Estimating Mutation Rate: How to Count Mutations?
Yun-Xin Fu
1and Haying Huai
Human Genetics Center, University of Texas, Houston, Texas 77030
Manuscript received August 28, 2002 Accepted for publication March 11, 2003
ABSTRACT
Mutation rate is an essential parameter in genetic research. Counting the number of mutant individuals provides information for a direct estimate of mutation rate. However, mutant individuals in the same family can share the same mutations due to premeiotic mutation events, so that the number of mutant individuals can be significantly larger than the number of mutation events observed. Since mutation rate is more closely related to the number of mutation events, whether one should count only independent mutation events or the number of mutants remains controversial. We show in this article that counting mutant individuals is a correct approach for estimating mutation rate, while counting only mutation events will result in underestimation. We also derived the variance of the mutation-rate estimate, which allows us to examine a number of important issues about the design of such experiments. The general strategy of such an experiment should be to sample as many families as possible and not to sample much more offspring per family than the reciprocal of the pairwise correlation coefficient within each family. To obtain a reasonably accurate estimate of mutation rate, the number of sampled families needs to be in the same or higher order of magnitude as the reciprocal of the mutation rate.
A
significant fraction of the genetic research of the direct estimate of the rate of mutation. Theseexperi-ments may be time consuming, but the statistical method last century has been to illuminate various aspects
of mutation (De Vries1901/1903;LuriaandDelbru¨ ck used for estimating mutation rate is straightforward and
should not be controversial. It appeared indeed to be
1943;McClintock1950;KeightleyandEyre-Walker
1999). This is natural because mutations are the ultimate the case for early geneticists (Bridges 1919; Wright
and Eaton 1926; Fisher 1930; Dobzhansky and source of genetic variation upon which natural selection
and other evolutionary forces can act (Kimura 1983; Wright1941). However, the increasing number of
ob-servations that some mutant offspring share the same
Lynch and Hill 1986; Johnson 1999). Early
experi-mutation has prompted many contemporary geneticists
ments on mutation rate include those byCastle(1905,
to reconsider how mutation rate should be estimated
1929),Muller(1920, 1928), andMorgan(1950). To
(Engels1979;RussellandRussell1996;Neel1998; date, extensive mutation data, from either mutation
Thompsonet al. 1998). experiments or surveys, are available for many species,
A clustered mutation means that two or more progeny
particularly fruit flies (Schalet1960;Crow and
Sim-of a family inherit the same mutation (Purdom et al.
mons1983), mice (FavorandNeuhauser-Klaus1994;
1968;HartlandGreen1970;FavorandNeuhauser
-Russelland Russell1996), and humans (Neel and
Klaus 1994). Mutation clusters have been widely
ob-Rothman 1978; Cooper and Krawczak 1993; Crow
served and are now considered as general rather than 1993, 1999). Due to the importance of mutation rate,
as the exception (Hall 1988; Drost and Lee 1995;
such experiments will be likely to continue in the future,
MohrenweiserandZingg1995;HuaiandWoodruff
with more and more details being revealed by the advent
1997; Paashuis-Lew and Heddle 1998; Lewis 1999;
of new molecular techniques (KondrashovandCrow
for reference, see Woodruff et al. 1996). The most
1993;Fu1994).
important issue created by mutation cluster is how to In a typical mutation experiment, some aspects of the
count the mutations for the purpose of estimating muta-progeny of well-characterized parents are examined. A
tion rate. Several ways of counting have been proposed. mutant is identified if an offspring differs from its
par-One is to count each mutant offspring as one mutation, ents in a way that can be explained only by invoking a
disregarding whether or not the mutation is shared
mutation (Auerbach1959;Drake1991, 1993). When
(Haldane1935;SpencerandStern1948;Auerbach
a large number of offspring of mating pairs have been
1962; Muller et al. 1963; Combes et al. 1989; Huai
examined, the proportion of mutant progeny yields a
1997). The second is to count each cluster as only one
mutation (Russell 1977;Shukla et al. 1979; Heddle
et al. 1996; Nishinoet al. 1996). The third is to count
1Corresponding author:Human Genetics Center, SPH, University of
only those mutations that are not clustered (
Abra-Texas, 1200 Herman Pressler, Houston, TX 77030.
E-mail: [email protected] hamsonandWolff1976;RussellandRussell1992),
but in many cases this third choice is made because only the induced mutations in limited stages of the life
cycle are measured (Masonet al. 1987;Arraultet al.
2002). Various arguments have been put forward to
support one method or the other (RussellandRussell
1992, 1996; Huai and Woodruff 1998a,b; Heddle
1999), but no resolution has been obtained to date (Thompsonet al. 1998;StuartandGlickman 2000). In this article, we show for the first time that counting each mutant as one mutation regardless of cluster is the correct way to obtain an unbiased estimate of the mutation rate. Of equal significance is the formula for the sampling variance of the estimator, which does not require knowing all family sizes. We also discuss two important issues in designing a mutagenesis experi-ment, the sampling strategy and sample size
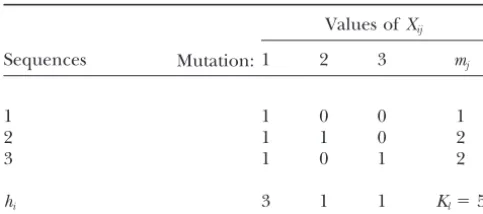
require-Figure1.—An example of mutations and their relationships
ment. Furthermore, we reanalyze several large data sets
withhiandmj. and show that some of the results in mutation rate
estimates have an undesirably large variance.
