0022-538X/92/031602-08$02.00/0
Copyright © 1992, American Society for Microbiology
Sequence
and Structural Elements That Contribute to Efficient
Encephalomyocarditis
Virus RNA Translation
GREGORY M. DUKE, MICHAEL A. HOFFMAN,AND ANN C. PALMENBERG*
Institutefor Molecular Virology and Department of Veterinary Science, University of Wisconsin,
Madison, Wisconsin53706
Received 23August1991/Accepted25November 1991
The nucleotide sequence of the5' nontranslatedregionofencephalomyocarditisvirus(EMCV-Rueckert)was
determined, and a consensus RNA structural model for this sequence (850 bases) and three other
poly(C)-containing cardioviruses (mengovirus, EMCV-B, and EMCV-D) was created through reiterative use of a
minimum-free-energyfolding algorithm.The RNA elements within thisregion which contribute to translation
ofEMCVproteinsweremappedin cell-free reactionsprogrammed with cDNA-derived RNAtranscripts. The data provide evidence that stem-loop motifs I, J and K, formed by viral bases 451 to 785, are important components of cap-independent translation. In contrast to other reports, a minimal role for stem-loop H (bases 406 to444) in translational activity is indicated. Small 5' nontranslated region fragments (bases 667 to 797) containing the J and K motifs proved strong competitive inhibitors when added to cell-free reactions
programmedwithexogenous capped or uncapped mRNAs. Theputativesequestering of required translational factors by this segment clearly contributes to translational activity, but also suggests a possiblecompetitive
mechanism for the downregulation of host proteinsynthesis duringviral infection.
Unlike most eucaryotic mRNAs, the positive-strand ge-nomesof picornavirusesareuncapped andhave verylong5'
nontranslated regions (600 to 1,300 bases), which contain extensive secondary structure and multiple noninitiating AUG triplets(9, 36). These uncommonfeatures, combined with the knowledge thatpoliovirus infection leads to inhibi-tion ofcap-dependenttranslationof normalcellularmRNAs, have long warrantedsuspicion about alternate translational mechanisms (9, 36). Ourrecent reports and those of others
have indeed confirmed that 5' non-translated regions (5'
NTRs) ofpoliovirus, encephalomyocarditis virus (EMCV) and foot-and-mouth disease viruspromote cap-independent translation initiation, whereby ribosome bindingoccurs via internalentry ontothe 5'NTRwithout scanning fromthe 5'
terminus.Internalentry,relativetothe 5'terminus,hasbeen clearly demonstrated with monocistronic and dicistronic cDNA-derived transcript RNAs (5, 10-12, 15, 16, 30), as
well as with virion RNA (36). The binding site has been
variously describedas aribosomal landingpad, translational
enhancer, orinternalribosomeentry site.
During poliovirus infection, cap-independent translation occursinconjunction with viral2Aprotease-induced
degra-dation of cellular proteinp220, a componentofthe
eucary-otic initiation factor 4F (eIF-4F) cap-binding complex. The
synergistic processes presumably promote viral translation while down regulating that of the host (18). Foot-and-mouth
disease virus infection also induces degradation of p220,
althoughadifferent viralprotease (Lrather than 2A) may be
responsible (3). With EMCV, the translational shift from host toviralmRNAislikewisevery proficient, but seems to resultfrom ahighly effective competition for cellular
trans-lational components rather than selective degradation of
p220(17, 21, 39).
The translational efficiency of EMCV RNA is especially
*Correspondingauthor.
tPresent address: Department of Microbiology/Immunology, StanfordUniversity, Stanford, CA 94305-5402.
apparent incell-freeextracts,inwhich 8to10copiesof viral
polyprotein aretypically obtained from eachtemplate (37).
We have shown thattheactivenature of these templates is
notuniqueto expression of viral codingsequences, but is a property that canbe qualitatively transferred to nearly any
eucaryotic cistron through ligation with viral fragments containing the internal ribosome entrysite element (12,27).
The method has been widely used to obtain high-level protein expression in a variety of eucaryotic cells (5, 41). The transferred EMCV element usually includes bases 335 to 837 (translation begins at base 834) (5, 12), although deletionanalyses with dicistronic chimericmRNAs suggest that bases 403 to 811 may provide the core fragment for efficient translation (6, 14).
In this report, EMCV enhancer sequences are further
defined through nested monocistronic RNAtranscripts de-rived from cDNA clones and expressed in reticulocyte lysates. Linker insertion and deletion mapping show that
translational functionality is achieved apparently through concertedcontributions of severalstructuresandsequences
within the required region. Moreover, the potent inhibitory effects ofoneparticular stem-loop element (theJ-Kdomain, bases680 to785)onexogenously translating capped mRNAs
suggest further biological involvement in putative
host-protein shutoff mechanisms during naturalviral infection.
MATERIALS AND METHODS
Sequence determination. EMCV Rueckert strain
(EMCV-R) was passaged in HeLa cells, and vRNA was
purified as described previously (33). Comprehensive se-quence data for the 5' noncoding region of EMCV-Rwere
generatedbythreeseparatetechniques: (i) primerextension
dideoxynucleotide reactions withreverse transcriptase,
us-ing EMCV vRNA templates and 5'-labeled
(32p
or35S)
oligonucleotide primers; (ii) Maxam-Gilbert determinations ofcDNAssynthesized from EMCVvRNA[poly(C) region];and(iii) primer extension dideoxynucleotide reactions with Klenow polymerase, using EMCV cDNA-containing plas-1602
on November 10, 2019 by guest
http://jvi.asm.org/
mids as templates (20, 25, 38). All nucleotides were
con-firmed by multiple determinations.
