Efficient Translation of Epstein-Barr Virus (EBV) DNA
Polymerase Contributes to the Enhanced Lytic Replication
Phenotype of M81 EBV
Trenton Mel Church,aDinesh Verma,aJacob Thompson,b Sankar Swaminathana,b
aDivision of Infectious Diseases, Department of Medicine, University of Utah School of Medicine, Salt Lake City,
Utah, USA
bDepartment of Pathology, University of Utah School of Medicine, Salt Lake City, Utah, USA
ABSTRACT Epstein-Barr virus (EBV) is linked to the development of both lymphoid and epithelial malignancies worldwide. The M81 strain of EBV, isolated from a Chi-nese patient with nasopharyngeal carcinoma (NPC), demonstrates spontaneous lytic replication and high-titer virus production in comparison to the prototype B95-8 EBV strain. Genetic comparisons of M81 and B95-8 EBVs were previously been performed in order to determine if the hyperlytic property of M81 is associated with sequence differences in essential lytic genes. EBV SM is an RNA-binding protein expressed dur-ing early lytic replication that is essential for virus production. We compared the functions of M81 SM and B95-8 SM and demonstrate that polymorphisms in SM do not contribute to the lytic phenotype of M81 EBV. However, the expression level of the EBV DNA polymerase protein was much higher in M81- than in B95-8-infected cells. The relative deficiency in the expression of B95-8 DNA polymerase was related to the B95-8 genome deletion, which truncates the BALF5 3= untranslated region (UTR). Similarly, the insertion of bacmid DNA into the widely used recombinant B95-8 bacmid creates an inefficient BALF5 3= UTR. We further showed that the while SM is required for and facilitates the efficient expression of both M81 and B95-8 mRNAs regardless of the 3= UTR, the BALF5 3=UTR sequence is important for BALF5 protein translation. These data indicate that the enhanced lytic replication and virus production of M81 compared to those of B95-8 are partly due to the ro-bust translation of EBV DNA polymerase required for viral DNA replication due to a more efficient BALF5 3=UTR in M81.
IMPORTANCE Epstein-Barr virus (EBV) infects more than 90% of the human popula-tion, but the incidence of EBV-associated tumors varies greatly in different parts of the world. Thus, understanding the connection between genetic polymorphisms from patient isolates of EBV, gene expression phenotypes, and disease is important and may help in developing antiviral therapy. This study examines potential causes of the enhanced lytic replicative properties of M81 EBV isolated from a nasopharyn-geal carcinoma (NPC) patient and provides new evidence for the role of the BALF5 gene 3= UTR sequence in DNA polymerase protein expression during lytic replica-tion. Variation in the gene structure of the DNA polymerase gene may therefore contribute to lytic virus reactivation and pathogenesis.
KEYWORDS DNA polymerase, DNA replication, EBV, Epstein-Barr virus, SM protein, herpesvirus, posttranscriptional RNA-binding proteins, posttranscriptional control mechanisms, transcriptional regulation, translational control
E
pstein-Barr virus (EBV) is a human herpesvirus that infects and establishes persistent latent infection in more than 95% of the human population worldwide. EBV infection is associated with several lymphomas and epithelial carcinomas, includingReceived12 October 2017Accepted17 December 2017
Accepted manuscript posted online20 December 2017
CitationChurch TM, Verma D, Thompson J, Swaminathan S. 2018. Efficient translation of Epstein-Barr virus (EBV) DNA polymerase contributes to the enhanced lytic replication phenotype of M81 EBV. J Virol 92:e01794-17. https://doi.org/10.1128/JVI.01794-17. EditorRichard M. Longnecker, Northwestern University
Copyright© 2018 American Society for Microbiology.All Rights Reserved. Address correspondence to Sankar Swaminathan, [email protected].
crossm
on November 6, 2019 by guest
http://jvi.asm.org/
Burkitt lymphoma, posttransplant lymphoproliferative syndromes, gastric carcinoma, and nasopharyngeal carcinoma (NPC) (1). Distinct geographic variations in the inci-dences of these malignancies have been observed and have been attributed to environmental, genetic, and viral factors (2–4). The incidence of EBV-associated NPC is highly geographically variable and is particularly prevalent in southern China. Strain polymorphisms in the LMP-1 gene that are regionally overrepresented in cases of NPC have been identified, but whether they actually represent pathogenic determinants is controversial.
In 1970, a B cell lymphoblastoid cell line (LCL) named HKLY-28 was established from NPC tissue from a Hong Kong Chinese patient (5). Virus from this line was passaged to B cells from the common marmoset,Callithrix jacchus, and two LCLs (M81 and M82) were generated (6). EBV virions from M81 were then used to generate LCLs from human cord blood and adult lymphocytes, and the properties of these LCLs were compared to those of LCLs made from B95-8, the prototype EBV strain (7). M81 LCLs spontaneously expressed early lytic antigens at a much higher rate than did B95-8 LCLs. Furthermore, M81 LCLs spontaneously expressed viral capsid antigen (VCA) and produced infectious virions from cord blood LCLs, which was never observed with the B95-8-derived cord blood LCLs. These enhanced lytic properties of the M81 EBV strain were confirmed in recent studies of a bacmid made from M81 (8). In addition, M81 was demonstrated to possess enhanced epithelial cell tropism compared to B95-8. The enhanced lytic properties of M81 were shown to be partly attributable to the functional properties of the M81 Zta transactivator protein, which activates the entry of latent EBV into the lytic phase of replication and is required for lytic DNA replication (9). Those studies were done by comparing the lytic replication of bacmid mutants in which the Zta gene was deleted when rescued by either M81 or B95-8 Zta. Although M81 Zta was more potent in its transactivation function than B95-8 Zta and led to increased virion production, it was not capable of fully reconstituting the hyperlytic M81 phenotype, suggesting that other M81 genes were also involved.
M81 was also shown to cluster with other Chinese NPC-derived EBV strains based on taxonomic comparison with several Burkitt lymphoma and NPC strains of different geographic origins (8). Those findings raised the possibility that the epitheliotropic and lytic properties of NPC strains may correlate with their oncogenic properties. Although EBV infection in NPC tumors is latent, serologic reactivity to lytic antigens, particularly IgA responses, is frequently evident in patients with NPC and serves as an indicator of recurrence after therapy (10–12). In order to investigate other EBV gene products that may play a role in the hyperlytic phenotype of M81, we examined the functional properties of the M81 SM gene, which is essential for lytic replication, infectious virus production, and the efficient expression of many late EBV genes (13–17). The EBV SM protein is a regulatory protein expressed by EBV during the early lytic phase of replication. SM acts as a posttranscriptional regulator to specifically enhance the accumulation of several EBV RNA transcripts that are required for infectious virion production (14, 17). EBV SM and its homologs in other human herpesviruses have been shown to play important roles in RNA biology, including the processes of RNA stabili-zation, splicing, polyadenylation, export, and translation (18–20). A comparison of the lytic replicative properties of B95-8 and M81 viruses and their SM functions was performed by using an SM knockout (SMKO) bacmid. In addition, these studies revealed that M81 DNA replication was more robust than that of B95-8. Further investigation demonstrated that the M81 DNA polymerase was more efficiently expressed. The basis of the enhanced expression of the M81 DNA polymerase was studied and determined to be correlated with genetic determinants that facilitate the translation of the M81 DNA polymerase mRNA.
RESULTS
Comparison of lytic replication between M81 and B95-8 EBV.In order to confirm the greater lytic replication of M81 EBV than of B95-8 EBV, we compared the levels of production of infectious virions in HEK293 cells infected with either M81 or B95-8 EBV.
on November 6, 2019 by guest
http://jvi.asm.org/
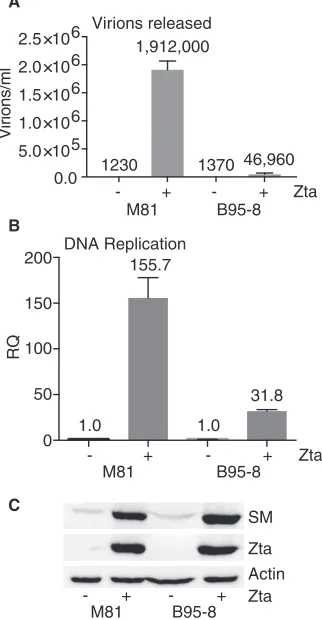
Each cell line was transfected with the lytic transactivator Zta (Z; BZLF1) to induce EBV lytic replication in parallel, and cell supernatants were harvested at 5 days postrepli-cation. The release of infectious viral particles was measured by infecting Raji cells with the supernatant obtained from either M81 EBV- or B95-8 EBV-infected HEK293 cells. Numbers of infected Raji cells expressing green fluorescent protein (GFP) due to EBV infection were quantitated by flow cytometry at 72 h postinfection. As shown in Fig. 1A, infectious virus production was observed in both M81 EBV- and B95-8 EBV-infected cells after the induction of lytic replication by Zta transfection. However, M81 EBV was more lytic and produced approximately 40-fold more infectious virion particles than B95-8 EBV. This result corresponds well with the previously reported findings that M81 EBV is more capable of producing infectious virions (6–8).
