0022-538X/87/082642-06$02.00/0
Copyright© 1987,AmericanSociety forMicrobiology
Virus Attenuation
and
Identification of Structural Proteins of
Vaccinia Virus That Are Selectively Modified during
Virus
Persistence
EDUARDO PAEZ,t SHATHA DALLO, AND MARIANO ESTEBAN*
Departments ofBiochemistry andMicrobiologyandImmunology, State UniversityofNew York,Health Science Center atBrooklyn, New York, New York 11203
Received29December1986/Accepted 1May 1987
Toinvestigate thegenetic stability of vacciniavirusDNA,wehave tested whetheralterations occurred in the polypeptide compositionof thiscomplexvirusduring persistentinfections. We found that variants isolatedat variouspassagesinFrienderythroleukemia cells persistently infectedwith vaccinia viruscontained,in addition to an8-megadalton (MDa) deletion onthe left terminusof theviralgenome,majoralterations in the sizes of three structuralproteins with molecularmassesof about39, 21, and 14 kDa. Alterations in isoelectricpoints werealsoobserved inproteins of48, 27,and 14 kDa. The 14-kDaproteinispartof the virusenvelope,and the variants increased thesizeof thisproteinfrom 0.5to3 kDawithincreasingpassage number. Alteration in size of the 14-kDaproteinisadominant trait sinceitappearedin the whole viruspopulationbypassage 48. With more passages, somevariantswerefoundtoincreaseordecreasethesizeofa39-kDacoreprotein byabout 2 kDa and todecrease the sizeofan envelopeprotein of21 kDa byabout 2 kDa. These three proteinswere immunogenic in mice and elicited a strong host immune response. Major alterations in the sizes of these proteinswereprevented by continuoustreatmentofthepersistentlyinfected cultures with interferon.However, after interferonwasremoved,proteinmodificationsappearedwithincreasingpassagenumber. Generationof the 8-MDa deletion and alterations inthesize ofthe 14-kDaprotein correlated witha marked decreasein virulence of these variants. Our findings suggest that during virus persistence, specific mutations are introducedin the vaccinia virusgenome that leadtoprotein alterations andtohighly attenuatedviruses.
Itis believed thatpoxviruses are wellconserved through evolution. With theexception of heterogeneityatthe endsof
the viral genome, aremarkable sequence conservation has
been observed withinthecentralregion ofthe genome(fora
review, see references 3 and 10). These previous studies were carried out by restriction enzyme analysis of viral genomes (9)obtained after afewcycles ofvirus multiplica-tion and with serially propagated viruses(10). Becauseofthe
cytocidal nature ofpoxviruses (1), most procedures used
would have selected for the virus population that grew
faster. Our aimwas toestablishpersistent infectionsto test the genetic stability of poxviruses. This method could
pro-vide geneticmarkers thatpredict with certainty the virulence or attenuation of these viruses. We have previously
de-scribedtwopersistently infected vaccinia virus-cell systems
from Friend erythroleukemia (FEL) cells. The first persist-entinfectionwasin theuntreatedcultures (11), whereas the second persistent infection was established in the presence
ofinterferon (IFN) (13).During virus persistence, a
charac-teristic8-megadalton (MDa) deletion is generated at the left terminus of the viral genome (11). By restriction enzyme analysis, no other changes except genetic alterations at the ends wereobserved during virus persistence. Because vac-cinia virus has a large DNA (185 kilobases) which encodes more than 100 structural polypeptides (5), it is likely that
genetic alterations other than deletions are also introduced
into the virus population during virus persistence. In this
investigation we havetested whether protein modifications are introduced in vaccinia virus when it is grown under
*Correspondingauthor.
tPresent address: Centrode Investigaciones Biologicas, 28006 Madrid, Spain.
stringentselective conditionsimposed byapersistent infec-tion in FEL cells and by IFN treatment. We have charac-terized representative virus mutants in the course of a
persistentvirus infection that lasted 2 years.
Structuralproteinmodifications in variants from untreated
persistently infected FEL cells and in variants from IFN-treated persistently infected FEL cells were examined by
one-and two-dimensionalgel electrophoresisof35S-labeled
proteins frompurified virions and by immunoblot analysis.