Aconsensus RNAsecondary structure for the 5'
noncod-ing regions of poly(C)-containnoncod-ingcardioviruses was created
through reiterativeuseof the folding algorithm of Zuker and
Stiegler (42), modified to accept large (e.g., >2,500
nucleo-tides) data fields. The minimum-free-energy structures for
the 5'-most 2,500 nucleotides fromEMCV-R, mengovirus-M
(24), EMCV-D, and EMCV-B (1, 2), were calculated
sepa-rately and then compared. Thefolding energies were those
defined by Freier et al. (7) and averaged to about -1,180
kcal/mol (ca. -4,937 kJ/mol) for each sequence. Elements
commontoalldeterminationswereretained, and intervening
segments wererefolded untilaconsensus structure maximiz-ing total paired nucleotides in analogous configurations for all strains was achieved. The model was adjusted until the
calculated minimum freeenergy didnotdiffer by more than
5%fromanyof the optimum individual sequencesolutions. Construction of cDNA. Standard DNA cloning reactions (34)wereused, and Escherichia coli JM101wasthebacterial hostfor propagation of recombinantDNA.Thetranscription
vector Bluescribe M13+ was purchased from Stratagene. Plasmid pEA1, a subclone of pE13 (4), contained EMCV bases 335 to 4229 inserted between the EcoRI and BamHI sites of Bluescribe M13+. This construction isvery similar
to plasmid pESA1 (28), except for the choice of vector sequences. The pBalAl plasmids (numbered 1 to 14) were
derived frompEAl andcontainedanested series of
sequen-tial deletions (fromprogressive Bal 31 reactions)within the 5' noncoding segments of the viral cDNA. Insertion
muta-tionscontaining EcoRI restriction sites were made at natu-rallyoccurring MaeII restriction sites in the viral5' noncod-ing region ofplasmidpEAL.The sequence at theinsertion is
ACGCGCGCGGAATTCGT (inserted sequences are under-lined). Additional 4-base insertionswerecreatedbyfillingin AvrII and HindIII restriction sites within the viral 5'
non-coding regionofpEAlwith Klenowfragment. An insertion-deletion plasmid was made byjoining (with EcoRI linkers)
sequencescleavedatKpnI(base701) and MaeII(base 776).
The construction of all plasmids was confirmed by nucleo-tide sequencing.
Transcription ofplasmid cDNA. Plasmid DNAs were lin-earizedbydigestionwithBamHIandusedastemplatesin T7
polymerase runoff transcription reactions containing
[a-32P]CTP
(13 mCi/mmol). Reactions were terminated byaddition of EDTA (to 10 mM), and the samples were extracted withphenol-chloroform, precipitatedwithethanol,
and then resuspended in water. Reaction efficiency was monitored by scintillationcounting afterabsorptionof sam-ples to Whatman DE81 filters. To determine the transcript
size andpurity, sample aliquotswere denaturedby incuba-tionat50°Cfor1hin thepresenceof1 Mdeionizedglyoxal
and 10 mM Na2HPO4 (pH 6.5) and then loaded onto 1%
agarose gels. Afterelectrophoresis (1 V/cm for11 h), dried
gelswerescanned forradioactivitywith anAmbis radiation detector. The radioactivity in full-length
transcript
bands relative tothatfrom the total samplewas used toestimatethe percentage offull-length RNA synthesized in the
reac-tion.
Invitroproteinsynthesis. Cell-free translation reactions in reticulocyte extracts were performed as described
previ-ously (29). Typically,vRNA(1to3,ug)or
transcript
RNA(1to3 ,ug)wasusedtodirectprotein synthesisinreactions(15
to30,ul) radiolabeledwith
[35S]methionine
(1 ,uCi/u). After60to90 minof incubationat30°C,reactionswere
stopped by
theaddition ofcycloheximide andpancreatic
RNase(to0.361
121
181
241
301 361
421
481
541
601
661
721
781
841
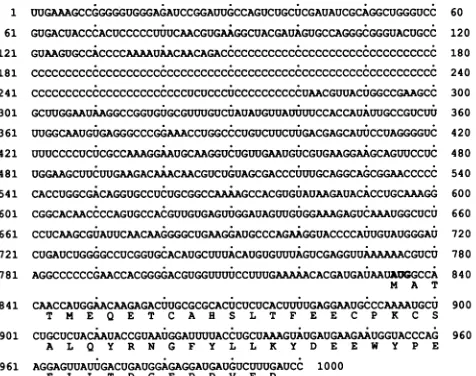
901
UUGAAAGCCGGGGGUGGGAGAUCCGGAUUGCCAGUCUGCUCGAUAUCGCAGGCUGGGUCC GUGACUACCCACUCCCCCUWUCAACGUGAAGGCUACGAUAGUGCCAGGGCGGGUACUGCC
GUAAGUGCCACCCCAAAAUAACAACAGACCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCCC
cCCCCCCCCcCCCCCCCcCCCCCCCCCCCCCCCCCCCCCCCCCCCCCcCcCCCCCCCCCC CCCCCCCCCCCCCCCCCCCCCCCUCUCCCUCCCCCCCCCCUAACGUUACUGGCCGAAGCC GCUUGGAAUAAGGCCGGUGUGCGUUUGUCUAUAUGUUAUWWUCCACCAUAUUGCCGUCUU
UUGGCAAUGUGAGGGCCCGGAAACCUGGCCCUGUCUUCUUGACGAGCAUUCCUAGGGGUC UUUCCCCUCUCGCCAAAGGAAUGCAAGGUCUGUUGAAUGUCGUGAAGGAAGCAGUUCCUC
UGGAAGCUUCUUGAAGACAAACAACGUCUGUAGCGACCCUWUGCAGGCAGCGGAACCCCC
CACCUGGCGACAGGUGCCUCUGCGGCCAAAAGCCACGUGUAUAAGAUACACCUGCAAAGG
CGGCACAACCCCAGUGCCACGUUGUGAGUWGGAUAGUUGUGGAAAGAGUCAAAUGGCUCU
CCUCAAGCGUAUUCAACAAGGGGCUGAAGGAUGCCCAGAAGGUACCCCAWUGUAUGGGAU CUGAUCUGGGGCCUCGGUGCACAUGCUUUACAUGUGUUUAGUCGAGGUUAAAAAACGUCU
AGGCCCCCCGAACCACGGGGACGUGGUUUUCCUUUGAAAAACACGAUGAUAAUAUGCCA M A T
CAACCAUGGAACAAGAGACWUGCGCGCACUCUCUCACUUWUGAGGAAUGCCCAAAAUGCU
T M E Q E T C A H S L T F E E C P K C S
CUGCUCUACAAUACCGUAAUGGAUUUUACCUGCUAAAGUAUGAUGAAGAAUGGUACCCAG
AL Q Y R N G F Y L L K Y D E E W Y P E
96AGAUA.AUAGAGGA UUUUAC 1000
60
120
180
240
300
360
420
480
540
600
660
720
780
840
900
960
9 61 AGGAGUUAW~GACUGAUGG'AGAGGAUGAUGUCUUUGAUCC 100 0
E L L T D G E D D V F D
FIG. 1. Nucleotide sequence of the5'-most1,000nucleotides of EMCV-R. Thepolyproteinreadingframebeginsatbase834,and the poly(C)tractcontains 132pyrimidines(C115UCUCCCUC10). mg/ml
each).