In order to determine if the increased lytic capability of M81 EBV was due to increased EBV DNA replication compared to that of B95-8 EBV, DNA replication was measured in HEK293 cells infected with M81 or B95-8 EBV. Lytic replication in both cell lines was induced by transfection with Zta plasmid DNA, and EBV genome amplification was measured in total cellular DNA purified at 60 h posttransfection. Relative quantities of EBV DNA were measured by quantitative PCR (qPCR) of M81 and B95-8 DNAs in parallel, as described previously (17, 21). As shown in Fig. 1B, there were approximately 156-fold and 32-fold increases in the EBV intracellular DNA levels after the induction of
A
B
C
- + - +
M81 B95-8
DNA Replication
1.0
155.7
1.0 31.8
RQ
- + - +
M81 B95-8
SM
Zta Actin Zta
Zta 200
150
100
50
0
- + - +
M81 B95-8
Zta 0.0
5.0 105 1.0 106 1.5 106 2.0 106 2.5 106
V
irions/
ml
1230
1,912,000
1370 46,960 Virions released
FIG 1Comparison of virion production by M81 EBV and B95-8 EBV. HEK293 cells stably infected with
either M81 EBV or B95-8 EBV strains were transfected with either the empty vector (⫺) or Zta plasmid
DNA (⫹) to induce EBV lytic replication. (A) Quantification of virus production. Supernatants were
harvested 5 days after the induction of replication, and virus-containing supernatants were used to infect Raji cells. The number of GFP-positive Raji cells, representing infectious viral particles, was counted by flow cytometry. (B) Quantification of EBV DNA replication by qPCR. Cells were harvested, and DNA was isolated 3 days after the induction of replication. Numbers of EBV genomes were measured by qPCR. RQ, relative quantity. (C) Western blotting was performed on cell lysates harvested 2 days after replication induction to measure Zta and SM protein expression levels. Western blots were stripped and reprobed with actin antibody as a loading control. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:3.585.124.285.77.387.2]replication in M81- and B95-8-infected cells, respectively. These results suggest that the greater efficiency of DNA replication in M81-infected cells may be one of the reasons for the overall increased production of infectious virus by M81 EBV.
It was previously reported that polymorphisms in the BZLF1 gene of M81 contrib-uted to the hyperlytic phenotype (8). In order to confirm that the overall levels of Zta were comparable in the two cell lines under these experimental conditions, we compared Zta protein expression levels in both cell lines by Western blotting of cell lysates harvested 48 h after lytic induction. Immunoblotting demonstrated that protein levels of Zta were similar (Fig. 1C) in M81- and B95-8-infected cells. Previous studies suggested that although M81 Zta may be more efficient, it did not fully explain the lytic phenotype of M81 and that other lytic genes may contribute (8). Since the SM protein is essential for lytic EBV production and specifically increases the expression levels of several other essential EBV lytic genes (14, 17), we compared the levels of SM protein expression by M81 and B95-8. The levels of SM protein expression were similar for M81 and B95-8, indicating that the level of the SM protein did not differ between the two strains and could not explain the difference in lytic phenotypes.
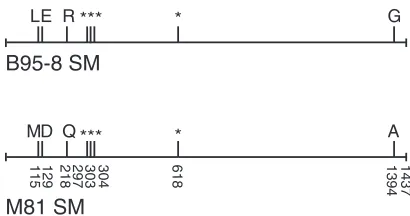
Polymorphism in EBV SM protein does not contribute to enhanced lytic repli-cation of M81 EBV.Since a higher level of expression of the SM protein by M81 was not observed, we considered the possibility that polymorphisms in the SM protein may result in a more active or functional M81 SM protein, which could contribute to the enhanced lytic phenotype of M81. Therefore, we compared the EBV SM sequences of the M81 and B95-8 genomes, and inspection revealed that there are eight nucleotide differences, four of which lead to amino acid changes in the predicted SM protein sequences of the two EBV strains (Fig. 2).
In order to determine if the four amino acid changes in the SM sequence are related to the lytic capabilities of M81 EBV versus B95-8 EBV, we cloned the SM open reading frames (ORFs) from the two different SM genes exactly in the same manner, adding an N-terminal FLAG tag in the mammalian expression vector pCDNA3. HEK293 cells infected with SM-null B95-8 EBV (EBVBMLF1-KO) (SMKO) (16, 17) were used to compare the functional abilities of the two different SM genes from M81 and B95-8.
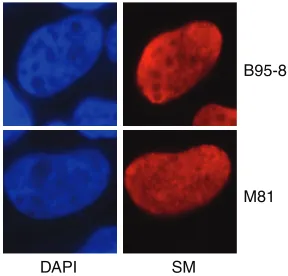
We first asked if there were any differences in the nuclear localization of SM due to sequence differences found between M81 SM and B95-8 SM. SMKO cells were trans-fected with Zta plasmid DNA to induce lytic replication with either B95-8 or M81 SM, as described above. Forty-eight hours after transfection, cells were fixed and stained with SM antibody to assess SM localization (red) and with DAPI (4= ,6-diamidino-2-phenylindole) nuclear stain (blue), as shown in Fig. 3. SM proteins from both virus strains exhibited the typical diffuse, speckled nuclear staining, with some nucleolar sparing. No differences in the localizations of the SM protein pattern were apparent between B95-8 SM and M81 SM during lytic replication.
*** *
*
B95-8 SM
M81 SM
129 297
115 218 303 304 618 1394 1437
G LE R
***
MD Q A
FIG 2Comparison of M81 SM and B95-8 SM sequences. A schematic diagram shows the alignment of SM sequences between M81 EBV and B95-8 EBV. The two DNA sequences show a total of eight nucleotide differences, and four of these nucleotide changes (positions 297, 303, 304, and 618) are conservative,
leading to no change in the amino acid sequence (marked by *). The remaining four nucleotide
polymorphisms (positions 115, 129, 218, and 1394) were nonconservative, leading to four amino acid differences between the two sequences (noted by letter references to amino acids).
on November 6, 2019 by guest
http://jvi.asm.org/
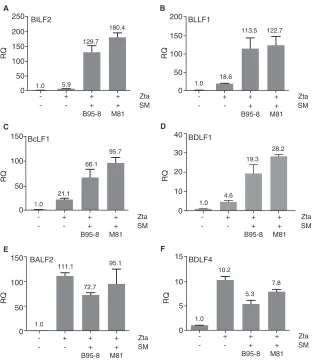
[image:4.585.101.306.70.178.2]SM is essential for infectious virus production by EBV because it specifically en-hances the accumulation of a subset of important EBV late mRNAs (17). We therefore compared the functions of both M81 SM and B95-8 SM in their abilities to enhance the accumulation of these specific SM-dependent EBV mRNAs. SMKO cells were transfected with the Zta plasmid alone to induce lytic replication or with the either M81 or B95-8 SM. Forty-eight hours after transfection, cells were harvested, and SM-dependent (BILF2, gp350, BcLF1, or BDLF1) and SM-independent (BALF2 and BDLF4) RNAs were tested by reverse transcription-qPCR (RT-qPCR), as described previously (17). As ex-pected, SM rescue led to a significant increase in the steady-state mRNA levels of SM-dependent EBV genes, and there was no significant functional difference observed for gene regulation by either M81 or B95-8 SM (Fig. 4A to F). Neither SM protein led to significant changes in the levels of SM-independent genes or led to a slight decrease, as previously described (17). These data indicate that the increased replicative prop-erties of M81 are not due to its SM protein being more active in transcriptional or posttranscriptional effects on its known RNA targets.