Unlabeledor[35S]methionine-labeled purifiedvaccinia virus orvariants weredisruptedat100°Cfor 3 min insamplebuffer
(62.5 mMTris hydrochloride [pH 6.8], 4% sodiumdodecyl sulfate [SDS], 10% 2-mercaptoethanol, 10% glycerol, 0.02% bromophenol blue). One-dimensional gel electrophoresis
was run on 12% gels as described elsewhere (12), and two-dimensional gel electrophoresis was carried out as
de-scribed previously (2). Isoelectric focusingwas done in the
first dimension withampholines (pH 5 to 7 and 3 to 10) and in the second dimensionwas doneby SDS-12%
polyacryl-amidegelelectrophoresis (PAGE). Thegels weredriedout and exposed to X-AR5 film for autoradiography. For
im-munoblotanalysis,40,ugofprotein from purified virions was
appliedper well and fractionated with SDS-12% polyacryl-amide gels. Theproteins were transferred to nitrocellulose and immunoblots were developed as described previously (16), except that 5% blotto (nonfat dry milk) was used
instead of fetal calf serum. Monoclonal antibodies against the14-kDa envelope protein of vaccinia virus wereprepared as described previously (16). Polyclonal antiserum was raised in rabbitsimmunizedwith either purified virus
enve-lopesorpurified virus cores.Viralenvelopes wereprepared from5mg ofpurified virus suspended in 1 ml bysonication (500 W for 10 s) in 50 mM Tris hydrochloride (pH 8.5)-10
2642
on November 10, 2019 by guest
http://jvi.asm.org/
mM
MgC12-10
mM dithiothreitol-0.5% Triton X-100. After incubation at37°C
for 30min,the extract was layered on top of a 36% sucrose cushion in the same buffer, and cores were pelleted by centrifugation at 20,000 x g for 60 min at4°C.
The upper phase, containing the viral envelopes, was recov-ered and dialyzed extensively against phosphate-buffrecov-ered saline. Cores were resuspended in the same buffer and pelleted through a sucrose cushion. This process was re-peated three times. Cores were finally suspended in phos-phate-buffered saline. Rabbits were immunized by three intramuscular injections of purified viral envelopes or cores (100,ug
of protein). The first injection was in Freund complete adjuvant, and the other two injections were admin-istered every 15 days but without adjuvant. One week later, rabbits were bled from the ear vein and serum samples were collected. The antibody levels remained high even 6 months after the last inoculation. When the pattern of35S-labeledvirion proteins was analyzed by one-dimensional SDS-PAGE (Fig. 1A), we observed changes in the sizes of proteins with molecular masses of about 39 (clones 65-16 and 101-14), 21 [clone (88)-24, not shown], and 14 [clones 53-8, 65-16, (74)-23, and 101-14] kDa. Two-dimensional gel elec-trophoretic analyses revealed additional information. For simplicity, only the results obtained with the wild-type virus and clone 101-14 are presented. Several proteins were found to change in mutant 101-14 compared with proteins of the
wild-type
virus. These proteins (Fig.1A, denoted by arrows in part WR) have molecular weights of about 64, 48, 39, 27, 24, and 14 kDa.Next, we tested whether these modified proteins were immunogenic. Purified virus mutants were run on one- and two-dimensional SDS-PAGE gels, blotted on nitrocellulose
paper, and reacted with anti-vaccinia rabbit antiserum, and the antigenic viral proteins were identified by the immuno-peroxidase staining method (16). By inspection of the im-munoblot of proteins run on one-dimensional SDS-PAGE gels (Fig. 1B), it was clear that three proteins of molecular weights of about 39, 21, and 14 kDa changed in size in variants from persistently infected FEL cells. In some variants, the 39-kDa protein either gained (variants 65-16 and 65-17) or lost (variants 101-12 and 101-14) about 2 kDa, the 21-kDa protein lost about 2 kDa [clone (88)-24], and the 14-kDa protein gained from 0.5 to 3 kDa inall variants at or after 48 passages. These protein modifications were not observed in variants isolated from IFN-treated persistently infected FEL cells. Significantly, when IFN treatment was discontinued and virus persistence was maintained for vari-ous passages, variants emerged that had protein modifica-tions similar to those of variants from untreated persistently infected FELcells. Immunoblot analysis of proteins run in two-dimensional gel electrophoresis (Fig. 1B, lower panel) confirmed that the changes occurred in proteins that have an
immunogenic role.