Samples (3RI)
wereanalyzed by
sodiumdodecyl
sulfate-polyacrylamide
(Maizel) gel
electrophoresis
(SDS-PAGE)
(26), and labeledproteins
were visualizedby
autoradiography
of the driedgel.
Incorporation
of [35S]me-thionine into acid-insolubleproducts
wasperformed
as de-scribedpreviously
(22).Nucleotide sequence accession number. The GenBank/
EMBL accession number for this sequence
(complete
EMCV-R genome)is M81861. RESULTS
Sequence and structure model for EMCV-R. The nucleo-tidesequenceof the 5'-most
1,000
nucleotides of EMCV-R ispresented
inFig.
1. Thedatacorrectminordiscrepancies
in theprevious
determination (23, 25) and complete the se-quence for this strain. Whenpoly(C) length
differences arediscounted,
the 5' NTRof EMCV-R shares95.2%nucleotideidentity with those of EMCV-B and EMCV-D
(1,
2)
and 92.7%identity
with the sameregion
frommengovirus-M
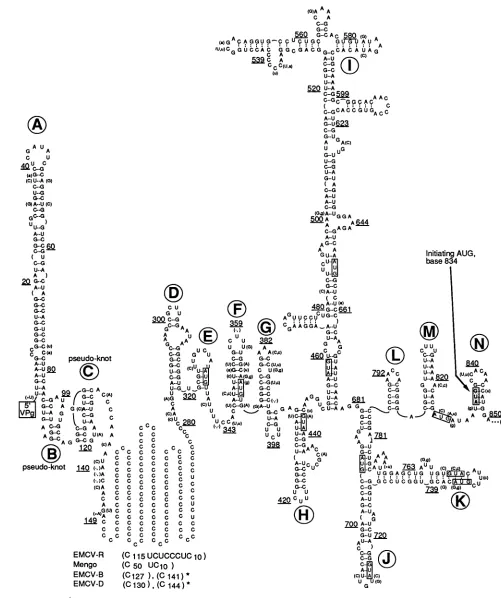
(24).AconsensusRNA
secondary
structurefor the 5' ends of four cardiovirusRNAs,
highlighting
straindifferences,
ispresented in Fig. 2. The model contains many elements similartothose
suggested
frompartial
sequencealignments
(13, 31), but it also differs in three
regards.
First,
thesedominant
patterns
were derived fromminimum-free-energy
calculations which consideredsimultaneously
allpossible
pairing
options
forlarge
segments of thegenome(the
5'-most2,500
nucleotides).
Second,
thepoly(C)
tracts and their discontinuitieswerenotexcludedfromconsiderationby
thealgorithm,
but were allowed to(potentially)
participate
in basepairing,
aslong
asthese interactions couldcontributeto thelowest-energy
solutions.Third,
calculations from thefinalconsensusmodel allowedamaximumdeviation of
only
5% from theoptimum folding
solution foreach of thefourparticipating
(2,500-base)
sequences. This model isconsis-tent with most available data
concerning
enzyme-sensitive
sites,
exposed loops,
etc.,for relatedisolates of EMCV(6,
13, 31,40).The structure suggests a linear series of 5'
noncoding
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.324.562.81.269.2]G AUA
C U
40U C
(a)G--C
(C)U--A(G)
C--G U--G
G--C
(0)A--U(C) C--G GC--G U)U--G A--U G--C
C--G
( UC -G
C--G U--A
A)
G2-C
A--U
(A
G--C
G--C G--C U--A G--C G--U G--C
G-C
cG--C(u)
C C (a)
G--c pseudo-knot
A--U A--U G--C
U--A A G A
(U--A AAC&
G-U U t U-A C
A C-G C-G A A
G--C G-c U (A) A
ACAG(G--CU)AG
0 (c)A
A C C
C C
(C)U C C
pseudo-knot 140 (-.)(A C C
(-)A C C
(-(C C C
(C)A C C
A C C
C C C
A C C
G(U)C0 C (+A)A C C
c C C
C C C
C C
EMCV-R (C
Mengo (C
EMCV-B (C
EMCV-D (C c c c
c
c c c
c
c c c c c c c c
c
c c c
rC
()A AA
C A
C-G
G-C
560 G-CAC 580 (G) A
(a)GA9AGGUG-I; 9U-riiG9; GLT~TAU
Mu) G6 AC GCG,AC,G,G-C AG
CAC .C(U.:CG A
U-A
-GCGG AC c
C-GCACC GU&ACC A-U
G-U62
C-G
G--UG
A GAPC
G-5U
U-G
C-G
U-A G-U
( A
C-G
A-U
A-U C-G
(G.g)A-UGA
SOA AGA
C-GG A-U
A G--C
A A
GU--A
U-G
C-G
G-U (Ac
c 480A--U{)
G U 8GGC661
300C-G A UUCC-UG A
GOGAU°U G-U G)G s(AU G
A-- A(-9 CGG- G-C G
Au u~~~~~00 A4
AA c u 382 U-~AAG
-G - U A -U (GaAC, G AU G
C UC--G(GucUCU UA)(GA- Mc)aCAU CU- - C GC-C U A C-Geu G-- AM- U--G
CGCC U UC-G(A) G C-GuA G--C
UC-GCCa)G--C(u)u C G--Croa) AA
'M'C(QU--
C--G U--AC--GG A~~~~~~~~~--M2
GC-CCCU20C G-C
C C (C.C)U-G G--C U-A
CACG U () 32 U-A c
c AA ) u (UCU-()()A. GCA 9 C
C-- UC U AU(OAA GG
CC--G--C UGC UA C-G-- AG--CAQU--GCA --C C 70 (-
-(C (CU } (A)-Cu C U.-(Ge) (A) G
C-G G c).A(I C-- A--U A A
A--U C-jCU-G
Ag)C--GUAG-c ccc UU(CAA.CG-G--CA
c (c) A UG-C A
c CCC GC-..-- G-CAAu
(
C) ccU U- (U)G--~~~~~C-IUlG-(A),CAU u
'C C C -CUGA
C C C U-AC440GUGCC
C c C UCU -G
C C C C-GAG
CC C c
C2
u G A-jUAC C CCA-C A--UlA
C C C C ~~AU(
--Cccc~ ~ ~ ~ ~ G- GO
C
C-C C CUG-C
C C C C G-U720
C,CC
~~~~~~~~~~~~~~AU-A_
115 UCUCCCUC10)
50 UC10)
27 ),(C141)* 130), (C144)*
E)cCG C-G
(C)U-I(C)
U (G)
G
* independentdeterminationsofsequence
FIG. 2. Consensus secondary structurefor 5' NTRsoffourrelatedcardioviruses. The sequence shown is for EMCV-R. Changes in
mengovirus and other EMCV strains (EMCV-B and EMCV-D) aregiven parentheticallyas capital and lowercase letters, respectively.