The equivalence of this counting to that of Equation
THEORY
1 can be proved as follows. LetXijbe the index variable
Counting mutations: Suppose a total of m haploid that takes value 1 if theith mutation is inherited by the
families are studied in an experiment. Let ni be the jth sequence and value 0 otherwise. Figure 1 (see Table
number of offspring examined in theith family andn⫽ 1 also) gives an example, from which we can easily see
n1⫹. . .⫹nmbe total sample size. Considering thelth the relationship betweenX
ijandhi andmjas family, an individual is a mutant if it differs from its
parent(s) by at least one mutation. It is important to hi ⫽Xi1⫹. . . ⫹Xin
l (5)
realize that mutations in different mutant individuals
mj⫽X1j⫹. . . ⫹XIj. (6)
are not necessarily distinct. LetKlbe the total number
of mutations counted as follows: each mutation that is
It follows that
inherited bykindividuals (more preciselyksequences)
is countedktimes. That is, suppose that there are in total K
l⫽h1 ⫹. . .⫹ hI
I independent mutation events, and the ith mutation
is inherited byhisequences. Then ⫽
兺
I
i⫽1
兺
nl
j⫽1
xij
Kl⫽h1⫹. . . ⫹hI. (1)
⫽
兺
nl
j⫽1
兺
I
i⫽1
xij Note that if there is only one mutant for each mutation
event,Kis simply the number of mutant individuals.
The total number of mutations in the experiment is ⫽m1⫹m2⫹. . . ⫹mnl.
defined as (7)
One special case deserves mentioning. When only
K⫽K1⫹. . . ⫹Km. (2)
one sequence is examined per family, each mutation
event will be counted exactly once. Therefore, K in
A mutation is said to be size iif there arei mutants
in the sample sharing that mutation. Letclbe the num- this situation is equal to the number of independent
ber of clusters of mutations of sizel. Then it is easy to mutations in the experiment.
see that RepresentingKby Equation 4 provides a convenient
basis for studying its statistical properties, and we discuss
K⫽c1⫹2c2⫹ . . .⫹ LcL, (3)
the mean and variance below.
whereLis the maximum cluster size. The mean and variance of K: Let R represent the
Consider thelth family again. Although Equation 1 number of DNA replications between two successive
indicates thatKis counted by considering each muta- generations andibe the mutation rate for theith cell
tion at a time, it can also be counted by considering replication. Let
one sequence at a time. Letmjbe the number of
muta-i⫽ 1⫹. . . ⫹ i. (8)
tions that occurred in individual (sequence)j. Then
TABLE 1
An example of mutations and their relationships with
hiandmjas shown in Figure 1
Values ofXij
Sequences Mutation:1 2 3 mj
1 1 0 0 1
2 1 1 0 2
3 1 0 1 2
hi 3 1 1 Kl⫽5
sequence (Figure 2). We assume thatRis a fixed number, Figure2.—Relationship between two sequences in a family.
which is appropriate for experiments in which individuals sampled are of the same sex and about the same age.
We assume that the number of mutations at
replica-tionifollows the Poisson distribution with both mean fore, whether a single gene is considered or multiple
and variance ofi. Thus for anyjsequence overRinde- genes are pooled,ˆ is an unbiased estimator of.
pendent cell divisions, We now consider the variance ofˆ . Consider first the
case in whichnl⫽1 for alli. As we mentioned earlier,
E(mj)⫽ Var(mj)⫽ , (9)
Kis equivalent to the number of mutation events. Since
the mutations in different families are independent, we
whereE( ) and Var( ) stand for expectation and
vari-have ance, respectively. We also assume that mutations in
different families are independent.
Var(K)⫽
兺
l
Var(Kl)⫽n. (13)
Since
E(Kl)⫽
兺
E(mj) (10) Therefore⫽nl, Var(ˆ )⫽ Var(k)/n2 ⫽ /n. (14)
it follows that In general, we have
E(K)⫽
兺
lE(Kl) Var(K
i)⫽
兺
Var(mi)⫹兺
i⬍jCov(mi,mj)
⫽
兺
l
nl ⫽n ⫹ φ
兺
l
nl(nl ⫺1) , (15)
⫽n. (11)
where
This equation suggests that an unbiased estimator ofis φ⫽
Cov(mi,mj) .
ˆ ⫽ K/n, (12)
So φis the covariance between mi and mj, that is, the
covariance of the numbers of mutations in any pair of which is exactly the way the mutation rate has been
sequences from the same family. Therefore
estimated in some empirical studies (e.g., Haldane
1935; Spencer and Stern 1948; Auerbach 1962;
Var(K)⫽n ⫹
兺
l
nl(nl⫺ 1) .
