©
DOI: 10.1534/genetics.104.040352
The
Bombyx mori
Karyotype and the Assignment of Linkage Groups
Atsuo Yoshido,* Hisanori Bando,* Yuji Yasukochi
†and Ken Sahara*
,1*Division of Applied Bioscience, Graduate School of Agriculture, Hokkaido University, Sapporo, 060-8589, Japan and †National Institute of Agrobiological Sciences, Tsukuba, 305-3934, Japan
Manuscript received December 26, 2004 Accepted for publication February 15, 2005
ABSTRACT
Lepidopteran species have a relatively high number of small holocentric chromosomes (Bombyx mori, 2n⫽56). Chromosome identification has long been hampered in this group by the high number and by the absence of suitable markers like centromere position and chromosome bands. In this study, we carried out fluorescencein situhybridization (FISH) on meiotic chromosome complements using geneti-cally mappedB. moribacterial artificial chromosomes (BACs) as probes. The combination of two to four either green or red fluorescence-labeled probes per chromosome allowed us to recognize unequivocally each of the 28 bivalents of theB. morikaryotype by its labeling pattern. Each chromosome was assigned one of the already established genetic linkage groups and the correct orientation in the chromosome was defined. This facilitates physical mapping of any other sequence and bears relevance for the ongoingB. mori genome projects. Two-color BAC-FISH karyotyping overcomes the problem of chromosome recognition in organisms where conventional banding techniques are not available.
T
HE silkworm,Bombyx mori, is one of the model or- apsed, and display chromomere patterns (Traut1976), but are still insufficient for general mapping purposes. ganisms in genetic research, second among insectsIn this study, we used pachytene chromosome com-only to the fruit fly,Drosophila melanogaster. It is an
eco-plements and fluorescence in situ hybridization with nomically important species with⬎3000 known strains
bacterial artificial chromosome probes (BAC-FISH), (Yamamoto 2000) and ⬎400 mutations reported for
which has recently been established inB. mori(Sahara silkworms, corresponding toⵑ230 mapped genes or loci
et al. 2003b) to identify all B. mori chromosomes and (Doira1983). Linkage groups have been established for
assign them to respective linkage groups. The basic re-gene mutants (Fujii et al. 1998) and densely spaced
quirements to achieve this goal were already fulfilled. RAPD (Promboonet al. 1995;Yasukochi1998, 1999),
BAC libraries, together consisting of 36,864 clones, have RFLP (Shi et al. 1995), and AFLP (Tan et al. 2001)
been constructed from two strains (Wuet al. 1999), and markers. Whole-genome sequencing projects are well
dense genetic map data, based on genes and RAPD under way (Mitaet al. 2004;Xiaet al. 2004).
Neverthe-markers (Yasukochi 1998, 1999), were available. We less, knowledge of the karyotype is still in its infancy.
screened the BAC libraries for suitable clones and deter-The chromosome number (n⫽28, Kawaguchi1928;
mined the loci on Yasukochi’s RAPD map. Using these 2n⫽56,Kawamura1979) is known and some progress
BACs we identified all B. mori autosomes and con-has been made with respect to the identification of
structed the complete karyotype ofB. mori. the sex chromosomes (Trautet al. 1999;Saharaet al.
2003a) but there has been no general basis for chromo-some identification and physical mapping.
MATERIALS AND METHODS Bombyx shares this problem with other moths and
butterflies (Lepidoptera). They are cytogenetically char- Isolation and genetic mapping of BAC clones:The two-step acterized by possessing small and numerous holokinetic PCR screening described inYasukochi(2002) was employed to isolate BACs that represent suitable loci of all of the 28 chromosomes. The chromosomes lack primary
constric-linkage groups ofB. mori. A BAC library (Wuet al. 1999) con-tions and are rather uniform in size during mitotic
meta-structed fromB. moristrain p50 with average insert size of 134.5 phase. No banding technique has yet been found to dif- kb was used for the PCR screening. Nine STS primer sets were ferentiate the chromosomes. Conditions are better for designed to isolate BACs with known genes [M24370⫹J04829, meiotic chromosomes, especially those in the pachytene AB007831, X04223, AB011497, D85134, D86601, AF287267, AB010825, andB. moriprothoracicotropic hormone (Shimada
stage when chromosomes are extended, pairwise
syn-et al. 1994)]. Partial sequencing was performed in another 60 BACs and STS primer sets designed from the resultant sequences. The STS primers amplify polymorphic DNA frag-ments between p50 and C108. Linkage analysis using these 1Corresponding author: Division of Applied Bioscience, Graduate
STSs was performed in the manner described previously School of Agriculture, Hokkaido University N9, W9, Kita-ku, Sapporo,
060-8589, Japan. E-mail: [email protected] (Yasukochi 1998) to determine the loci of the BACs. We
isolated 7 additional BACs and ascertained their map positions indeed hybridized to the Z chromosome, which was by means of two independent but closely linked RAPD mark- identified independently as the pairing partner of the ers. All BACs are listed in Table 1.
