Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Interaction of Poly(rC) Binding Protein 2 with the 5
9
Noncoding
Region of Hepatitis A Virus RNA and Its Effects
on Translation
JUDITH GRAFF,* JOHN CHA, LAWRENCE B. BLYN,†ANDELLIE EHRENFELD‡
Department of Molecular Biology and Biochemistry, University of California, Irvine, Irvine, California 92697
Received 26 March 1998/Accepted 24 August 1998
Utilization of internal ribosome entry segment (IRES) structures in the 5* noncoding region (5*NCR) of picornavirus RNAs for initiation of translation requires a number of host cell factors whose distribution may vary in different cells and whose requirement may vary for different picornaviruses. We have examined the requirement of the cellular protein poly(rC) binding protein 2 (PCBP2) for hepatitis A virus (HAV) RNA translation. PCBP2 has recently been identified as a factor required for translation and replication of poliovirus (PV) RNA. PCBP2 was shown to be present in FRhK-4 cells, which are permissive for growth of HAV, as it is in HeLa cells, which support translation of HAV RNA but which have not been reported to host replication of the virus. Competition RNA mobility shift assays showed that the 5*NCR of HAV RNA competed for binding of PCBP2 with a probe representing stem-loop IV of the PV 5*NCR. The binding site on HAV RNA was mapped to nucleotides 1 to 157, which includes a pyrimidine-rich sequence. HeLa cell extracts that had been depleted of PCBP2 by passage over a PV stem-loop IV RNA affinity column supported only low levels of HAV RNA translation. Translation activity was restored upon addition of recombinant PCBP2 to the depleted extract. Removal of the 5*-terminal 138 nucleotides of the HAV RNA, or removal of the entire IRES, eliminated the dependence of HAV RNA translation on PCBP2.
Members of the family Picornaviridae employ a unique ini-tiation mechanism for translation of their uncapped RNA ge-nomes (for reviews, see references 13 and 40). In contrast to
ribosomes scanning from the capped 59ends of the majority of
cellular mRNAs, the long (600- to 1,000-nucleotide [nt]) and
highly structured 59noncoding region (59NCR) of picornavirus
RNAs mediates the binding of ribosomal subunits internally at an internal ribosome entry segment (IRES), a region consist-ing of 400 to 500 nt. Despite little similarity in primary nucle-otide sequences and marked variations in predicted secondary
structures of the 59NCRs among the different genera of
picor-naviruses, the IRES elements are functionally equivalent.
Analysis of potential higher-order folds in the 59NCRs of both
viral and cellular RNAs suggest the presence of a common
core structure at the 39end of the IRES that may represent the
actual site of ribosome entry (27). Binding of the ribosomal initiation complex to the IRES element is presumably facili-tated by one or more cellular trans-acting protein factors dis-tinct from the canonical initiation factors. These factors may vary with respect to distribution in different cells and require-ment by different picornaviruses. Rabbit reticulocyte lysates (RRL) support accurate and efficient translation in vitro of cardiovirus and aphthovirus IRESes. In a reconstituted in vitro system, a set of purified canonical initiation factors alone were able to mediate internal ribosomal entry of encephalomyocar-ditis virus (EMCV) RNA, although some enhancement of ini-tiation complex formation was observed upon addition of the
cellular polypyrimidine tract binding protein (PTB) (34). In contrast, poliovirus (PV) and rhinovirus RNAs require supple-mentation of RRL with HeLa cell factors for correct and efficient translation (7, 12, 21). Function of the hepatitis A virus (HAV) IRES in RRL may require additional factors present in mammalian liver at higher concentrations than present in RRL (17). Addition of ribosomal salt wash (RSW) from other mouse tissues or cell lines did not stimulate HAV translation in RRL (17, 22). However, recently we showed that HeLa cells support the translation of HAV RNA in vivo and that translation of HAV RNA is efficient in HeLa cell extract (18). None of the picornavirus IRESes has been reported to function in a wheat germ translation system. Several proteins, most of which were isolated from HeLa cells, interact with
distinct regions in picornavirus 59NCRs and have functional
specificity regarding IRES utilization. These include the La autoantigen (p52), specific for PV RNA translation (29, 30); PTB (p57), which shows interaction with several picornavirus IRESes (9, 21, 24, 31, 32); and poly(rC) binding protein 2 (PCBP2; p39), shown to be required for PV translation (5, 6, 14).
The human cellular protein PCBP2 was identified in RSW of HeLa cells by RNA affinity column chromatography using PV stem-loop IV RNA (5). PCBP2 is present in most mammalian tissues, and it is related to a class of RNA binding proteins which contain three internal peptide repeats corresponding to K-homologous (KH) domains, originally identified in hetero-geneous nuclear ribonucleoprotein K (28, 39). PCBP2 does not contain other known nucleic acid binding motifs, and therefore the KH domain is believed to determine its RNA binding activity. Previous demonstration of binding to homopolymeric RNA sequences showed strong binding to poly(rC) in vitro and lesser binding to poly(rG) and poly(U) (28). The longest ho-mopolymeric sequence in the PV stem-loop IV binding site is four C residues. It is likely that PCBP2 has other RNA recog-nition specificities as well. Preliminary data from competition
* Corresponding author. Mailing address: National Institutes of Health, NIAID, LID, Molecular Hepatitis Section, Building 7, Room 200, 7 Center Dr., Bethesda, MD 20892-0740. Phone: (301) 496-6227. Fax: (301) 402-0524. E-mail: [email protected].
† Present address: ISIS Pharmaceuticals, Carlsbad, CA 92008. ‡ Present address: National Institutes of Health, National Institute of Allergy and Infectious Diseases, LVD, Picornavirus Section, Be-thesda, MD 20892.
9668
on November 9, 2019 by guest
http://jvi.asm.org/
assays showed that the IRES elements of different picornavi-ruses such as coxsackievirus B3, human rhinovirus 14, EMCV, and HAV were able to compete with PV RNA for binding of PCBP2 (4a). Dildine and Semler (11) showed the competition
of 59NCRs of coxsackievirus B3, human rhinovirus 14, and
Theiler’s murine encephalomyelitis virus for formation of a complex between PV stem-loop IV sequences and protein ex-tract from HeLa cells, later shown to represent PCBP2 (5).
HAV, a distant relative of PV, was chosen to examine a possible broader role of PCBP2 in IRES-mediated translation of other picornaviruses. The genome of HAV is composed of a single-stranded positive-sense RNA approximately 7,500 nt in length with a structural organization and gene order char-acteristic of picornaviruses. It encodes a predicted polyprotein of 2,227 amino acids, cleaved to functional polypeptides,
pre-ceded by a 734-nt-long 59NCR. Computer-assisted folding
pre-dictions and biochemical probing showed that the HAV
59NCR forms extensive higher-order structures, typical for the
long 59NCRs of picornaviruses, which usually contain about six
predicted stem-loop domains (8, 27). Also as found for the
other picornaviruses, the HAV 59NCR contains multiple AUG
codons upstream of the translation initiation codon, which are not used for translation initiation (41). However, as the sole member of the genus Hepatovirus within the family
Picorna-viridae, HAV shows many unique features. The nucleotide and
amino acid sequences of the HAV genome are only distantly related to those of other picornaviruses. HAV grows ineffi-ciently in cell cultures, usually establishing a persistent, non-lytic infection in the cultured cells and failing to inhibit host cell protein synthesis. Only one HAV-encoded proteinase, 3C, has been identified (16, 20, 23, 35, 36), and details of the polyprotein processing scheme in vivo have not been demon-strated. At the level of RNA translation, there is evidence that capsid coding sequences downstream of the initiation codon are involved in translation of the RNA in vitro, unlike the case for other picornaviruses (18, 45). No specific function of any
trans-acting factor has been identified to date for translation or
RNA replication. Proteins with molecular masses of 30, 39, and 110 kDa, isolated from RSW of BS-C-1 cells and FRhK-4 cells, as well as PTB, present in RRL and HeLa cell extract,
were found to bind to different regions within the 59NCR of
HAV (9), although the biological significance of these inter-actions has not been tested. A 39-kDa protein which interacted
with stem-loop IIIa of the HAV 59NCR was identified as
glyceraldehyde-3-phosphate dehydrogenase (37); a functional role in translation of HAV of this protein also remains to be determined. The HAV proteinase 3C was found to bind to the
59terminus of the HAV genome (26).