Muller et al. 1963; Woodruff and Thompson 1992;
Huai1997; Drakeet al. 1998;Thompsonet al. 1998).
And then Counting each cluster as one mutation or counting only
independent mutations will result in underestimation
Var(ˆ )⫽
n ⫹
兺
lnl(nl⫺ 1)
n2 , (16)
of the true mutation rate; in some cases the
underesti-mation may well be a few fold (Paashuis-Lewand
Hed-dle1998;Selby1998a,b;Thompsonet al. 1998;Heddle where ⫽φ/ is the correlation coefficient between
the numbers of mutations in two different sequences
1999). Since the mutation ratein this article is defined
as the sum of mutation rates over cell replications that from the same family.
Two special cases are illuminating. The first is that
are not necessarily equal, it does not matter if
exam-ined,i.e.,nl⫽c,l⫽1,. . .,m. It follows from Equation it is thus the proportion of mutations that are shared by two sequences from the same family.
16 that
Let rl,ij be the number of shared mutations for
se-Kl⫽K/(mc)
quencesiandjin thelth family. It thus follows that
and
E[rl,ij]⫽ E(t)
. (27)
Var(ˆ )⫽ 1
m
冢
1
c ⫹ c ⫺1
c
冣
. (17)The above equation suggests that an unbiased
estima-The second case is that all the offspring of each family tor ofE(t)is
are examined. Since offspring number of a family is
typically not a predetermined quantity, we have ˆE(t)⫽
2
兺
lnl(nl⫺1)兺
l兺
i⬍jrl,ij, (28)
E(K)⫽
兺
E(nl) (18)wherenl(nl⫺1)/2 is the number of pairs of sequences
Var(K)⫽
兺
E(nl)⫹φ兺
E[nl(nl⫺ 1)]. (19)for all the sample from the lth family. Suppose that
The most common practice is to assume that offspring there are in totalLobserved clusters and c
i is the size
number of a family follows a Poisson distribution with of clusteri. The above estimator can then be written as
meanf. Then
ˆE(t)⫽
兺
L
i⫽1ci(ci⫺ 1)
兺
lnl(nl⫺1). (29)
E(K)⫽mE(Kl)⫽ mf (20)
Var(K)⫽mVar(Kl)⫽mf ⫹mφf2. (21)
Substituting this into Equation 16 yields an unbiased
estimate of the standard error ofˆ as
So we have
ˆ ⫽ 1
n
冪
K⫹兺
L
i⫽1
ci(ci⫺1) . (30)
Var(ˆ )⫽ 1
m
冢
1
f ⫹
冣
. (22)Estimating the sampling variance:Sinceˆ is an unbi- Since many experiments on mutation rate were done
ased estimator, the precision of estimation is thus deter- over the many decades, detailed cluster sizes may not
mined by its variance. To compute the variance ofˆ , be available. So the above formula is difficult to apply
we need to know the value of the covariance between in many situations. However, boundaries of the standard
two mutations (see Figure 2). error ofˆ can be obtained on the basis of partial
infor-Suppose that sequence i and sequence j shared a mation as follows.
common ancestortcell replications ago. Then we can If only the numberSof singletonsLandKis known,
expressmiandmjas then the minimum value attainable by兺ci(ci⫺1) is (K⫺
S)(K ⫺ S ⫺ L)/L, corresponding to the situation in
mi⫽ rij⫹ri (23)
which all clusters are of equal size (K⫺S)/L. A
lower-mj⫽ rij⫹rj, (24) bound of the standard error is thus given by
where rij represents the number of mutations in the
min⫽
1
n
冪
K⫹(K⫺S)(K⫺S⫺L)
L . (31)
common ancestor (shared mutations), andriandrjare
the numbers of mutations in sequencesiandj,
respec-Letcminandcmaxbe the minimum and maximum
clus-tively, since the separation from their common ancestor
ter sizes, respectively. Then we have 2ⱕ cmin ⱕ cmax ⱕ
(Figure 2). Conditional on thetvalue,rij,ri, andrjare
K⫺S⫺cmin(L⫺1). The maximum value of兺ci(ci⫺ 1)
independent Poisson variable with means equal to t,
corresponds to the situation in which there are as many
⫺ t, and ⫺ t, respectively. Therefore
clusters ofcminsize as possible. An upper bound of the
E(mimj)⫽ Et[E(r2ij ⫹rij(ri⫹ rj)⫹ rirj)]
standard error is therefore given by
⫽ Et[t⫹ 2t ⫹2t( ⫺ t)⫹( ⫺ t)2]
max⫽
1
n
√
K⫹cmin(L⫺b)⫹ bcmax(cmax ⫺1) , (32)⫽ Et[t⫹ 2]
⫽ E(t)⫹ 2. (25) whereb ⫽(K⫺ S⫺cminL)/(cmax⫺cmin).
