelyassembled virionsarenecessarily extracellular,since the
section could havebeencutthroughthetransverseaxisofsomeofthe virions. (B)Accumulationofmature
particles in the intercellularspaceof ts3-infectedcellsgrownat thepermissivetemperatureshowingvariation in theshapeand sizeofthecoresofthe virions. Scale=200nm.Virion witharegular pentagonalcore(single
black arrow); virion with an elongated core (double black arrow); spherical virion with twocores (white
arrow).
226
...*
4 8
.t
.
b
Izi
I-I.--
-,W..I
c
-"'k
a
I,-I&;
'W"W ?
-14f
I,Qtwt
on November 10, 2019 by guest
http://jvi.asm.org/
ts MUTANT OF MuLV RESTRICTED IN ASSEMBLY 227
trarily separated into four groups according to their stage inassembly: early (Fig. 2a), inter-mediate (Fig. 2b), late (Fig. 4b), and very late (Fig. 2d and4c).
The results of this investigation are summa-rized in Table 2. In thets3-infected cells grown atthe permissive temperature, almost 90% of the particles observed were non-cellassociated,
and more than 90% of these were mature parti-cles with electron-dense nucleoid (Fig. 3B). Only 2% of the cell-associated virions were mul-tiploids. The normal C-typecell-associated par-ticlesappeared to be evenly distributed among the four stages of assembly.
Some marked differences were observed be-tween ts3 virions from cells grown at the per-missive temperature as compared with those grown at the nonpermissive temperature. There were 22 cell-associated virions per cell section in ts3-infected cells grown at 390C as compared with 0.5 virion per cell section in ts3-infected cells grown at 34WC. In contrast to the virions found in cells grown at the permissive temperature, 96%of the virions observed in the nonpermissive culture were cellassociated. The
few extracellularvirionsobservedwere
imma-ture (Fig. 3A). Maturevirions may have been
present insuchlownumbersthattheywere not
detected. In the culture grown at the
nonper-missive temperature, 29% of the total virions
observedweremultiploids, whereasonly2%of
thevirions inthe culturegrownatthe permis-sive temperature were multiploids. Fifty
per-cent of the normal C-type virions observed in
the culture grown at the nonpermissive
tem-perature were
early-budding
virions, whereasless than20%of them were in a verylatestage
inassembly.
Acomposite viewof ts3-infected cellsgrown
at390C for48hand then shiftedto34WCfor30
minisshowninFig.4.Severalmarkedchanges
were observed. The number of cell-associated
virions decreased to about 2 virions per cell
section,whereas theratioofmultiploidsto
nor-malparticles remainedthe same. The number
of extracellularparticles increased fromabout
4%toalmost50%of the total number of
parti-clesobserved, andmorethan80%of thesewere
immatureparticles. Amongthe cell-associated
normal C-type virions, over30%were late-bud-ding particles, and almost60%wereinthe final stagesofassembly.
As compared with a ts3-infected culture
shifted to the permissive temperature for 30
min, alonger shiftdown period (60 min) further reduced thenumber of cell-associated virions to about 0.5 virion per cell section. There was a
further increase in the proportion of non-cell-associatedvirions tocell-associated virions and
areduction in theproportion of multiploids to
total virions. Among the normal C-type parti-cles, there appeared to be a slight increase in the proportion of early-budding virions.
The above findings may be summarized as follows: multiploids were present in ts3-in-fected cells grown at both the permissive and nonpermissive temperature. Consistent with ourSEMfindings, the proportion of multiploids
to normal virions was higher in ts3-infected
cells grown at the nonpermissive than at the
permissive temperature. In spite of the fact
that 50% of the normal particles in the very
earlystages inassembly couldnotbe identified
inthe SEMstudies,ahigherproportionof mul-tiploids was observed in the TEM studies. This suggests that the duplexes, triplexes, etc.,
ob-served bySEM wereonlyaportionof the total
multiploids present. The proportion of
multi-ploids calculated from the TEMstudies didnot
exceed30%of the totalvirionsobservedin
ts3-infectedcells grown at the nonpermissive
tem-perature. Virionassembly rapidlyoccurred on
temperatureshiftdown with both normal parti-cles andmultiploids dissociatingfrom the cell
surface.
