1535-9778/07/$08.00⫹0 doi:10.1128/EC.00213-06
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Divergence of Protein Kinase A Catalytic Subunits in
Cryptococcus neoformans
and
Cryptococcus gattii
Illustrates
Evolutionary Reconfiguration of a Signaling Cascade
䌤
†
Julie K. Hicks
1and Joseph Heitman
1,2,3*
Department of Molecular Genetics and Microbiology,1Medicine,2and Pharmacology and Cancer Biology,3
Duke University Medical Center, Durham, North Carolina 27710
Received 6 July 2006/Accepted 11 December 2006
Gene duplication and divergence via both the loss and gain of gene activities are powerful evolutionary forces underlying the origin of new biological functions. Here a comparative genetics approach was applied to examine the roles of protein kinase A (PKA) catalytic subunits in three closely related varieties or sibling species of the pathogenic fungus genusCryptococcus. Previous studies revealed that two PKA catalytic subunits, Pka1 and Pka2, control virulence factor production and mating. However, only one of the two plays the predominant physiological role, and this function has been exchanged between Pka1 and Pka2 in strains of the
Cryptococcus neoformansvar. grubiiserotype A lineage compared to divergent C. neoformansvar.neoformans
serotype D isolates. To understand the basis for this functional plasticity, here the activities of Pka1 and Pka2 were defined in the two varieties and the related sibling speciesCryptococcus gattii by gene disruption and characterization, heterologous complementation, and analysis of serotype AD hybrid mutant strains. The findings provide evidence for a shared ancestral role of PKA in governing mating and virulence factor production and indicate that the exchange of catalytic subunit roles is attributable to loss of function. Our studies illustrate the plasticity of signaling networks enabling rapid rewiring during speciation of a clade of common human fungal pathogens.
Gene duplication and divergence are significant forces in the evolution of new genes (21). Gene duplication events can oc-cur both on a macrogenomic scale, such as that involving the ancestral whole genome duplication that gave rise to
Saccha-romyces cerevisiaeand related sensu stricto strains (31), and on
the microgenomic scale of individual genes. Several models regarding the mechanisms by which new functions arise fol-lowing gene duplication have been posited. The model origi-nally postulated by Ohno (21), now termed “neofunctionaliza-tion,” suggests that following duplication, one gene of the pair retains the original function while functional constraints on the other gene are relaxed, allowing it to undergo accelerated evolution and eventually develop novel functions.
A well-documented example of neofunctionalization is that of theORC1/SIR3gene pair that arose inS. cerevisiaefollowing the whole-genome duplication that occurred approximately 100 to 300 million years ago (mya) (15, 28). In this instance,S.
cerevisiaeOrc1 shares 48% identity with a protein found in the
related species,Saccharomyces kluyveri, while Sir3 shares only 24% identity with this protein, suggesting that Orc1 has re-tained the ancestral function while Sir3 has been subject to accelerated evolution. Consistent with this is the observation that theS. kluyveri ORC1/SIR3 gene can complement theS.
cerevisiae orc1⌬mutant but not theS. cerevisiae sir3⌬mutant.
Another example of gene duplication inS. cerevisiaeinvolves the genes encoding the three protein kinase A (PKA) catalytic subunits, Tpk1, Tpk2, and Tpk3. All three catalytic subunits share a redundant function yet also have novel functions as well. While all three catalytic subunits share redundant roles in viability, as demonstrated by the fact that atpk1⌬tpk2⌬tpk3⌬
triple mutant is inviable, they play opposing roles in pseudohy-phal growth, with Tpk2 activating and Tpk1 and Tpk3 repress-ing the filamentous dimorphic transition (22, 24). The abun-dance of well-studied gene duplications inS. cerevisiaeand the presence of a well-characterized lineage (including the closely related sensu stricto strains that can be used as outgroups)
makeS. cerevisiaean ideal model system for examining gene
duplication events in ascomycete fungi.
Similarly, Cryptococcus neoformans has several attributes that render it a facile system in which to study gene duplication events in basidiomycete fungi. First, there are multiple exam-ples of gene duplications in this fungus, including the laccase genesLAC1and LAC2, the Ras genes RAS1andRAS2, the carbonic anhydrase genesCAN1andCAN2, and the cyclophi-lin A genesCPA1andCPA2(2, 4, 23, 29, 30). Another example of gene duplication inCryptococcusinvolves the protein kinase A catalytic subunits, Pka1 and Pka2, which share 35% identity at the protein level. Our previous work showed that both the serotype A and D lineages have both subunits but that the functions of these subunits differ between the two lineages. Deletion of thePKA1gene in serotype A results in a loss of mating and melanin and capsule production, whereas it is the deletion of thePKA2gene in serotype D that results in the loss of these functions (3, 11). The second attribute that renders
Cryptococcus neoformansan amenable model system for
study-ing gene duplication and evolution in basidiomycete fungi is * Corresponding author. Mailing address: Department of Molecular
Genetics and Microbiology, 322 CARL Bldg., Duke University Med-ical Center, Research Dr., Durham, NC 27710. Phone: (919) 684-2824. Fax: (919) 684-5458. E-mail: [email protected].
† Supplemental material for this article may be found at http://ec .asm.org/.
䌤Published ahead of print on 22 December 2006.
413
on September 8, 2020 by guest
http://ec.asm.org/
the presence of full genome sequences for the closely related varietiesC. neoformansvar. neoformans(serotype D) andC.
neoformansvar.grubii(serotype A) and sister species
Crypto-coccus gattii(18). Much like the sensu stricto strains of
Sac-charomyces, theCryptococcusspecies complex enables detailed
analyses of gene evolution and function (13).