Stem-loopfeatures arelettered consecutively, and AUG sequences(EMCV-R)areboxed.
Initiating AUG, base834
L0c UUA82 CA84c A
- U--A
C U--A (~C A U--A~a PCAC A
i--C G A(Cx) G-C
-G G--C GC(u)
-G U--A A
-G G-C U
,-G C--G C (U--GO
A u Au) A G 0
kA> AUA A..(3.)
(G.g)
763AUU (C) cA
;AGCUG UG IGU AICU
: : U(C)
UCGGUGC A U
Z3
(G))~
,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.43.544.86.684.2]T7 RNA Transcripts.
E-Al p3alAl -14
4tk
-3 kb
-6kb
FIG. 3. RNA transcript integrity. T7 transcript samples from pBalAl-1 to pBalAl-14 wereprepared, denatured, fractionatedby electrophoresis, and detected asdescribed in Materialsand Meth-ods.
stem-loop motifs (Fig. 2, letters A to N), within which at least65% of the nucleotides [exclusive of the poly(C) tract] participate in base pairing. Surprisingly, the poly(C) region did not make a major contribution to the most favorable configuration ofany sequence (5'-most 2,500 bases), even
whenallowed topair with distant regions. The pseudoknot motifs in B and C differ from apublished structure of this region (40), but thisarrangementis energeticallyvery
favor-able and has been independently suggested by algorithms sensitivetopseudoknot configurations (31a).
Among the considered viral strains, transitions (e.g., U-*CorA--G)accountfor60% of thesequence variation.
More than two-thirds of theseexchanges occurinstemsand
cause little (if any) perturbation of the common structure. The model has 10 examples of double substitutions within paired bases, althoughsomeof thesecompensating changes are separated by more than 40 to50 nucleotides within the
sequence(seestemsA, F, H, I, and K). Transversions (e.g., U-iAorG-*C), insertions,anddeletionsaremore common
totheunpaired regions (77%) andaretypicallyatthetopsof stemsorinbulge-loops between pairedstems.
Translation of RNAs containing 5' NTR mutations. To defineimportant translational elements within theEMCV5' NTR, weconstructedaseries of cDNAplasmids containing progressive 5'--3' deletions. The constructions, designated pBalAl-1 topBalAl-14, each includedcodingsequencesfor the amino-terminal half of the polyprotein open reading frame (1,131 codons). RNA transcripts from these cDNAs
wereprepared in parallel and assayed by gel electrophoresis
(Fig. 3) to ensure minimum variation in the amount of full-length material (ca. 70%). Thetranscripts wereused to
program reticulocyte lysate reactions, takingcarethateach sample received the same amount of RNA (2.6 ,ug), at a
concentration (86 ,ug/ml) previously determined to give optimal, saturating protein synthesis.
The pEAl transcripts which began at viral base 335
A.
d
LI|1AI|
1B
1C
I|
1DI2AI12B
Al
virlon RNA pEAl
pBalAl
1-14
B.
400c 3007
x
v-2
200-21
CIO
"6
- 363 387
406 416
443 454 466 485
505 513 546
620
776-L
802-CPM Incorporated
perul of lysate
UG(834) (x 103) 302 (1 00%)
1 253 (84%)
^, 251 (83%)
* 254 (84%h)
.4 230 (76%)
"= 95 (31%)
.= 64 (21%°h)
.= 82 (27%)
,= 169 (56%)
,= 76 (25%)
,~ 14 (5%)
,= 18 (6%)
.= 21 (7%)
,= 23 (8%)
.= 45 (15%)
.= 105 (35%)
-\&-_406
l-_466
416 1485 ----802
AUG (base 834)
200 460 600 800 1000
5' most viral nucleotide
FIG. 4. 5' deletion mutations. (A)EMCV virion RNA(1 ,ug)or T7 RNAtranscripts (2.6 ,g)asshown inFig.3[pEA1andpBalAl-1
topBalAl-14]wereusedtoprogramtranslation reactions (30RI)in rabbit reticulocyte extracts containing [35S]methionine (1 ,uCi/,ul). After 90 min at 30°C, triplicate sampleswere removed(2 ,ul)and quantitated for trichloroacetic acid-insoluble radioactivity. Aver-agedresults(standard deviation, <2%)arepresentedascountsper minute incorporated per microliter oflysate and also as percent
incorporation relative to virion RNA. (B) Data from panel A displayed graphically.
directedprotein synthesistoabout 84% of the level obtained withvirion RNA. However, this is probably nota
substan-tivedifference intranslational efficiency (6), sincethe trun-catedcoding regions of the transcripts containonly halfas manymethionines(24versus51)asintactpolyproteins.The pEAl incorporationrepresents about threetofive copiesof protein product from each transcript template (pEAl), a
significant activity consideringthe saturatingconcentrations ofRNA,andprovidesthetruebaseline forcomparisonwith othertranscripts.
