Biology Theses Department of Biology
5-10-2019
Region-Specific Effects Of Microglial Depletion
On Developmental Neuronal Cell Death In The
Mouse Brain
Andrew Jacobs
Follow this and additional works at:https://scholarworks.gsu.edu/biology_theses
This Thesis is brought to you for free and open access by the Department of Biology at ScholarWorks @ Georgia State University. It has been accepted for inclusion in Biology Theses by an authorized administrator of ScholarWorks @ Georgia State University. For more information, please contact
Recommended Citation
Jacobs, Andrew, "Region-Specific Effects Of Microglial Depletion On Developmental Neuronal Cell Death In The Mouse Brain." Thesis, Georgia State University, 2019.
NEURONAL CELL DEATH IN THE MOUSE BRAIN
ANDREW JACOBS
Under the Direction of Nancy Forger, PhD
ABSTRACT
Naturally occurring perinatal cell death is a major developmental process in the formation
of the brain. Microglia, the primary immunocompetent cells of the brain, have recently been
implicated in actively contributing to perinatal cell death. However, the current research is
contradictory. Some studies suggest that microglia induce neuronal death, while others suggest
that microglia protect neurons from intrinsic cell death programs. These studies utilize many
methods of manipulating microglia, they examine single regions of interest, and analyze at
different times in development. These differences in analysis may be responsible for the different
conclusions. In this study, we selectively reduced microglia brain wide via clodronate liposomes
and measured the amount of microglial reduction and the subsequent effects on cell death. We
also analyzed regional differences in cytokine expression as a possible underlying mechanism for
effects of microglia on cell death.
NEURONAL CELL DEATH IN THE MOUSE BRAIN
by
ANDREW JACOBS
A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of
Master of Science
in the College of Arts and Sciences
Georgia State University
Copyright by Andrew James Jacobs
NEURONAL CELL DEATH IN THE MOUSE BRAIN
by
ANDREW JACOBS
Committee Chair: Nancy Forger
Committee: Aaron Roseberry
Anne Murphy
Electronic Version Approved:
Office of Graduate Studies
College of Arts and Sciences
Georgia State University
DEDICATION
I dedicate this work to Dr. Alexandra Castillo-Ruiz. Without her unending mentorship
ACKNOWLEDGEMENTS
I would like to thank my thesis advisor and mentor Nancy Forger, to whom I owe most of
my progress toward any of my academic goals. I’d also like to acknowledge my committee
TABLE OF CONTENTS
ACKNOWLEDGEMENTS ... V
LIST OF TABLES ... VIII
LIST OF FIGURES ... IX
1 INTRODUCTION... 1
1.1 Background ... 1
1.2 Purpose of the Study ... 2
2 EXPERIMENT ... 3
2.1 Material and Methods ... 3
2.1.1 Animals ... 3
2.1.2 Depletion of Microglia ... 3
2.1.3 Immunohistochemistry ... 4
2.1.4 Brain Regions Analyzed... 5
2.1.5 Quantification of Iba1 and AC3 labeling... 6
2.1.6 Brain Cytokine Expression ... 6
2.1 Statistics ... 7
3 RESULTS ... 8
3.1 Clodronate liposome treatment reduces microglial cell number ... 8
3.3 Effect of clodronate treatment on microglial cell size varies by region ... 14
3.4 Elevated expression of inflammatory cytokines in the hippocampus ... 14
4 CONCLUSIONS ... 17
LIST OF TABLES
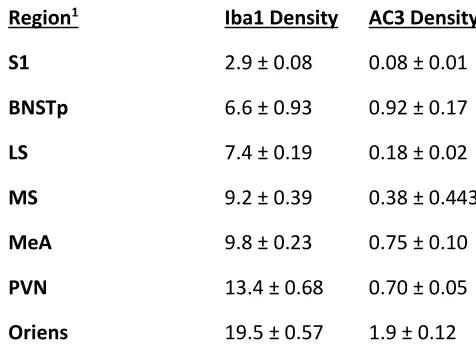
Table 3.1 Microglial density (Iba1+ cells per mm3) and cell death density (AC3+ cells per mm3)
across seven brain regions of saline-treated newborns ... 8
LIST OF FIGURES
Figure 3.1 Photomicrographs of Iba1-labeled cells in neonates receiving intracerebroventricular
injections of vehicle- (left) or clodronate-filled (right) liposomes. ... 9
Figure 3.2 Intracerebroventricular injections of clodronate liposomes significantly reduced
microglial density in all brain regions examined: the medial septum (MS), lateral septum
(LS), principal bed nucleus of the stria terminalis (BNSTp), medial amygdala (MeA),
paraventricular nucleus of the hypothalamus (PVN), oriens layer of the hippocampus, and
the primary somatosensory cortex (S1). **P < 0.01; ***P < 0.001; ****P < 0.0001. ... 10
Figure 3.3 Photomicrographs of AC3-labeled cells (black) in neonatal mice treated with
liposomes containing vehicle or clodronate. ... 12
Figure 3.4 Neonatal treatment with clodronate liposomes caused region-specific changes in
perinatal cell death. ... 13
Figure 3.5 Average microglial cell size was increased in most brain regions of clodronate-treated
pups, but was reduced in the hippocampal oriens... 14
Figure 3.6 The expression of cytokines in the septum and hippocampus of neonates treated with
saline- (black bars) or clodronate-filled (gray bars) liposomes. ... 16
1 INTRODUCTION
1.1 Background
Microglia are the primary immunocompetent cells of the brain. In adults, microglia
survey their immediate environment, are activated by infection or trauma, and respond by
releasing cytokines and chemokines (Davalos et al., 2005; Smith et al., 2012; Hristovska and
Pascual, 2015; Tucker et al., 2016). Microglia also have important roles during development
such as phagocytizing synapses and cellular debris from apoptotic neurons (Paolicelli et al.,
2011; Schwarz et al., 2012). However, the role of microglia in development has expanded to
encompass active involvement in perinatal cell death.