In this study, we investigated the possibility of PCBP2
bind-ing within the HAV 59NCR. Competition RNA mobility assays
conducted with recombinant PCBP2 (rPCBP2), 32P-labeled
PV stem-loop IV RNA, and the HAV 59NCR as competitor
RNA clearly demonstrate the interaction of PCBP2 with the
59NCR of HAV. Binding appears to be restricted to the 59
-terminal region of the 59NCR. Furthermore, the depletion of
PCBP2 from HeLa cell extract, used to support cell-free trans-lation of HAV RNA, resulted in reduced transtrans-lational capacity
of the 59NCR, which was restored by the addition of rPCBP2.
The data support a functional role for PCBP2 in HAV RNA translation.
MATERIALS AND METHODS
Plasmid construction.Plasmids pPsP* (18), pT220-460 (11), pH139-4977 (18), and pT7-HAV1 (19) were described previously. Plasmids constructed in this study were derived from pHAV/7(735) (41), containing the complete cDNA sequence of the cell culture-adapted HAV, HM175p35, under SP6 promoter
control. It contains substitutions at nt 735 and 736, changing the AUG codon at this position to GCG in order to direct translation initiation to the following start codon at nt 741, which is the predominant one used in infected cells (41).
The numbering used to describe nucleotide positions in the HAV genome in the following plasmids is adjusted to the sequence of the wild-type HAV strain HM175 (10).
The HAV subclone pH749 contains the entire 59NCR. It was derived from pHAV/7(735) by digestion with XbaI downstream of the 59NCR (nt 744) and in the polylinker sequence. Separation of the fragments by agarose gel electro-phoresis and gel purification (QIAquick; Qiagen Inc.) eliminated the coding sequences and the 39NCR. Ligation of the remaining 3.5-kb fragment yielded the subclone pH749, harboring nt 1 to 749 of the HAV genome under SP6 promoter control. From this parental subclone we generated a series of 59-end and internal deletions to construct pH148-749, pH292-749, pH533-749, pH667-749, and pH749D158-292, using the following strategies.
To delete nt 1 to 147, 1 to 291, 1 to 532, or 1 to 666, respectively, from the 59
end of the HAV 59NCR, plasmid pH749 was digested with HindIII, which cuts 7 nt downstream the SP6 promoter start site, and with an appropriate second restriction endonuclease within the 59NCR (SspI, XcmI, NsiI, or EcoO109I, respectively). The protruding ends were treated with the Klenow fragment of DNA polymerase I, and the cDNAs lacking the particular 59-end sequences were used for blunt-end ligation after gel purification. If SspI and EcoO109I, restric-tion endonucleases which cut pH749 twice, were used, a two-point ligarestric-tion was performed. Each resulting plasmid, pH148-749, pH292-749, pH533-749, and pH667-749, harbors the nucleotide sequences of the HAV genome indicated by numbers in the name.
The internal deletion from nt 158 to 292 within the HAV 59NCR was created by digestion of pH749 with PstI and XcmI, which cut in the HAV 59NCR at nt 157 and 284, respectively. The large 3.4-kb fragment was isolated by agarose gel electrophoresis and gel purification following incubation with Klenow enzyme to produce blunt ends. Ligation with T4 DNA ligase generated plasmid pH749D158-292.
The construct pH667-4977, lacking most of the 59NCR, was generated by digestion of pH667-749 with XbaI and EcoRI, followed by insertion of the 4.2-kb
XbaI-EcoRI fragment comprising nt 744 to 4977 of pT7-HAV1. The resulting
plasmid contained nt 667 to 4977 of the HAV genome under SP6 promoter control.
All plasmids were propagated by standard procedures in Escherichia coli DH5acells, using ampicillin selection. The appropriate nucleotide sequences throughout the junctions of the ligated fragments were verified by analysis using Sequenase version 2.0 (U.S. Biochemical) and [35S]dATP according to the
man-ufacturer’s instructions.
RNA synthesis in vitro.Circular plasmids were linearized with the appropriate restriction enzyme. The linear DNAs were then used to direct transcription in a 20-ml reaction mixture in the presence of [3H]UTP with a MEGAscript kit
(Ambion) according to the supplier’s instructions to produce transcripts up to 750 nt in length. For production of transcripts longer than 750 nt, a 50-ml reaction mixture was prepared by using a MAXIscript kit (Ambion) in the presence of [3H]UTP. Following transcription, the reaction mixtures were
treated with RNase-free DNase I at 37°C for 15 min. Transcripts were phenol-chloroform extracted prior to analysis of the integrity of the RNA by agarose gel electrophoresis and quantification by measuring the incorporated3H. T7 or SP6
RNA polymerase was used, depending on the promoter sequence present in the plasmid to allow the production of sense transcripts. Although all RNAs were3H
labeled, they are referred to as unlabeled RNA in competition experiments. Plasmid pT220-460 was linearized with HindIII and used for synthesis of32
P-labeled PV stem-loop IV RNA by using a MAXIscript T7 kit (Ambion). Twenty microcuries of [32P]CTP (3,000 Ci/mmol) was added to 20ml of transcription
reaction mixture. Unincorporated nucleotides were removed by passage over a G-50 Sephadex Quick Spin column (Boehringer Mannheim), and measurement of incorporated32P was used for quantification of the transcripts.
Preparation of cell extracts. RSW from HeLa S3 cells and FRhK-4 cell monolayers were prepared as described by Brown and Ehrenfeld (7). The RSW was concentrated by precipitation with ammonium sulfate, and the 0 to 40% saturation precipitate (A-cut) was resuspended and dialyzed against 20 mM Tris (pH 7.4)–100 mM KCl–0.2 mM EDTA–7 mMb-mercaptoethanol–5% glycerol overnight at 4°C. Total protein concentration in the A-cut preparations was determined by the Bradford method, using a commercial kit (Pierce).
HeLa S3 cell extract (S10) and HeLa S3 cell translation initiation factors (RSW) for cell-free translation reactions were prepared as described previously (3, 42) and combined (HeLa S10-RSW extract) in a relative volume-to-volume ratio (74:26) used for translation assays in vitro prior to translation and deple-tion.