DISCUSSION
Our SEM and TEM studies confirmed the findings of Wong and MacLeod (8) that there were substantially more virions on the cell
surface ofts3-infected TB cells grown at the
nonpermissive temperature(39°C) than of cells grown atthepermissive temperature(34°C) or
ofMuLV-infected TB cells grown at39°C. Our
SEMstudiesagreewiththeir observationsthat
morethan 90% of the virions that had
accumu-latedatthe cell surface at 39°C dissociated from
the cell surface during the first 60 min after
temperature shiftdown. However, our TEM
studiesindicatethatwhen the accumulation of
virions on the cell surface is maximal, the total
cellsurfaceareathat can be occupied by virus
particles is greaterthan the 10% estimated by
Wong andMacLeod (8), since neither virions in
theveryearlystagesof assembly (about 50% of
the total number of normal cell-associated
C-type particles) nor many of the multiploids
(suchastherod-shapedvirions,the abnormally
large spherical forms, or the atypically shaped virions) were identified by SEM.
Ourpresentfindingssuggest thatthe
produc-tionofatypically shapedparticles
(multiploids)
atthenonpermissive temperature may not be a direct result of the ts defectof ts3. Less than
30% of the virionsobserved by TEMfrom
ts3-infected cells grownatthenonpermissive
tem-perature were multiploids. Moreover, multi-ploids, primarily duplexes, were also
consist-VOL. 24, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
ZIAMOR~ ~ ~ 4'
04,
.Yet
w ; --**.41r
-S
P
g
5",
a
c
b
*i
I
.14 s,.
1{Z
d
I.g
=.I
e
..
'
I
X .1
UI,4
it w~~~~~~~~~~~~~T-,
Ais. rb&* .>
VA~~~~~~~~~~~9
4'~~~~~~~A
vs W 4 0
4
*
fX
Y
gi
to *4
V''
E*L;s;t '
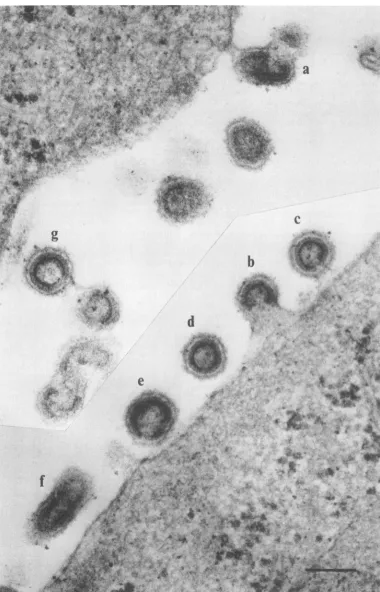
*~~~~~~~~~~~-FIG. 4. Ts3-infected TB cellsgrown at the nonpermissive temperaturefor48 h andthen shifted to the
permissivetemperaturefor30minshowingextracellularimmaturenormal virions(d)andmultiploids(e-g)
andnormalvirions(bandc)andmultiploids (a)at a lateorverylatestageinassembly.Scale =100nm.
228
I?'i
.!:..- I
P. d;
MkK-T.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.77.457.48.640.2]ts MUTANT OF MuLV RESTRICTED IN ASSEMBLY 229 TABLE 2. Relative proportionsofnormal virionstomultiploidsand virions in variousstages ofassembly in
differentts3-infectedculturesdeterminedbyTEM
%Mem- %Freeim- % of different development stages of Totalno. No. ofcell- brane-as- % Multi- mature membrane-associated normal
parti-ofparti- associated sociated ploidsof
particles
ofclesa
Tyofculture clesob- virions/ particles totalpar-non-mem-served cellsection of total tides
brane-as-
Inter- Late Ver particles sociated Early mediate latets3,340C 548 0.5 11 2 8 18 34 28 20
ts3, 390C 622 22 96 29 100b 50 13 18 19
ts3, 39- 34WC 559 2 53 28 82 3 7 31 59
(30min)
ts3, 39 . 340C 644 0.5 36 16 90 16 9 44 31
(60 min)
a SeelegendtoFig. 2and4.
bMatureparticles wereprobablypresent insuch low numbers thattheywerenotdetectable.
ently observed, althoughinlowernumbers, in
ts3-infected cells grown at 340C, in
MuLV-in-fected cellsgrownat 390C (12), andin a
Molo-ney murine sarcomavirus (Mo-MuSV) cellline
(unpublished data). The higher proportion of
multiploids to total particles found in
ts3-in-fectedcells grownatthenonpermissive
temper-ature maybeduetothe greater chance oftwo
or more particles assembling injuxtaposition becauseoftheaccumulationofbuddingviruses at the cell surface. The above suggestion is consistent with the observed decrease in the proportion ofmultiploids to total particles in
interferon-treated cells infected with ts3
(un-published data). Itwasshowninanotherstudy
(10) that the total number of ts3 virions
de-creasedbytwo- tothreefoldininterferon-treated
cells as compared with the controls. The
pro-duction ofatypically shaped virions in Friend
leukemia virus-infected cells after dimethyl
sulfoxide treatment has also been reported by
Sato et al. (5). In these dimethyl
sulfoxide-treated cells,therewas asignificantincreasein
the number of budding virions as compared
withthe controls.