Cell death is a major neurodevelopmental process that occurs primarily during the first
week of life in mice, and eliminates roughly 50% of the neurons initially produced
(Oppenheim, 1985; Ahern et al., 2013; Mosley et al., 2017). Previous work in our lab shows
that the highest rate of cell death in most regions of the mouse brain directly follows birth
(Mosley et al., 2017). This high incidence of cell death correlates with the presence and
colonization of the brain by microglia (Perry et al., 1985; Ashwell, 1990).
Manipulating the presence or competence of microglia during this developmental cell
death period has been shown to influence cell death, but in contradictory ways. For instance,
culturing chick retina with microglia increases cell death (Frade and Barde, 1998). In the
hippocampus, knocking-out immunoregulatory integrin genes involved in activation of
microglia, leads to decreases in developmental cell death and in the production of neurotoxic
superoxide ions (Wakselman et al., 2008). However, in the somatosensory cortex, selective
elimination of microglia or administration of a microglia inhibitor increases cell death of
As the primary immune cell of the brain, microglia release proinflammatory cytokines
(Giulian et al., 1988) and microglia can induce neuronal death by releasing inflammatory
cytokines in vitro (Guadagno et al., 2013) and in vivo (Kaur et al., 2014). Microglial cytokine
release may be an important mechanism by which microglia exert effects on developmental
cell death.
1.2 Purpose of the Study
Each of the previous studies examining effects of microglia on cell death utilized
different methods to manipulate microglia, focused on different areas of interest, and varied with
respect to developmental age analyzed. It is not clear whether these experimental variables
contributed to the contradictory outcomes, or whether there may be actual regional differences in
the involvement of microglia in perinatal cell death. Age and regional differences in microglial
gene expression (Crain et al., 2013; Hanamsagar et al., 2017) support the latter possibility.
To address this, we depleted microglia throughout the neonatal mouse brain using
clodronate liposomes, and examined effects on cell death across multiple brain regions of each
animal. Following this evaluation, we examined cytokine expression in two regions of interest
with opposite effects on cell death to associate regional differences in cytokine expression with
cell death influences. We hypothesized that microglia actively protect or kill neurons during the
developmental cell death period in a region-specific manner and that cytokine expression reflects
2 EXPERIMENT
2.1 Material and Methods
2.1.1 Animals
All procedures were performed in accordance with the National Institutes of Health
animal welfare guidelines and approved by the Institutional Animal Care and Use Committee of
Georgia State University. Wild-type C57BL/6 mice were housed in a 12:12 light: dark cycle at
22°C with food and water available ad libitum. Cages of breeding pairs were checked daily for
births.
2.1.2 Depletion of Microglia
Microglia were depleted in newborn mice by the injection of clodronate-filled liposomes
(Encapsula NanoSciences, Brentwood, TN). Clodronate liposomes are selectively internalized by
microglia when injected into the brain or applied to brain slices, causing the demise of microglia
while sparing other cell types (Faustino et al., 2011; Kumamaru et al., 2012; Marín-Teva et al.,
2004). Cryoanesthetized newborn pups received bilateral intracerebroventricular injections of
undiluted clodronate- or saline-filled liposomes targeted to the lateral ventricles. The needle tip
of a 5 µl Hamilton syringe controlled by a micromanipulator was positioned approximately 1
mm rostral and 1 mm lateral to bregma, as described previously (Mosley et al., 2017b). An
incision was not required, as bregma could be visualized through the scalp. The needle was
lowered 2 mm beneath the surface of the skull, and a microsyringe pump (Micro4, World
Precision Instruments, Sarasota, FL) delivered injections (0.5 µL each side) over a span of 30
seconds.
In pilot work, we first determined an injection regime for microglial ablation in newborn
or vehicle-filled (N = 13) liposomes to male and female newborns on P1 (day of birth = P0) and
collecting brains 12, 24, or 48 h after injection. Immunohistochemical labeling for ionized
calcium binding adapter molecule 1 (Iba1) was performed to identify microglia, as described
below, and microglial cell density was measured in the septum, hippocampus and hypothalamus.
A consistent reduction of microglia was observed by 24 h after injection of clodronate
liposomes. A second cohort of mice was used to compare the effectiveness of injections on P1
only (N = 14) to injections on the first two days of life (P0 and P1; N = 12); in both cases
microglia were quantified 24 h after the last injection. Because the two-day injection regime was
significantly more effective at reducing microglia than injections on P1 alone (P < 0.001; not
shown), we used mice injected on P0 and P1 in all analyses below.
2.1.3 Immunohistochemistry
Brains were collected, immersion fixed in 5% acrolein (in 0.1M phosphate buffer) for 24
h, and transferred to 30% sucrose. Brains were then frozen-sectioned into four, 40 µm coronal
series that were stored in cryoprotectant at -20 °C until processed for immunohistochemistry.
One series of sections was stained for Iba1, a marker constitutively expressed by microglia
(Ahmed et al., 2007), and a second series for activated caspase 3 (AC3). Almost all AC3+ cells
in the newborn rodent brain exhibit other markers of apoptosis (Srinivasan et al., 1998;
Zacharaki et al., 2010), and AC3 labeling the forebrain is eliminated in neonatal mice lacking the
pro-death gene, Bax (Ahern et al., 2013). Staining for both markers was performed as described
previously (Strahan et al. , 2017). Briefly, antigen retrieval was performed using 0.05 M sodium
citrate and excess unreacted aldehyde was neutralized with 0.1M glycine. Tissue was incubated
in blocking solution (20% normal goat serum and 2% hydrogen peroxide), and primary antibody:
or rabbit anti-AC3 (Cell Signaling, Beverly, MA, USA; AB_2341188; 1:20,000). Sections were
subsequently incubated with goat anti-rabbit secondary antibodies (Vector Laboratories,
Burlingame, CA, USA; AB_2313606; 1:1,000 for Iba1 or 1:500 for AC3) and avidin-biotin
(Vector Laboratories; 1:1,000 for Iba1 or 1:500 for AC3). Staining was visualized using
3,3’-diaminobenzidine tetrahydrochloride, nickel sulfate, and hydrogen peroxide. Sections were then
mounted on gelatin-coated slides, dehydrated in alcohol and coverslipped. AC3-labeled sections
were counter-stained with thionin.