To identify whether mutations occur within the same protein, we used monoclonal antibodies prepared against vaccinia virus (16). One such monoclonal antibody (mAbC3) was found to react against the modified 14-kDa protein, as revealed by immunoblot analysis (Fig. 1C); a breakdown product was also detected (16). The 14-kDa polypeptide is an envelope protein that must play an important role in virus-host cell interactions. It has been demonstrated that this protein causescellfusion and is involved in virus penetration and that monoclonal antibodies reactive against this protein have neutralizing activity (16, 17). In variants from untreated persistently infected FEL cells, the 14-kDa protein gained from 0.5 to 3 kDa with increasing passage numbers. In
contrast, no change oraminor increase in size was observed in variantsfrom IFN-treated persistently infected FEL cells. For example, at passage 48, the 14-kDa protein gained 1.5 kDa in size with variants from untreated persistently
in-fected FEL cells and only about 0.5 kDa in variants from
IFN-treated persistently infected FEL cells. After removal ofIFNandanadditional40passages in the absenceofIFN, variants were generated that had gained 1.5 to 2 kDa insize, whereas if IFN treatment was maintained, no additional changes were observed.
Next we examined thesevariants for a number of proper-ties relevantto virulence. These findings are summarized in Table 1. Wefoundthat variants from untreatedpersistently infected cellsorfrom persistently infected cultures afterIFN
was removed had the same deletion of about 8 MDa atthe left end of the viral genome as previously described (11). This deletion appeared at relatively early passages and was maintained with later passages. Significantly, continuous treatment of the persistently infected cultures with IFN
completely blocked the generation of deletions, even 2 years after the establishment of virus persistence. Another obser-vation was that variants isolated after later passages of untreated persistently infected FEL cells had greatly
re-duced plaque sizes compared with the wild-type virus or
with variants isolated at similar passages but from
IFN-treated persistently infected FEL cells. This reduction in plaque size was greater at later cell passages (clone 101-14) than that at passage 48 described previously (11). However, in IFN-treated cultures, the plaque size was only slighty reduced even at late cell passages. An enhanced cell fusion ability of the virus isolates was observed with increasing passage number, independently of the IFN treatment. For example, mutant 30-10 from untreated cultures and mutant
IFN 87-4 from IFN-treated cultures caused cellstofuse into large polykaryocytes, a cytopathic effect completely dif-ferentfromthatof the wild-type virus, which usually caused cells to round and clump together. However, virus isolates from late cell passages of untreated cultures lost this char-acteristic (clones 101-12 and 65-16). This effect may be due to reversion to the wild-type pheotype, because mutations occur within the same gene or within another gene(s) that blocks this cell fusion capacity. These mutants induced a
typical early cell rounding, with the exception of clonesIFN
87-4, 53-8, and (56)-14, which showed a delay of several hours in the appearance of cell rounding. The degrees of
infectivity and virulence of these variants were tested with culture cells and with experimental animals. Some of these mutants had greatly reduced infectivity and virulence. Sig-nificantly, a good correlation was established between the appearance of deletions at the left terminus of the viral genome and greatly reduced morbidity compared with that
of the wild-type virus. Mutant 14-3 contained an 8-MDa deletion at the left end of the genome, produced a normal plaque size, and had a 2-log reduction invirulence. With the appearance of the small plaque size (mutant 48-7), the virulence decreased by 3 logs,suggesting thattheplaque size may be another genetic markerresponsible for virus
atten-uation.
The particular relevance of this study is that we have identified specific protein modifications in vaccinia variants from untreated persistently infected FEL cells. These mod-ifications do not occur in variants from IFN-treated persis-tently infected FEL cells. Changes in size have been ob-served primarily in structural viralproteins of about 39, 21, and 14 kDa. These are immunogenic proteins (Fig. 1). We have also observed alterations in the isoelectric points of
on November 10, 2019 by guest
http://jvi.asm.org/
WR
101-14A
,, e-
IV-594 4" 0^ i..
am
'wa
*- a164 SI.