Engineered deletionof thenext52 viral bases(tobase387)
gave transcript activities similar to those of pEAl and confirmedpreviousobservations thatstem-loops A to G do notcontributesignificantlytoEMCV translational function-ality (12, 36).Furthertruncations, however,whichimpinged
on structure H, showed significantdecline in incorporation (Fig. 4B). Computer-generated structural models suggest thattranscripts shortened within this region (i.e., beginning
pEAl (1131 codons)
on November 10, 2019 by guest
http://jvi.asm.org/
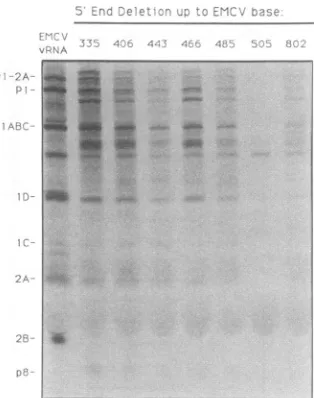
[image:4.612.97.275.80.316.2] [image:4.612.325.560.82.442.2]J n--rc: .ee et -.. 'T ..
--2IA
P'-FIG. 5. Proteinproducedin cellextractsis viral. Samplesfrom thetranslation extracts inFig.4wereprocessedwithexogenous3C proteaseandanalyzed by SDS-PAGE andautoradiography.
with bases 406to450)may assumestablepairing
configura-tionsquitedissimilartotheconsensus structure(notshown).
The predictedalterations would disrupt stem-loopI as well
as remaining H fragment, but it is also anticipated that
slightly larger deletions (i.e., to bases 451 to 500) might
restore most of the I configuration (see also reference 6).
Indeed, complete removal ofthe H domain (to base 466)
from thepBalAl-7transcript partiallyreestablished transla-tion in areproducible manner. This level ofincorporation
was alwayslower(e.g., 56 versus 84%)than thatachieved with pEA1, but clearly indicates thatintactHsequencesare not anabsolute requirementfortranslationalactivity.
Tran-scriptsbeginningwith base505,513, 546,or620werepoorly
translated in the reactions, although very shorttranscripts
with only 32 5' NTR bases had somewhat better activity.
These highly abbreviated RNAs (e.g., beginningwith base
802) have been suggested to initiate synthesis in a
5'-end-dependentmannerunrelatedtointernalribosome entrysite function (15).
The translation data presented in Fig. 4 were obtained withoptimal concentrations of added templateRNA.
How-ever, the differential levels of incorporation were clearly
dependent on template sequence, not concentration,
be-causesimilar resultswerealsoobtained undersubsaturating
RNA transcript conditions (e.g., 50 ,ug/ml), as long as
compared samples contained similar molar equivalents of
template(multiple determinations notshown).
To confirmthatprotein synthesis bythe transcript series
was uniformly initiating at the same AUG (base 834), we
processed selected product samples with exogenous,
unla-beled 3C protease and then analyzed them by SDS-PAGE (Fig. 5). Although thepatternsvaried in intensity according
to incorporation, all lanes showed bands characteristic of L-P1-2AB* sequences and there was no clear evidence for
significantlevels ofinitiationatalternate AUG triplets.Even
the shortest transcripts, whicharemissingmajor portions of
the internal ribosome entry site, seemed to direct most
residual synthesis towards the correct, in-frame AUG and
hence reinforcefindingsthat sequencesand/orstructures in the immediate vicinity of this codon probably play strong
selectiveroles in synthesis initiation(15).
pEAI (1131codons)
A 1B IC
I
1D I 2A126B|
Relati' inco(p
nomutation UG834)
335 415
Hin tillin
335 488
EcoRIlinker
ln"rfit
335 5
EcoRIlinker
insertion
335
EcoRIlinker
insertion
335 77
ive35-Met wrporation
ercent)
100
88
90
85
15
8
777 EcoRIlinker
insertion/deletlon
335
A4
701-776 2
FIG. 6. Localinsertion and deletion mutations. Mutationswere
created in pEAl through restriction enzyme digest (AvrII at base 415;HindIl atbase488;MaeIIatbases506, 577, and777;orKpnI at base 701) followed by EcoRI linker insertion, filling in with Klenowpolymerase, orsequentialdigestion (with KpnIandMaeII) and EcoRI linker insertion. T7 RNAtranscriptssynthesized from these cDNAs(1.6to1.7 jig)weretranslated inreticulocyte lysates (15 iul) as in Fig. 4, and protein synthesis was quantitated by trichloroacetic acid-insoluble radioactivity. Presented values are relative to unmutated pEAl transcript incorporation (ca. 250,000 cpm/,ul),whichwas set at100o.
The deletion experiments placed the 5' boundary for minimum translational activity within the lower regions of
stem-loop I (roughly between bases 466 and 505). Local
perturbations affectingthisregionwereexamined with
engi-neeredlinker insertions orinternaldeletions (Fig.6).
Four-base insertionsatposition415(stem H)or488(lowerstemof I)decreased translation of the downstream cistron by only
10 to12%, consistent with the idea thatanintactHelement
and the lower portion of stem-loop I are not required for translational functionality. Since deletions past base 485 gave virtually inactive templates (Fig. 4), it was surprising
that insertions creatinganextended bulge loopatbase 506
were quite effective templates; this suggests that sizable alterationsin the middle ofstem I mayalso betolerable, if
stemintegrity is generally maintained.In contrast, insertion
at position 577, which disrupts the top of this stem (from bases527 to591), strongly decreased translationalactivity. TheJ-K domain also contributestoefficient translational function. Deletions within thisregion (e.g., bases701 to776)
or insertional augmentation near the A-rich bulge loop at
base 777 significantly decreased protein synthesis (Fig. 6). The detrimental effects occurredwithout alteration of
(pre-dicted) base pairs within adjacentstems (H, I, L, M, etc.) and are consistent with data implicating this region as a required translational element and possible protein-binding site(35).