Western blot analysis.Protein extracts mixed with Laemmli sample buffer were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes (Schleicher & Schuell). The blot was incubated for 60 min with Tris-buffered saline containing 3% milk and then for 60 min with the primary antibody raised against rPCBP2 (6). After a wash with Tris-buffered saline containing 0.05% Tween 20, the membrane was incubated for 30 min with alkaline phosphatase-conjugated goat anti-rabbit immunoglobulin G Fc (Promega) as the secondary antibody, followed by color detection as recommended by the manufacturer.
on November 9, 2019 by guest
http://jvi.asm.org/
Depletion of HeLa cell extract.PCBP2-depleted extract was prepared by RNA affinity column chromatography using PV stem-loop IV RNA as described pre-viously (6). Briefly, 2 ml of combined HeLa S10-RSW extract was depleted of PCBP2 by passage over immobilized PV stem-loop IV RNA, derived from pT220-460, at 4°C. Approximately 1 mg of PV stem-loop IV RNA was covalently coupled to preswollen CNBr-activated Sepharose 4B (Pharmacia) in 0.1 M NaCH3CO2for 24 h at 4°C (1-ml packed-bed volume). The column was
equil-ibrated with translation buffer containing 20 mM HEPES-KOH (pH 7.4), 91 mM KCH3CO2, 2.6 mM KCl, 3.4 mM Mg(CH3CO2)2, and 4 mM dithiothreitol. The
flowthrough was passed four additional times over the column. To reuse the column, proteins bound to PV stem-loop IV RNA were eluted with 1 M KCl, and the column was washed with 2 M KCl and stored in translation buffer. Control extract (mock-depleted HeLa S10-RSW extract) was passaged five times over a column without coupled RNA.
Translation assay in vitro.Translation assays of various viral transcripts were performed in HeLa S10-RSW extracts (2, 42) that were either mock depleted or depleted of PCBP2 (6). The cell-free translation assays were performed in a 10-ml reaction mixture containing 500 ng of transcript in the presence of a mixture of [35S]methionine and [35S]cysteine. The final concentration of
potas-sium in the translation mixture was reduced to 100 mM instead of 138 mM, adjusted for optimal translation of HAV RNA. Reactions performed with PCBP2-depleted extracts were supplemented with 200 nM rPCBP2 for restora-tion assays. The control assays were supplemented with the appropriate buffer. After incubation for 90 min at 30°C, samples were diluted in Laemmli sample buffer and analyzed by SDS-PAGE (11% gel). Following electrophoresis, the gel was stained with Coomassie blue, dried and subsequently subjected to autora-diography.
RNA mobility shift assay and competition assay.RNA mobility shift assays were performed as previously described (4) except that rPCBP2 was used instead of protein extract from HeLa cells. Briefly, purified rPCBP2 (2 pmol), generously provided by Todd B. Parsley (6, 33), was preincubated with tRNA (1mg/ml)– heparin (0.25mg/ml)–40 mM KCl–5 mM HEPES (pH 7.4)–2 mM MgCl2–0.1 mM
EDTA–2 mM dithiothreitol for 10 min at 30°C before addition of 0.05 pmol of
32P-labeled RNA or labeled RNA mixed with competitor RNA and further
incubation for 15 min at 30°C. Glycerol was added to a final concentration of 10%, and the RNA-protein complex was separated from free RNA on a 4% nondenaturing polyacrylamide gel in Tris-borate-EDTA buffer at 50 V and 4°C overnight. The gel had been preequilibrated for 30 min at 280 V and 4°C. Following electrophoresis, the gel was dried and exposed to X-ray film (Kodak BioMax, Kodak Corp.).
RESULTS
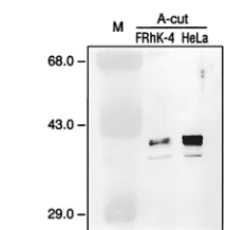
Presence of PCBP2 in FRhK-4 cells. The cellular factor PCBP2, which has been shown to play a role in translation of PV RNA, was isolated from RSW of HeLa cells, a cell line which has not been reported to be permissive for HAV repli-cation. We wanted to determine whether PCBP2 was required for translation of HAV RNA. Although we have recently shown that HAV RNA translated efficiently in HeLa cells (18), it was important to determine whether PCBP2 was expressed in cells known to support growth of HAV, such as the fetal rhesus monkey cell line FRhK-4. The A-cut of FRhK-4 cell RSW was prepared and compared with the HeLa cell A-cut for detection of PCBP2. Equal amounts of protein from both A-cut preparations were separated by SDS-PAGE and sub-jected to analysis by Western blot using antiserum raised against rPCBP2. As shown in Fig. 1, the anti-rPCBP2 serum recognized the same protein pattern, thought to represent phosphorylated variants of PCBP2 (28) in both HeLa and FRhK-4 cells. The immunoreactive proteins appeared to be present at a lower concentration in FRhK-4 cell RSW A-cut than in the corresponding HeLa cell fraction. There may be less PCBP2 present in FRhK-4 cells than in HeLa cells; how-ever, there may be weaker recognition of FRhK-4 PCBP2 by this antiserum, since it was raised against the human PCBP2 and FRhK-4 cells are of rhesus monkey origin. In addition, the immunoreactive proteins could be a combination of PCBP2 and PCBP1, since the antiserum used is cross-reactive (6). PCBP1 and PCBP2 differ in amino acid sequence by only about 10% and are closely related members of the same gene family (1, 25).
PCBP2 binding to the HAV 5*NCR.To determine whether
PCBP2 binds to the 59NCR of HAV RNA and to assess its
binding affinity to HAV sequences compared to PV sequences, competition RNA mobility shift assays were performed. As shown previously (5), PV stem-loop IV RNA, representing nt
220 to 460 from the PV 59NCR, forms a stable complex with
purified rPCBP2 (Fig. 2, lanes 1 and 2). Addition of increasing amounts of unlabeled HAV RNA containing nt 1 to 744
(H1-744), representing the entire 59NCR of HAV, to the binding
reaction resulted in competition for binding of rPCBP2 with PV stem-loop IV RNA (Fig. 2, lanes 8 to 12). Effective com-petition for rPCBP2 binding was achieved in this assay with a
20-fold molar excess of the unlabeled HAV RNA over 32
P-labeled PV stem-loop IV RNA. The same ratio of unP-labeled
RNA of the PV 59NCR (P1-816) to32P-labeled PV stem-loop
IV RNA was necessary for competition for rPCBP2 (Fig. 2,
lanes 3 to 7), indicating that the HAV 59NCR interacts with
PCBP2 and that the affinity for binding of PCBP2 to the HAV
59NCR is similar to that for binding of PCBP2 to the PV
59NCR.
Mapping of PCBP2 binding site within the HAV 5*NCR.
PCBP2 has been shown to bind preferentially to homopoly-meric poly(rC) sequences (28), and specifically to the PV
stem-loop IV RNA (5) and the 59-terminal 90 nt of the PV 59NCR
(stem-loop I) (14, 33), that form a cloverleaf-like structure. Lesser binding activity was demonstrated to poly(rG) and
poly(U) (28). The 59NCR of HAV contains two
pyrimidine-rich tracts located at the 59end between nt 99 to 138 (pY1 [38])
and at the 39end between nt 712 to 726 (pY2 [38]), in addition
[image:3.612.372.485.71.186.2]to several stable stem-loop structures, which could serve as
FIG. 1. Western blot analysis of HeLa and FRhK-4 cell fractions for PCBP2. Thirty micrograms of protein from the A-cut preparations derived from FRhK-4 (lane 2) and HeLa S3 (lane 3) cell RSW were resolved by SDS-PAGE and transferred to a nitrocellulose membrane for immunoblot analysis using serum raised against rPCBP2. Immunoreactive proteins recognized by anti-rPCBP2 antiserum are indicated to the right; molecular masses of marker pro-teins (M; lane 1) are indicated in kilodaltons to the left.