W chromosome (Figure 1A). The W chromosome had
Chromosome preparation:Pachytene chromosome
prepa-been painted with genomicin situ hybridization (GISH) rations fromB. moristrain p50 were carried out according to
Saharaet al. (1999, 2003b). Briefly, the ovaries of last instar (Saharaet al. 2003b). The result shows that our BAC-larvae were dissected in an insect saline (Glaser1917) and FISH procedure reliably identifies the Z chromosome pretreated in hypotonic solution (83 mmKCl and 17 mmNaCl; and that the chromosomal sites of the BACs
corre-Marec and Traut 1993) followed by fixation in Carnoy’s
sponded well with their loci on the RAPD map of linkage fluid (ethanol, chloroform, acetic acid, 6:3:1). Cells were
disso-group 1 (Figure 1, B and C). ciated in 60% acetic acid and spread on a glass slide placed on
a heating plate at 50⬚. The preparations were passed through a In this manner, all 27 autosomes were identified by graded ethanol series (70, 80, and 98%) and stored in the two to four red or green double-dot signals (one dot
freezer (–30⬚) until further use. per homolog) depending on the number of Cy3- (red)
Probe labeling and BAC-FISH:BAC-FISH was carried out
or fluorescein- (green) labeled linkage group-specific according to the method described inSaharaet al. (2003b)
BACs used as probes. The chromosomes are shown in with slight modifications. Briefly, BAC-containing clones were
cultured in LB medium containing 20g/ml chlorampheni- Figure 2 with each bivalent arranged together with the col at 37⬚for 16 hr. DNA was extracted with a Plasmid Midi kit corresponding linkage map oriented with position 0 cM (QIAGEN, Tokyo). DNA labeling was done by nick translation at the top. In a few cases, we produced yellow double-using the Invitrogen nick translation system (Invitrogen,
dot signals. This was intended when we mixed Cy3- and Tokyo) with Cy3-dCTP (Amersham, Tokyo) or
fluorescein-fluorescein-labeled probe from the same BAC (Figure 12-dCTP (Perkin Elmer, Boston).
After removal from the freezer, chromosome preparations 2, bivalents of linkage groups 14 and 15). The same were passed through an ethanol series and air dried. Denatur- effect was caused inadvertently by two differently labeled ation was done at 72⬚for 3.5 min in 70% formamide, 2⫻SSC. BACs with overlapping hybridization signals (Figure 2, The probe cocktail for one slide consisted of 100 ng labeled
bivalents of linkage groups 18 and 20). We never de-BAC, 25 g sonicated salmon sperm DNA (Sigma-Aldrich,
tected double-dot signals on other autosomes. The W Tokyo) and 10g (for single chromosome identification) or
100g (for karyotyping) sonicatedB. morimale genomic DNA chromosome, however, displayed extra signals with in 10l hybridization solution (50% formamide, 10% dextran many of the autosome-specific BAC probes (see below). sulfate, 2⫻SSC). After incubation in a moist chamber at 37⬚ In most bivalents, there was good correspondence of for 3 days, slides were washed at 62⬚in 0.1⫻SSC containing 1%
the labeling pattern on the chromosome with the deter-Triton X-100. The slides were counterstained and mounted in
mined positions on the respective linkage map. Excep-antifade [0.233 g 1,4-diazabicyclo(2.2.2)-octane, 1 ml 0.2m
Tris-HCl, pH 8.0, 9 ml glycerol] containing 0.5g/ml DAPI tions are BACs 8H2A in LG8, 3A3C in LG13, 5E8C in (4⬘,6-diamidino-2-phenylindole; Sigma-Aldrich). LG16, and 3H6F in LG18, which mapped in the correct
Image processing and measurement: Black-and-white im- order but not in the expected distance from one
an-ages were taken with a Photometrics CoolSNAP CCD camera
other (Figure 2). attached to a Leica DMRE HC fluorescence microscope,
The chromosomes were routinely stained with DAPI. through the A, L5, and N2.1 filters of the fluorescence filter
set. Pseudocoloring and superimposing of the images were We found that two of the 28 bivalents, those correspond-done using Adobe Photoshop, version 7.0. Routinely, red col- ing to linkage groups 11 and 24 (Figure 2), could also be oring was used for Cy3, green for fluorescein, and light blue reliably discriminated by the DAPI pattern. The bivalent for DAPI images.