2.1.4 Brain Regions Analyzed
Labeled cell densities were measured in seven brain regions where we have previously
quantified cell death density during neonatal life (Ahern et al., 2013a; Mosley et al., 2017a and
unpublished data): the medial septum (MS), lateral septum (LS), principal nucleus of the bed
nucleus of the stria terminalis (BNSTp), medial amygdala (MeA), paraventricular nucleus of the
hypothalamus (PVN), oriens layer of the hippocampus (oriens), and primary somatosensory
cortex (S1). For the septum, sections were analyzed from Plate 57 in the mouse brain atlas of
Paxinos et al. (2007) to the midline crossing of the anterior commissure (Plate 59). The BNSTp
was analyzed between Plates 61-63. For the MeA, we included sections where the nucleus
appears lateral to the optic tract (Plate 70) until the point where the fimbria joined the cortex
(Plate 74). The PVN was analyzed in its entirety. For the oriens, analyses included the first
section in which the rostral dentate gyrus formed (Plate 65) to the point where the hippocampus
started to tip ventrally (Plate 70), as previously described (Mosley et al., 2017a). S1 was
analyzed in three sections: the rostral-most section in which the dentate gyrus was clearly
2.1.5 Quantification of Iba1 and AC3 labeling
To quantify Iba1 expressing cells, photomicrographs were taken of sections containing
the regions of interest listed above. Regions were outlined and the number of labeled cells within
each region was quantified using the particle counter function in ImageJ (version 1.51t, National
Institutes of Health). We also calculated average microglial size in each brain region by dividing
the total number of Iba1+ pixels by the total number of Iba1+ cells (i.e., number of pixels per
cell). To quantify AC3 positive cells, StereoInvestigator software (MBF Bioscience, Williston,
VT) was used to outline each region of interest and all labeled cells within the outline were
manually counted. All cell counts are expressed as densities (# of labeled cells /mm3).
2.1.6 Brain Cytokine Expression
A separate cohort of mice was treated with control or clodronate-filled liposomes as
above and brains were collected on P2 for analysis of cytokine expression by Real-Time PCR.
We contrasted expression in the septum and hippocampus because these two areas had a similar
percent reduction in microglia after clodronate liposome treatment, but were divergent with
respect to the effect on cell death (see Results). Brains were stored at -80°C until sectioning on a
cryostat to reach the areas of interest. For the septum, one punch (containing the medial and
lateral nuclei) was collected (inner diameter = 2 mm; Plates 57-59 in the Paxinos et al., 2007
atlas). For the hippocampus, four punches were taken (two per side, inner diameter = 1 mm)
from the region represented in Plates 68-72 of Paxinos et al. (2007). Tissue was placed in an
RNAase-free tube and homogenized in TRIzol (Invitrogen, Carlsbad, CA, USA). RNA was
precipitated, and reverse transcription performed with SuperScript IV First-Strand Synthesis
System (Invitrogen) in a thermal cycler (Applied Biosystems Inc., Foster City, CA, USA). Real
FastStart Essential DNA Green Master Kit (Roche, Basel, Switzerland) according to the
manufacturer’s instructions. Validated primers were used to detect expression of interleukin-1β
(IL-1 β), IL-6, tumor necrosis factor-α (TNF-α), IL-10, transforming growth factor-β (TGF-β)
(Qiagen Inc., Valencia, CA, USA), and two control genes: glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; Qiagen) and 18S rRNA (Integrated DNA Technologies Inc., Skokie,
IL, USA). Cytokines were expressed relative to an average of both control genes. N = 10-13
mice per group for the septum and 7-13 per group for the hippocampus.
2.1 Statistics
Because we found no effect of sex in our pilot experiments, and previous studies from our
lab also found no sex difference in cell death between P0-P2 in the brain regions analyzed here
(Ahern et al., 2013; Mosley et al., 2017a), we combined data from males and females in our
analyses. Two-tailed independent t-tests were used to compare saline- and clodronate-liposome
groups for density of Iba1+ and AC3+ cells. Regional differences in microglial density and cell
size were analyzed using repeated measures analysis of variance (ANOVA), and two-way
3 RESULTS
3.1 Clodronate liposome treatment reduces microglial cell number
We observed regional differences in microglial density among control animals (P <
0.0001), with the lowest density in S1 (2.9 ± 0.08 x 103 cells/mm3) and the highest density in the
[image:19.612.176.414.297.471.2]hippocampal oriens layer (19.5 ± 0.57 x 103 cells/mm3; Table 3.1).
Table 3.1 Microglial density (Iba1+ cells per mm3) and cell death density (AC3+ cells per mm3)
across seven brain regions of saline-treated newborns
Intracerebroventricular administration of clodronate liposomes on P0 and P1 depleted
microglia at P2 in all brain regions examined (Figures 3.1 & 3.2). Compared to controls, Iba1+
cell density was reduced by an average of 44% across regions in clodronate-treated animals: MS
(50 ± 5%, P < 0.0001), LS (54 ± 4%, P < 0.0001), BNSTp (32 ± 7%; P < 0.01), MeA (33 ± 3%,
P < 0.0001), PVN (32 ± 3%, P < 0.001;), oriens (51 ± 6 %, P < 0.001), and S1 (54 ± 6%, P <
0.001).