FIG. 2. Mobility and competition assay of RNAs from PV and HAV 59NCR.
32P-labeled PV stem-loop IV RNA (nt 220 to 460) was incubated without (lane
1) and with rPCBP2 (lane 2) and separated on a 4% native polyacrylamide gel. Competition for complex formation between32P-labeled PV stem-loop IV RNA
and rPCBP2 was tested in the presence of 1- to 40-fold molar excesses of transcripts representing the PV 59NCR (P1-816; lanes 3 to 7) or transcripts representing the HAV 59NCR (H1-744; lanes 8 to 12). Free labeled PV stem-loop IV (stIV) RNA and the RNA-protein complex are indicated to the left.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.316.538.575.659.2]potential binding sites for PCBP2. To identify the region(s) of
the HAV 59NCR that interacts with the cellular protein
PCBP2, a series of RNA fragments of the HAV 59NCR,
de-picted in Fig. 3, were used as unlabeled competitor RNAs
during RNA mobility shift assays with 32P-labeled PV
stem-loop IV RNA and rPCBP2.
In the first analysis, an RNA segment comprising the 59half
of the 59NCR (nt 1 to 354) was used as competitor RNA at
10-and 40-fold molar excesses over the radioactive labeled probe. As shown in Fig. 4A, lanes 5 and 6, the first 354 nt of the HAV
59NCR (H1-354) showed competition for rPCBP2 binding to
PV stem-loop IV RNA similar to that seen with the entire
59NCR (H1-744) (Fig. 4A, lanes 3 and 4). The RNA
repre-senting the entire HAV 59NCR competes slightly better than
the RNA of the fragment H1-354, likely due to an increase in stability of the relevant RNA structure when presented in the context of the larger RNA. A similar effect was observed for
competition between the intact PV 59NCR and an RNA
rep-resenting only the stem-loop IV domain (4). The region H1-354 includes several predicted stem-loop structures thought to be upstream of the IRES (nt 1 to 94), the pY1 pyrimidine-rich tract, and stem-loop IIIa,b structure (nt 150 to 315), consid-ered an essential part of the IRES (8). Further fragmentation
(nt 1 to 160 or 140 to 248) of the 59 half resulted in poor
competition (data not shown), possibly due to disruption of the overall structure.
Another set of RNAs used for competition represented
in-creasing lengths of 39-end sequences of the HAV 59NCR.
Transcripts representing small RNA fragments from the 39
third, nt 533 to 744 derived from pH533-749 or nt 677 to 744 derived from pH667-749, which include the pY2
pyrimidine-rich tract of the HAV 59NCR, failed to compete for binding of
rPCBP2 (data not shown), as did transcripts of the 39half of
the HAV 59NCR derived from pH292-749 (Fig. 4B, lanes 7 to
10). Although, H292-744 failed to displace the PV probe from the PCBP2 complex, some interaction between these macro-molecules appears to occur at high HAV RNA concentrations, causing the appearance of new bands with an increased shift in mobility. Finally, transcripts comprising nt 148 to 744 of the
HAV 59NCR (H148-744) showed a weak competition for
bind-ing of rPCBP2, causbind-ing complete displacement of the PV stem-loop IV RNA only at a molar ratio of 80:1 (Fig. 4B, lanes 3 to 6). Nonspecific binding of rPCBP2 was tested by using RNA transcribed from pGEM digested with RsaI as the competitor. There was no competition for rPCBP2 binding detectable even at a molar ratio of competitor RNA to specific probe of 80:1 (data not shown and shown previously [4]).
The results of the competition assays indicated that the
binding site for PCBP2 within the 59NCR of HAV is in the 59
half of the 59NCR and that the 39half of the HAV 59NCR does
not compete for PCBP2 binding. To localize the binding region
more precisely, an RNA fragment of the HAV 59NCR with an
internal deletion from nt 158 to 292 (H744D158-292) was
pre-pared. This construct removes the stem-loop IIIa,b structure. Figure 4B, lanes 11 to 14, shows the results obtained in assays
using the internally deleted HAV 59NCR as competitor RNA.
H744D158-292 competed efficiently for binding of rPCBP2
with PV stem-loop IV RNA at a molar ratio of 10:1, indicating
that the first 157 nt, including the 59-terminal pyrimidine-rich
tract of the HAV 59NCR, are involved in binding of PCBP2.
An RNA fragment comprising only the 59-terminal 160 nt
demonstrated direct binding to rPCBP2 by mobility shift (data not shown), confirming the interaction of this region of the
HAV 59NCR with PCBP2.
Effect of PCBP2 on translation in vitro of HAV RNA.To determine whether the presence of PCBP2 affected translation
directed by the 59NCR of HAV RNA, translation assays were
[image:4.612.134.459.69.316.2]performed with cell extracts which had been depleted of
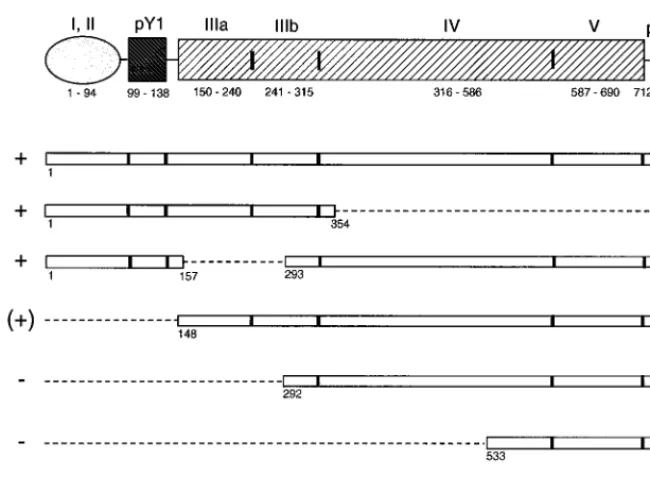
FIG. 3. Schematic representation of RNA fragments of the HAV 59NCR used as competitor RNAs in mobility shift assays with32P-labeled PV stem-loop IV RNA and rPCBP2. Open bars represent sequences of the 59NCR comprising nucleotides indicated below each fragment. The dotted lines indicate the extent of deletions in the 59NCR. The vertical lines indicate the borders of the predicted stem-loop structures I to V and the pyrimidine-rich tracts at the 59(pY1) and 39(pY2) ends of the 59NCR (8, 38) as depicted above the diagram of the RNA fragments.1, (1), or2at the left indicates whether the particular RNA fragment competes, competes weakly, or does not compete with PV stem-loop IV RNA for binding of rPCBP2.
on November 9, 2019 by guest
http://jvi.asm.org/
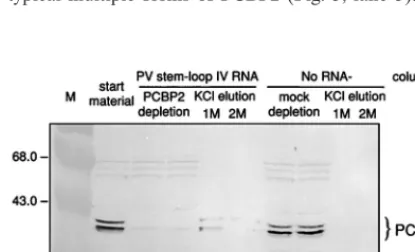
PCBP2. Successful depletion of PCBP2 from combined HeLa S10-RSW extracts by PV stem-loop IV RNA affinity column chromatography was verified by Western blot analysis with anti-rPCBP2 serum (Fig. 5). The RNA affinity column re-moved PCBP2 from the cell extract almost completely (Fig. 5, lanes 3 and 4). Minor residual amounts of PCBP2 were vari-ably detectable after depletion in different preparations. Elu-tion of the column with 1 M KCl recovered PCBP2 from the PV stem-loop IV RNA column. The high-salt eluate showed the typical multiple forms of PCBP2 (Fig. 5, lane 5). PCBP2
was not removed when cell extract was passed over a column without coupled RNA (mock-depleted HeLa S10-RSW ex-tract) (Fig. 5, lanes 7 and 8), and subsequently no PCBP2 was recovered in the 1 or 2 M KCl eluate from the control column (Fig. 5, lanes 9, 10).