corresponding to linkage group 11 was easily recogniz-Chromosome length was measured by using free software,
able by the attached nucleolus, which divided the chro-ImageJ (http://rsb.info.nih.gov/ij/index.html). The results
presented are average lengths of measurements repeated five mosome into two arms (Figure 2). The arm ratio,ⵑ2:1
times. (the end of the long arm corresponding to the proximal
end of the linkage map), was similar to that given in previous reports (Rasmussen 1976; Traut 1976). In RESULTS
the bivalent corresponding to linkage group 24, a seg-ment ofⵑ10% of the chromosome length was deeply
Selection of BAC clones:TheB. moriBAC library of
Wuet al. (1999) was screened to identify suitable clones stained with DAPI. This conspicuous, presumably het-erochromatic, segment was located at approximately for BAC-FISH mapping. Sixty-nine BACs were isolated
with polymorphic STSs whose loci were confirmed by two-thirds of the chromosome length (Figure 2, LG24). The DAPI-positive segment had already been detected linkage analysis on the same population of 166 F2
indi-viduals described inYasukochi(1998), and 7 BACs were in a previous study when it was recognized as a conspicu-ous autosomal heterochromatic block strongly painted isolated with two independent and closely linked RAPD
markers (Yasukochi1998). In total, we selected 76 BACs, by GISH (Saharaet al. 2003b).
KaryotypingB. mori:For karyotyping, we used a probe
2–6 from each of the 28 linkage groups (Table 1).
Identification of individual chromosomes:To test the cocktail consisting of 62 BACs labeled with Cy3-dCTP
and/or fluorescein-dCTP. The respective BACs and system, we hybridized the three BACs 9A5H, 14I7D, and
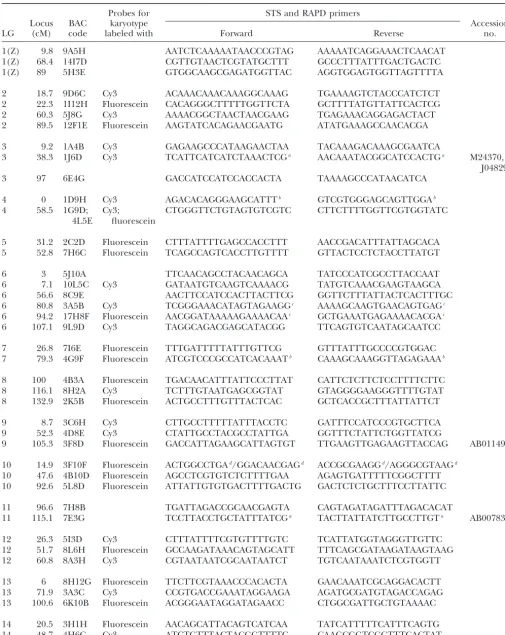
TABLE 1
B. moriBAC clones and STS primers used in this study
Probes for STS and RAPD primers
Locus BAC karyotype Accession
LG (cM) code labeled with Forward Reverse no.
1(Z) 9.8 9A5H AATCTCAAAAATAACCCGTAG AAAAATCAGGAAACTCAACAT
1(Z) 68.4 14I7D CGTTGTAACTCGTATGCTTT GCCCTTTATTTGACTGACTC
1(Z) 89 5H3E GTGGCAAGCGAGATGGTTAC AGGTGGAGTGGTTAGTTTTA
2 18.7 9D6C Cy3 ACAAACAAACAAAGGCAAAG TGAAAAGTCTACCCATCTCT
2 22.3 1I12H Fluorescein CACAGGGCTTTTTGGTTCTA GCTTTTATGTTATTCACTCG
2 60.3 5J8G Cy3 AAAACGGCTAACTAACGAAG TGAGAAACAGGAGACTACT
2 89.5 12F1E Fluorescein AAGTATCACAGAACGAATG ATATGAAAGCCAACACGA