Region1 Iba1 Density AC3 Density
S1 2.9 ± 0.08 0.08 ± 0.01
BNSTp 6.6 ± 0.93 0.92 ± 0.17
LS 7.4 ± 0.19 0.18 ± 0.02
MS 9.2 ± 0.39 0.38 ± 0.443
MeA 9.8 ± 0.23 0.75 ± 0.10
PVN 13.4 ± 0.68 0.70 ± 0.05
Oriens 19.5 ± 0.57 1.9 ± 0.12
Figure 3.1 Photomicrographs of Iba1-labeled cells in neonates receiving
intracerebroventricular injections of vehicle- (left) or clodronate-filled (right) liposomes.
(A,B) Lateral and medial septum (LS, MS); (C,D) medial amygdala (MeA); (E,F) hippocampus, including hippocampal oriens. Abbreviations: cc, corpus callosum; LV, lateral ventricle. Scale
3.2 Reduction of microglia is associated with region-dependent changes in cell death
Consistent with our previous observations, cell death in vehicle-treated animals was much
higher in the oriens layer of the hippocampus than in any other region examined, and the rates of
cell death in other brain regions was also similar to that reported previously (Ahern et al., 2013;
Mosley et al., 2017a). We observed a significant correlation between AC3+ cell density and
microglial cell density across regions (r = 0.84; P = 0.01; Table 2.1), which is consistent with
past reports that microglia are concentrated in brain regions with high levels of cell death
(Marín-Teva et al., 2004; Wakselman et al., 2008).
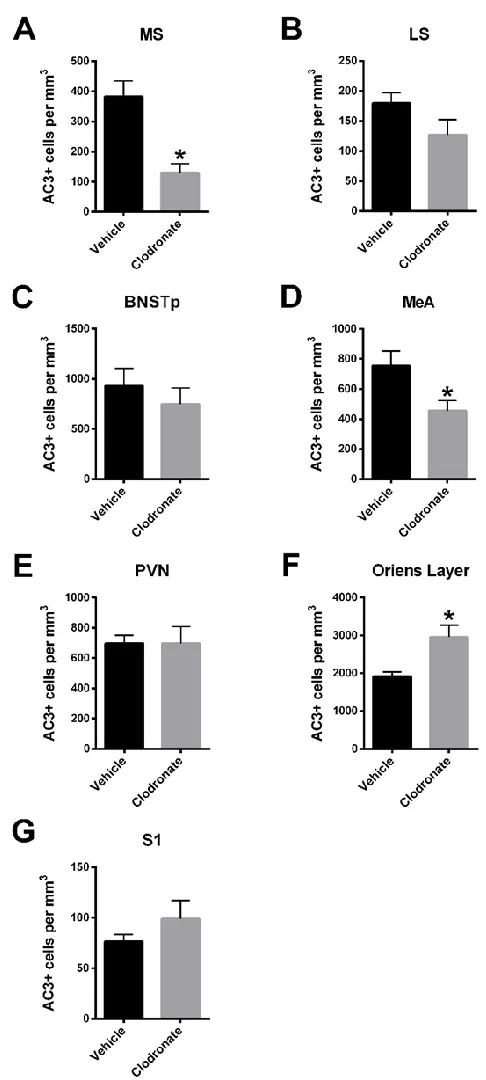
Animals treated with clodronate liposomes had fewer AC3+ cells compared to
vehicle-treated controls in five of the seven regions examined, although this was significant only for the
MS (66 ± 8% reduction in AC3 cells, P < 0.05) and MeA (40 ± 9% reduction, P < 0.05) (Figures
3.3 & 3.4). In striking contrast, cell death was significantly increased in clodronate-treated pups
Figure 3.3 Photomicrographs of AC3-labeled cells (black) in neonatal mice treated with liposomes containing vehicle or clodronate.
Clodronate liposomes decreased AC3+ cell number in the medial septum (A,B) and medial amygdala (C,D) but increased cell death in the oriens layer (E,F). Tissue has been counterstained with thionin (blue). Abbreviations: cc, corpus callosum; LV, lateral ventricle.
Figure 3.4 Neonatal treatment with clodronate liposomes caused region-specific changes in perinatal cell death.
AC3+ cell density was significantly reduced in the medial septum (MS) and medial amygdala (MeA), but was significantly increased in the oriens layer of the hippocampus of
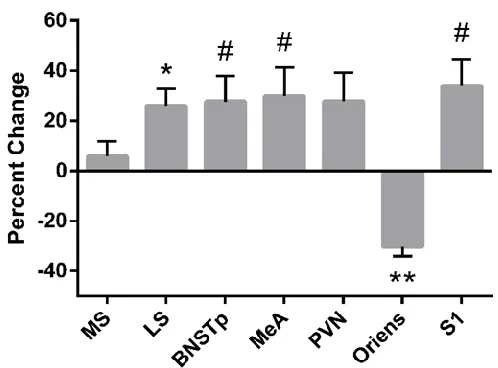
3.3 Effect of clodronate treatment on microglial cell size varies by region
We found no significant difference across brain regions in microglial size in control
animals. However, clodronate liposome treatment affected microglial size, and in a manner that
varied by region (Figure 3.5). In most brain regions, the size of remaining microglia was
increased; this was significant for the LS (26.1%, P < 0.05) and approached significance for the
BNSTp, MeA, and S1 (27.7% to 33.9% increase; P < 0.1 in all cases). In contrast, microglial cell
size was significantly reduced in clodronate-treated pups in the oriens layer of the hippocampus
[image:25.612.198.447.302.486.2](30.5%, P < 0.01).
Figure 3.5 Average microglial cell size was increased in most brain regions of clodronate-treated pups, but was reduced in the hippocampal oriens.
Data are expressed relative to microglial cell size in the corresponding brain region of vehicle-treated controls. # < 0.10; *P < 0.05; **P < 0.01.