We can also construct a confidence interval for
esti-It follows thatφ⫽E(mimj)⫺E(mi)E(mj)⫽ E(t). Soφis
matingˆ . Note thatˆ is the average of many variables
the expected number of shared mutations between two
with the same mean and variance and 0 or small covari-sequences from the same family. Since the correlation
ance; therefore, by the central limit theorem of
proba-coefficientis defined as
bility,ˆ can be approximated by a normal distribution
with meanand variance2. The 95% confidence
inter- ⫽ E(t)
TABLE 2
Examples of estimates of mutation rate and standard error
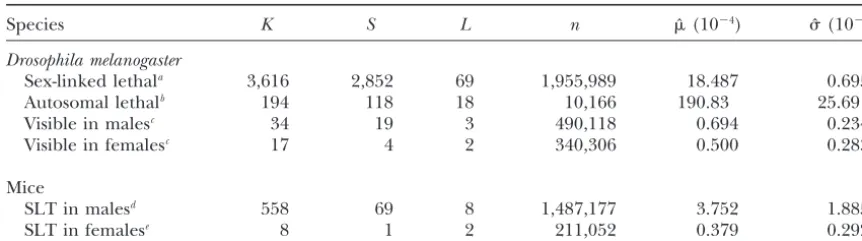
Species K S L n ˆ (10⫺4) ˆ (10⫺4)
Drosophila melanogaster
Sex-linked lethala 3,616 2,852 69 1,955,989 18.487 0.695
Autosomal lethalb 194 118 18 10,166 190.83 25.69
Visible in malesc 34 19 3 490,118 0.694 0.234
Visible in femalesc 17 4 2 340,306 0.500 0.283
Mice
SLT in malesd 558 69 8 1,487,177 3.752 1.885
SLT in femalese 8 1 2 211,052 0.379 0.292
aDetails in tables ofMasonet al. (1985) andWoodruffandThompson(1992).
bDetails inSchalet(1960),Shuklaet al. (1979),Woodruffet al. (1996), and also in Table 3 ofThompson et al. (1998).
cSchalet(1960) andShuklaet al. (1979); see also Table 3 ofThompsonet al. (1998).
dFavorandNeuhauser-Klaus (1994),Russelland Russell (1996),Drostand Lee (1995, 1998), and
Selby (1998a,b).
eRussell(1964),RussellandRussell (1992), andSelby (1998a,b).
Even though the mutation rate estimates of male and assumption of Bernoulli (Poisson) distribution due to
premeiotic mutation events. Even though his formulas female mice have a not so small variance, we note that
the mean ratio of the mutation rate estimates of mouse appear to be more influential than Muller’s, especially
in Drosophila mutagenesis involving transposable ele-males over mouse feele-males is 10 in Table 2, compared
to the value of 2 in Changet al. (1994). The sample ments (Margulieset al. 1986;EhlingandNeuhauser
-Klaus1988;Brownet al. 1989;BadgeandBrookfield
size for female mice in the specific locus test (SLT) is
far below that for male mice. Thus cluster mutations in 1998), most of his conclusions and parameter
estima-tions are not relevant to the premeiotic cluster muta-female mice may well be underestimated or not
recov-ered. More experiments are needed for a specific locus tions.
Sampling strategy: Since ˆ is an unbiased estimator test of female mice.
of, sampling strategy should aim at reducing the
vari-ance of the estimate. When to sample and how to sample both play important roles in determining the variance.
DISCUSSION
Before we examine these two issues in turn, we note
Relation to others’ work:Muller(1952, 1962)
pre-that the best possible strategy is to examine as many
sented an estimator of the standard error ofˆ asˆ ⫽ families as possible but only one sequence per family.
q(1/n)
√
兺Li⫽1ci(ci⫺1) (q ⫽ 1 ⫺ ˆ , and singletons are This sampling strategy will minimize the sampling
vari-considered as special cases of cluster mutations), which ance for a fixed total number of individuals examined
is close to but tends to underestimateˆ compared with (compare Equation 13 with Equation 16). Apparently
Equation 30 of this article. The derivation of this for- this strategy is neither practical in most situations for
mula was never given, but Muller appeared to have multicellular organisms nor possible in somatic or
bacte-obtained this formula with the assumption that the pre- ria mutagenesis (LuriaandDelbru¨ ck1943; Heddle
meiotic mutations are not common; his brief explana- 1999; H.Huaiand Y.-X.Fu, unpublished results). There
tion of the formulas suggested that it may be the result are good reasons for choosing large family sizes in
cer-of a compound or generalized Poisson process, which tain situations. For example, it is important to know
is the sum of independent Poisson variables. the value of for the purpose of understanding the
Engels(1979) also gave formulas for both mean and clustering phenomenon of mutations, and can be
variance of mutation rates when cluster mutations are estimated better with larger family sizes. Another reason
present. He focused mainly on a clustering model vio- favoring larger family size is that there may be a nearly
lating homogeneity of mutation probabilities among fixed amount of effort or expense for each family
different parents. Engels visualized a conceptual two- screened regardless of its size.