14-euiseca0
-\
a
as-4~~~
S-a
a we
-._
_i*qp-- i
b
0
9
B
. 4
i..-r
--- L, .-l ,T
-59
r-:
IW.~~~~~ ~ ~ -- .* -; -38 -2740
eeefl.S&-~~~~~Aa 16CS05014
go * _JIb010 *o, W .:" f *Xa _t
.. .. *,¢ . * w W^ *do *Mo.
V
a..
t
65-16
-59
-.36
-27
-14
IEF
0
059-
- X0
0'
36-36- _ _490
27--59
21-j
i4-S
nr
4_O'
-x _
-36
-27
-21
-14
...
w
40
WR
-T
9
9
=4*14.
on November 10, 2019 by guest
http://jvi.asm.org/
C
N
I
.o m 1
*
*w-.-- -I
-4P
-FIG. 1. Alterations in the electrophoretic mobilities of several structural vaccinia virus proteinsduring virus persistence. Conditions for virus persistence in FEL cells, treatment with IFN, virus purification, and virus titration weredone asdescribed previously (11-13). (A) [35S]methionine-labeled purified viruses isolated at different passages from persistently infected cells run in one-dimensional (left) and two-dimensional (right) gel electrophoreses and analyzed by autoradiography. Numbersatthetopindicate variantsasdescribed inTable1. Abbreviations: WR, wild-type virus; SDS, sodium dodecyl sulfate; IEF, isoelectric focusing. (B) Alterations in immunogenic structural proteins of vaccinia virus mutants. Purified viruses run in one-dimensional (top) and two-dimensional(bottom) SDS-PAGE areshown. (C) Immunoblot of virus proteins reacted with monoclonal antibody mAbC3 specific for the 14-kDa envelope protein and visualized bythe immunoperoxidasestaining method. The low-molecular-weight product is thoughttobegeneratedby specific proteolytic cleavage. Numbers atthe top of thegels indicate variants asdescribed in Table1. Arrowspointtopolypeptides (39, 27,24,and 14kDa) whicharealtered in mutantviruses. The molecular masses (kDa) of some structuralpolypeptides of vaccinia virusareindicatedatboth sides.
TABLE 1. Propertiesofspontaneous vaccinia virus mutantsfrompersistently infectedFELcellsa
Reduced
Treatment Virus Deletion Plaque size Amt of cell infectivity LD50
variantb occurrence (mm)' fusiond incultured (PFUY
cells'
None WT No Normal (3) + No 106
5-3 No Normal (3) ++ No 106
14-1 Yes Normal (3) + + No
14-3 Yes Normal (3) + + No 108
30-1 Yes Normal (3) + + + + No 108
30-10 Yes Normal (3) + +++ No
48-7 Yes Small(1) ++ + + Yes 109
48-21 Yes Small(1) ++ + + Yes
101-12 Yes Verysmall (0.5) + Yes
101-14 Yes Verysmall (0.5) + Yes >109
IFN 48-42 No Normal(3) + + + No 107
48-47 No Medium(2) ++ + + No
87-1 No Medium(2) + + + + No
Noneg 53-7 Yes Small(1) + + Yes
53-8 Yes Small(1) + + Yes
65-16 Yes Very small(0.5) + Yes >109
65-17 Yes Very small(0.5) + Yes
IFNremovedh (56)-12 No Small (1) + + Yes i07
(56)-14 Yes Small(1) ++ Yes
(74)-22 Yes Small(1) ++++ Yes
(74)-23 Yes Small(1) ++++ Yes
(88)-24 No Small(1) ++++ Yes
aVariantswereisolated fromuntreatedandIFN-treatedpersistentlyinfectedFELcells.Theoccurrenceof deletionswasdeterminedbySouthernblots,as
describedpreviously (13),andtheplaquesizewasestimated in BSC-40 cells.
bThefirst number indicates the cell passage, and the secondnumberindicatestheclone. Relativetothenormalsize ofWT virus.
dScreenedat24hafter infectionwith 1 PFUofeachvariantpercell. +,clumpsof cells with littlecellfusion; ++,scatteredpolykaryocytes; + ++, many
polykaryocytes;++++,allcells fused intolargepolykaryocytes.