3.4 Elevated expression of inflammatory cytokines in the hippocampus
Microglia are the main source of inflammatory cytokines in the brain (Doorn et al., 2015)
and cytokines have been linked to neuronal cell death following hypoxia or injury (Hasturk et al.,
in the septum and oriens layer of the hippocampus, we asked whether baseline cytokine
expression, or cytokine expression following treatment with clodronate liposomes, might differ
between these brain regions. For each of the inflammatory cytokines examined, we found a
main effect of brain region with no significant effect of liposome treatment or
region-by-treatment interaction: the expression of IL-1β (P < 0.01), IL-6 (P < 0.01), and TNF-α (P < 0.05)
were all higher in the hippocampus than in the septum (Figure 3.6). There were no significant
effects of region or treatment for IL-10, which is generally anti-inflammatory, or for the
Figure 3.6 The expression of cytokines in the septum and hippocampus of neonates treated with saline- (black bars) or clodronate-filled (gray bars) liposomes.
Expression of the inflammatory cytokines interleukin IL-1β, IL-6, and tumor necrosis factor alpha (TNF-α) was greater in the hippocampus than in the septum, independent of liposome treatment. Data are presented relative to expression of control genes GAPDH and 18S RNA. *P
4 CONCLUSIONS
Treatment of neonatal mice with clodronate liposomes decreased microglial number in all
brain regions examined. The reduction in microglia seen here is similar to that reported
previously in rats following injections of clodronate liposomes during the first week of life
(VanRyzin et al., 2016; Nelson and Lenz, 2017). These authors also found that although
microglial density returned to normal within a week of treatment, the neonatal depletion of
microglia had lasting effects on locomotor behavior, anxiety, and prosocial behaviors (VanRyzin
et al., 2016; Nelson and Lenz, 2017). The mechanism(s) underlying effects of transient, early-life
microglia depletion on later behavior is not known but could involve changes in developmental
processes such as cell death.
Previous studies reducing microglia, or preventing the interaction between microglia and
neurons, have variously concluded that microglia kill or protect developing neurons (Marín-Teva
et al., 2011). Most of the prior studies have been performed in vitro, however, which could be
problematic because cultured microglia produce higher levels of inflammatory cytokines than are
seen in the neonate, and differ from brain-resident microglia in the expression of other genes
(Crain et al., 2013; Hurley et al., 1999). Two other studies using in vivo models have reached
conflicting conclusions regarding microglial effects on perinatal cell death (Ueno et al., 2013;
Wakselman et al., 2008). In addition to differences in whether the subject tissue is in vivo or in
vitro, differences in effects could be due to species, mode of manipulating microglia, or other
differences in experimental design. Since each study focused on a single brain region, the
possibility of identifying regional differences are diminished.
Here, we addressed this by examining multiple brain regions at a constant developmental
developmental cell death. Cell death was significantly reduced in the medial septum and medial
amygdala however, cell death was increased in the oriens layer of the hippocampus. Interestingly
the magnitude of microglial reduction was not related to the effect on cell death (e.g., a 50%
reduction in microglia in the medial septum decreased cell death, while a 51% reduction of
microglia in the hippocampal oriens increased cell death). Instead, microglia appear to have
opposite effects on neuronal cell death in different regions of the brain.
Although our study demonstrates microglial influence on developmental cell death, it
focuses on one developmental time point (P2), which is a limitation of this study. Wakselman et
al. (2008) observed dynamic effects of microglial influence on cell death over time, with
significant effects occurring on P0 and P1, and what appears to be almost a reverse effect on P2.
This signals that perhaps microglial influence on cell death is not only regionally linked but also
temporally linked. A longitudinal study looking at the effect of microglia depletion on cell death
at earlier and later timepoints would test whether there are age dependent differences in
microglia’s influence on development.
A second limitation is that glia other than microglia may influence developmental cell death.
Astrocytes are involved in developmental events such as promoting angiogenesis in the brain,
axonal guidance, and promoting developmental neuronal survival (Ma et al., 2012; Powell and
Geller, 1999; Schwartz and Nishiyama, 1994). It has also been shown that astrocytes are highly
reactive to microglial signaling (Riddler, 2017), so it is possible that clodronate treatment
indirectly affected astrocyte number or gene expression and that this led to the examined effects
on cell death. Examination of astrocytes after clodronate treatment in future studies could
Microglia also play dual roles in other aspects of neural circuit formation (Konishi &
Kiyama, 2018). For example, they have been reported to either reduce or promote neurogenesis
(Arnò et al., 2014; Cunningham et al., 2013) and to increase or eliminate synapses (Paolicelli et
al., 2011; Parkhurst et al., 2013; Schafer et al., 2012). Whether these are regional differences is
not clear, but microglia isolated from different brain regions display distinct inflammatory
profiles under basal and stimulated conditions (de Haas et al., 2008; Kim et al., 2000; Kostuk et
al., 2018; Lai et al., 2011; Olah et al., 2011), which could account for differences in the effect of
inhibiting or eliminating microglia.
Given that we found a two-fold higher density of microglia in the hippocampus than in the
septum and also higher expression in the hippocampus than in the septum of all three
inflammatory cytokines examined (IL-1 β, IL-6, and TNF-α) this corroborates previous evidence
that microglia exhibit regional differences. The high inflammatory cytokine expression could be
due to the greater density of microglia, and could contribute to the very high rate of cell death in
the hippocampal oriens: IL-1β exacerbates neuronal cell death due to ischemia or excitotoxicity
(Loddick & Rothwell, 1996; Relton & Rothwell, 1992), and TNF-α induces apoptosis of neurons
and neural precursor cells (Guadagno et al., 2013; Kaur et al., 2014; Sedel et al., 2004).
However, a comparison between our cytokine data and regional cell death data is not perfect.