step experiment. The first step consists of choosing from It is clear from the variance equation (16) that the
a pool of possible mutation rates for all different par- best sampling time is the one that minimizes the
correla-ents. The second step is the independent Bernoulli sam- tion coefficientbetween any pair of progeny within a
pling within each family. So Engels did not address family. Apparently, this indicates that the best time to
TABLE 3
The variation coefficient␣(␣⫽ˆ /ˆ ) when all themfamilies are of the same sizefand when⫽0.10
Family size (f)
m⫻u 1 5 10 50 100 1,000 10,000
0.01 10.000 5.292 4.359 3.435 3.302 3.176 3.164
0.1 3.162 1.673 1.378 1.086 1.044 1.004 1.000
1 1.000 0.529 0.436 0.344 0.330 0.318 0.316
3 0.577 0.306 0.252 0.198 0.191 0.183 0.183
10 0.316 0.167 0.138 0.109 0.104 0.100 0.100
tion in multicellular species when germ cell number is f (orcmax)Ⰷ ⫺1, the further increase of mean family
size does not help to reduce the variance much (see small. Also note that males, especially older males in
many species, usually have much more germ cell divi- Table 3 and Figure 3). Therefore a reasonable strategy
is to examine as many progeny per family as possible sions than females have; hence it may be possible that
the correlation coefficientsare much lower in males, but not much more than⫺1.
Because of the important role ofin the estimate of
especially in older males.
Given that the sampling time is determined, that is, mutation rate and genetic counseling (Hartl 1971;
Wijsman 1991; Young1991), it is useful to obtain an
is fixed, how many individuals to sample from each
family depends on the value of. To demonstrate, con- estimate of from experimental data (Van Essen et
al. 1992; CooperandKrawczak 1993;Bridges1994;
sider the case wherenl⫽cmax⫽f,l⫽1, . . . ,mor that
family size follows Poisson distribution. We note from Zlotogora1998). From Equation 29, we can estimate
ˆE(t)by
both Equations 17 and 22 that the variance of the esti-mate of mutation rate is approaching
ˆE(t)⫽
兺
L
i⫽1ci(ci⫺ 1)
(m⫺1)2
f ⫹n(f⫺1)
, (34)
1
mˆE(t) (33)
where fis the mean family size and2
f is the sampling
variance of the family size,i.e.,
with an increasing and large enough family size. Once
a useful experiment for estimating mutation rate should
2
f ⫽ 1
m⫺ 1
兺
i(ni⫺ f)2 (35)
examine at least as many families as the reciprocal of the mutation rate (Table 3, Figures 3 and 4). This con-clusion will reintroduce the problem of how to avoid
⬇ 1
m
兺
in2
i ⫺f2. (36)
preexisting mutations if the sampled family number is at least as large as the reciprocal of the mutation rate.
So can be estimated by
Can we find a way that de novocluster mutations can
be discriminated confidently from preexisting
muta- ⫽ E(t)
tions? It is relatively easy to eliminate the preexisting
mutations that are from grandparents’ heterozygosity (Yang et al. 2001), but if one of the grandparents is
⫽ n
兺
L
i⫽1ci(ci⫺1)
K[(m⫺1)2
f ⫹n(f⫺ 1)]
(37)
genetic mosaic for a new mutation, then we need careful
analysis of its timing and effects (H.Huaiand Y.-X.Fu,
unpublished results).
⬇
兺
Li⫽1ci(ci⫺1)K(2
f/f⫹f)
. (38)
Estimating from the number of mutation events:
Let Ibe the number of mutation events in an
experi-Kis the total number of mutants recovered in the
ex-ment that examinedsoffspring. As we have shown, when
periment, counting all members of any clusters.
one sequence is examined per family,Iis the same as
Sample size requirement: Besides sampling strategy
Kso usingI/syields an unbiased estimate of. When
to reduce variance in the estimate, it is important to
multiple offspring from a family are examined, I can
determine the sample size required in achieving a given
be ⬍K; the magnitude of difference depends on the
precision in estimation. It is obvious that the standard
number of offspring examined as well as onˆE(t).
error of an estimate should not be larger than the
esti-SinceK/nis an unbiased estimate of, it follows that
mate itself. For a good estimate, the standard error
I/nis an underestimate of mutation rate. It is tempting
should probably be an order of magnitude smaller than
to suggest an estimate as the estimate.
Kl ⫽I/n⬘, (41) For simplicity, consider the case that an equal number
of progeny is examined for each family. Suppose we
wheren⬘is a constant satisfyingmⱕn⬘ ⬍n. However,
want to ensure that the standard error is as small as␣. it is not obvious what valuen⬘should be. Hence
count-Then family numbermand family size f(cmax) need to ing each mutant as one mutation regardless of the
clus-satisfy ter’s origin is the more straightforward way to obtain
an unbiased estimate of the mutation rate. Nevertheless, 1
m
冢
1
f ⫹ f⫺ 1
f
冣
⫽ ␣22 (39)
it should be worth exploring efficient ways to use the frequencies of various mutation events.
Alternative ways to combine information: We note or
thatKl/nlis an unbiased estimate of. So
m⫽ ␣⫺2⫺1
冢
1f ⫹ f⫺ 1
f
冣
. (40) ˆ⬘ ⫽兺
iiKl/nl (42)is an unbiased estimator, where i ⱖ 0 and 兺ii ⫽ 1.