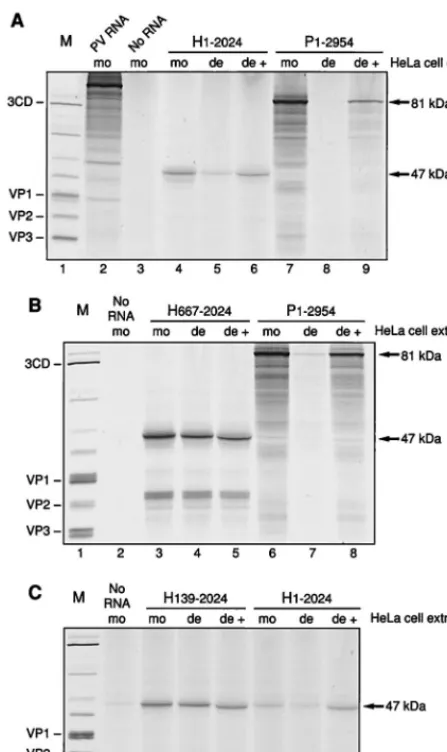
HAV RNA was translated in the PCBP2-depleted and the mock-depleted HeLa S10-RSW extracts. Truncated HAV RNA representing nt 1 to 2024 derived from pT7-HAV1 was used to program PCBP2-depleted HeLa S10-RSW extracts or mock-depleted control extract; for comparison, truncated PV RNA representing nt 1 to 2954 derived from pPsP* was trans-lated. To simplify the pattern of translation products, tran-scripts were prepared from plasmids linearized so as to encode only capsid protein sequences. Figure 6A shows the translation products obtained in this assay derived from HAV RNA H1-2024 (lanes 4 to 6) and from PV RNA P1-2954 (lanes 7 to 9) with predicted molecular masses of 47 and 81 kDa, respec-tively. Translation of HAV RNA was decreased in PCBP2-depleted extract (lane 5) compared to mock-PCBP2-depleted extract (lane 4). Restoration of this decreased translation efficiency was achieved by addition of rPCBP2 (Fig. 6A, lane 6), indicat-ing a specific effect of PCBP2 for translation of HAV. How-ever, depletion of PCBP2 did not cause as complete a reduc-tion in translareduc-tion as that seen using PV RNA (Fig. 6A, lanes 7 to 9). Thus, HAV RNA requires PCBP2 for efficient trans-lation, although it can still be translated at a low level when the majority of the cellular factor PCBP2 has been depleted from the extract. The incomplete restoration of translation with rPCBP2 in this assay for PV RNA may be due to addition of nonsaturating amounts of rPCBP2. In the case of HAV RNA, saturating amounts of rPCBP2 were used, as determined in a dose-response assay (data not shown).
The specificity of PCBP2 stimulation of HAV IRES-medi-ated translation was tested by construction of an HAV RNA
with a short, relatively unstructured 59NCR, designed to be
translated by scanning from the 59end. HAV RNA containing
nt 667 to 2024, lacking almost the entire 59NCR, was then
translated in PCBP2-depleted or mock-depleted HeLa S10-RSW extract. Figure 6B shows that translation of HAV RNA comprising nt 667 to 2024 occurred with the same efficiency in the two extracts (lanes 3 and 4), and the addition of rPCBP2 to the depleted extract showed no stimulation (lane 5), verifying that the effect of PCBP2 on HAV RNA is a function of IRES-mediated translation. Supplementation of PCBP2-depleted ex-tract with rPCBP2 yielded a translation product which ap-peared to migrate slightly faster than the products obtained with mock- or PCBP2-depleted extract alone. This effect was caused by the high protein concentration of added rPCBP2, which migrates at about 43 kDa, and could be reproduced by adding increasing amounts of rPCBP2 to mock-depleted or PCBP2-depleted extract (data not shown). Translation prod-ucts (about 30 kDa) of the H667-2024 RNA smaller than the expected 47-kDa protein were detectable, most probably due to aberrant internal initiations on this RNA.
The mobility shift competition assays described above
indi-cated that the first 157 nt of the HAV 59NCR mediated
bind-ing of PCBP2. Transcripts containbind-ing nt 139 to 2024 with a disrupted potential binding site for rPCBP2 were prepared and used to program HeLa cell extract which was mock depleted or depleted of PCBP2. As predicted, depletion of PCBP2 had no effect on translation of this RNA (Fig. 6C; compare lanes 3 and 4). Similarly, addition of exogenous rPCBP2 to the depleted extract had no influence on the efficiency of translation (lane 5). Consistent with the assignment of sequences or structures
formed by the 59-terminal 138 nt in the binding of PCBP2,
[image:5.612.51.292.70.262.2]HAV RNA comprising nt 139 to 744, used as competitor RNA
FIG. 4. RNA mobility shift and competition assay with32P-labeled PV
stem-loop IV RNA (nt 220 to 460), rPCBP2, and various HAV 59NCR fragments as competitor RNA analyzed on a 4% native polyacrylamide gel. (A) Labeled RNA (PV nt 220 to 460) was incubated with rPCBP2 in the absence (lane 2) or in the presence of unlabeled competitor HAV RNA comprising nt 1 to 744 (lanes 3 and 4) or comprising the 59half of the HAV 59NCR from nt 1 to 354 (lanes 5 and 6) at the molar excess indicated. Lane 1 was loaded with32P-labeled PV stem-loop
IV (stIV) RNA only. (B) Labeled RNA (PV nt 220 to 460) was incubated with rPCBP2 in the presence of competitor RNA comprising nt 148 to 744 (lanes 3 to 6), nt 292 to 744 (lanes 7 to 10), and nt 1 to 744 with the internal deletion from nt 158 to 292 (lanes 11 to 14). Labeled PV stem-loop IV RNA alone was loaded in lane 1 and as complex with rPCBP2 in lane 2. Free RNA and the RNA-protein complex are indicated to the left.