The ts defect of ts3 appears to be due to some restriction in virion assembly. At the nonper-missive temperature, both normal and multi-ploid virions in all stages of assembly were observed. Only about 20%ofthenormal
parti-cles observed were at a very late stage in
as-sembly (Fig.2d).Four percentof the total
parti-clesobservedwereextracellular, noneof which were mature virions. However, aftera 30-min
shifttopermissive conditions, about 90% of the normal C-type virions observed were
late-bud-dingvirions(Fig. 4b), with almost60%of these
at avery late stageinassembly (Fig. 4d). This rapid increaseintheproportion of budding
viri-ons inthe finalstages of assembly on
tempera-ture shiftdown, inconjunctionwith a simulta-neousdecrease inthe number ofcell-associated
virions and an increase in the number of extra-cellular immature virions, suggests that rapid
assembly and release of the virions occurred
duringlte 30 min after shift to the permissive
temperature. On temperatureshiftdown,rapid
assembly and release of the multiploid virions
alsoseem tohave occurred. In the390C-infected
culture, mostof the multiploids observed were
broadly linked tothe cell at their bases (Fig.
2c). On temperature shiftdown, most of them
showed completeassembly of viral components,
resealing of the cellular membrane, and
shrinkingof the cytoplasmic linkage to the cell
(Fig. 2g and 4a). The large number of incom-pletely assembled ts3 virions in all stages of
assembly at the nonpermissive temperature
suggests that restriction in assembly only
oc-cursafterithas been initiated and that the ts
lesionis a step common to the whole assembly
process.
We have as yet no conclusive evidence as to how virion assembly is restricted at the nonper-missive temperature. Previous findings (7)
in-dicate that ts3 virions partially assembled on
the cell membrane of cultures held at
390C
be-fore the addition of actinomycin D or cyclohexi-mide are able to complete assembly and disso-ciatefrom the cell membrane on temperatureshiftdownto 34C inthe presence of the
inhibi-tors.Thissuggests that the ts defect liesbeyond events associated with the synthesis of viral
DNA, DNA-dependent RNA, and protein. The
ts defecthasalso been demonstrated (7) to be
reversible, because even in an established in-fection a shift from 34 to
390C
turns offvirus production almost immediately. In the same study, the ability of ts3 to rescue MuSV from an MuSV-infected nonproducer cell line was also examined. Whereas ts3 was found to rescue MuSV at340C,
a shift to390C
immediately blocked releaseofboth viruses. A subsequent shift from39 to340C
again resultedintheacti-VOL. 24, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
230 YUEN AND WONG
nomycin D- and cycloheximide-insensitive
release of both ts3 and MuSVduring the first hour aftershiftdown. Recently, sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (J. A. Ptak and P. K. Y. Wong, manuscript in
preparation) indicated that the mobilityof the glycoproteinfraction of ts3 virions grown atthe
permissive temperature isdifferent fromthat of wild-type MuLV. In addition, the mobility of viral proteins precipitated by anti-MuLV gp7l from ts3-infected cells grown atthe nonpermis-sive temperature was also different from that precipitated from wild-type MuLV-infected
cells. Furthermore, the proteins precipitated
fromts3-infectedcells grown at the
nonpermis-sive temperature showed an increase in the amount of the 85,000-molecular-weight
glyco-proteinsand a concomitantdecreaseingp69/71.
All the above findings indicate that the de-fect of ts3 mayresideineitherthe processingor
conformation of the viral envelope
compo-nents(s). Ithasbeensuggestedthat the
assem-blyofenvelopedviruses isregulated by
protein-protein interactions (4). The conversion of a membrane area into a budding patch may be
brought aboutbyinsertionofglycoprotein
mol-ecules into the lipidbilayer (1). Itwasfurther
suggested that the shape and rigidity of the
viral envelope is maintainedby aviral struc-tural componentlaid downinnumerical
corre-spondencetotheenvelope
glycoprotein(s).