We can see from Table 3 and Figure 3 that it is obvious
Because
that once the uniformly sampled family sizefis above
the reciprocal of, further increases offwill not help Var(ˆ⬘)⫽
兺
i (2
i/nl)( ⫹ E(t)(nl⫺1)) , (43) reduce the standard error of mutation rate estimation
very much. In these cases there is an optimum family
to obtain a best linear estimate of,needs to satisfy
size (a little bit ⬎⫺1) at which the experiment is most
efficient. So it is not always true that the larger the 2
i
ni( ⫹ E(t)(ni⫺1))
⫽ 2m
nm( ⫹ E(t)(nm⫺1))
, (44)
sample size, the more precise the experiment.
On the other hand, given that the total sample size
wherei⫽1, . . . ,m⫺1. That is,
is fixed (shown in Figure 4), the best sampling strategy
is to examine as many families as possible where the 
i
m
⫽ ni( ⫹ E(t)(ni⫺ 1))
nm( ⫹ E(t)(nm⫺1))
. (45)
family sizefis fixed at one, where each sample is
inde-pendent from others. If this is not feasible, try to fix
So the family size as low as possible when there is no need
for estimation of.
i⫽
冢
兺
ini
1⫹ (ni⫺1)
冣
⫺1
ni
1⫹ (ni⫺1)
. (46)
Also family size f can affect the number of families
that need to be studied, but the most important factor
Figure4.—The total sample size fixed at
m⫻f⫽100/, and all themfamilies are of the same sizef. The designed family sizes vary from 1.0 to 1000 to visualize their ef-fects on the variation coefficient of muta-tion rate estimamuta-tion.
Combes, R. D., J. Bootman, M. G. Ford, J. HepworthandD. W. combine data from different families is to give each
Salt, 1989 Statistical method for the design and analysis of
individual equal weight regardless of family affiliation, mutation experiments with the fruit flyDrosophila melanogaster, pp.
251–283 inStatistical Evaluation of Mutagenicity Test Data, edited by because each provides the same amount of independent
D. J.Kirkland. Cambridge University Press, Cambridge, UK.
information; whenis close to one, the best way is to
Cooper, D. N., andM. Krawczak, 1993 Human Gene Mutation. BIOS
give equal weight to each family, because each family Scientific Publishers, Oxford.
Crow, J. F., 1993 How much do we know about spontaneous human provides the same amount of information regardless of
mutation rates? Environ. Mol. Mutagen.21:122–129. size.
Crow, J. F., 1999 Spontaneous mutation in man. Mutat. Res.437: 5–9.
This work was supported in part by National Institutes of Health
Crow, J. F., andM. J. Simmons, 1983 The mutation load in Drosoph-grants R29 GM50428 (Y.X.F.) and R01 HGO1708 (Y.X.F.).
ila, pp. 1–35 inThe Genetics and Biology of Drosophila, Vol. 3c, edited by M.Ashburner, H. L.Carsonand J. N.Thompson. Academic Press, London.
De Vries, H., 1901/1903 Die Mutations Theorie, Vols. 1 and 2, Veit,
LITERATURE CITED
Leipzig, Germany (English translation, 1909/1910. Open Court, Abrahamson, S., and S. Wolff, 1976 Re-analysis of radiation- Chicago).
induced specific locus mutations in mouse. Nature264:715–719. Dobzhansky,T. H., and S.Wright, 1941 Genetics of natural popu-Arrault, X., V. Michel, P. Quillardet, M. HofnungandE. Touati, lations. V. Relations between mutation, rate and accumulation 2002 Comparison of kinetics of induction of DNA adducts and of lethals in populations ofDrosophila pseudoobscura. Genetics26: gene mutations by a nitrofuran compound, 7-methoxy-2-nitro- 23–51.
naphtho[2,1-b]furan (R7000), in the caecum and small intestine Drake, J. W., 1991 Spontaneous mutation. Annu. Rev. Genet.25:
of Big BlueTM mice. Mutagenesis17:353–359. 125–146.
Auerbach, C., 1959 Spontaneous mutations in dry spores of Neuro- Drake, J. W., 1993 Rates of spontaneous mutation among RNA
spora crassa. Z. Vererbungsl.90:335–346. viruses. Proc. Natl. Acad. Sci. USA90:4171–4175.
Auerbach, C., 1962 Mutation: An Introduction to Research in Mutagene- Drake, J. W., B. Charlesworth, D. CharlesworthandJ. F. Crow,
sis. Part I. Methods.Oliver & Boyd, Edinburgh. 1998 Rates of spontaneous mutation. Genetics148:1667–1686. Badge, R. M., andJ. F. Y. Brookfield, 1998 A novel repressor of Drost, J. B., andW. R. Lee, 1995 Biological basis of germline muta-P-element transposition inDrosophila melanogaster.Genet. Res.71: tion: comparisons of spontaneous germline mutation rates
21–30. among Drosophila, mouse, and human. Environ. Mol. Mutagen.
Bridges, C. B., 1919 The stages at which mutations occur in the 25(Suppl. 26): 48–64.
germ tract. Proc. Soc. Exp. Biol. Mod.17:1–2. Drost, J. B., and W. R. Lee, 1998 The developmental basis for
Bridges, P. J., 1994 The Calculation of Genetic Risks. Johns Hopkins the germline mosaicism in mouse andDrosophila melanogaster.