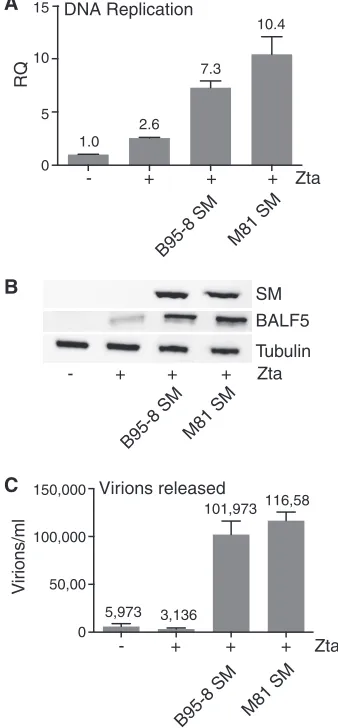
We have previously shown that SM leads to increased EBV DNA replication despite having no significant effects on the mRNA levels of the EBV genes encoding proteins directly involved in EBV lytic DNA replication (17). To ask if the sequence differences in M81 EBV SM could potentially be related to the increased EBV DNA replication observed in the wild-type (WT) virus comparison (Fig. 1B), we compared the effects of M81 SM and B95-8 SM on DNA replication when rescuing SMKO EBV. We used SMKO–B95-8-infected HEK293 cells and transfected them with the Zta plasmid to induce lytic replication with or without each SM plasmid. Sixty hours after transfection, the cells were harvested, total DNA was isolated, and relative EBV DNA copy numbers were determined by qPCR (Fig. 5A). The results indicate that both M81 SM and B95-8 SM enhanced DNA replication. Although the level of DNA replication was slightly higher in the presence of M81 SM-transfected samples than in the presence of B95-8 SM-transfected samples, it did not explain the 5-fold-higher level of replication seen in M81-infected cells (Fig. 1B). Since M81 cells exhibited greater DNA replication, we also compared the expression levels of the polymerase protein in cells rescued with M81 or B95-8 SM. SMKO cells were transfected with Zta and either M81 or B95-8 SM. Forty-eight hours after transfection, cells were harvested, and lysates were immunoblotted to measure SM and DNA polymerase (BALF5) protein expression levels. As shown in Fig. 5B, similar levels of BALF5 protein expression were seen in M81 SM- and B95-8 SM-transfected cells. Although BALF5 protein expression was clearly increased by SM, this effect was similar in the presence of either M81 or B95-8 SM, showing that SM sequence differences in the M81 strain are not responsible for the increased DNA polymerase protein levels or DNA replication observed with M81 EBV. We confirmed that SM expression levels were similar in both cases.
DAPI
M81 B95-8
SM
FIG 3Nuclear localization patterns of M81 SM and B95-8 SM during lytic replication. HEK293 cells stably infected with SMKO EBV were transfected with Zta and either M81 EBV SM or B95-8 EBV SM. Cells were fixed 48 h after transfection and stained with SM antibody (red). Nuclei were stained with DAPI (blue). Cells were visualized by fluorescence microscopy.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:5.585.132.276.72.210.2]Finally, we compared M81 SM and B95-8 SM for their abilities to rescue infectious virus production from SMKO EBV-infected cells. The B95-8 SMKO-infected HEK293 cell line was transfected with Zta plasmid DNA to induce EBV lytic replication along with pCDNA3-FLAG constructs containing either M81 SM or B85-8 SM. Numbers of infectious viral particles were measured in cell supernatants harvested at 5 days postreplication. Numbers of released infectious viral particles were measured by infecting Raji cells with the supernatants, and numbers of GFP-expressing infected Raji cells were quantitated by flow cytometry at 72 h postinfection (Fig. 5C). As expected, SMKO EBV did not release infectious viral particles without SM despite the induction of replication, whereas abundant virus production was observed in M81 SM- and B95-8 SM-rescued samples. As shown in Fig. 5C, there was no difference in virus yields; approximately 1⫻ 105 GFP-inducing units/ml were released from both M81 SM- and B95-8 SM-rescued samples. These results suggest that the differences observed in the SM ORF sequences are not functionally significant and are not related to the lytic properties of M81. These results indicated that both M81 SM and B95-8 SM functionally behave similarly and that the highly lytic properties of M81 EBV were not due to functional differences due to sequence polymorphisms observed between B95-8 SM and M81 SM.
Role of EBV DNA polymerase in M81 lytic replication. The replication of EBV genomes and the production of infectious viral particles require six viral genes that act
E RQ BALF2 150 100 50 0 1.0 111.1 72.7 95.1 C RQ BcLF1 150 100 50 0 1.0 21.1 66.1 95.7 RQ BDLF4 0 5 10 15 1.0 10.2 5.3 7.8 A 129.7 180.4 BILF2 250 200 150 RQ 1.0 5.9 100 50 0
- + + + Zta
- - + + SM
B95-8 M81 B 113.5 122.7 BLLF1 200 150 RQ 1.0 18.6 100 50 0
- + + + Zta
- - + + SM
B95-8 M81 F D RQ BDLF1 1.0 4.6 19.3 28.2 40 30 20 10 0
- + + + Zta
- - + + SM
B95-8 M81
- + + + Zta
- - + + SM
B95-8 M81
- + + + Zta
- - + + SM
B95-8 M81
- + + + Zta
- - + + SM
B95-8 M81
FIG 4Effect of B95-8 SM versus M81 SM on EBV RNA accumulation during lytic replication. SMKO HEK293
cells were transfected with the empty vector (⫺), Zta (⫹), or Zta with either B95-8 SM or M81 SM, as
shown. Transfection of Zta induces EBV replication, which is fully rescued by cotransfection of SM. RNA was harvested 48 h after transfection. Expression levels of SM-responsive (BILF2, BLLF1, BcLF1, and BDLF1) (A to D) and SM-independent (BALF2 and BDLF4) (E and F) RNAs were measured by RT-qPCR. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
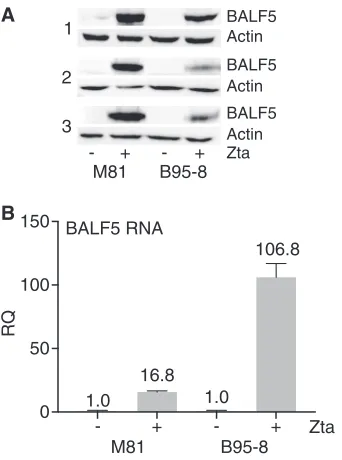
[image:6.585.50.362.69.429.2]as replication factors (22, 23). One of these, BALF5, encodes the EBV DNA polymerase and is essential for lytic EBV DNA replication. Previous studies have shown that the EBV DNA polymerase 3=untranslated region (UTR) contains a noncanonical poly(A) signal, UAUAAA, and that SM enhances the processing of this transcript (24, 25). It is also known that B95-8 has a deletion in this region of the genome so that the polyadenyl-ation signal used by BALF5 is a serendipitous alternate signal (24, 26). Thus, it was possible that M81 EBV expresses larger amounts of either the BALF5 protein or mRNA than B95-8. We therefore compared BALF5 protein and RNA expression levels in M81-and B95-8-infected cells. We used HEK293 cells stably infected with either M81 or B95-8 EBV, transfected them with the lytictrans-activator (Zta) to induce EBV lytic replication in parallel, and harvested them 48 h after transfection. BALF5 protein and mRNA levels were measured by Western blotting and RT-PCR, respectively. As shown in Fig. 6A, the level of the BALF5 protein found in M81 EBV-infected cells was consistently and significantly higher than that in B95-8 EBV-infected cells, suggesting that the enhanced lytic characteristics of M81 EBV may be related to increased BALF5 protein expression.