3 9.2 1A4B Cy3 GAGAAGCCCATAAGAACTAA TACAAAGACAAAGCGAATCA
3 38.3 1J6D Cy3 TCATTCATCATCTAAACTCGa AACAAATACGGCATCCACTGa M24370,
J04829
3 97 6E4G GACCATCCATCCACCACTA TAAAAGCCCATAACATCA
4 0 1D9H Cy3 AGACACAGGGAAGCATTTb GTCGTGGGAGCAGTTGGAb
4 58.5 1G9D; Cy3; CTGGGTTCTGTAGTGTCGTC CTTCTTTTGGTTCGTGGTATC
4L5E fluorescein
5 31.2 2C2D Fluorescein CTTTATTTTGAGCCACCTTT AACCGACATTTATTAGCACA
5 52.8 7H6C Fluorescein TCAGCCAGTCACCTTGTTTT GTTACTCCTCTACCTTATGT
6 3 5J10A TTCAACAGCCTACAACAGCA TATCCCATCGCCTTACCAAT
6 7.1 10L5C Cy3 GATAATGTCAAGTCAAAACG TATGTCAAACGAAGTAAGCA
6 56.6 8C9E AACTTCCATCCACTTACTTCG GGTTCTTTATTACTCACTTTGC
6 80.8 3A5B Cy3 TCGGGAAACATAGTAGAAGGc AAAAGCAAGTGAACAGTGAGc
6 94.2 17H8F Fluorescein AACGGATAAAAAGAAAACAAc GCTGAAATGAGAAAACACGAc
6 107.1 9L9D Cy3 TAGGCAGACGAGCATACGG TTCAGTGTCAATAGCAATCC
7 26.8 7I6E Fluorescein TTTGATTTTTATTTGTTCG GTTTATTTGCCCCGTGGAC
7 79.3 4G9F Fluorescein ATCGTCCCGCCATCACAAATb CAAAGCAAAGGTTAGAGAAAb
8 100 4B3A Fluorescein TGACAACATTTATTCCCTTAT CATTCTCTTCTCCTTTTCTTC
8 116.1 8H2A Cy3 TCTTTGTAATGAGCGGTAT GTAGGGGAAGGGTTTTGTAT
8 132.9 2K5B Fluorescein ACTGCCTTTGTTTACTCAC GCTCACCGCTTTATTATTCT
9 8.7 3C6H Cy3 CTTGCCTTTTTATTTACCTC GATTTCCATCCCGTGCTTCA
9 52.3 4D8E Cy3 CTATTGCCTACGCCTATTGA GGTTTCTATTCTGGTTATCG
9 105.3 3F8D Fluorescein GACCATTAGAAGCATTAGTGT TTGAAGTTGAGAAGTTACCAG AB011497
10 14.9 3F10F Fluorescein ACTGGCCTGAd/GGACAACGAGd ACCGCGAAGGd/AGGGCGTAAGd
10 47.6 4B10D Fluorescein AGCCTCGTGTCTCTTTTGAA AGAGTGATTTTTCGGCTTTT
10 92.6 5L8D Fluorescein ATTATTGTGTGACTTTTGACTG GACTCTCTGCTTTCCTTATTC
11 96.6 7H8B TGATTAGACCGCAACGAGTA CAGTAGATAGATTTAGACACAT
11 115.1 7E3G TCCTTACCTGCTATTTATCGa TACTTATTATCTTGCCTTGTa AB007831
12 26.3 5I3D Cy3 CTTTATTTTCGTGTTTTGTC TCATTATGGTAGGGTTGTTC
12 51.7 8L6H Fluorescein GCCAAGATAAACAGTAGCATT TTTCAGCGATAAGATAAGTAAG
12 60.8 8A3H Cy3 CGTAATAATCGCAATAATCT TGTCAATAAATCTCGTGGTT
13 6 8H12G Fluorescein TTCTTCGTAAACCCACACTA GAACAAATCGCAGGACACTT
13 71.9 3A3C Cy3 CCGTGACCGAAATAGGAAGA AGATGCGATGTAGACCAGAG
13 100.6 6K10B Fluorescein ACGGGAATAGGATAGAACC CTGGCGATTGCTGTAAAAC
14 20.5 3H1H Fluorescein AACAGCATTACAGTCATCAA TATCATTTTTCATTTCAGTG
14 48.7 4H6C Cy3 ATCTGTTTACTACGGTTTTC CAAGCGGTCGGTTTCACTAT
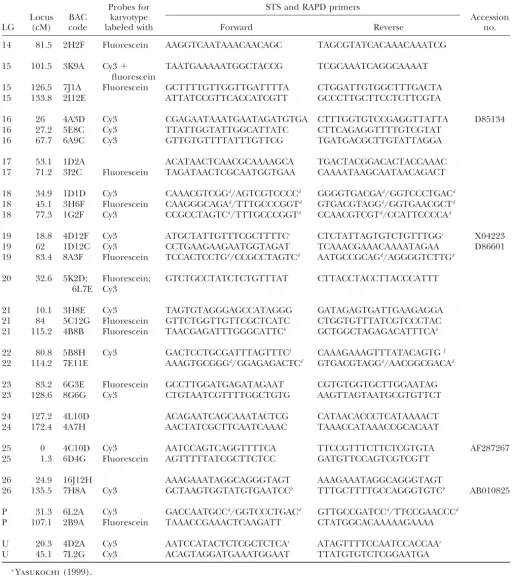
TABLE 1
(Continued)
Probes for STS and RAPD primers
Locus BAC karyotype Accession
LG (cM) code labeled with Forward Reverse no.