FIG. 5. Western blot analysis of combined HeLa S10-RSW extract, using antiserum raised against rPCBP2. The extract was depleted of PCBP2 by RNA affinity column chromatography with immobilized PV stem-loop IV RNA (nt 220 to 460). Lane 2, untreated extract; lanes 3 and 4, extract passaged five times over the RNA affinity column; lanes 5 and 6, 1 and 2 M KCl eluates; lanes 7 to 10, control extract passaged over a column without any immobilized RNA (mock-depleted extract) and its high-salt eluates. Molecular masses of marker proteins (M; lane 1) are indicated in kilodaltons to the left; immunoreactive proteins recognized by anti-rPCBP2 antiserum are indicated to the right.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.66.274.526.652.2]in the mobility shift assay with PV stem-loop IV RNA and rPCBP2, also failed to compete for rPCBP2 binding at a molar ratio of competitor RNA to specific probe of 20:1 (data not
shown). RNA of the entire HAV 59NCR competes efficiently
at this molar concentration. Comparison of the efficiencies of translation of HAV RNAs H1-2024 and H139-2024 in the control, mock-depleted extract showed that deletion of the
59-terminal 138 nt of the HAV 59NCR resulted in more
effi-cient translation under these conditions than the HAV RNA
containing the entire 59NCR (Fig. 6C, lanes 3 and 6). This
effect has been observed previously (18, 44). Interestingly, ad-dition of rPCBP2 to the depleted extract restored translation
of H1-2024 to levels similar to those seen when the PCBP2 binding sequences were not present (lane 8 and 5).
DISCUSSION
Internal initiation of translation mediated by picornavirus IRES structures may be facilitated by multiple host cell pro-teins in addition to the canonical initiation factors (for a re-view, see reference 40). Addition of PTB to a set of purified initiation factors enhanced the formation of the translation initiation complex of a variant of EMCV IRES-driven
trans-lation (24, 34). The 59NCR of HAV RNA has been shown to
bind specifically to several proteins derived from the RSW of BS-C-1, FRhK-4, and HeLa cells (9), although most of these proteins were not identified. A 39-kDa protein that was present in BS-C-1 and FRhK-4, but not in HeLa, cells and that
cross-linked to several regions of the HAV 59NCR and bound
specifically to stem-loop IIIa of the HAV 59NCR was
subse-quently identified as glyceraldehyde-3-phosphate dehydroge-nase (37). It was not determined whether these interactions provide a functional role in HAV translation.
A cellular RNA binding protein, PCBP2, has been shown
previously to bind to stem-loop IV in the 59NCR of PV RNA,
and this binding appears essential for IRES-mediated transla-tion of PV RNA (5, 6). PCBP2 binds also to sequences in the
59-terminal cloverleaf-like structure (stem-loop I) formed by
the first 90 nt of PV RNA and has been implicated in replica-tion of PV RNA (14, 33). To determine whether PCBP2 had a potential role in HAV RNA translation, we investigated the
binding of rPCBP2 to the 59NCR of HAV RNA and examined
the translation of this RNA in an extract depleted of PCBP2. We made use of the specific binding between PV stem-loop IV RNA and rPCBP2 in a competition RNA mobility shift assay
with competitor RNAs representing the 59NCR of HAV or
segments of this 59NCR.
The results showed that the HAV 59NCR competes for
binding of PCBP2 to PV stem-loop IV and that the binding site
is located within the 59third of the HAV 59NCR. Maintenance
of the overall structure of the 59NCR proved to be important
for PCBP2 interaction, since most fragments of this RNA region tested for direct binding to PCBP2 resulted in only weak and nonspecific retarded migration of the RNA (data not shown). Additional cellular or viral proteins could also affect the binding of PCBP2 to HAV RNA, a possibility not tested in
this study. The 59-terminal 157-nt sequence of the HAV
59NCR was able to specifically compete for binding of PCBP2
if presented in the context of the HAV 59NCR harboring an
internal deletion from nt 158 to 292. The 59-terminal
se-quences of the HAV 59NCR contain a pyrimidine-rich tract
from nt 99 to 138 which may provide a favorable binding site for PCBP2, since PCBP2 was originally characterized as
dem-onstrating strong binding to poly(rC) (28, 43). The 39half of
the 59NCR is predicted to form several stem-loop structures
and also contains a pyrimidine-rich tract from nt 712 to 726. These sequences did not compete for binding of PCBP2. Even the very small RNA probe from nt 667 to 744, designed to expose the pyrimidine-rich tract, did not bind PCBP2. Thus, the cellular protein PCBP2 seems to interact with HAV RNA containing the pyrimidine-rich tract from nt 99 to 138 yet not
with the pyrimidine-rich tract located at the 39end between nt
712 to 726. If the pyrimidine-rich tract is the site of PCBP2 binding, the secondary structure and the primary sequence context surrounding the tract may be responsible for the dif-ferential binding of the two pyrimidine-rich tracts. Additional investigations are necessary to specify the exact binding site.
[image:6.612.57.281.71.447.2]The affinities of binding to PCBP2 appear quite comparable
FIG. 6. Translation of truncated HAV and PV RNAs driven by different 59NCR regions in HeLa S10-RSW extracts. The H1-2024 (lanes 4 to 6) and P1-2954 (lanes 7 to 9) (A), H667-2024 (lanes 3 to 5) and P1-2954 (lanes 6 to 8) (B), and H139-2024 (lanes 3 to 5) and H1-2024 (lanes 6 to 8) (C) transcripts were used to program translation extracts in the presence of [35S]methionine and
[35S]cysteine. Translation assays were carried out in mock-depleted extract (mo),
PCBP2-depleted extract (de), and PCBP2-depleted extract supplemented with rPCBP2 (de1). PV-encoded polypeptides obtained from PV-infected HeLa cells labeled with [35S]methionine served as protein marker (M; lane 1) and are
identified on the left. HeLa S10-RSW extract programmed with 100 ng of PV RNA isolated from purified PV virions was used as internal translation control (panel A, lane 2). Due to the short incubation time used in this assay, the PV polyprotein is mainly uncleaved. HeLa cell extract programmed with no RNA is analyzed in lane 3 of panel A and lanes 2 of panels B and C. The predicted protein sizes of the translation products are indicated to the right.
on November 9, 2019 by guest
http://jvi.asm.org/
between the PV and the HAV 59NCRs, as indicated by the
equal molar concentrations of the two 59NCRs required to
compete with the stem-loop IV probe (Fig. 2). However, the binding sites on the two picornaviral RNAs appear to be quite
different. The PV 59NCR binds PCBP2 at stem-loop IV, a
large and complex internal domain (nt 235 to 440) within the IRES element (5), as well as at a subloop of the cloverleaf-like
structure (nt 1 to 90) at the 59terminus of the 59NCR upstream
of the IRES (33). The HAV RNA binding site could be
local-ized only to within the 59one-third of the 59NCR (nt 1 to 157).
Neither a terminal cloverleaf-like structure nor anything re-sembling the downstream PV stem-loop IV domain is pre-dicted to form in this region of the HAV RNA, and the
pyri-midine-rich tract including the 59-terminal region in HAV
RNA that is likely responsible for its PCBP2 binding is not present in PV RNA. Despite this apparent difference in bind-ing sites, translation of both PV and HAV RNAs is signifi-cantly reduced in HeLa cell extracts depleted of PCBP2. These observations suggest that the role of PCBP2 in facilitating IRES-mediated initiation of translation is to stabilize the com-plex secondary and tertiary structure of the IRES, rather than to identify similar structural determinants in the RNA. It is, of course, also possible that PCBP2 needs only to bind some-where within or near the IRES in order to direct other factors
to the initiation complex. Binding of PCBP2 to the 59end of
the HAV RNA may also have a role in RNA replication, as is the case for PV RNA (14, 15, 33); however, no obvious simi-larity between the two viruses exists in this region.