A
B
C
Virions released- + + +
M81 SM B95-8 SM
Zta
5,973 3,136
101,973116,58
150,000
100,000
50,00
0
V
ir
ions
/ml
- + + +
M81 SM B95-8 SM
Zta DNA Replication
1.0 2.6
7.3 10.4 15
10
5
0
RQ
- + + +
M81 SM B95-8 SM
Zta SM BALF5
Tubulin
FIG 5Comparison of SM functions in lytic EBV replication. SMKO HEK293 cells were transfected with the
empty vector (⫺), Zta (⫹), or Zta with either B95-8 SM or M81 SM. (A) Quantification of EBV DNA
replication by qPCR. Cells were harvested, total DNA was isolated 3 days after the induction of replication, and numbers of EBV genomes were measured by qPCR. (B) Western blotting was performed on cell lysates harvested 2 days after the induction of replication to measure SM, Zta, and BALF5 protein expression levels. Western blots were stripped and reprobed for tubulin as a loading control. (C) Quantification of virus production. Cell supernatants were harvested 5 days after lytic induction, and virus-containing supernatants were used to infect Raji cells. The number of GFP-positive Raji cells, representing infectious viral particles, was counted by flow cytometry. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.120.289.77.441.2]Somewhat surprisingly, the BALF5 mRNA level in B95-8 EBV-infected cells was much higher than that in M81-infected cells (Fig. 6B), suggesting that B95-8 BALF5 mRNA may be translated less efficiently.
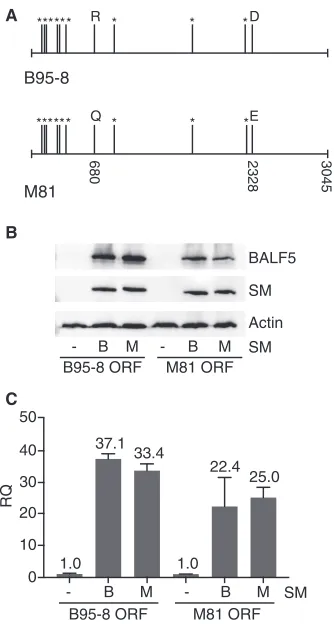
Increased BALF5 protein expression in M81 EBV is not due to polymorphism in the BALF5 coding sequence.One explanation for the difference in the efficiencies of protein expression from M81 versus B95-8 BALF5 mRNAs is the known differences in translation that are derived from differences in codon usage. Optimal codon usage is correlated with enhanced translation elongation (27). Examination of the M81 and B95-8 genomes and inspection of the BALF5 ORFs revealed that there are 11 nucleotide differences with 9 conservative codon differences in BALF5, as shown in Fig. 7A. These polymorphisms raised the possibility that M81 BALF5 mRNA was translated more efficiently. It was also possible that there were strain-specific differences in the ability of M81 SM to enhance M81 BALF5 translation, since SM has been reported to facilitate the translation of other EBV mRNAs (28). We therefore compared the amounts of RNA and protein produced from each BALF5 coding domain cloned into an expression vector and in the presence of SM from either strain. The pCDNA3 expression vector was used, which contains a cytomegalovirus (CMV) promoter and a bovine growth hormone polyadenylation signal. This experimental design allowed us to isolate and compare the efficiencies of translation of each BALF5 ORF independent of noncoding regulatory regions present upstream and downstream of the gene in the different viral genomes. The M81 and B95-8 BALF5 expression plasmids were transfected into HEK293 cells with either an empty vector, M81 SM, or B95-8 SM. Forty-eight hours after transfection, cells were harvested, and BALF5 protein levels were measured by Western blotting. As shown in Fig. 7B, BALF5 protein expression was highly SM dependent, but BALF5 was equally well expressed in the presence of either M81 SM or B95-8 SM.
To compare the BALF5 mRNA abundances from M81 and B95-8 and their depen-dence on SM, we measured BALF5 mRNA levels in the cells in which BALF5 protein expression levels were compared, as described above (Fig. 7C). This experiment showed
B A
- + - +
M81 B95-8
Zta BALF5 RNA
1.0 16.8
106.8
1.0
RQ
150
100
50
0
- + - +
M81 B95-8
Zta BALF5
BALF5
BALF5 Actin Actin Actin
1
2
3
FIG 6Comparison of BALF5 gene expression levels in M81- and B95-8-infected cells during lytic replication. HEK293 cells stably infected with either the M81 EBV or B95-8 EBV strain were induced to
permit lytic replication by transfection of a Zta plasmid (⫹) or were transfected with the empty vector
(⫺). (A) Expression of endogenous BALF5 proteins was analyzed by Western blotting of cell lysates
harvested 48 h after transfection. Results from three independent experiments are shown. Blots were stripped and reprobed for actin as a loading control. (B) RT-qPCR was performed to measure BALF5 RNA levels in M81 EBV- or B95-8 EBV-infected cells at 2 days posttransfection. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.122.293.71.301.2]that BALF5 RNAs from both strains were poorly expressed in the absence of SM and were upregulated by SM. However, M81 SM and B95-8 SM were equally efficient in enhancing BALF5 RNA expression. These two sets of experiments therefore indicate that the more robust BALF5 protein expression in the M81 viral strain is not due to either increased RNA accumulation or more efficient translation due to differential codon usage between M81 BALF5 and B95-8 BALF5.
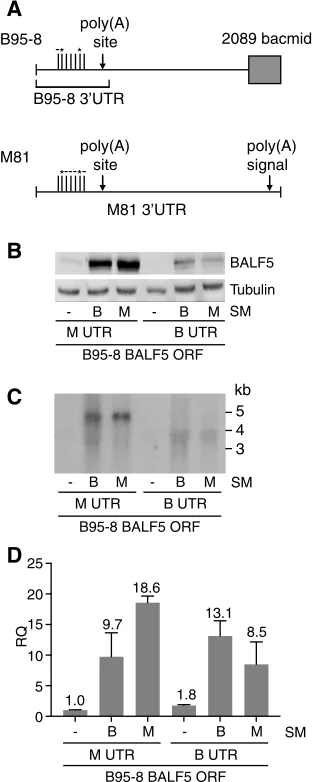
The EBV M81 BALF5 3=UTR sequence is important for SM-dependent BALF5 gene expression and lytic replication. Previous studies reported that B95-8 EBV expresses an RNA transcript of 3.7 kb, but other EBV strains such as Raji and P3HR1 produce an additional major transcript of 5.03 kb (26). The longer transcript is due to the usage of a preferred noncanonical UAUAAA signal also present in M81. The shorter transcript is derived by the use of an alternative upstream polyadenylation site, which, due to the deletion of the downstream polyadenylation site in the B95-8 genome, is the sole BALF5 transcript in B95-8 (26). In the p2089 bacmid, which is the strain used in this study, the bacmid sequence was inserted into the B95-8 deletion, as shown in Fig. 8A (29). Close examination of the 3=UTR BALF5 nucleotide sequence (Fig. 8A) revealed seven nucleotide differences between B95-8 and M81 (comprised of one deletion in B95-8, four deletions in M81, and two nucleotides that are different). In order to
A
B
R * D
B95-8
C
RQ
50
40
30
0 20
10 1.0
37.1 33.4
1.0 22.4
25.0
B M B M
B95-8 ORF
SM
-M81 ORF
B M B M
B95-8 ORF
SM
-M81 ORF SM BALF5
Actin
* * * * * *
* *
Q E
M81
680 2328 3045
* * * * *
* * * *
FIG 7Enhanced M81 BALF5 protein expression is not due to polymorphisms in M81 and B95-8 BALF5 coding sequences. (A) Schematic diagram showing an alignment of M81 and B95-8 EBV BALF5 coding sequences. The two DNA sequences contain a total of 11 nucleotide differences, and 9 of these nucleotide differences (positions 105, 135, 156, 276, 300, 369, 898, 2205, and 2211) are conservative,
leading to no difference in amino acid sequences (denoted by*). Two polymorphisms at nucleotide
positions 680 and 2328 lead to a difference in the amino acid (noted by letter references to the amino acid). (B) Expression levels of BALF5 proteins determined by Western blotting. HEK293 cells were transfected with either a B95-8 or M81 BALF5 coding sequence expression vector along with either B95-8 SM or M81 SM plasmids or empty vector DNA. Cell lysates were harvested 48 h after transfection and analyzed for BALF5 protein expression with anti-BALF5 antibody. Actin was used as a loading control. (C) RNA was harvested from cells at the same time as protein was harvested, and RT-qPCR was performed to measure the expression level of BALF5 RNA. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.123.290.70.382.2]examine if sequence differences in BALF5 3= UTRs are associated with BALF5 gene regulation, we cloned BALF5 3=UTR sequences from M81 and p2089 bacmids into the B95-8 BALF5 expression vector after deleting the bovine growth hormone polyadenyl-ation signal in pCDNA3. The effects of each 3=UTR sequence on B95-8 BALF5 protein expression were then compared in HEK293 cells by transfecting a B95-8 BALF5 ORF containing either the p2089 3=UTR or the M81 3=UTR alone or in combination with each SM (M81 or B95-8), as described above. Forty-eight hours after transfection, cells