14 81.5 2H2F Fluorescein AAGGTCAATAAACAACAGC TAGCGTATCACAAACAAATCG
15 101.5 3K9A Cy3⫹ TAATGAAAAATGGCTACCG TCGCAAATCAGGCAAAAT
fluorescein
15 126.5 7J1A Fluorescein GCTTTTGTTGGTTGATTTTA CTGGATTGTGGCTTTGACTA
15 133.8 2I12E ATTATCCGTTCACCATCGTT GCCCTTGCTTCCTCTTCGTA
16 26 4A3D Cy3 CGAGAATAAATGAATAGATGTGA CTTTGGTGTCCGAGGTTATTA D85134
16 27.2 5E8C Cy3 TTATTGGTATTGGCATTATC CTTCAGAGGTTTTGTCGTAT
16 67.7 6A9C Cy3 GTTGTGTTTTATTTGTTCG TGATGACGCTTGTATTAGGA
17 53.1 1D2A ACATAACTCAACGCAAAAGCA TGACTACGGACACTACCAAAC
17 71.2 3I2C Fluorescein TAGATAACTCGCAATGGTGAA CAAAATAAGCAATAACAGACT
18 34.9 1D1D Cy3 CAAACGTCGGd/AGTCGTCCCCd GGGGTGACGAd/GGTCCCTGACd
18 45.1 3H6F Fluorescein CAAGGGCAGAd/TTTGCCCGGTd GTGACGTAGGd/GGTGAACGCTd
18 77.3 1G2F Cy3 CCGCCTAGTCd/TTTGCCCGGTd CCAACGTCGTd/CCATTCCCCAd
19 18.8 4D12F Cy3 ATGCTATTGTTTCGCTTTTCe CTCTATTAGTGTCTGTTTGGc X04223
19 62 1D12C Cy3 CCTGAAGAAGAATGGTAGAT TCAAACGAAACAAAATAGAA D86601
19 83.4 8A3F Fluorescein TCCACTCCTGd/CCGCCTAGTCd AATGCCGCAGd/AGGGGTCTTGd
20 32.6 5K2D; Fluorescein; GTCTGCCTATCTCTGTTTAT CTTACCTACCTTACCCATTT
6L7E Cy3
21 10.1 3H8E Cy3 TAGTGTAGGGAGCCATAGGG GATAGAGTGATTGAAGAGGA
21 84 5C12G Fluorescein GTTCTGGTTGTTCGCTCATC CTGGTGTTTATCGTCCCTAC
21 115.2 4B8B Fluorescein TAACGAGATTTGGGCATTCb GCTGGCTAGAGACATTTCAb
22 80.8 5B8H Cy3 GACTCCTGCGATTTAGTTTCf CAAAGAAAGTTTATACAGTGf
22 114.2 7E11E AAAGTGCGGGd/GGAGAGACTCd GTGACGTAGGd/AACGGCGACAd
23 83.2 6G3E Fluorescein GCCTTGGATGAGATAGAAT CGTGTGGTGCTTGGAATAG
23 128.6 8G6G Cy3 CTGTAATCGTTTTGGCTGTG AAGTTAGTAATGCGTGTTCT
24 127.2 4L10D ACAGAATCAGCAAATACTCG CATAACACCCTCATAAAACT
24 172.4 4A7H AACTATCGCTTCAATCAAAC TAAACCATAAACCGCACAAT
25 0 4C10D Cy3 AATCCAGTCAGGTTTTCA TTCCGTTTCTTCTCGTGTA AF287267
25 1.3 6D4G Fluorescein AGTTTTTATCGCTTCTCC GATGTTCCAGTCGTCGTT
26 24.9 16J12H AAAGAAATAGGCAGGGTAGT AAAGAAATAGGCAGGGTAGT
26 135.5 7H8A Cy3 GCTAAGTGGTATGTGAATCCb TTTGCTTTTGCCAGGGTGTCb AB010825
P 31.3 6L2A Cy3 GACCAATGCCd/GGTCCCTGACd GTTGCCGATCCd/TTCCGAACCCd
P 107.1 2B9A Fluorescein TAAACCGAAACTCAAGATT CTATGGCACAAAAAGAAAA
U 20.3 4D2A Cy3 AATCCATACTCTCGCTCTCAe ATAGTTTTCCAATCCACCAAe
U 45.1 7L2G Cy3 ACAGTAGGATGAAATGGAAT TTATGTGTCTCGGAATGA
Figure 1.—Detection of the WZ bivalent (Z is linkage group 1) using Z-BACs 9A5H, 14I7D, 5H3E, and a female-derived whole-genomic probe for the W chromosome (A). Red signals are from Z-BACs and green sig-nals are from the whole-genomic probe. The chromosomes are counterstained with DAPI (light blue). The arrow points to the heterochromatic block of an au-tosomal bivalent (see text). N, nucleolus. Bar, 10m. (B and C) Linkage map with RAPD loci (in black) from Yasukochi
(1998) and the map positions of BACs (in red) corresponding to the signals on the Z chromosome.
