The role of purinergic signalling in the musculoskeletal system
Full text
Figure

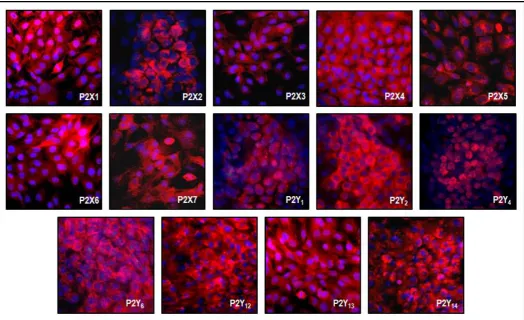
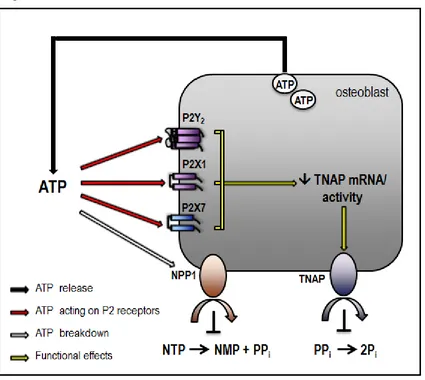
Related documents
Also, HPLC methods were reported for determination of Candesartan cilexetil in tablets, as a single component 3,7-9 , in combination with Candesartan and a
BRBS depends on local outlier factor (LOF) algorithm to specify points' importance score; then it uses these scores in selecting the border and rare points to be included in
Failing that, even a constant factor im- provement through the use of elliptic curves would be very welcome 3 We remark that Krohn-Freedman-Mazires’ homomorphic hash function can
(2019) / Survey of medicinal plants in Jyoti Nivas College (Autonomous), Bengaluru, Karnataka, India 11 campus and the information gathered was noted in.. a field note
establised coupled coincidence and coupled common fixed point results for a mixed g-monotone mapping satisfying Non-linear contraction in partially ordered G-metric
In the paper Customer behavior analysis using big data analytics by khade, they used decision tree algorithm to predict the customer behavior and to analysis
The addition of rituximab to a combination of fludarabine, cyclophosphamide, mitoxantrone (FCM) significantly increases the response rate and prolongs survival as
The goal of clinical trials for MDS is to improve treatment and quality of life and to increase survival. Patients’ participation in clinical trials is needed to improve