It isconceivable that other structural proteins,
in-cludingtheribonucleoprotein,maybesimilarly
assembled in their correct order. We suggest
that if the glycoprotein(s) of ts3 is unable to
assume its correctconformationatthe nonper-missive temperature, thenthe assemblyof the rest of theviral components may behampered
by either an inefficiency or inaccuracy in the protein-protein interactions. However, once
permissive conditions are restored, the
defec-tive conformation of the glycoprotein is
cor-rected andnormalassemblycanbe resumed.
Among the late ts mutants ofRauscher
leu-kemiavirusisolatedby Stephenson and
Aaron-son (6), ts24 exhibits characteristics similar tothose of ts3. Recentmicroscopic studies (11) suggest that ts24is defective either inthe late
stages of assembly of the particles or in the release of the virus particles from the cell sur-face. Ts24 resemblests3 inthatthereare more
cell-associated virions at the nonpermissive than at the permissive temperature and that virions are rapidly released on temperature
shiftdown. However,unlike ts3, there appearto
be far fewer cell-associated virions in
ts24-in-fectedcellsgrown for6daysatthe nonpermis-sivetemperature (4 to 5virions/cell section)as
compared with ts3-infected cells grown for48h
at 39°C (22 virions/cell section). Furthermore,
allthebudding ts24 virions observed in infected cells grown atthe nonpermissive temperature werealmost completely assembled, normal vir-ions. By contrast, both normal particles and multiploids in all stages of assembly were ob-served ints3-infected cells grown at the nonper-missive temperature.
Itis possible that although ts3 and ts24 have certaincharacteristics in common, they are in fact temperature sensitive in different func-tions. Alternatively, they may be defective in the same function to varying degrees. An ap-proach in resolving this possibility may be achievedby means ofcomplementationand re-combination tests between thesemutants.
ACKNOWLEDGMENTS
Wewish to thank H. C. Chen and L. Jones for their skillfulassistance and M.Weber for critically reviewing the manuscript.
This investigation was supported by Public Health Serv-iceresearch grant CA-17695 awarded by the National Can-cerInstitute to P.K.Y.W.
LITERATURE CITED
1. Blough, H. A., and J. M. Tiffany. 1975. Theoretical aspectsof structure andassembly of viral envelopes. Curr.Top.Microbiol.Immunol. 70:1-30.
2. Friis, R. R., H. Ogura, H. Gelderblom, and M. S. Halpern.1976.Thedefectivematurationof viral prog-eny with a temperature-sensitive mutant of avian sarcomavirus.Virology 73:259-272.
3. Hunter, E., M. J. Hayman, R. W. Rongey, and P. K. Vogt. 1976. An avian sarcoma virus mutant that is temperature sensitive forvirionassembly. Virology 69:35-49.
4. Klenk, H.-D. 1974. Viral envelopes and their relation-ship to cellularmembranes. Curr. Top. Microbiol. Immunol. 68:29-58.
5. Sato, T.,C.Friend,and E. de Harven. 1971. Ultrastruc-tural changes in Friend erythroleukemia cells treated with dimethyl sulfoxide. Cancer Res.
31:1402-1417.
6. Stephenson,J.R.,andS.A.Aaronson. 1973. Character-ization oftemperature-sensitive mutants of murine leukemiavirus. Virology 54:53-59.
7. Wong,P.K.Y.,and J. A. McCarter.1974.Studies oftwo
temperature sensitive mutants of Moloney murine leukemiavirus. Virology 58:396-408.
8. Wong, P. K. Y., and R. MacLeod. 1975. Studies on the buddingprocessof a temperature-sensitive mutant of murineleukemiaviruswith a scanning electron mi-croscope.J.Virol.16:434-442.
9. Wong, P. K. Y., L. J. Russ, and J. A. McCarter. 1973. Rapid, selective procedure for isolation of sponta-neoustemperature-sensitive mutants ofMoloney mu-rineleukemiavirus.Virology 51:424-431.
10. Wong, P. K. Y., P. H. Yuen, R. MacLeod, E. H. Chang,M. W.Myers,and R. M.Friedman.1977.The effectofinterferonondenovo infectionofMoloney leukemia virus. Cell 10:245-252.
11. Yeger, H., V. I. Kalnins, and J. R. Stephenson. 1976.
Electron microscopyofmammalianType-CRNA vi-ruses: useofconditional lethalmutants instudies of virion muturation and assembly. Virology
74:459-469.
12. Yuen, P. H., and P. K. Y. Wong. 1977. Amorphologic studyontheultrastructure andassemblyofmurine
leukemia virus using a temperature-sensitive mu-tantrestrictedinassembly. Virology80:260-274.
on November 10, 2019 by guest
http://jvi.asm.org/