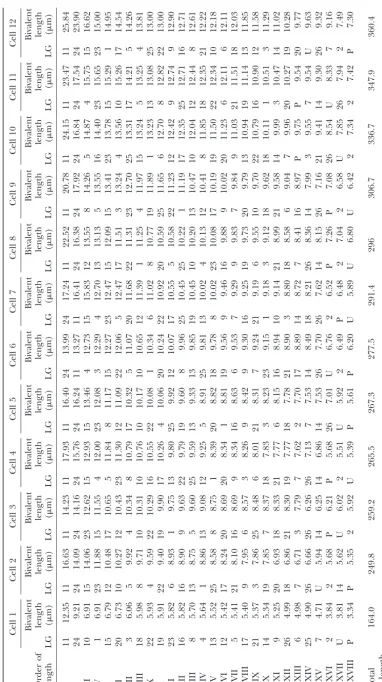
well-spread pachytene nuclei could be discriminated BAC-FISH had previously been applied to fine map-ping in extended chromatin fibers (Weier2001). The (Figure 3A). We arranged the bivalents of one cell
(Fig-ure 3B, cell 4 in Table 2) according to their average method commonly used to identify chromosomes by FISH is either chromosome painting (Cremer et al. length calculated from 12 karyotyped complements.
The probe cocktail did not include markers for the 1988;Lichteret al. 1988;Pinkelet al. 1988) or by using centromeric chromosome-specific repetitive sequence chromosomes corresponding to linkage groups 1(Z),
11, and 24. Those corresponding to linkage groups 11 probes (Hizume et al. 2002;Vischiet al. 2003), which are not available in B. mori. Karyotyping of the whole and 24 could be recognized from the DAPI pattern
alone. And, Z-specific probes proved unnecessary as the chromosome complement has been achieved in the hu-man and in the mouse with sophisticated color schemes probe cocktail reliably produced extra label on the W
chromosome to which the Z chromosome was synapsed and probe mixes of five and seven different fluoro-in the pachytene stage. phores. M-FISH (Speicher et al. 1996; Jentsch et al. Length measurements of 12 karyotyped complements 2001) and spectral karyotyping (Liyanageet al. 1996; are listed in Table 2. It is obvious that the various chro- Schro¨ ck et al. 1996) are two of these methods. We mosomes occupy similar but not constant positions in had decided against chromosome painting. Probes for the order of length. Systematic individual length changes chromosome painting are usually generated from sorted during pachytene development are not apparent but chromosomes (Van Dilla et al. 1986; Collins et al. cannot be excluded. Considering the rather shallow 1991;Vooijset al. 1993), from microdissected chromo-length gradient especially in the middle part of theB. somes (Guan et al. 1994), or from a densely spaced
mori complement, however, variation in position may chromosome-specific BAC array (Lysak et al. 2001). be merely due to measurement error (the median of Sorting chromosomes is probably not possible in Bom-standard error is 0.101 with the range from 0.019 to byx due to the similarity of chromosome sizes (see Table 0.340) and/or variation in individual chromosome 2). Microdissection of chromosomes is confronted with
spreading. the same problem if one does not wish to rely on a
single microdissected anonymous chromosome. A col-lection of 28 sufficiently dense chromosome-specific
DISCUSSION BAC arrays would have been a feasible alternative for
probe generation. But this would have been a rather The high number and small size of the chromosomes
expensive alternative with respect to costs and experi-and the absence of suitable cytogenetic markers like
mental work and it would have given less information. bands and localized centromeres have hitherto
inhib-The obvious advantage of using only a few BACs and ited chromosome identification inB. mori. We
circum-two different fluorophores per chromosome is the gen-vented the problem by (1) hybridizing selected
fluores-eration of anchor points that relate to the genetical cence-labeled BACs to the chromosomes (BAC-FISH)
map. Besides mere identification of the chromosome, and (2) using pachytene instead of mitotic
chromo-they provide a framework for physical mapping of other somes. Pachytene chromosomes have the advantage of
BACs and allow us to distinguish between the two an extended length (4.3 times longer than mitotic
chro-mosome ends. mosomes on average) and a reduced number (28
biva-The two B. mori whole-genome shotgun sequencing lents instead of 56 single mitotic chromosomes). In this
projects,Mitaet al. (2004) andXiaet al. (2004), pres-way, we were able to identify all 28 chromosomes of the
cover-Figure
2.—Identification
o
f
individual
chromosomes
a
nd
comparison
w
ith
the
linkage
map.
Note
that
the
lengths
of
chromosome
bivalents
cannot
be
compared
int
h
is
figure
a
s
they
are
derived
from
d
ifferent
cells.
Numbering
o
f
linkage
groups
follows
Fujii
et
al
.
(1998)
and
Yasukochi
(1998)
(for
d
etails
see
Yasukochi
et
al
.
2
005).
L
inkage
maps
show
RAPD
loci
from
Yasukochi
(1998)
(in
black),
including
the
m
arkers
used
to
select
the
BACs
for
F
ISH
(red,
g
reen,
o
r
yellow
text
o
n
the
right).
The
sites
of
BAC-FISH
signals
correspond
nearly
always
with
those
m
ap
positions.
For
the
BACs
used,
see
Table
1
Figure
3.—BAC-FISH
karyotype
o
f
B.
mori
.