Tissue punches included all hippocampal cell layers, whereas the microglial and AC3 counts in
the present study were specific to the oriens. We also cannot rule out the possibility that the
elevated cytokine expression in the hippocampus is a consequence, rather than cause, of the high
rate of cell death.
Another interesting result was the effect of treatment on cytokine expression in these two
significant differences were seen in cytokine expression. This result implies a compensation
effect by either the remaining microglia or other glia (Sofroniew, 2014). This compensation is
consistent with a previous report in microglia-depleted adult mice (Parkhurst et al., 2013). Given
that an effect was seen on developmental cell death as a result of microglia depletion and
cytokine expression remained unchanged, this leads us to conclude that the cytokines measured
here may not be responsible for inducing developmental cell death but may contribute to overall
regional heterogeneity.
Furthermore, the cytokine expression levels we observed may not represent a clear picture
of what specifically microglia in the tissue are expressing. The punches that we collected
included all cell types that exist in brain tissue. Although microglia are the primary producers of
brain cytokines, it is possible that upon microglial number reduction, other glia such as
astrocytes upregulate cytokine expression. Furthermore, it is possible that the remaining
microglia do upregulate expression and thus differences in cytokine expression would not be
observed, however protein turnover may not keep pace with the higher expression levels. Future
studies looking at protein levels instead of mRNA expression would provide a clearer picture of
how cytokines may influence cell death.
Interestingly, an effect of increased microglial cell size was seen across the brain in the
clodronate-treated group. As far as we know, this effect of clodronate treatment has not been
described previously. Microglia in adults normally “tile” the brain, covering all areas without
overlap of territory (Jinnoet al., 2007). It’s possible that a reduction in microglial number may
trigger an increase in size of the remaining microglia, and in some way may reflect the
compensation in cytokine expression. It is worth noting, that microglia impacts on
microglial cell size. It is unclear whether one precedes the other, but it furthers the theme of
regional heterogeneity. One limitation of this finding however, is that we analyzed microglial
size by dividing pixels above threshold by microglial counts. A more in-depth analysis looking at
phenotypic measures such as branch length and complexity, number of phagocytic cups, and
soma size could provide a nuanced look at microglial activation than our gross measure of size.
In summary, these findings add to a growing body of work suggesting regional differences in
REFERENCES
Ahern, T. H., Krug, S., Carr, A. V., Murray, E. K., Fitzpatrick, E., Bengston, L., McCutcheon, J.,
De Vries, G.J., Forger, N. G. (2013). Cell death atlas of the postnatal mouse ventral
forebrain and hypothalamus: Effects of age and sex. Journal of Comparative Neurology,
521(11), 2551–2569. https://doi.org/10.1002/cne.23298
Arnò, B., Grassivaro, F., Rossi, C., Bergamaschi, A., Castiglioni, V., Furlan, R., Greter, M.,
Favaro, R., Comi, G., Becher, B., Martino, G., Muzio, L. (2014). Neural progenitor cells
orchestrate microglia migration and positioning into the developing cortex. Nature
Communications, 5(1), 5611. https://doi.org/10.1038/ncomms6611
Ashwell, K. (1990). Microglia and cell death in the developing mouse cerebellum. Brain
Research. Developmental Brain Research, 55(2), 219–230.
https://doi.org/10.1016/0165-3806(90)90203-B
Brown, G. C., & Neher, J. J. (2014). Microglial phagocytosis of live neurons. Nature Reviews
Neuroscience, 15(4), 209–216. https://doi.org/10.1038/nrn3710
Crain, J. M., Nikodemova, M., & Watters, J. J. (2013). Microglia express distinct M1 and M2
phenotypic markers in the postnatal and adult central nervous system in male and female
mice. Journal of Neuroscience Research, 91(9), 1143–1151.
https://doi.org/10.1002/jnr.23242
Cunningham, C. L., Martinez-Cerdeno, V., & Noctor, S. C. (2013). Microglia regulate the
number of neural precursor cells in the developing cerebral cortex. Journal of Neuroscience,
33(10), 4216–4233. https://doi.org/10.1523/JNEUROSCI.3441-12.2013
mediates rapid microglial response to local brain injury in vivo. Nature Neuroscience, 8(6),
752–758. https://doi.org/10.1038/nn1472
de Haas, A. H., Boddeke, H. W. G. M., & Biber, K. (2008). Region-specific expression of
immunoregulatory proteins on microglia in the healthy CNS. Glia, 56(8), 888–894.
https://doi.org/10.1002/glia.20663
Doorn, K. J., Brevé, J. J. P., Drukarch, B., Boddeke, H. W., Huitinga, I., Lucassen, P. J., & van
Dam, A.-M. (2015). Brain region-specific gene expression profiles in freshly isolated rat
microglia. Frontiers in Cellular Neuroscience, 9, 84.
https://doi.org/10.3389/fncel.2015.00084
Faustino, J. V., Wang, X., Johnson, C. E., Klibanov, A., Derugin, N., Wendland, M. F., &
Vexler, Z. S. (2011). Microglial cells contribute to endogenous brain defenses after acute
neonatal focal stroke. The Journal of Neuroscience, 31(36), 12992–13001.
https://doi.org/10.1523/JNEUROSCI.2102-11.2011
Frade, J. M., & Barde, Y.-A. (1998). Microglia-derived nerve growth factor causes cell death in
the developing retina. Neuron, 20(1), 35–41.