Translation competition assays. Cardiovirus RNAs have
fj,
e.-2...
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.341.532.80.347.2] [image:5.612.102.259.81.280.2]Re (p Pr Fh (a Competing RNA
Fragment
(none)
335 '848
elative Incorporation lesspronounced with thefragment frombases 335 to775or ercent) of 35S-Met in whenthe competitor includeda 13-base insertion at base 777 resence of Competing RNA (within J-K). The fragment from bases 335 to 619 had almost
Template RNA no inhibitory effect, which makes it unlikely that the ob-IVRNA B-globin pEAl served results were a nonspecific consequence of added
apped) (capped) (EMCV) RNA.Therelativeinhibitoryactivityof thetested fragments was also evident at higher and lower molar excess of 100 100 100
competitor
RNAs(e.g.,
molar ratios of1,
2.5,
and10)
and confirma dependence on sequence, rather thanconcentra-36 12 42 tion (all data not shown).
620-848 620-797
667-797
335 619
335 775
EcoRi hiker
whieron
335
%-7
848EcoRt hiker
335 77
1.48
FIG. 7. Translation competition scripts containing the indicated El added toreticulocyte reactions (15 excess(oratwo- tothreefold molar overthetemplate RNA. The templat (FHV) RNA (1.2 ,ug), cellular 3-glo
T7 transcripts (1.8 ,ug). Synthesizt
trichloroacetic acid-insoluble radioa tion at 30°C. The results (average ( incorporation (percent) relative to s tor RNA(typically, for FHV300,00 125,000cpm/,ul=100%,andfor pEA results are reproducible to a stand done.
been demonstrated to be effit eucaryotic mRNAs in cell-fre Regions within the EMCV 5'
proteinswereinvestigated intra RNAtranscriptscontainingbase thereofwereaddedtoreticulocy
lation-competent mRNAs from
,B-globin,orintactpEAl (Fig.7)
sequences wereomitted from th interference fromsynthesizedv assays on initial translation evc
Since the molar ratios of the
deliberately kept low (molarral
proteinsynthesisto50% of thel1
offragment wasindicative ofst
Theassaysrevealed that intac and all smaller segments contai
reasonably effective competito withinthesefragmentscenters c
predictions suggest that these inhibitory RNAs and make it lik
teringofrequiredtranslational c
ble for this effect. Both capp
3-globin)
and uncapped(pEA1)competition, although
1-globin
tory than pEAl and could typimolarratios offragment. With
-37 13 42
cc 'OA AR
DISCUSSION
Do Am Translation is initiated on
picornavirus
RNA bycap-nd 44 52 independent, internal entry ofribosomes onto the
5'
NTR.Previous analyses of EMCV bases involved in efficient 88 95 114 translation were carried out predominantly with dicistronic mRNAs (14). We have further defined the required RNA 71 72 125 sequences by deletion and insertion analysis with
monocis-tronic mRNAs. Although our results generally agree with
13
<5
20 those of Jang and Wimmer (14), they do call into question therole of stem-loopH. These authors suggestedthat integrity 68 62 105 of H correlates with translational efficiency of the down-streamcistron in dicistronicmRNAs and also with theability
MCV 5' NTR sequences were ofa57-kDa cellular
protein
tobindtothisspecific
structure.
RI)
ina four- to sixfold molar However, their tested deletions within this region were less excesswithP-globin
reactions) comprehensive than our pEAlBal series, and we have Le RNAs were:flockhouse virus createdatleastonetranscriptin which this entire domainis obin mRNA(0.8Rg),
andpEAl cleanly removed(deletiontobase466) and yet the RNAstill ed protein was quantitated by translates with reasonable efficiency (70% of that ofpEA1).ictivity after 60 min of incuba- We can onlyspeculate about the reasons for these exper-ofduplicate samples) represent imentaldiscrepancies. However, it might be considered that
camples
withnoaddedcompeti- monocistronic and dicistronic contexts could place entirely)0
cpm25
l = 10%, for1
T-globin
different constraints
on the
ability
of the
5'
NTR
to
initiate
lard deviation of10%.
nd, not translation. Indeed, Jang et al. showed previously (12) that some EMCV deletion mutants are moreeffective inmono-cistronic than dimono-cistronic contexts. Moreover, it ispossible
thatengineered constructions which only partiallyremove a
localRNAmotif have unanticipated configuration effectson
cient competitors of other adjacent structural elements. Thus, certain transcripts,
ei-e synthesis reactions (32). thermonocistronic ordicistronic, could potentially exhibit NTR that might sequester altered activities independent of the deleted sequences.
,nslation competitionassays. Computer-aided RNA folding supports this idea and sug-s335to848orsubfragments geststhat thedeletions within stem-loopH (topositions421 (te extractscontainingtrans- and 425) performed by Jang and Wimmer (14) might have
i flock house virus, rabbit disturbed the natural folding of adjacentstemI, ina manner
.Except for14bases,coding analogousto ourpEAlBal constructions which beginatbase
etestedfragmentstoreduce 406, 416, or443. Insertions specific to stem-loop H (linker iralproteinand tofocus the insertion atbase415)orremoval ofHwithout disruption of
ents rather thanelongation. I(deletion ofbase 466) allowedus to independently assess
competing segments were these motifs and to map the 5'-most required translational tio of2 to 6), depression of elementtothelowerregionsofstemIrather thanstem-loop
evelachieved in the absence H. trongcompetition.
St 5' NTRs(bases 335to848)
ining bases 667 to 797were trs. The common sequence Dn the J-Kregion. Computer
stems are conserved in all ;ely thatJ-K-specific
seques-components (35)is
responsi-ted
(flock house virus andtemplateswere sensitiveto RNA was much less refrac-ically be inhibited by lower all templates, inhibitionwas
Our dataare in agreement with those of Shih etal. (36), whofound that cDNAfragmentshybridizingtonucleotides
420 to 449 caused only a slight translational inhibition of EMCVvRNA, whereas cDNAshybridizingbetween
nucle-otides 450 and 834 caused a high degree of inhibition. Correlative results were also obtained with the related
foot-and-mouth disease virus, in which the RNA
region
controlling translation appears to maintain the same basic structuralpatternasthatof EMCV(31).Thefoot-and-mouth diseasevirusanalogof EMCVstemHwasmutatedsuch that the 57-kDaprotein boundvery weakly, yet translationwasdecreasedonly50% whencomparedwith thatofunmutated transcript RNA (16, 18a, 19). This lends support to our
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.71.289.83.312.2]conclusion that theintegrityofstemHdoesnot seemcrucial forproper translationalfunction.