Previous results showed that a 39-kDa protein identified as glyceraldehyde-3-phosphate dehydrogenase bound to the
pre-dicted stem-loop IIIa (nt 155 to 235) of the HAV 59NCR (37).
This 39-kDa protein could also be cross-linked to nt 1 to 148
and nt 597 to 740 of the HAV 59NCR (9). The cross-linking
activity was detected in RSW of BS-C-1 and FRhK-4, but not HeLa, cells. We have shown that PCBP2, also an approxi-mately 39-kDa protein, binds to HAV RNA by a competition RNA mobility shift assay. PCBP2 is present in both HeLa and FRhK-4 cells. It is not known whether PCBP2 cross-links to HAV RNA. It appears, therefore, that the observed binding of these two similar-sized proteins may not be related.
Translation assays with PCBP2-depleted HeLa cell extract demonstrated a requirement for the binding of PCBP2 for efficient translation of HAV RNA, when the translation is
directed by the complete HAV 59NCR. Addition of rPCBP2 to
the depleted extracts specifically restored the reduced tion activity of the PCBP2-depleted extract. Residual transla-tion activity of HAV RNA was still detectable in the PCBP2-depleted extract and was greater than that detected with PV RNA in the same extract preparation. It is possible that HAV RNA needs a lower concentration of PCBP2 for functional translation than PV RNA, and since the depletion procedure fails to remove all of the PCBP2, there may be enough PCBP2 remaining in the extract to support a low level of translation of HAV RNA. Alternatively, removal of PCBP2 from the trans-lation extract may allow another trans-acting factor present
only in minor amounts to interact with the HAV 59NCR in a
manner to support viral RNA translation on a low level.
The finding that translation promoted by the HAV 59NCR
lacking the 59-terminal 138 nt, a sequence which is upstream of
the HAV IRES element, was independent of PCBP2 and more efficient was rather surprising. Considering that the possible role of PCBP2 binding is to promote the correct
three-dimen-sional organization of the 59NCR of HAV RNA necessary to
facilitate internal ribosome entry, it may be that the HAV
59NCR sequences lacking the 59-terminal 138 nt adopt the
correct structure spontaneously without the need for PCBP2
binding to promote the appropriate folding. This may suggest another reason for the finding that HAV RNA containing the
entire 59NCR translates, albeit poorly, in PCBP2-depleted
ex-tracts. This activity might reflect a minor population of HAV RNA molecules already in a conformation folded in the ap-propriate secondary structure without the help of PCBP2. Re-cently, Gamarnik and Andino (15) proposed that the RNA
element at the 59end of the PV genome contains overlapping
signals for translation and replication of RNA. Their model suggests that the binding of the cellular protein PCBP to the RNA enhances translation and stimulates production of viral proteins. When sufficient concentrations of the viral protein 3CD become available, both 3CD and PCBP are bound, re-sulting in repression of translation and formation of a ribonu-cleoprotein complex that initiates viral RNA replication. The data presented here suggest a similar model for regulation of
the utilization of the HAV 59NCR RNA as a template for
translation and transcription. In the absence of other viral or cellular proteins, PCBP binding facilitates translation, perhaps by stabilizing the IRES conformation. Binding of other
pro-teins to the 59-terminal RNA sequence might displace or alter
PCBP binding so as to favor a replication structure over a
translation structure. The presence of the 59-terminal RNA
sequences were observed to inherently downregulate transla-tion, perhaps because of their potential to form the replication-specific structure.
ACKNOWLEDGMENTS
We thank Todd B. Parsley for generously providing purified rPCBP2, and we are grateful to Ollie Richards and Yolanda Bell for helpful discussions.
This work was supported by Public Health Service grants AI26350 and AI12387 from the National Institutes of Health.
REFERENCES
1. Aasheim, H.-C., T. Loukianova, A. Deggerdal, and E. B. Smeland. 1994. Tissue specific expression and cDNA structure of a human transcript encod-ing a nucleic acid bindencod-ing [oligo(dC)] protein related to the pre-mRNA binding protein K. Nucleic Acids Res. 22:956–964.
2. Barton, D. J., and J. B. Flanegan. 1993. Coupled translation and replication of poliovirus RNA in vitro: synthesis of functional 3D polymerase and infectious virus. J. Virol. 67:822–831.
3. Barton, D. J., E. P. Black, and J. B. Flanegan. 1995. Complete replication of poliovirus in vitro: preinitiation RNA replication complexes require soluble cellular factors for the synthesis of VPg-linked RNA. J. Virol. 69:5516–5527. 4. Blyn, L. B., R. Chen, B. L. Semler, and E. Ehrenfeld. 1995. Host cell proteins binding to domain IV of the 59noncoding region of poliovirus RNA. J. Virol.
69:4381–4389.
4a.Blyn, L. B., and J. Graff. Unpublished data.
5. Blyn, L. B., K. M. Swiderek, O. Richards, D. C. Stahl, B. L. Semler, and E.
Ehrenfeld.1996. Poly(rC) binding protein 2 binds to stem-loop IV of the poliovirus RNA 59 noncoding region: identification by automated liquid chromatography-tandem mass spectrometry. Proc. Natl. Acad. Sci. USA
93:11115–11120.
6. Blyn, L. B., J. Towner, B. L. Semler, and E. Ehrenfeld. 1997. Requirement of poly(rC) binding protein 2 for translation of poliovirus RNA. J. Virol.
71:6243–6246.
7. Brown, B. A., and E. Ehrenfeld. 1979. Translation of poliovirus RNA in vitro: changes in cleavage pattern and initiation sites by ribosomal salt wash. Virology 97:396–405.
8. Brown, E. A., S. P. Day, R. W. Jansen, and S. M. Lemon. 1991. The 59
nontranslated region of hepatitis A virus RNA: secondary structure and elements required for translation in vitro. J. Virol. 65:5828–5838. 9. Chang, K. H., E. A. Brown, and S. M. Lemon. 1993. Cell type-specific
proteins which interact with the 59nontranslated region of hepatitis A virus RNA. J. Virol. 67:6716–6725.
10. Cohen, J. I., J. R. Ticehurst, R. H. Purcell, A. Buckler-White, and B. M.
Baroudy.1987. Complete nucleotide sequence of wild-type hepatitis A virus: comparison with different strains of hepatitis A virus and other picornavi-ruses. J. Virol. 61:50–59.
11. Dildine, S. L., and B. L. Semler. 1992. Conservation of RNA-protein inter-actions among picornaviruses. J. Virol. 66:4364–4376.
12. Dorner, A. J., B. L. Semler, R. J. Jackson, R. Hanecak, E. Duprey, and E.
on November 9, 2019 by guest
http://jvi.asm.org/
Wimmer.1984. In vitro translation of poliovirus RNA: utilization of internal initiation sites in reticulocyte lysate. J. Virol. 50:507–514.
13. Ehrenfeld, E. 1996. Initiation of translation by picornavirus RNAs, p. 549– 573. In J. W. B. Hershey, M. B. Mathews, and N. Sonenberg (ed.), Trans-lational control. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
14. Gamarnik, A. V., and R. Andino. 1997. Two functional complexes formed by KH domain containing proteins with the 59noncoding region of poliovirus RNA. RNA 3:882–892.