https://doi.org/10.1016/S0896-6273(00)80432-8
Giulian D, Young DG, Woodward J, Brown DC, Lachman LB. 1988. Interleukin-1 is an
astroglial growth factor in the developing brain. J Neurosci [Internet] 8:709–14. Available
from: http://www.ncbi.nlm.nih.gov/pubmed/3257519
Guadagno, J., Xu, X., Karajgikar, M., Brown, A., & Cregan, S. P. (2013). Microglia-derived
TNFα induces apoptosis in neural precursor cells via transcriptional activation of the Bcl-2
https://doi.org/10.1038/cddis.2013.59
Hasturk, A. E., Gokce, E. C., Yilmaz, E. R., Horasanli, B., Evirgen, O., Hayirli, N., Gokturk, H.,
Erguder, I., Can, B. (2018). Therapeutic evaluation of tumor necrosis factor-alpha
antagonist Etanercept against traumatic brain injury in rats: ultrastructural, pathological, and
biochemical analyses. Asian Journal of Neurosurgery, 13(4), 1018–1025.
https://doi.org/10.4103/ajns.AJNS_29_17
Hristovska, I., & Pascual, O. (2015). Deciphering Resting Microglial Morphology and Process
Motility from a Synaptic Prospect. Frontiers in Integrative Neuroscience, 9, 73.
https://doi.org/10.3389/fnint.2015.00073
Hurley, S. D., Walter, S. A., Semple-Rowland, S. L., & Streit, W. J. (1999). Cytokine transcripts
expressed by microglia in vitro are not expressed by ameboid microglia of the developing
rat central nervous system. Glia, 25(3), 304–309.
https://doi.org/10.1002/(SICI)1098-1136(19990201)25:3<304::AID-GLIA10>3.0.CO;2-W
Jinno, S., Fleischer, F., Eckel, S., Schmidt, V., & Kosaka, T. (2007). Spatial arrangement of
microglia in the mouse hippocampus: A stereological study in comparison with astrocytes.
Glia, 55(13), 1334–1347. https://doi.org/10.1002/glia.20552
Kaur, C., Sivakumar, V., Zou, Z., & Ling, E.-A. (2014). Microglia-derived proinflammatory
cytokines tumor necrosis factor-alpha and interleukin-1beta induce Purkinje neuronal
apoptosis via their receptors in hypoxic neonatal rat brain. Brain Structure and Function,
219(1), 151–170. https://doi.org/10.1007/s00429-012-0491-5
Kim, W. G., Mohney, R. P., Wilson, B., Jeohn, G. H., Liu, B., & Hong, J. S. (2000). Regional
of microglia. The Journal of Neuroscience, 20(16), 6309–6316.
https://doi.org/10.1523/JNEUROSCI.20-16-06309.2000
Konishi, H., & Kiyama, H. (2018). Microglial TREM2/DAP12 signaling: a double-edged sword
in neural diseases. Frontiers in Cellular Neuroscience, 12, 206.
https://doi.org/10.3389/fncel.2018.00206
Kostuk, E. W., Cai, J., & Iacovitti, L. (2018). Regional microglia are transcriptionally distinct
but similarly exacerbate neurodegeneration in a culture model of Parkinson’s disease.
Journal of Neuroinflammation, 15(1), 139. https://doi.org/10.1186/s12974-018-1181-x
Kumamaru, H., Saiwai, H., Kobayakawa, K., Kubota, K., van Rooijen, N., Inoue, K., Iwamoto,
Y., Okada, S. (2012). Liposomal clodronate selectively eliminates microglia from primary
astrocyte cultures. Journal of Neuroinflammation, 9(1), 647.
https://doi.org/10.1186/1742-2094-9-116
Lai, A. Y., Dhami, K. S., Dibal, C. D., & Todd, K. G. (2011). Neonatal rat microglia derived
from different brain regions have distinct activation responses. Neuron Glia Biology, 7(01),
5–16. https://doi.org/10.1017/S1740925X12000154
Loddick, S. A., & Rothwell, N. J. (1996). Neuroprotective effects of human recombinant
interleukin-1 receptor antagonist in focal cerebral ischaemia in the rat. Journal of Cerebral
Blood Flow & Metabolism, 16(5), 932–940.
https://doi.org/10.1097/00004647-199609000-00017
Ma, S., Kwon, H. J., & Huang, Z. (2012). A Functional Requirement for Astroglia in Promoting
Blood Vessel Development in the Early Postnatal Brain. PLoS ONE, 7(10).
Marín-Teva, J. L., Dusart, I., Colin, C., Gervais, A., van Rooijen, N., & Mallat, M. (2004).
Microglia promote the death of developing Purkinje cells. Neuron, 41(4), 535–547.
https://doi.org/10.1016/S0896-6273(04)00069-8
Marín-Teva JL, Cuadros MA, Martín-Oliva D, Navascués J. (2011). Microglia and neuronal cell
death. Neuron Glia Biology, 7(1), 25-40. https://doi.org/10.1017/S1740925X12000014
Mosley, M., Shah, C., Morse, K. A., Miloro, S. A., Holmes, M. M., Ahern, T. H., & Forger, N.
G. (2017a). Patterns of cell death in the perinatal mouse forebrain. Journal of Comparative
Neurology, 525(1), 47–64. https://doi.org/10.1002/cne.24041
Mosley, M., Weathington, J., Cortes, L. R., Bruggeman, E., Castillo-Ruiz, A., Xue, B., & Forger,
N. G. (2017b). Neonatal Inhibition of DNA Methylation Alters Cell Phenotype in Sexually
Dimorphic Regions of the Mouse Brain. Endocrinology, 158(6), 1838–1848.
https://doi.org/10.1210/en.2017-00205
Nelson, L. H., & Lenz, K. M. (2017). Microglia depletion in early life programs persistent
changes in social, mood-related, and locomotor behavior in male and female rats.
Behavioural Brain Research, 316, 279–293. https://doi.org/10.1016/j.bbr.2016.09.006
Olah, M., Biber, K., Vinet, J., & Boddeke, H. W. G. M. (2011). Microglia phenotype diversity.