A
B
C
B M B M
B UTR
SM
-RQ
25
20
15
10
5
0
M UTR M81 3’UTR B95-8 3’UTR
poly(A) site
poly(A) signal poly(A)
site 2089 bacmid
*---*-* * -B95-8
M81
B M B M
B95-8 BALF5 ORF B UTR
SM
-M UTR
BALF5
Tubulin
1.0 9.7
18.6
1.8 13.1
8.5
D
B M B M
B UTR
-M UTR
SM 5
3 4 kb
B95-8 BALF5 ORF
B95-8 BALF5 ORF
FIG 8Effect of the BALF5 3=UTR on protein expression. (A) Diagram showing an alignment of the
corresponding regions of the B95-8 and M81 bacmid BALF5 3=UTR sequences. In the B95-8 bacmid, GFP
has been inserted into the naturally occurring deletion in B95-8. Two nucleotide differences between the
BALF5 3=UTRs of M81 and B95-8 are shown with asterisks (T to C at B95-8 nucleotide 153012 and G to
A at B95-8 nucleotide 152962). One gap in B95-8 (nucleotide 153081) and two gaps in M81 (B95-8 nucleotides 152974 to 152976 and nucleotide 152946) are shown with dashes. The proximal cleavage and polyadenylation sites at nucleotide 153215 (present in both genomes) and the preferred distal site
in M81 are shown with arrows. (B) Western blotting comparing the effects of B95-8 and M81 3=UTRs on
BALF5 protein expression. The B95-8 BALF5 coding sequence was cloned into the pCDNA3 expression
vector with either the B95-8 3=UTR or the M81 3=UTR sequence downstream of the BALF5 ORF. Each
construct was transfected into HEK293 cells along with either B95-8 SM (B), M81 SM (M), or the empty
vector (⫺). BALF5 protein expression was analyzed by Western blotting. Blots were stripped and
reprobed with tubulin antibody as a loading control. (C) Northern blotting of BALF5 mRNAs produced
with either B95-8 or M81 3=UTRs. RNAs from transfected cells in panel B were isolated and analyzed by
Northern blotting. RNA marker sizes are shown at the right in kilobases. (D) RT-qPCR was performed with RNAs harvested from the samples in panel C, as described above. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:10.585.128.284.74.465.2]were harvested, and BALF5 and SM protein expression levels were measured by Western blotting. As shown in Fig. 8B, BALF5 protein was expressed more efficiently from an mRNA with the M81 3=UTR than with the B95-8 bacmid 3=UTR. While both SM proteins enhance BALF5 expression, they are equally effective in this regard, with the 3=UTR determining the level of expression.
We also measured BALF5 RNA levels 48 h after transfection in the above-described experiments to see how the 3=UTRs might affect BALF5 RNA regulation by SM. Again, BALF5 RNA was expressed poorly in the absence of SM and was upregulated by either SM (Fig. 8C and D). Interestingly, BALF5 mRNA with the B95-8 3=UTR was upregulated more by B95-8 SM than by M81 SM, suggesting that there may be complex interactions between the 3=UTR and SM that affect RNA processing and stability.
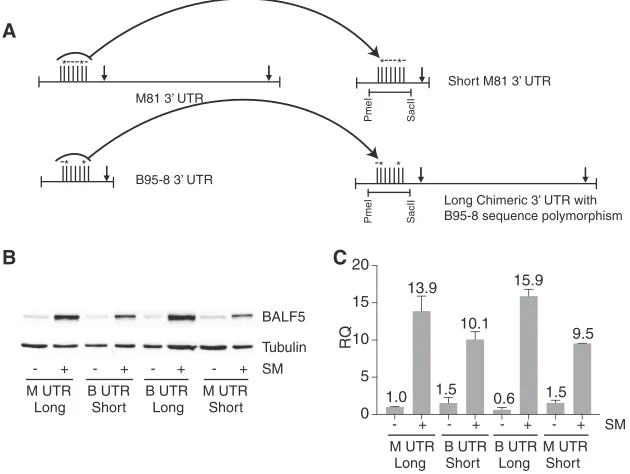
We further asked whether sequence polymorphisms observed between M81 and B95-8 BALF5 3= UTRs or the length of the BALF5 3= UTR was important for BALF5 translation, as shown in Fig. 8. We constructed additional BALF5 genes in which the sequence polymorphisms between M81 and B95 3=UTRs were exchanged, as shown in Fig. 9A. Briefly, we constructed a short M81 3=UTR that was the same length as the B95-8 3=UTR but that contained the unique M81 sequence in the 5=portion of the UTR. We also made a long 3= UTR in which the 5=-proximal B95-8 polymorphism was exchanged for the M81 sequence and therefore was the same length as the M81 3=
UTR. BALF5 protein expression in the presence of these four different 3=UTRs was then analyzed. The M81 3=UTR, the B95-8 3=UTR, the M81 3=UTR with the transposed B95-8 sequence (long chimeric 3=UTR), and the short M81 3=UTR were compared to assess their effects on BALF5 protein expression. Western blot analysis of BALF5 protein expression from transfected HEK293 cells showed that switching 3= UTR sequence polymorphisms between M81 and B95-8 did not change BALF5 protein expression compared to that with their parent UTR sequences (Fig. 9B). BALF5 protein expression
A
B
Long Chimeric 3’ UTR with B95-8 sequence polymorphism
Short M81 3’ UTR
*---*-BALF5
Tubulin
* * -M81 3’ UTR
B95-8 3’ UTR
*---*-* *
-C
SacII
PmeI
SacII
PmeI
+ SM
-M UTR +
-B UTR +
-B UTR +
-M UTR
Long Short Long Short
+ SM
-M UTR +
-B UTR +
-B UTR +
-M UTR Long Short Long Short
1.0 13.9
1.5 10.1
0.6 15.9
1.5 9.5
RQ
0 5 10 15 20
FIG 9Effect of BALF5 3=UTR length and sequence polymorphisms on BALF5 protein expression. (A)
Diagram showing the construction of an artificial short M81 3=UTR and a chimeric long B95-8 3=UTR
composed of the 5=B95-8 sequence fused to the 3=M81 sequence. BALF5 expression vectors with either
a native M81 (M UTR long), native B95-8 (B UTR short), chimeric long B95-8 (B UTR long), or artificial short
M81 (M UTR short) 3=UTR were transfected into HEK293 cells. Each construct was also cotransfected with
either the empty vector control (⫺) or a B95-8 SM expression plasmid (⫹). Restriction enzyme sites used
for the construction of 3=UTRs (PmeI and SacII) are shown. (B) Comparison of BALF5 protein expression
levels from each expression vector was performed by Western blotting. Blots were stripped and reprobed with tubulin antibody as a loading control. (C) RT-qPCR was performed with RNAs harvested from the
samples in panel B, as described above. Error bars represent standard errors of the means.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:11.585.49.364.74.311.2]was most efficient with the longer 3=UTR derived from M81 regardless of the proximal sequence polymorphism. We also measured BALF5 RNA expression levels in the above-described experiment by RT-qPCR 48 h after transfection. RT-PCR analysis showed that while RNA expression is not markedly different between the four con-structs and is upregulated by SM (Fig. 9C), BALF5 protein translation is primarily dependent on the length of the 3= UTR. These data thus demonstrate that the M81 BALF5 mRNA is translated more effectively than the B95-8 BALF5 mRNA because of the longer 3= UTR that is present in M81 EBV but truncated in B95-8 EBV. The resultant increased efficiency of DNA replication likely contributes to the M81 lytic phenotype. EBV strains derived from Chinese NPC isolates tend to cluster together taxonomi-cally and are represented by a variant strain distinct from lymphoma-associated strains and NPC strains from other geographic locales (8). The BALF5 protein-coding sequences from a variety of EBV strains from different sources (GD2, Raji, Akata, M81, C666, HKNPC1, and AG876) are highly conserved and have only 0 to 2 amino acid polymor-phisms out of 1,015 amino acids. Since the BALF5 3=UTR appears to be involved in the expression of BALF5, which may correlate with biological characteristics of the parent viral strain, we performed a taxonomic analysis of the 3= UTRs from a variety of EBV strains (Fig. 10). The results revealed that the 3=UTRs of Chinese NPC strains clustered in a manner essentially identical to that demonstrated in a broad phylogenetic analysis of these strains performed previously by Tsai et al. (8). GD2, HKNPC1, M81, and C666.1 were very close to each other and varied significantly from Akata, Raji, and AG876. GD1, a Chinese NPC isolate, was further removed from the other isolates, which was also true of its overall phylogenetic position when the entire genomes were compared (8). Thus, the 3=UTR of BALF5 appears to be highly conserved in a variant-specific manner that correlates with the level of conservation seen in coding regions of proteins used to distinguish between geographic variants.