'Viralinfectivitywasmeasuredbytitrationvirusyieldforat24hafterinfectionwith 1 PFUof eachvariant percell;variantsthatproducedless than70% ofthe
wild-type virus yieldwereconsideredtohave reducedinfectivity.Evaluationofvirulencewasestimated inwhitefemaleBALB/cmice(CharlesRiverBreeding
Laboratories, Inc.)6 to 8weeksold afterintraperitonealinoculation(0.2ml)with differentdilutionsinphosphate-bufferedsaline of wildtypeand variants. fVirulencewasscored14daysafter virusinoculation(10micepergroup),and the dose that caused50%mortality (LD_%)wasestimated(21).
gVariants isolated fromindependentlyestablishedculturesof untreatedpersistentlyinfected FELcells.
hCultures were treated with IFN continuously during 14, 30, or45 passages; thereafter, IFN treatment was discontinued and cultures were passaged continuouslywithoutIFNuntil cellpassages56, 74,or88,respectively.
qml.4.
on November 10, 2019 by guest
http://jvi.asm.org/
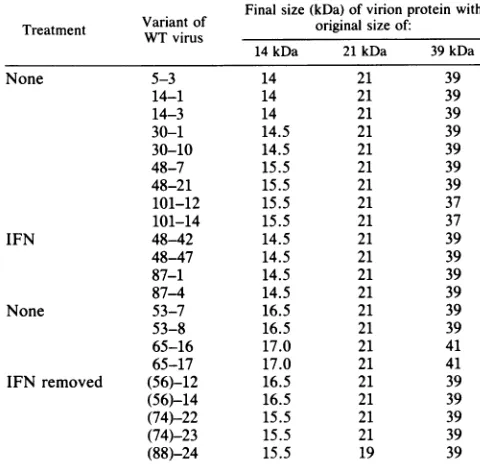
[image:4.612.59.557.375.613.2]TABLE 2. Alterations in sizes of structuralproteinsfrom spontaneous vaccinia virusmutants'
Finalsize(kDa)of virionprotein with
Treatment Variant of original size of:
WTvirus
14 kDa 21kDa 39 kDa
None 5-3 14 21 39
14-1 14 21 39
14-3 14 21 39
30-1 14.5 21 39
30-10 14.5 21 39
48-7 15.5 21 39
48-21 15.5 21 39
101-12 15.5 21 37
101-14 15.5 21 37
IFN 48-42 14.5 21 39
48-47 14.5 21 39
87-1 14.5 21 39
87-4 14.5 21 39
None 53-7 16.5 21 39
53-8 16.5 21 39
65-16 17.0 21 41
65-17 17.0 21 41
IFN removed (56)-12 16.5 21 39
(56)-14 16.5 21 39
(74)-22 15.5 21 39
(74)-23 15.5 21 39
(88)-24 15.5 19 39
a Alterations of the 14-, 21-, and 39-kDa virionproteinsweremeasuredby
one-andtwo-dimensional PAGE ofpurifiedvariants with vesicular stomatitis virusand vaccinia virusproteinsasmarkers. Theoriginandtreatmentofthe
variantsare asdescribed in Table1footnotes.