We suggest instead that the J-K element and the ever-present stem-loop I play more critical roles in translation initiation. Independent of other 5' NTR regions, the J-K sequences (bases 667 to 797) were remarkably active in
competitive inhibitionassays. It is notable that linker inser-tions instemsIand J-K(atbases 577and777,respectively)
both destroyed translational activity but showed quite dis-tinct effects when used as inhibitors in the competition
assays. The insertion at base 577 (stem I) was an active
competitor,whereas theinsertionatbase777(stem J-K)had lost inhibitory activity. The terminal loop of stem K
(CU-UUA,bases746to750)wasrecently reportedas anefficient
bindingsite foreucaryotic initiation factor eIF-2/2B (35), a
protein complex presumed to function in initiator AUG
recognition (8). Consistentwith ourdata, astrong
competi-tiveJ-K sequence-dependent sequesteringof these required translational elements may explain the overall
efficiency
ofEMCV
fragments
atdirecting
protein synthesis. Moreover,such interactions could provide a plausible mechanism by which EMCV vRNA may outcompete host mRNAs for ribosomesorinitiationfactors(eIF-2/2B?)andeffect shutoff ofcellularprotein synthesis during infection. We are
cur-rently examining the role ofpurified eIF-2 and eIF-2B on translational
activity
andspecifically
onJ-K-dependent inhi-bitionofexogenous mRNA translation.ACKNOWLEDGMENTS
We thank DingShih for critical readingof the manuscript and CornelisPleijfor corroborative dataconcerningpseudoknot struc-turesBandC.
This workwassupportedbyPublic HealthService grant AI-17331 from the NIH.
REFERENCES
1. Bae, Y. S., H. M. Eun, and J. W. Yoon. 1989. Molecular identification ofadiabetogenicviral gene. Diabetes 38:316-320. 2. Cohen,S.H.,R. K.Naviaux,K. M. V.Brink,andG.W.Jordan. 1988. Comparison ofthe nucleotide sequences ofdiabetogenic andnondiabetogenic encephalomyocarditisvirus.Virology166: 603-607.
3. Devaney,M.A.,V. N.Vakharia,R. E.Lloyd,E.Ehrenfeld,and M.J.Grubman.1988.Leaderproteinoffoot-and-mouth disease virus is required for cleavage of the p220 component of the cap-bindingprotein complex.J.Virol. 62:4407-4409.
4. Duke,G.M.,and A. C.Palmenberg.1989.Cloningand synthe-sis ofinfectious cardiovirus RNAs containing short, discrete poly(C)tracts. J.Virol. 63:1822-1826.
5. Elroy-Stein, O.,T.R.Fuerst,and B.Moss. 1989. Cap-indepen-dent translation ofmRNA conferred by encephalomyocarditis virus 5' sequence improves the performance of the vaccinia
virus/bacteriophage T7 hybrid expression system. Proc. Natl. Acad. Sci. USA86:6126-6130.
6. Evstafieva, A. G., T. Y. Ugarova, B. K. Chernov, and I. N.
Shatsky.1991. AcomplexRNAsequencedetermines the inter-nal initiation ofencephalomyocarditis virus RNA translation. NucleicAcids Res. 19:665-671.
7. Freier, S. M., R. Kierzek, J. A. Jaeger, N. Sugimoto, M. H. Caruthers, T. Neilson, and D. H. Turner. 1986. Improved free-energyparametersforpredictions ofRNAduplex stability. Proc.Natl. Acad. Sci. USA 83:9373-9377.
8. Hershey, J. W. B., R. Duncan, and M. B. Mathews. 1986. Introduction: mechanisms of translationalcontrol, p. 1-18. In M. B. Mathews(ed.), Translational control:current communi-cations in molecularbiology. Cold SpringHarborLaboratory, ColdSpring Harbor, N.Y.
9. Jackson, R. J. 1986.Adetailed kinetic analysis of thein vitro
synthesis and processingofencephalomyocarditis virus
prod-ucts. Virology149:114-127.
10. Jackson, R.J., M. T.Howell,andA. Kaminski. 1990. The novel mechanism of initiation of picornavirus RNA translation. TrendsBiochem. Sci. 15:477-483.
11. Jang, S. K., M. V. Davies, R. J. Kaufman, and E. Wimmer. 1989. Initiation of protein synthesis by the internal entry of ribosomes into the 5' nontranslated region of encephalomyo-carditis virus RNA in vivo. J. Virol. 63:1651-1660.
12. Jang,S. K., H.-G. Krausslich, M.J. H. Nicklin,G. M. Duke, A. C.Palmenberg, and E. Wimmer. 1988. A segment of the 5' nontranslated region ofencephalomyocarditis virus RNA di-rectsinternal entry of ribosomesduringinvitro translation. J. Virol.62:2636-2643.
13. Jang, S.K., T. Pestova, C. U. T.Hellen, G. W. Witherell, and E. Wimmer. 1990. Cap-independent translation of picornaviral RNAs: structure andfunction of the internal ribosomal entry site. Enzyme44:292-309.
14. Jang, S.K., and E. Wimmer. 1990. Cap-independent translation ofencephalomyocarditis virus RNA: structural elements of the internalribosomal entry site and involvement ofacellular57-kD RNA-bindingprotein. Genes Dev. 4:1560-1572.
15. Kaminski,A., M. T. Howell, and R. J. Jackson. 1990. Initiation ofencephalomyocarditis virus RNA translation: the authentic initiationsite isnotselectedbyascanning mechanism. EMBO J. 9:3753-3759.