DISCUSSION
It has long been hypothesized that strain-specific variation in EBV may contribute to the geographic variability in the incidence of NPC. EBV replication at high levels is characteristic of NPC, and the presence of EBV DNA or antibodies against replicative EBV antigens in blood is predictive of NPC (10–12). Since NPC patients may carry EBV strains with increased lytic and epitheliotropic potentials, polymorphisms in specific genes may correlate with pathogenicity and essentially serve as potential virulence
C666.1
AG876 Raji
Akata
GD1
HKNPC1
M81 GD2
FIG 10Taxonomy of EBV BALF5 3=UTR sequences from NPC and lymphoma isolates. Shown is the
phylogenetic relatedness of the BALF5 3=UTRs from NPC strains (M81, C666, HKNPC.1, GD1, and GD2)
and lymphoma strains (Raji, AG876, and Akata). The 3=UTR sequences of BALF5 from each strain
extending from the stop codon to the mapped cleavage site were compared by using ClustalW, and distance trees were constructed by using PHYLIP and DrawTree.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:12.585.145.262.73.260.2]factors for NPC. The M81 EBV strain, isolated from an EBV-positive nasopharyngeal carcinoma from a Chinese patient, exhibits increased, persistent lytic replication in B lymphocytes and epithelial cells and greater epitheliotropism than other previously described EBV strains (6–8). This study investigates the potential mechanisms un-derlying the enhanced lytic replication characteristic of the M81 strain of EBV. Tsai et al. showed that the sustained and increased lytic replication of M81 is partly due to a polymorphism in the Zta protein (8). While M81 Zta was more efficient in inducing lytic replication than B95-8 Zta, the enhanced lytic phenotype of M81 could not be entirely recapitulated by the M81 Zta protein alone, leading those authors to conclude that other M81 viral genes were required (8). Those authors also ruled out polymorphisms in the BZLF1 promoter as contributing to the enhanced lytic replicative properties of M81.
Since the EBV SM gene is essential for lytic replication and infectious virus produc-tion (13), we asked whether the M81 SM protein might contribute to the increased lytic properties of M81. We first confirmed that cells infected with recombinant M81 indeed produced larger amounts of virus after the induction of lytic replication than did cells infected with recombinant B95-8 virus. Importantly, we noted that M81 virus-infected cells undergoing lytic EBV replication contained larger amounts of EBV DNA than did B95-8-infected cells, suggesting that at least part of the M81 replicative phenotype stems from increased DNA replication. Despite differences in amino acid sequences of M81 SM and B95-8 SM, there were no significant functional differences between the two SM proteins, compared by their ability to complement an SMKO B95-8 strain. M81 SM was no more efficient than B95-8 SM in enhancing the accumulation of SM-dependent EBV mRNAs. SM also has an enhancing effect on lytic EBV DNA replication (17), and we therefore asked whether M81 SM might be more potent than B95-8 SM in its ability to stimulate EBV DNA replication, but again, the functions of both SM proteins were similar.
The EBV DNA polymerase (BALF5) mRNA is unusual in having two potential cleavage and polyadenylation sites, and SM has been shown to facilitate the processing of this RNA transcript (24–26). Although the similarity in overall DNA replication after com-plementation of SMKO EBV with either M81 SM or B95-8 SM suggested that BALF5 regulation by SM was unlikely to be the cause of the increased DNA replication in M81, we measured BALF5 expression levels to determine whether BALF5 gene polymor-phisms might contribute to the superior M81 replication. Interestingly, the BALF5 protein expression level was markedly higher in M81 despite much lower BALF5 RNA levels than those of B95-8, suggesting that the translation of the M81 BALF5 transcript was particularly robust. Since codon usage bias may contribute to differences in translation efficiencies (30), we compared the protein expression levels of the isolated BALF5 ORFs from both viral strains but found no significant differences. However, a similar comparison of the BALF5 3=UTRs revealed that the M81 3=UTR led to higher expression levels of the BALF5 protein in chimeric constructs, primarily attributable to its greater length. Comparison of the BALF5 3=UTRs of other NPC EBV strains revealed that these 3=UTR sequences are highly conserved and differ from those of lymphoma strains, suggesting that the 3=UTR of BALF5 is functionally important. The 3=UTR affects translational regulation by participation in the formation of a pseudoloop in which the 5= and 3= UTRs are brought into proximity by the interaction of eIF4G and PAPB, respectively (31). RNA-binding proteins that localize to the 3= UTR may also affect translation by potentiating or antagonizing silencing by microRNAs (miRNAs) (32, 33). The 3=UTR of BALF5 may therefore play a role in the lytic replicative capacity of EBV strains in general. The regulation of both RNA processing and the translation of BALF5 transcripts thus appears to be important in lytic EBV DNA replication. The SM protein is clearly important for enhancing BALF5 RNA accumulation, but the limiting factor in protein expression appears to be the translatability of the resultant transcript. Despite the accumulation of similar or even larger amounts of the B95-8 BALF5 transcript, the level of production of the polymerase protein is lower than that of M81. Taken together, these findings imply that EBV DNA polymerase protein production is regulated by
on November 6, 2019 by guest
http://jvi.asm.org/
translational control, which may play a role in strain-specific differences in lytic repli-cative potential and pathogenesis.
MATERIALS AND METHODS
Cells, plasmids, and DNA transfections.HEK293 cells carrying B95-8 EBV or M81 EBV bacmids, expressing GFP and hygromycin resistance genes, were maintained as monolayer cultures in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum, 1% GlutaMAX supplement (Life
Technologies), and 100g/ml of hygromycin (8, 29). Cloned HEK293 cells carrying an SM-null EBV 2089
bacmid with the SM gene inactivated by the insertion of a kanamycin resistance gene (EBVBMLF1-KO)
(SMKO) were maintained in parallel (13, 34). Raji is an EBV-positive human Burkitt lymphoma cell line carrying a replication-defective EBV strain (35). Raji cells were grown in RPMI 1640 containing 10% fetal bovine serum (Gibco). Transfections of HEK293 cells were performed with TransIT293 reagent (Mirus) according to the manufacturer’s protocol. SM, BALF4, and BZLF1 (Zta) expression vectors were previously described (16, 36, 37). FLAG-M81 SM, FLAG–B95-8 SM, M81 BALF5, and B95-8 BALF5 expression vectors were constructed by PCR amplification of the respective ORFs from B95-8 DNA or M81 DNA and cloned
into pCDNA3. The B95-8 3=UTR and the M81 3=UTR of BALF5 were synthesized by PCR, fused to the
B95-8 BALF5 ORF by Gibson assembly (NEB), and cloned into pCDNA3. By using PmeII and SacII
restriction enzymes, a short M81 3=UTR was constructed, in which the unique M81 sequences differing
from those of B95-8 were exchanged for those in the short B95-8 3=UTR. Conversely, a long B95-8 3=UTR
was constructed by exchanging the unique B95-8 region for M81 sequences in the long M81 3=UTR. All
constructs were verified by DNA sequencing.
Immunoblotting.Protein cell lysates were made from cells 48 h after transfection. Samples were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. Blots were probed with anti-Zta (Argene), anti-SM, or anti-BALF5 (a kind gift from Friedrich Grasser, Germany) polyclonal antibodies. The signal was visualized by incubation with horseradish peroxidase-conjugated secondary antibody and a chemiluminescent substrate (Bio-Rad), as previously described (38).