(A)
An
oocyte
pachytene
nucleus.
Bar,
10
m.
(B)
The
bivalents
from
this
nucleus
arranged
according
to
their
lengths
together
with
their
diagnostic
macket al., 1996 Multicolor spectral karyotyping of mouse chor-age, respectively, of the genome. The karyotype and the
omosomes. Nat. Genet.14:312–315.
anchor points provided in this article afford a basis to Lysak, M. A., P. F. Fransz, H. B. AliandI. Schubert, 2001 Chromo-map contigs directly or with little experimental effort some painting inArabidopsis thaliana.Plant J.28:689–697.
Marec, F., andW. Traut, 1993 Synaptonemal complexes in female
to the respective chromosomes and linkage groups. Our
and male meiotic prophase ofEphestia kuehniella(Lepdoptera).
results already disclose that the map of 28 linkage Heredity71:394–404.
groups mainly based on RAPD markers (Yasukochi Mita, K., M. Kasahara, S. Sasaki, Y. Nagayasu, T. Yamadaet al.,
2004 The genome sequence of silkworm,Bombyx mori.DNA Res.
1998) truly reflects the chromosome status inB. mori.
11:27–35.
Since the BACs have been proven to give chromo- Pinkel, D., J. Landegent, C. Collins, J. Fuscoe, R. Segraveset al., some-specific signals, identification of individual mitotic 1988 Fluorescencein situhybridization with human chromo-some specific libraries: detection of trisomy 21 and translocation (as opposed to meiotic pachytene) chromosomes by
of chromosome 4. Proc. Natl. Acad. Sci. USA85:9138–9142.
BAC-FISH will probably also be possible inB. mori.
BAC-Promboon, A., T. Shimada, H. FujiwaraandM. Kobayashi, 1995
FISH karyotyping of mitotic chromosomes or mapping Linkage map of random amplified polymorphic DNAs (RAPDs) other probes relative to our anchor points, however, in the silkworm,Bombyx mori.Genet. Res.66:1–7.
Rasmussen, S. W., 1976 The meiotic prophase inBombyx mori
fe-will be difficult or impossible due to the small size of
males analyzed by three-dimensional reconstructions of synapto-the mitotic chromosomes. On synapto-the osynapto-ther hand, synapto-the BAC- nemal complexes. Chromosoma54:245–293.
FISH karyotyping method is probably easily adaptable Sahara, K., F. MarecandW. Traut, 1999 TTAGG telomeric re-peats in chromosomes of some insects and other arthropods. to other species with similar problems of chromosome
Chromosome Res.7:449–460.
recognition. Sahara, K., F. Marec, U. EickhoffandW. Traut, 2003a Moth sex chromatin probed by comparative genomic hybridization (CGH). We thank Walther Traut (Luebeck, Germany) for critical
discus-Genome46:339–342.
sions and Yasuhiro Yamada for technical assistance. This study was
Sahara, K., A. Yoshido, N. Kawamura, A. Ohnuma, H. Abeet al.,
partially supported by Grant-in-Aid for Scientific Research no.
2003b W-derived BAC probes as a new tool for identification
15380227 from the Japan Society for the Promotion of Science (K.S.
of the W chromosome and its aberrations inBombyx mori.
Chro-and Y.Y.) Chro-and by the Insect Technology Project from the Ministry of
mosoma112:48–55.
Agriculture, Forestry and Fisheries in Japan, no. 2102 (Y.Y.) and no. Schro
¨ ck, E., S. Du Manoir, T. Veldman, B. Schoell, J. Wienberg
2013 (K.S.). et al., 1996 Multicolor spectral karyotyping of human
chromo-somes. Science273:494–497.
Shi, J., D. G. HeckelandM. R. Goldsmith, 1995 A genetic linkage
map for the domesticated silkworm,Bombyx mori, based on
restric-LITERATURE CITED tion fragment length polymorphisms. Genet. Res.66:109–126.
Shimada, T., T. Hasegawa, K. Matsumoto, N. AguiandM.
Koba-Collins, C. C., W. L. Kuo, R. Segraves, J. C. Fuscoe, D. Pinkelet
yashi, 1994 Polymorphism and linkage analysis of the
protho-al., 1991 Construction and characterization of plasmid libraries
racicotropic hormone gene in the silkworm,Bombyx mori.Genet.
enriched in sequences from single human chromosomes.
Geno-Res.63:189–195.
mics11:997–1006.
Speicher, M. R., S. G. BallardandD. C. Ward, 1996 Karyotyping
Cremer,T., P.Lichter, J.Borden, D. C.Wardand L.Manuelidis,
human chromosomes by combinatorial multi-fluor FISH. Nat.