University Press, Baltimore. Genetica102/103:421–443.
Brown, A. J. L., L. S. Alphey, A. J. Flavell, T. I. Gerasimovaand Ehling, U. H., andA. Neuhauser-Klaus, 1988 Induction of spe-S. J. Ross, 1989 Instability in the Ctmr2 strain of Drosophila cific-locus and dominant-lethal mutations by cyclophosphamide
melanogaster: role of P-element functions and structure of re- and combined cyclophosphamide-radiation treatment in
male-vertants. Mol. Gen. Genet.218:208–213. mice. Mutat. Res.199:21–30.
Castle, W. E., 1905 The mutation theory of organic evolution: from Engels, W. R., 1979 The estimation of mutation rates when premei-the standpoint of animal breeding. Science21:521–525. otic events are involved. Environ. Mutagen.1:37–43.
Castle, W. E., 1929 A mosaic(intense-dilute) coat pattern in the Favor, J., and A. Neuhauser-Klaus, 1994 Genetic mosaicism in the house mouse. Annu. Rev. Genet.28:27–47.
Fisher, R. A., 1930 Note on a tricolour (mosaic) mouse. J. Genet. Radiosensitivity in Germ Cells, edited byF. Sobels. Pergamon Press,
23:77–81. Oxford.
Fu, Y. X., 1994 A phylogenetic estimator of population size or muta- Neel, J. V., 1998 A reappraisal of studies concerning the genetic
tion rate. Genetics136:685–692. effects of the radiation of humans, mice, and Drosophila. Environ.
Haldane, J. B. S., 1935 The rate of spontaneous mutation in a Mol. Mutagen.31:4–10.
human gene. J. Genet.31:317–326. Neel, J. V., andE. D. Rothman, 1978 Indirect estimates of mutation
Hall, J. G., 1988 Somatic mosaicism: observations related to clinical rates in tribal Amerindians. Proc. Natl. Acad. Sci. USA75:5585–
genetics. Am. J. Hum. Genet.46:1187–1193. 5588.
Heddle, J. A., 1999 On clonal expansion and its effects on mutant Nishino, H., D. J. Schaid, V. L. Buettner, J. HaavikandS. Sommer, frequencies, mutation spectra and statistics for somatic mutations 1996 Mutation frequencies but not mutant frequencies in big
in vivo. Mutagenesis14:257–260. blue mice fit a Poisson distribution. Environ. Mol. Mutagen.28:
Heddle, J. A., L. Cosentino, G. Dawood, R. R. SwigerandY. Paas- 414–417.
huis-Lew, 1996 Why do stem cells exist? Environ. Mol. Muta- Paashuis-Lew, Y. R. M., andJ. A. Heddle, 1998 Rates of mutation
gen.28:334–341. during embryogenesis and growth. Mutagenesis13:613–617.
Hartl, D. L., 1971 Recurrence risks for germinal mosaics. Am. J. Purdom, C. C., K. F. DyerandD. G. Papworth, 1968 Allelic clusters
Hum. Genet.23:124–134. among spontaneous mutations in Drosophila. Mutat. Res.5:305–
Hartl, D. L., andM. M. Green, 1970 Genetic studies of germinal 307. mosaicism inDrosophila melanogasterusing the mutablewcgene.
Russell, L. B.,1964 Genetic and functional mosaicism in the mouse,
Genetics65:449–456. pp. 153–181 inThe Role of Chromosomes in Development, edited by
Huai, H., 1997 The evolutionary implication of premeiotic clusters M. Locke. Academic Press, New York.
of mutation. Ph.D. Thesis, Bowling Green State University, Bowl- Russell, L. B., andW. L. Russell, 1992 Frequency and nature of
ing Green, OH. specific-locus mutations induced in female mice by radiation and
Huai, H., andR. C. Woodruff, 1997 Clusters of identical new muta- chemical: a review. Mutat. Res.296:107–127.
tions can account for the “overdispersed” molecular clock. Genet- Russell, L. B., andW. L. Russell, 1996 Spontaneous mutations
ics147:339–348. recovered as mosaics in the mouse specific-locus test. Proc. Natl.
Huai, H., andR. C. Woodruff, 1998a Clusters of new identical Acad. Sci. USA93:13072–13077.
mutants and the fate of underdominant mutations. Genetica Russell, W. L., 1977 Mutation frequencies in female mice and the
102/103:489–505. estimation of genetic hazards of radiation in women. Proc. Natl.
Huai, H., andR. C. Woodruff, 1998b With the correct concept of Acad. Sci. USA74:3523–3527.
mutation rate, cluster mutations can explain the overdispersed Schalet, A., 1960 A study of spontaneous visible mutations in
Dro-molecular clock. Genetics149:467–469. sophila melanogaster. Ph.D. Thesis, Indiana University,
Blooming-Johnson, T., 1999 Beneficial mutations, hitchhiking and the evolu- ton, IN.
tion of mutation rates in sexual populations. Genetics151:1621– Selby, P. B., 1998a Major impacts of gonadal mosaicism on
heredi-1631. tary risk estimation, origin of hereditary diseases, and evolution.