Immunofluorescence microscopy.For immunofluorescence, SMKO cells were cultured on cover-slips for 24 h before transfection. Cells were transfected with Zta and either FLAG-M81 SM or FLAG–B95-8 SM by using TransIT293 reagent (Mirus Bio) according to the manufacturer’s specifications. Cells were fixed with 4% paraformaldehyde 48 h after transfection and stored at 4°C until staining. Cells were permeabilized with 0.1% Triton X-100, blocked with 20% goat serum, and stained with anti-SM polyclonal antibody (1:500 dilution). The cells were washed and incubated with Alexa Fluor 594 conjugated with goat anti-rabbit IgG. Nuclei were stained with ProLong Gold antifade reagent with DAPI (catalog number P36935; Molecular Probes), and mounted slides were visualized and imaged by using an Axio Imager M2 microscope (Zeiss).
Measurement of viral DNA replication and infectious virus titers.Lytic EBV replication in HEK293 cells infected with either WT B95-8 2089 EBV, WT M81 EBV, or SM knockout EBV was induced by transfection with a Zta expression plasmid and other plasmids as indicated. Cell pellets were harvested 60 h after transfection, and DNA was purified by using a Qiagen blood and tissue DNeasy kit and used for measurement of EBV DNA copy numbers. Fifty nanograms of total cell DNA was analyzed by qPCR using EBV gene-specific primers (BILF2) to quantitate the viral genome copy number, which was
normalized to cellular-actin (17). Infectious viral particles were measured in cell supernatants 5 days
after the induction of EBV replication. Cleared supernatants were filtered through 0.8-m-pore-size
cellulose acetate filters, and serial dilutions of supernatants were used to infect Raji cells and incubated at 37°C. The numbers of GFP-positive Raji cells representing infectious EBV particles were visualized under a fluorescence microscope at 2 days postinfection, and the numbers of virus-infected cells were quantitated by flow cytometry as previously described (17, 39, 40).
RNA isolation, qPCR, and Northern blotting.Total RNA was purified from pellets of HEK293 cells by using QIAzol and miRNeasy columns (Qiagen) according to the manufacturer’s protocols. A DNase treatment step was performed after purification (Promega). The RNA level was quantitated by spectro-photometry, and qPCR was performed with 50 ng cellular RNA and SYBR green RNA-to-CT mix (ABI) according to the manufacturer’s protocol, using an Applied Biosystems StepOne Plus real-time PCR thermocycler. Controls with no template were included in each analysis. PCR amplification was per-formed in triplicate with gene-specific primers, as described previously (17). Cellular
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or-actin was used as the endogenous control for all reactions. For
Northern blotting, RNA was purified from HEK293 cells transfected with plasmid DNA by using QIAzol and miRNeasy columns (Qiagen) according to the manufacturer’s protocols. RNA was quantitated by
spectrophotometry, and 3g of RNA was electrophoresed on 1% denaturing agarose gels and
trans-ferred to membranes, as previously described (36).32P-labeled BALF5 DNA probes were generated by
using a random oligonucleotide labeling kit (Amersham), and membranes were hybridized with probes overnight at 65°C. The blots were washed, and the radioactive signal was visualized by autoradiography.
Phylogeny analysis.DNA sequences of the BALF5 3=UTRs were aligned by using ClustalW to generate distance maps and to generate PHYLIP-compatible output trees (41). Trees were visualized by using the DrawTree program in PHYLIP (42). Protein sequences were translated and aligned by using BLAST (43). EBV genome sequences used in the analyses were as follows: GD1 (GenBank accession
number AY961628.3), GD2 (GenBank accession number HQ020558.1), HKNPC1 (GenBank accession
number JQ009376.1), Akata (GenBank accession number KC207813.1), the EBV reference genome
(GenBank accession numberNC_007605.1), and B95-8 (GenBank accession numberV01555.2).
on November 6, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant RO1 81133 (S.S.) from the National Cancer Institute and grant 1I01BX002262 (S.S.) from the ORD BLR&D.
DNA sequencing and flow cytometry were performed at the University of Utah Health Sciences Core Facilities.
REFERENCES
1. Longnecker RM, Kieff E, Cohen JI. 2013. Epstein-Barr virus, p 1898 –1959.
InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA,
Racaniello VR, Roizman B (ed), Fields virology, 6th ed, vol 2. Lippincott Williams & Wilkins, Philadelphia, PA.
2. Rickinson AB, Kieff E. 2007. Epstein-Barr virus, p 2655–2700.InKnipe
DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 2. Lippincott Williams & Wilkins, Philadelphia, PA.
3. Chang CM, Yu KJ, Mbulaiteye SM, Hildesheim A, Bhatia K. 2009. The extent of genetic diversity of Epstein-Barr virus and its geographic and
disease patterns: a need for reappraisal. Virus Res 143:209 –221.https://
doi.org/10.1016/j.virusres.2009.07.005.
4. Hsu JL, Glaser SL. 2000. Epstein-Barr virus-associated malignancies: ep-idemiologic patterns and etiologic implications. Crit Rev Oncol Hematol
34:27–53.https://doi.org/10.1016/S1040-8428(00)00046-9.
5. De-The G, Ho HC, Kwan HC, Desgranges C, Favre MC. 1970. Nasopha-ryngeal carcinoma (NPC). I. Types of cultures derived from tumour biopsies and non-tumorous tissues of Chinese patients with special reference to lymphoblastoid transformation. Int J Cancer 6:189 –206. https://doi.org/10.1002/ijc.2910060206.
6. Desgranges C, Lenoir G, de-The G, Seigneurin JM, Hilgers J, Dubouch P. 1976. In vitro transforming activity of EBV. I. Establishment and proper-ties of two EBV strains (M81 and M72) produced by immortalized Callithrix jacchus lymphocytes. Biomedicine 25:349 –352.
7. Desgranges C, Lavoue MF, Patet J, de-The G. 1979. In vitro transforming activity of Epstein-Barr virus (EBV). II. Differences between M81 and B95-8 EBV strains. Biomedicine 30:102–108.
8. Tsai MH, Raykova A, Klinke O, Bernhardt K, Gartner K, Leung CS, Geletneky K, Sertel S, Munz C, Feederle R, Delecluse HJ. 2013. Spon-taneous lytic replication and epitheliotropism define an Epstein-Barr
virus strain found in carcinomas. Cell Rep 5:458 – 470.https://doi.org/
10.1016/j.celrep.2013.09.012.
9. Swaminathan S, Kenney S. 2009. The Epstein-Barr virus lytic life cycle, p
285–315.InDamania B, Pipas J (ed), DNA tumor viruses. Springer, New
York, NY.
10. Chan KCA, Woo JKS, King A, Zee BCY, Lam WKJ, Chan SL, Chu SWI, Mak C, Tse IOL, Leung SYM, Chan G, Hui EP, Ma BBY, Chiu RWK, Leung SF, van Hasselt AC, Chan ATC, Lo YMD. 2017. Analysis of plasma Epstein-Barr virus DNA to screen for nasopharyngeal cancer. N Engl J Med 377:
513–522.https://doi.org/10.1056/NEJMoa1701717.
11. Chien YC, Chen JY, Liu MY, Yang HI, Hsu MM, Chen CJ, Yang CS. 2001. Serologic markers of Epstein-Barr virus infection and nasopharyngeal
carcinoma in Taiwanese men. N Engl J Med 345:1877–1882.https://doi
.org/10.1056/NEJMoa011610.
12. Cao SM, Liu Z, Jia WH, Huang QH, Liu Q, Guo X, Huang TB, Ye W, Hong MH. 2011. Fluctuations of Epstein-Barr virus serological antibodies and risk for nasopharyngeal carcinoma: a prospective screening study with a
20-year follow-up. PLoS One 6:e19100.https://doi.org/10.1371/journal
.pone.0019100.
13. Gruffat H, Batisse J, Pich D, Neuhierl B, Manet E, Hammerschmidt W, Sergeant A. 2002. Epstein-Barr virus mRNA export factor EB2 is essential
for production of infectious virus. J Virol 76:9635–9644.https://doi.org/
10.1128/JVI.76.19.9635-9644.2002.