1988 Detection of chromosome aberrations in metaphase and
Genet.12:368–375.
interphase tumor cells byin situhybridization using chromosome
specific library probes. Hum. Genet.80:235–246. Tan, Y., C. Wan, Y. Zhu, C. Lu, Z. Xianget al., 2001 An amplified
Doira, H., 1983 Linkage maps ofBombyx moristatus quo in 1983. fragment length polymorphisms map of the silkworm. Genetics
Sericologia23:245–269. 157:1277–1284.
Fujii, H., Y. Banno, H. Doira, H. KiharaandY. Kawaguchi, 1998 Traut, W., 1976 Pachytene mapping in the female silkworm,Bombyx
Genetical Stocks and Mutations of Bombyx mori: Important Genetic moriL. (Lepidoptera). Chromosoma58:275–284.
Resources, Ed. 2. Institute of Genetic Resources, Faculty of Agricul- Traut, W., K. Sahara, T. D. OttoandF. Marec, 1999 Molecular
ture, Kyushu University, Fukuoka, Japan. differentiation of sex chromosome probed by comparative
geno-Glaser, R. W., 1917 ‘Ringer’ solutions and some notes on the physio- mic hybridization. Chromosoma108:173–180.
logical basis of their ionic composition. Comp. Biochem. Physiol. Van Dilla, M. A., L. L. Deaven, K. L. Albroght, N. A. Allen,
2:241–289. M. R. Aubuchonet al., 1986 Human chromosome specific DNA
Guan, X. Y., P. S. MeltzerandJ. M. Trent, 1994 Rapid generation libraries: construction and availability. Biotechnology4:537–552.
of whole chromosome painting probes (WCPs) by chromosome Vischi, M., I. Jurman, G. BianchiandM. Morgante, 2003
Karyo-microdissection. Genomics22:101–107. type of Norway spruce by multicolor FISH. Theor. Appl. Genet.
Hizume, M., F. Shibata, Y. MatsusakiandZ. Garajova, 2002 Chro- 107:591–597.
mosome identification and comparative karyotypic analyses of Vooijs, M., L. C.Yu, D.Tkachuk, D.Pinkel, D.Johnsonet al., 1993
fourPinusspecies. Theor. Appl. Genet.105:491–497.
Libraries for each human chromosome, constructed from
sorter-Jentsch, I., I. D. Adler, N. P. CarterandM. R. Speicher, 2001
enriched chromosomes by using linker-adaptor PCR. Am. J. Hum. Karyotyping mouse chromosomes by multiple-FISH (M-FISH).
Genet.52:586–597.
Chromosome Res.9:211–214.
Weier, H. U. G., 2001 DNA fiber mapping techniques for the
assem-Kawaguchi, E., 1928 Zytologische Untersuchungen am
Seidenspin-bly of high-resolution physical maps. J. Histochem. Cytochem.
ner und seinen Verwandten. I. Gametogenesese vonBombyx mori
49:939–948.
L. undBombyx mandarinaM. und ihres Bastarde. Z. Zellforsch.
Wu, C., S. Asakawa, N. Shimizu, S. KawasakiandY. Yasukochi,
7:519–552.
1999 Construction and characterization of bacterial artificial
Kawamura, N., 1979 Cytological studies on the mosaic silkworm
chromosome libraries from the silkworm,Bombyx mori.Mol. Gen.
induced by low temperature treatment. Chromosoma74:179–
Genet.261:698–706.
188.
Xia, Q., Z.Zhou, C.Lu, D. Cheng,F.Dai et al., 2004 A draft
Lichter, P., T.Cremer, J.Borden, L.Manuelidisand D. C.Ward,
sequence for the genome of the domesticated silkworm (Bombyx
1988 Delineation of individual human chromosomes in
meta-mori). Science306:1937–1940.
phase and interphase cells byin situsuppression hybridization
Yamamoto, T., 2000 Silkworm strains, pp. 45–49 inStrain
Mainte-using chromosome specific library probes. Hum. Genet.80:224–
nance and Databank for Life Science, edited by N.Nakatsuji. Kyori-234.
Yasukochi, Y., 1998 A dense genetic map of the silkworm,Bombyx Yasukochi, Y., L. A. Ashakumary, C. Wu, A. Yoshido, J. Nohataet mori, covering all chromosomes based on 1018 molecular mark- al., 2004 Organization of the Hox gene cluster of the silkworm,
ers. Genetics150:1513–1525. Bombyx mori: a split of the Hox cluster in a non-Drosophilainsect.
Yasukochi, Y., 1999 A simple and accurate method for generating Dev. Genes Evol.214:606–614.
co-dominant markers: an application of conformation-sensitive Yasukochi, Y., Y.Banno, K.Yamamoto, M. R.Goldsmithand H.
gel electrophoresis to linkage analysis in the silkworm. Mol. Gen. Fujii, 2005 Integration of molecular and classical linkage
Genet.261:796–802. groups of the silkworm,Bombyx mori(n⫽28). Genome (in press).
Yasukochi, Y., 2002 PCR-based screening for bacterial artificial