proteins of about 48, 27, and 14 kDa; other alterations included proteins of about 64 and 24 kDa which were
apparentlymissingin some mutants,possiblybecauseoneof
the proteins was altered in size and the mutated protein comigrated with theotherprotein. By examiningthe distri-bution of these proteins between the envelope and core componentsof the virus(16), wehave foundthatproteins of
24, 21, and 14kDaarepartof theenvelope,whereasproteins of39and 27 kDa arepartofthe core structureof vaccinia virus. It is significant that the most frequent protein
alter-ation occurs in the 14-kDa protein. Modifications in this
proteinareintroduced intheviralpopulation atpassage48. Thechangeconsists of anincreasein size from 0.5 to 3 kDa with increasing passage number of untreated persistently
infected FEL cells. In contrast, no change or a minor increase insize was observed in variants from IFN-treated
persistently infected FEL cells. Significantly, when IFN treatmentwasdiscontinuedandviruspersistence was
main-tained for various passages, variants emerged that have
14-kDa-protein modifications similar to those of variants
from untreated persistently infectedFEL cells. Thus, there is a cause-and-effect relationship between IFN treatment andprevention ofprotein modifications. It should be noted
thatthe8-MDadeletionis the first apparent mutational event
introduced in the virus population at relatively early pas-sages.Thereafter, protein modifications follow (Tables 1 and
2). How IFN blocks DNA sequence alterations is not known. This inhibition could result from IFN-mediated
changes in the topology of DNA, in the activity of DNA
replicating enzymes, orin other cellular factors involved in DNA metabolism (7). In fact, various effects of IFN at the DNA level have been described. These various effects
include decreased elongation (6), impaired homologous DNA recombination (14), and inhibition on stabilization or
integration of viral and cellular DNAs into animal cells (4, 15).
Inthisstudy wehave also defined a number ofbiological properties of vaccinia virus that are relevant to virulence. Compared with thewild-type virus or with variants isolated at similar passages but from IFN-treated persistently in-fected FELcells, variants isolated afterlong-termpassages of untreated persistently infected FEL cells have greatly reduced plaque sizes in cells in culture and reduced morbid-ity wheninoculated intraperitoneally into mice. The small-plaque-sizephenotypeand an 8-MDa deletionatthe left end of the viral DNA are two genetic determinants that correlate with a decreased virulence of vacciniavirus (Table 1). We have also observed that some of these variants produce
extensive cell fusion.Analysis of the cytopathic effect at late timespostinfection revealed an enhanced cell fusion ability of the virus isolates withincreasingpassagenumber, despite
the IFN treatment. Cell-cell fusion ability has been associ-ated with the production of nonglycosylated forms of the virushemagglutinin present on thesurface of virions(8, 20).
However, we have observed agoodcorrelation between the enhancedcell-cellfusionability and alterationsin the 14-kDa viralenvelopeprotein (Tables 1and 2). Infact, thisprotein
is involved in virus-induced cell-cell fusion at late times
postinfection, because the formation ofpolykaryocytes can beblockedcompletely by monoclonal antibodiesagainstthe 14-kDaprotein(17). Since some mutants isolatedatlate cell passages have lost theabilitytoformpolykaryocytes despite
altered 14-kDa proteins, it is possible that this viral mem-brane protein forms interactive complexes and that muta-tions in any one component of the complex may alter the structure andfunction ofthe entire complex. This process has been observed inherpesvirus (18, 19).
The significance of mutational changes in the structural
polypeptides of vacciniavirusduring persistent infectionsis that some of these changes might contribute to decreased virus pathogenesis and, in turn, provide specific genetic
markers relevant to virus attenuation. In addition, some of these changes may have contributed to strain variation of
poxvirus through evolution. In this regard, we have noted
thatamutated 41-kDa protein of vaccinia virus comigrates
and cross-reacts with amajor structural protein ofcowpox
virus.Itwillbeof interesttodeterminetheroleofthealtered
vacciniaviruspolypeptidesinvirusattenuationand whether
genetic diversionby members oftheorthopoxvirusgroup is related to any of the mutated proteins described in this report.
Thisinvestigation was supported by PublicHealth Servicegrant CA44262fromtheNational Institutes of Health andbygrantCCA 8510from theUSA-Spain Joint Committee for Scientific and Tech-nologicalCooperation. E.P.was arecipient ofafellowship fromthe ConsejoSuperiordeInvestigaciones Cientificas, Spain.
We thank VictoriaJimenez for her skilledtechnical assistance.
LITERATURE CITED
1. Bablanian, R. 1984. Poxvir-us cytopathogenicity: effects on cellularmacromolecularsynthesis, p.391-429. In H. Fraenkel-Conrat andR. G. Wagner(ed.), Comprehensive virology, vol. 19. PlenumPublishing Corp., NewYork.
2. Celis, J. E., and R. Bravo. 1984. Two-dimensional gel electro-phoresis of proteins: methods and applications. Academic Press, Inc., NewYork.