15. Gamarnik, A. V., and R. Andino. 1998. Switch from translation to RNA replication in a positive-stranded RNA virus. Genes Dev. 12:2293–2304. 16. Gauss-Mu¨ller, V., D. Ju¨rgensen, and R. Deutzmann. 1991. Autoproteolytic
cleavage of recombinant 3C proteinase of hepatitis A virus. Virology 182: 861–864.
17. Glass, M. J., and D. F. Summers. 1993. Identification of a trans-acting activity from liver that stimulates HAV translation in vitro. Virology 193: 1047–1050.
18. Graff, J., and E. Ehrenfeld. 1998. Coding sequences enhance internal initi-ation of transliniti-ation by hepatitis A virus RNA in vitro. J. Virol. 72:3571–3577. 19. Harmon, S. A., O. C. Richards, D. F. Summers, and E. Ehrenfeld. 1991. The 59-terminal nucleotides of hepatitis A virus RNA, but not poliovirus RNA, are required for infectivity. J. Virol. 65:2757–2760.
20. Harmon, S. A., W. Updike, X.-Y. Jia, D. F. Summers, and E. Ehrenfeld. 1992. Polyprotein processing in cis and in trans by hepatitis A virus 3C protease cloned and expressed in Escherichia coli. J. Virol. 66:5242–5247. 21. Jackson, R. J., S. L. Hunt, C. L. Gibbs, and A. Kaminski. 1994. Internal
initiation of translation of picornavirus RNAs. Mol. Biol. Rep. 19:147–159. 22. Jia, X.-Y., G. Schepper, D. Brown, W. Updike, S. Harmon, O. Richards, D.
Summers, and E. Ehrenfeld.1991. Translation of hepatitis A virus RNA in vitro: aberrant internal initiations influenced by 59noncoding region. Virol-ogy 182:712–722.
23. Jia, X.-Y., E. Ehrenfeld, and D. F. Summers. 1991. Proteolytic activity of hepatitis A virus 3C protein. J. Virol. 65:2595–2600.
24. Kaminski, A., and R. J. Jackson. 1998. The polypyrimidine tract binding protein (PTB) requirement for internal initiation of translation of cardiovi-rus RNAs is conditional rather than absolute. RNA 4:626–638.
25. Kiledjian, M., X. Wang, and S. A. Liebhaber. 1995. Identification of two KH domain proteins in thea-globin mRNP stability complex. EMBO J. 14:4357– 4364.
26. Kusov, Y., and V. Gauss-Mu¨ller. 1997. In vitro RNA binding of the hepatitis A virus proteinase 3C (HAV 3Cpro) to secondary structure elements within the 59terminus of the HAV genome. RNA 3:291–302.
27. Le, S. Y., A. Sidiqui, and J. V. Maizel. 1996. A common structural core in the internal ribosome entry site of picornavirus, hepatitis c virus, and pestivirus. Virus Genes 12:135–147.
28. Leffers, H., K. Delgaard, and J. E. Celis. 1995. Characterization of two major cellular poly(rC)-binding human proteins, each containing three K-homolo-gous (KH) domains. Eur. J. Biochem. 230:447–453.
29. Meerovitch, K., J. Pelletier, and N. Sonenberg. 1989. A cellular protein that binds to the 59-noncoding region of poliovirus RNA: implications for inter-nal translation initiation. Genes Dev. 3:1026–1034.
30. Meerovitch, K., Y. V. Svitkin, H. S. Lee, F. Lejbkowicz, D. J. Kenan, E. K. L.
Chan, V. I. Agol, J. D. Keene, and N. Sonenberg.1993. La autoantigen enhances and corrects aberrant translation of poliovirus RNA in reticulocyte lysate. J. Virol. 67:3798–3807.
31. Niepmann, M. 1996. Porcine polypyrimidine tract-binding protein stimulates translation initiation at the internal ribosome entry site of foot-and-mouth-disease virus. FEBS Lett. 388:39–42.
32. Niepmann, M., A. Petersen, K. Meyer, and E. Beck. 1997. Functional in-volvement of polypyrimidine tract-binding protein in translation initiation complexes with the internal ribosome entry site of foot-and-mouth disease virus. J. Virol. 71:8330–8339.
33. Parsley, T. B., J. S. Towner, L. B. Blyn, E. Ehrenfeld, and B. L. Semler. 1997. Poly (rC) binding protein 2 forms a ternary complex with the 59-terminal sequences of poliovirus RNA and the viral 3CD proteinase. RNA 3:1124– 1134.
34. Pestova, T., C. U. T. Hellen, and I. N. Shatsky. 1996. Canonical eukaryotic initiation factors determine initiation of translation by internal ribosomal entry. Mol. Cell. Biol. 16:6859–6869.
35. Schultheiss, T., Y. Y. Kusov, and V. Gauss-Mu¨ller. 1994. Proteinase 3C of hepatitis A virus (HAV) cleaves the HAV polyprotein P2-P3 at all sites including VP1/2A and 2A/2B. Virology 198:275–281.
36. Schultheiss, T., W. Sommergruber, Y. Kusov, and V. Gauss-Mu¨ller. 1995. Cleavage specificity of purified recombinant hepatitis A virus 3C proteinase on natural substrates. J. Virol. 69:1727–1733.
37. Schultz, D. E., C. C. Hardin, and S. M. Lemon. 1996. Specific interaction of glyceraldehyde 3-phosphate dehydrogenase with the 59-nontranslated RNA of hepatitis A virus. J. Biol. Chem. 271:14134–14142.
38. Shaffer, D. R., E. A. Brown, and S. M. Lemon. 1994. Large deletion muta-tions involving the first pyrimidine-rich tract of the 59nontranslated RNA of human hepatitis A virus define two adjacent domains associated with distinct replication phenotypes. J. Virol. 68:5568–5578.
39. Siomi, H., M. J. Matunis, W. M. Michael, and G. Dreyfuss. 1993. The pre-mRNA binding K protein contains a novel evolutionary conserved motif. Nucleic Acids Res. 21:1193–1198.
40. Stewart, S. R., and B. L. Semler. 1998. RNA determinants of picornavirus cap-independent translation initiation. Semin. Virol. 8:242–255.
41. Tesar, M., S. A. Harmon, D. F. Summers, and E. Ehrenfeld. 1992. Hepatitis A virus polyprotein synthesis initiates from two alternative AUG codons. Virology 186:609–618.
42. Todd, S., J. S. Towner, and B. L. Semler. 1997. Translation and replication properties of the human rhinovirus genome in vivo and in vitro. Virology
229:90–97.
43. Wang, X., M. Kiledjian, I. M. Weiss, and S. A. Liebhaber. 1995. Detection and characterization of a 39untranslated region ribonucleoprotein complex associated with humana-globin mRNA stability. Mol. Cell. Biol. 15:1769– 1777.
44. Whetter, L. E., S. P. Day, O. Elroy-Stein, E. A. Brown, and S. M. Lemon. 1994. Low efficiency of the 59nontranslated region of hepatitis A virus RNA in directing cap-independent translation in permissive monkey kidney cells. J. Virol. 68:5253–5263.
45. Zhang, Y., and G. G. Kaplan. 1998. Characterization of replication-compe-tent hepatitis A virus constructs containing insertions at the N terminus of the polyprotein. J. Virol. 72:349–357.