CNS & Neurological Disorders Drug Targets, 10(1), 108–118.
https://doi.org/10.2174/187152711794488575
Oppenheim, R. W. (1985). Naturally occurring cell death during neural development. Trends in
Neurosciences, 8, 487–493. https://doi.org/10.1016/0166-2236(85)90175-4
Ferreira, T.A., Guiducci, E., Dumas, L., Ragozzino, D., Gross, C. T. (2011). Synaptic
Pruning by Microglia Is Necessary for Normal Brain Development. Science, 333(6048),
1456–1458. https://doi.org/10.1126/science.1202529
Parkhurst, C. N., Yang, G., Ninan, I., Savas, J. N., Yates, J. R., Lafaille, J. J., Hempstead, B.L.,
Littman, D.R., Gan, W.-B. (2013). Microglia promote learning-dependent synapse
formation through brain-derived neurotrophic factor. Cell, 155(7), 1596–1609.
https://doi.org/10.1016/j.cell.2013.11.030
Paxinos G, Halliday G, Watson C, Koutcherov Y, Wang HQ. Atlas of the developing mouse
brain at E17.5, P0 and P6. San Diego: Elsevier; 2007.
Perry, V. H., Hume, D. A., & Gordon, S. (1985). Immunohistochemical localization of
macrophages and microglia in the adult and developing mouse brain. Neuroscience, 15(2),
313–326. https://doi.org/10.1016/0306-4522(85)90215-5
Powell, E. M., & Geller, H. M. (1999). Dissection of astrocyte-mediated cues in neuronal
guidance and process extension. Glia, 26(1), 73-83.
doi:10.1002/(sici)1098-1136(199903)26:13.0.co;2-s
Relton, J. K., & Rothwell, N. J. (1992). Interleukin-1 receptor antagonist inhibits ischaemic and
excitotoxic neuronal damage in the rat. Brain Research Bulletin, 29(2), 243–246.
https://doi.org/10.1016/0361-9230(92)90033-T
Ridler, C. (2017). Microglia-induced reactive astrocytes — toxic players in neurological
disease? Nature Reviews Neurology, 13(3), 127-127. doi:10.1038/nrneurol.2017.17
Ransohoff, R.M., Greenberg, M.E., Barres, B.A., Stevens, B. (2012). Microglia sculpt
postnatal neural circuits in an activity and complement-dependent manner. Neuron, 74(4),
691–705. https://doi.org/10.1016/j.neuron.2012.03.026
Schwartz, J. P., & Nishiyama, N. (1994). Neurotrophic factor gene expression in astrocytes
during development and following injury. Brain Research Bulletin, 35(5-6), 403-407.
doi:10.1016/0361-9230(94)90151-1
Schwarz JM, Sholar PW, Bilbo SD. 2012. Sex differences in microglial colonization of the
developing rat brain. J Neurochem [Internet] 120:no-no. Available from:
http://www.ncbi.nlm.nih.gov/pubmed/22182318
Sedel, F., Béchade, C., Vyas, S., & Triller, A. (2004). Macrophage-derived tumor necrosis
factor, an early developmental signal for motoneuron death. Journal of Neuroscience, 24(9),
2236–2246. https://doi.org/10.1523/JNEUROSCI.4464-03.2004
Smith, J. A., Das, A., Ray, S. K., & Banik, N. L. (2012). Role of pro-inflammatory cytokines
released from microglia in neurodegenerative diseases. Brain Research Bulletin, 87(1), 10–
20. https://doi.org/10.1016/j.brainresbull.2011.10.004
Sofroniew M V. 2014. Multiple Roles for Astrocytes as Effectors of Cytokines and
Inflammatory Mediators. Neurosci [Internet] 20:160–172. Available from:
http://journals.sagepub.com/doi/10.1177/1073858413504466
Srinivasan A, Roth KA, Sayers RO, Shindler KS, Wong AM, Fritz LC, Tomaselli KJ. (1998). In
situ immunodetection of activated caspase-3 in apoptotic neurons in the developing nervous
system. Cell Death and Differentiation, 5(12), 1004–1016.
Strahan, J. A., Walker, W. H., Montgomery, T. R., & Forger, N. G. (2017). Minocycline causes
widespread cell death and increases microglial labeling in the neonatal mouse brain.
Developmental Neurobiology, 77(6), 753–766. https://doi.org/10.1002/dneu.22457
Tucker EW, Pokkali S, Zhang Z, DeMarco VP, Klunk M, Smith ES, Ordonez AA, Penet M-F,
Bhujwalla Z, Jain SK, Kannan S. 2016. Microglia activation in a pediatric rabbit model of
tuberculous meningitis. Dis Model Mech [Internet] 9:1497–1506. Available from:
http://www.ncbi.nlm.nih.gov/pubmed/27935825
Ueno, M., Fujita, Y., Tanaka, T., Nakamura, Y., Kikuta, J., Ishii, M., & Yamashita, T. (2013).
Layer V cortical neurons require microglial support for survival during postnatal
development. Nature Neuroscience, 16(5), 543–551. https://doi.org/10.1038/nn.3358
VanRyzin, J. W., Yu, S. J., Perez-Pouchoulen, M., & McCarthy, M. M. (2016). Temporary
depletion of microglia during the early postnatal period induces lasting sex-dependent and
sex-independent effects on behavior in rats. ENeuro, 3(6), ENEURO.0297-16.2016.
https://doi.org/10.1523/ENEURO.0297-16.2016
Wakselman, S., Bechade, C., Roumier, A., Bernard, D., Triller, A., & Bessis, A. (2008).
Developmental neuronal death in hippocampus requires the microglial CD11b integrin and
DAP12 immunoreceptor. Journal of Neuroscience, 28(32), 8138–8143.
https://doi.org/10.1523/JNEUROSCI.1006-08.2008
Zacharaki T, Sophou S, Giannakopoulou A, Dinopoulos A, Antonopoulos J, Parnavelas JG, Dori
I. (2010). Natural and lesion-induced apoptosis in the dorsal lateral geniculate nucleus
during development. Brain Research, 1344, 62-76.