3. Dales, S., and B. G. T. Pogo. 1981. Biology of poxviruses. In D. W. Kingsbury and H. zur Hausen (ed.), Virology mono-graphs,vol. 18. Springer-Verlag, New York.
on November 10, 2019 by guest
http://jvi.asm.org/
4. Dubois, M. F., M. Vignal, M. Le Cunff, and C. Chany. 1983. Interferon inhibits transformation ofmousecellsbyexogenous cellularorviralgenes.Nature(London) 303:433-435.
5. Essani,K., and S. Dales. 1979.Biogenesis ofvaccinia:evidence formorethan 100 polypeptides. Virology95:385-394.
6. Esteban, M. 1984. Analysis of replicating vaccinia DNA in interferon-treated,virusinfectedcells. J.Interferon Res. 4:179-192.
7. Esteban, M., and E. Paez. 1985. Antiviral and antiproliferative properties of interferons: mechanism of action. Prog. Med. Virol. 32:159-173.
8. Ichihashi, Y., and S. Dales. 1971. Biogenesis of poxviruses: interrelationships between hemagglutinin production and polykaryocytosis. Virology46:533-543.
9. Mackett, M., and L. C. Archard. 1979. Conservation and variation in Orthopoxvirus genome structure. J. Gen. Virol. 45: 683-701.
10. Moss, B. 1985. Replication ofpoxviruses,p. 685-703. In B. N. Fields (ed.), Virology.RavenPress, New York.
11. Paez, E., S. Dallo, and M. Esteban. 1985. Generation of a dominant8MDa deletionatthe leftterminus of vaccinia virus DNA. Pro.Natl. Acad. Sci. USA82:3365-3369.
12. Paez, E., and M. Esteban. 1984. Resistance ofvaccinia virus to interferon is relatedto aninterferencephenomenonbetween the virusandtheinterteronsystem.Virology 134:12-28.
13. Paez, E., and M. Esteban. 1985. Interferonpreventsthe
gener-ation ofspontaneous deletions at the left terminus ofvaccinia virus DNA.J.Virol. 56:75-84.
14. Paez, E., and M. Esteban. 1985. Interferon inhibits marker rescueof vaccinia virus.J. InterferonRes. 5:247-256. 15. Perucho, M.,andM. Esteban. 1985. Inhibitory effect of
inter-feron on thegeneticandoncogenic transformation byviral and cellulargenes.J. Virol. 54:229-232.
16. Rodriguez, J.F., R. Janeczko, and M. Esteban. 1985. Isolation and characterization ofneutralizing monoclonal antibodies to vaccinia virus. J. Virol. 56:482-488.
17. Rodriguez, J. F., E. Paez, and M. Esteban. 1987. A14,000-Mr envelope protein of vacciniavirus is involved in cell fusionand formscovalently linkedtrimers. J.Virol.61:395-404.
18. Roizman, B. 1%2. Polykaryocytosis induced by viruses. Proc. Natl. Acad. Sci.USA48:228-234.
19. Ruyechan, W. T., L. S. Morse,D. M.Knipe,andB. Roizman. 1979. Molecular geneticsofherpes simplex virus.II.Mappingof themajor viral glycoproteins and of thegenetic loci specifying the social behavior ofinfected cells.J.Virol. 29:677-697. 20. Stern, W., and S. Dales. 1976. Biogenesis of vaccinia. Isolation
and characterization ofa surface component that elicits anti-body suppressing infectivity and cell-cell fusion. Virology 75: 232-241.
21. Zavagno, G., B. Jaffe, and M. Esteban. 1987. Role of prosta-glandinsandnonsteroidalanti-inflammatory drugs in the patho-genicityofvaccinia virus. J. Gen. Virol.68:593-600.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG.1.virustwo-dimensionalAbbreviations:proteins[35S]methionine-labeledatImmunoblotimmunoperoxidasemutant the Alterations in the electrophoretic mobilities of several structural vaccinia virus proteins during virus persistence](https://thumb-us.123doks.com/thumbv2/123dok_us/1356171.89162/4.612.59.557.375.613/dimensionalabbreviations-methionine-labeledatimmunoblotimmunoperoxidasemutant-alterations-electrophoretic-mobilities-structural-persistence.webp)