14. Batisse J, Manet E, Middeldorp J, Sergeant A, Gruffat H. 2005. Epstein-Barr virus mRNA export factor EB2 is essential for intranuclear capsid
assembly and production of gp350. J Virol 79:14102–14111.https://doi
.org/10.1128/JVI.79.22.14102-14111.2005.
15. Ruvolo V, Sun L, Howard K, Sung S, Delecluse HJ, Hammerschmidt W, Swaminathan S. 2004. Functional analysis of Epstein-Barr virus SM protein: identification of amino acids essential for structure, transactiva-tion, splicing inhibitransactiva-tion, and virion production. J Virol 78:340 –352. https://doi.org/10.1128/JVI.78.1.340-352.2004.
16. Han Z, Marendy E, Wang Y-D, Yuan J, Sample JT, Swaminathan S. 2007.
Multiple roles of Epstein-Barr virus SM protein in lytic replication. J Virol
81:4058 – 4069.https://doi.org/10.1128/JVI.02665-06.
17. Thompson J, Verma D, Li D, Mosbruger T, Swaminathan S. 2015. Iden-tification and characterization of the physiological gene targets of the essential lytic replicative Epstein-Barr virus SM protein. J Virol 90:
1206 –1221.https://doi.org/10.1128/JVI.02393-15.
18. Winkler M, Rice SA, Stamminger T. 1994. UL69 of human cytomegalovi-rus, an open reading frame with homology to ICP27 of herpes simplex virus, encodes a transactivator of gene expression. J Virol 68:3943–3954. 19. Sandri-Goldin RM. 2011. The many roles of the highly interactive HSV protein ICP27, a key regulator of infection. Future Microbiol 6:1261–1277. https://doi.org/10.2217/fmb.11.119.
20. Han Z, Swaminathan S. 2006. Kaposi’s sarcoma-associated herpesvirus lytic gene ORF57 is essential for infectious virion production. J Virol
80:5251–5260.https://doi.org/10.1128/JVI.02570-05.
21. Verma D, Thompson J, Swaminathan S. 2016. Spironolactone blocks Epstein-Barr virus production by inhibiting EBV SM protein function.
Proc Natl Acad Sci U S A 113:3609 –3614.https://doi.org/10.1073/pnas
.1523686113.
22. Fixman ED, Hayward GS, Hayward SD. 1992.trans-Acting requirements
for replication of Epstein-Barr virus ori-Lyt. J Virol 66:5030 –5039. 23. Fixman ED, Hayward GS, Hayward SD. 1995. Replication of Epstein-Barr
virus oriLyt: lack of a dedicated virally encoded origin-binding protein and dependence on Zta in cotransfection assays. J Virol 69:2998 –3006. 24. Silver Key SC, Pagano JS. 1997. A noncanonical poly(A) signal, UAUAAA, and flanking elements in Epstein-Barr virus DNA polymerase mRNA function in cleavage and polyadenylation assays. Virology 234:147–159. https://doi.org/10.1006/viro.1997.8647.
25. Silver Key SC, Yoshizaki T, Pagano JS. 1998. The Epstein-Barr virus (EBV) SM protein enhances pre-mRNA processing of the EBV DNA polymerase transcript. J Virol 72:8485– 8492.
26. Furnari FB, Adams MD, Pagano JS. 1993. Unconventional processing of
the 3=termini of the Epstein-Barr virus DNA polymerase mRNA. Proc Natl
Acad Sci U S A 90:378 –382.https://doi.org/10.1073/pnas.90.2.378.
27. Yu CH, Dang Y, Zhou Z, Wu C, Zhao F, Sachs MS, Liu Y. 2015. Codon usage influences the local rate of translation elongation to regulate
co-translational protein folding. Mol Cell 59:744 –754.https://doi.org/10
.1016/j.molcel.2015.07.018.
28. Ricci EP, Mure F, Gruffat H, Decimo D, Medina-Palazon C, Ohlmann T, Manet E. 2009. Translation of intronless RNAs is strongly stimulated by the Epstein-Barr virus mRNA export factor EB2. Nucleic Acids Res 37:
4932– 4943.https://doi.org/10.1093/nar/gkp497.
29. Delecluse HJ, Hilsendegen T, Pich D, Zeidler R, Hammerschmidt W. 1998. Propagation and recovery of intact, infectious Epstein-Barr virus from prokaryotic to human cells. Proc Natl Acad Sci U S A 95:8245– 8250. https://doi.org/10.1073/pnas.95.14.8245.
30. Quax TE, Claassens NJ, Soll D, van der Oost J. 2015. Codon bias as a
means to fine-tune gene expression. Mol Cell 59:149 –161.https://doi
.org/10.1016/j.molcel.2015.05.035.
31. Szostak E, Gebauer F. 2013. Translational control by 3=-UTR-binding
proteins. Brief Funct Genomics 12:58 – 65.https://doi.org/10.1093/bfgp/
els056.
32. Kedde M, van Kouwenhove M, Zwart W, Oude Vrielink JA, Elkon R, Agami
R. 2010. A Pumilio-induced RNA structure switch in p27-3=UTR controls
miR-221 and miR-222 accessibility. Nat Cell Biol 12:1014 –1020.https://
doi.org/10.1038/ncb2105.
33. Kedde M, Strasser MJ, Boldajipour B, Oude Vrielink JA, Slanchev K, le Sage C, Nagel R, Voorhoeve PM, van Duijse J, Orom UA, Lund AH, Perrakis A, Raz E, Agami R. 2007. RNA-binding protein Dnd1 inhibits
microRNA access to target mRNA. Cell 131:1273–1286.https://doi.org/
10.1016/j.cell.2007.11.034.
34. Gruffat H, Manet E, Sergeant A. 2002. MEF2-mediated recruitment of class II HDAC at the EBV immediate early gene BZLF1 links latency and
on November 6, 2019 by guest
http://jvi.asm.org/
chromatin remodeling. EMBO Rep 3:141–146.https://doi.org/10.1093/
embo-reports/kvf031.
35. Pulvertaft JV. 1965. A study of malignant tumours in Nigeria by
short-term tissue culture. J Clin Pathol 18:261–273.https://doi.org/10.1136/jcp
.18.3.261.
36. Ruvolo V, Wang E, Boyle S, Swaminathan S. 1998. The Epstein-Barr virus nuclear protein SM is both a post-transcriptional inhibitor and activator
of gene expression. Proc Natl Acad Sci U S A 95:8852– 8857.https://doi
.org/10.1073/pnas.95.15.8852.
37. Neuhierl B, Feederle R, Hammerschmidt W, Delecluse HJ. 2002. Glyco-protein gp110 of Epstein-Barr virus determines viral tropism and
effi-ciency of infection. Proc Natl Acad Sci U S A 99:15036 –15041.https://
doi.org/10.1073/pnas.232381299.
38. Verma D, Kim EA, Swaminathan S. 2013. Cell-based screening assay for antiviral compounds targeting the ability of herpesvirus posttran-scriptional regulatory proteins to stabilize viral mRNAs. J Virol 87:
10742–10751.https://doi.org/10.1128/JVI.01644-13.
39. Verma D, Li DJ, Krueger B, Renne R, Swaminathan S. 2015. Identification of the physiological gene targets of the essential lytic replicative Kapo-si’s sarcoma-associated herpesvirus ORF57 protein. J Virol 89:1688 –1702. https://doi.org/10.1128/JVI.02663-14.
40. Li DJ, Verma D, Mosbruger T, Swaminathan S. 2014. CTCF and Rad21 act as host cell restriction factors for Kaposi’s sarcoma-associated herpesvi-rus (KSHV) lytic replication by modulating viral gene transcription. PLoS
Pathog 10:e1003880.https://doi.org/10.1371/journal.ppat.1003880.
41. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, Mc-William H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. 2007. Clustal W and Clustal X version 2.0. Bioinformatics
23:2947–2948.https://doi.org/10.1093/bioinformatics/btm404.
42. Felsenstein J. 1989. PHYLIP—Phylogeny Inference Package (version 3.2). Cladistics 5:164 –166.
43. NCBI Resource Coordinators. 2016. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res 44:D7–D19.
https://doi.org/10.1093/nar/gkv1290.