Keightley, P. D., andA. Eyre-Walker, 1999 Terumi Mukai and Genetica102/103:445–462.
the riddle of deleterious mutation rates. Genetics153:515–523. Selby, P. B., 1998b Discovery of numerous clusters of spontaneous Kimura, M., 1983 The Neutral Theory of Molecular Evolution. Cam- mutations in the specific-locus test in mice necessitate major
bridge University Press, Cambridge, UK. increases in estimates of doubling doses. Genetica102/103:463–
Kondrashov, A. S., andJ. F. Crow, 1993 A molecular approach to 487.
estimating the human deleterious mutation rate. Hum. Mutat. Shukla, P. T., K.Sankaranarayanan and F. H.Sobels, 1979 Is
2:229–234. there a proportionality between the spontaneous and the
x-ray-Lewis, S. E., 1999 Life cycle of the mammalian germ cell: implication induced rates of mutation? Mutat. Res.61:229–248.
for spontaneous mutation frequencies. Teratology59:205–209. Spencer, W. P., andC. Stern, 1948 Experiments to test the validity Luria, S. E., andM. Delbru¨ ck, 1943 Mutations of bacteria from
of the linear r-dose/ mutation frequency relation in Drosophila virus sensitivity to virus resistance. Genetics28:491–511. at low dosage. Genetics33:43–74.
Lynch, M., andW. G. Hill, 1986 Phenotypic evolution by neutral
Stuart, G. R., andB. W. Glickman, 2000 Through a glass, darkly:
mutation. Evolution40:915–935. reflections of mutation from
lacItransgenic mice. Genetics155:
Margulies, L., D. I.Briscoeand S. S.Wallace, 1986 The
relation-1359–1367. ship between radiation-induced and transposon-induced
genetic-Thompson, J. N., R. C. Woodruffand H. Huai, 1998 Mutation damage during Drosophila oogenesis. Mutat. Res.162:55–68.
rate: a simple concept has become complex. Environ. Mol. Muta-Mason, J. M., R. Valencia, R. C. WoodruffandS. Zimmering, 1985
gen.32:292–300. Genetic drift and seasonal variation in spontaneous mutation
Van Essen, A. J., S. Abbs, M. Baiget, E. Bakker, C. Boileauet al., frequencies in Drosophila. Environ. Mutagen.7:663–676.
1992 Parental origin and germline mosaicism of deletions and Mason, J. M., R. Valencia, R. C. Woodruffand S. Zimmering,
duplications of the dystrophin gene: a European study. Hum. 1987 A guide for performing germ cell mutagenesis assays using
Genet.88:249–257. Drosophila melanogaster. Mutat. Res.189:93–102.
Wijsman, E. M., 1991 Recurrence risk of a new dominant mutation McClintock, B., 1950 The origin and behavior of mutable loci in
in children of unaffected parents. Am. J. Hum. Genet.48:654– maize. Proc. Natl. Acad. Sci. USA36:344–355.
661. Morgan, W. C., 1950 A new tail-short mutation in the mouse. J.
Woodruff, R.C., andJ. N. Thompson, 1992 Have premeiotic clus-Hered.41:208–215.
ters of mutation been overlooked in evolutionary theory? J. Evol. Mohrenweiser, H., andB. Zingg, 1995 Mosaicism: the embryo as
Biol.5:457–464. a target for induction of mutations leading to cancer and genetic
Woodruff, R. C., H. HuaiandJ. N. Thompson, 1996 Clusters of new disease. Environ. Mol. Mutagen.25(Suppl. 26): 21–29.
mutation in the evolutionary landscape. Genetica98:149–160. Muller, H. J., 1920 Further changes in the white-eye series of
Dro-Wright, S., andO. N. Eaton, 1926 Mutational mosaic coat patterns sophila and their bearing on the manner of occurrence of
muta-of the guinea pig. Genetics11:333–351. tion. J. Exp. Zool.31:443–472.
Yang, H. P., A. Y. Tanikawa, W. A. Van Voorhies, J. C. Silvaand Muller, H. J., 1928 The measurement of gene mutation rate in
A. S. Kondrashov, 2001 Whole-genome effects of ethyl meth-Drosophila, its high variability, and its dependence on
tempera-anesulfonate-induced mutation on nine quantitative traits in out-ture. Genetics13:279–357.
bredDrosophila melanogaster.Genetics157:1257–1265. Muller, H. J., 1952 The standard error of the frequency of mutants
Young, I. D., 1991 Introduction to Risk Calculations in Genetic
Con-some of which are of common origin. Genetics37:608.
sulting. Oxford University Press, Oxford. Muller, H. J., 1962 Studies in Genetics, p. 301. University of Indiana
Zlotogora, J., 1998 Germ line mosaicism. Hum. Genet.102:381– Press, Bloomington, IN.
386. Muller, H. J., I.Osterand S.Zimmering, 1963 Are chronic and
acute gamma irradiation equally mutagenic in Drosophila?, pp.