16. Kuhn, R., N. Luz, and E. Beck. 1990. Functional analysis of the internal translation initiation site of foot-and-mouth disease virus. J. Virol. 64:4625-4631.
17. Lawrence, C., and R. E. Thach. 1974. Encephalomyocarditis virus infection of mouse plasmacytoma cells. I. Inhibition of cellularproteinsynthesis. J. Virol. 14:598-610.
18. Lloyd, R. E., H. Toyoda, D. Etchison, E. Wimmer, and E. Ehrenfeld. 1986. Cleavage of the cap binding proteincomplex polypeptide p220 is not effected by the second poliovirus protease2A. Virology 150:299-303.
18a.Luz,N.Personal communication.
19. Luz, N.,andE. Beck. 1990. A cellular 57kDaprotein bindsto tworegions of the internal translation initiation site of foot-and-mouth disease virus. FEBS Lett. 269:311-314.
20. Maxam, A.M., andW. Gilbert. 1980. Sequencing end-labeled DNA withbase-specific chemical cleavages. MethodsEnzymol. 65:499-560.
21. Mosenkis, J., S. Daniels-McQueen, S. Janovec, R. Duncan, J.W. B.Hershey, J. A.Grifo,W.C. Merrick, andR.E. Thach. 1985.Shutoff of hosttranslationbyencephalomyocarditis virus infection does notinvolvecleavage of the eucaryotic initiation factor 4Fpolypeptide that accompanies poliovirus infection. J. Virol. 54:643-645.
22. Palmenberg, A. C. 1982. In vitro synthesis and assembly of picornaviral capsid intermediate structures. J. Virol. 44:900-906.
23. Palmenberg,A.C. 1987.Comparative organization and genome
structure in picornaviruses, p. 25-34. In M. A. Brinton, and R. R. Rueckert (ed.), Positive strand RNA viruses. Alan R. Liss, Inc., NewYork.
24. Palmenberg,A.C.,andG. M.Duke. Unpublished data. 25. Palmenberg, A.C., E. M. Kirby, M. R. Janda, N. L. Drake,
G. M. Duke, K. F. Potratz, and M. S. Collett. 1984. The nucleotide anddeduced sequences of the encephalomyocarditis viral polyprotein coding region. Nucleic Acids Res. 12:2969-2985.
26. Palmenberg,A.C.,M. A.Pallansch, and R. R. Rueckert.1979. Protease requiredfor processing picornaviral coat protein re-sides in theviralreplicase gene. J. Virol. 32:770-778.
27. Parks, G. D., G. M. Duke, and A. C. Palmenberg. 1986. Encephalomyocarditis 3C protease: efficient cell-free expres-sion fromclones which link viral5'noncoding sequences to the P3region. J. Virol.60:376-384.
28. Parks, G.D., andA.C. Palmenberg. 1987. Site-specific muta-tionsat apicornavirus VP3/VP1 cleavage site disrupt in vitro processing and assembly of capsid precursors. J. Virol. 61: 3680-3687.
29. Pelham,H. R.B.,and R.J. Jackson. 1976. An efficient
on November 10, 2019 by guest
http://jvi.asm.org/
dependent translation system from reticulocyte lysates. Eur.J. Biochem. 67:247-256.
30. Pelletier, J., and N. Sonenberg. 1988. Internal initiation of translation of eukaryotic mRNA by a sequence derived from poliovirus RNA. Nature (London) 334:320-325.
31. Pilipenko, E. V.,V.M.Bfinov, B. K. Chernov,T.M.Dmitrieva, and V. I. Agol. 1989. Conservation of the secondary structure elements ofthe 5'-untranslated region of cardio- and aphthovi-rusRNAs. Nucleic Acids Res. 17:5701-5711.
31a.Pleij, C. Personal communication.
32. Rosen, H., G. DiSegni, andR. Kaempfer. 1982. Translational controlby messenger RNA competition for eukaryotic initiation factor 2. J. Biol. Chem. 257:946-952.
33. Rueckert, R. R., and M. A. Pallansch. 1981. Preparation and characterization of encephalomyocarditis virus. Methods En-zymol. 78:315-325.
34. Sambrook, J., E. F. Fritsch,and T.Maniatis. 1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, ColdSpringHarbor, N.Y.
35. Scheper, G. C., A. Thomas, and H. 0. Voorma. 1991. The 5' untranslated region of encephalomyocarditis virus contains a sequencefor very efficient binding of eukaryotic initiation factor eIF-2/2B. Biochim.Biophys. Acta 1089:220-226.
36. Shih, D. S.,I.-W. Park, C.L.Evans, J.M. Jaynes,and A. C. Palmenberg.1987.Effects of cDNA hybridizationontranslation
ofencephalomyocarditis virus RNA. J. Virol. 61:2033-2037. 37. Shih, D.S., C. T. Shih, D. Zimmern, R. R. Rueckert, and P.
Kaesberg. 1979. Translation of encephalomyocarditis virus RNAin reticulocyte lysates: kinetic analysis of theformation of virion proteins and a protein required for processing. J. Virol. 30:472-480.
38. Smith, A. J. H. 1980. DNA sequence analysis by primed synthesis. Methods Enzymol. 65:560-580.
39. Svitkin,Y.V., V. A.Ginevskaya, T. Y. Ugarova, and V. I. Agol. 1978. A cell-free model of the encephalomyocarditis virus-induced inhibition of host cellprotein synthesis. Virology87: 199-233.
40. Vartapetian, A. B., A. S. Mankin, E. V. Shripkin, K. M. Chumakov, V. D. Smirnov, and A. A. Bogdanov. 1983. The primary and secondary structure of the 5' end region of enceph-alomyocarditis virus RNA. A novel approach to sequencing long RNA molecules. Gene 26:189-195.
41. Zhou, Y., T. J. Giordano,R. K.Durbin,and W. T.McAllister. 1990. Synthesis of functional mRNA in mammalian cells by bacteriophage T3 RNA polymerase. Mol. Cell. Biol. 10:4529-4537.
42. Zuker, M., and P. Stiegler. 1981. Optimal computerfolding of large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Res. 9:133-148.
on November 10, 2019 by guest
http://jvi.asm.org/