In this study, we conducted a comparative genetics study of the Pka1 and Pka2 catalytic subunits inC. gattii. Based on our results, we propose a model in which Pka1 and Pka2 played a shared ancestral role in mating and melanin and capsule pro-duction. InC. gattii, Pka1 has lost its role in melanin produc-tion while retaining roles in capsule producproduc-tion and mating, and Pka2 functions in all three pathways. InC. neoformansvar.
grubii(serotype A), Pka1 has retained its roles in melanin and
capsule production and mating, whereas Pka2 has lost all three functional roles. Finally, inC. neoformansvar.neoformans (se-rotype D), Pka1 has lost all of these functions, whereas Pka2 retained all three functions. These findings reveal rapid and plastic rewiring of signaling cascades controlling virulence and development during fungal speciation.
MATERIALS AND METHODS
C. neoformansandC. gattiistrains and media.All strains used in this study are listed in Table 1.C. neoformans(serotypes A and D) andC. gattii(serotypes B and C) strains were grown on standardS. cerevisiaemedia (26). The selective medium for biolistic transformation, proline medium for serotype AD hybrid selection, Niger seed medium, V8 medium, and Dulbecco’s modified Eagle’s (DME) medium were prepared as described previously (1, 8, 12, 14, 27).
pka1B⌬::NATandpka1D⌬::NATtransformants were selected on yeast extract-peptone-dextrose (YPD) medium containing 100 g/ml nourseothricin and pka2B⌬::NAT,pka2B⌬::NEO, andpka2D⌬::NEOwere selected on YPD me-dium containing 200g/ml G418. Genotypes were confirmed both by Southern hybridization and expression analysis.
Identification of theC. gattii PKA1andPKA2genes.TheC. gattiiserotype B PKA1andPKA2genes were identified by performing a BLASTn search of the Broad Institute serotype B database (strain R265) by using theC. neoformans serotype APKA1andPKA2open reading frame nucleotide sequences.
Disruption of theC. gattiiandC. neoformansserotype DPKA1andPKA2
genes.A prototrophic serotype B wild-type strain (R265) was biolistically trans-formed with the gel-extractedPKA1BandPKA2Bdisruption alleles as described previously (5) to create strains JKH290 and JKH293, respectively. Details on the construction of the overlap constructs can be found in the supplemental material. An auxotrophic serotype D strain containing aura5mutation (JEC34) was transformed with thePKA1D disruption allele to create strain JKH313. To TABLE 1. Strains used in this study
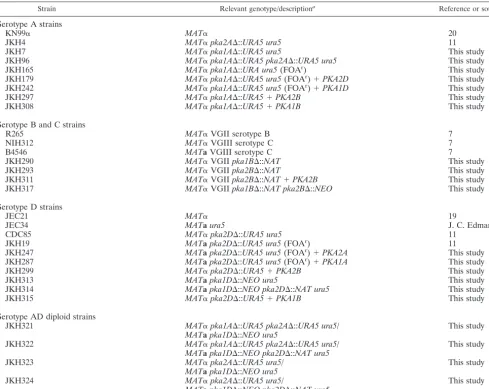
Strain Relevant genotype/descriptiona Reference or source
Serotype A strains
KN99␣ MAT␣ 20
JKH4 MAT␣pka2A⌬::URA5 ura5 11
JKH7 MAT␣pka1A⌬::URA5 ura5 This study
JKH96 MAT␣pka1A⌬::URA5 pka2A⌬::URA5 ura5 This study
JKH165 MAT␣pka1A⌬::URA ura5(FOAr) This study
JKH179 MAT␣pka1A⌬::URA5ura5(FOAr)⫹PKA2D This study
JKH242 MAT␣pka1A⌬::URA5 ura5(FOAr)⫹PKA1D This study
JKH297 MAT␣pka1A⌬::URA5⫹PKA2B This study
JKH308 MAT␣pka1A⌬::URA5⫹PKA1B This study
Serotype B and C strains
R265 MAT␣VGII serotype B 7
NIH312 MAT␣VGIII serotype C 7
B4546 MATaVGIII serotype C 7
JKH290 MAT␣VGIIpka1B⌬::NAT This study
JKH293 MAT␣VGIIpka2B⌬::NAT This study
JKH311 MAT␣VGIIpka2B⌬::NAT⫹PKA2B This study
JKH317 MAT␣VGIIpka1B⌬::NAT pka2B⌬::NEO This study
Serotype D strains
JEC21 MAT␣ 19
JEC34 MATaura5 J. C. Edman
CDC85 MAT␣pka2D⌬::URA5 ura5 11
JKH19 MATapka2D⌬::URA5 ura5(FOAr) 11
JKH247 MATapka2D⌬::URA5 ura5(FOAr)⫹PKA2A This study
JKH287 MATapka2D⌬::URA5 ura5(FOAr)⫹PKA1A This study
JKH299 MAT␣pka2D⌬::URA5⫹PKA2B This study
JKH313 MATapka1D⌬::NEO ura5 This study
JKH314 MATapka1D⌬::NEO pka2D⌬::NAT ura5 This study
JKH315 MAT␣pka2D⌬::URA5⫹PKA1B This study
Serotype AD diploid strains
JKH321 MAT␣pka1A⌬::URA5 pka2A⌬::URA5 ura5/ This study
MATapka1D⌬::NEO ura5
JKH322 MAT␣pka1A⌬::URA5 pka2A⌬::URA5 ura5/ This study
MATapka1D⌬::NEO pka2D⌬::NAT ura5
JKH323 MAT␣pka2A⌬::URA5 ura5/ This study
MATapka1D⌬::NEO ura5
JKH324 MAT␣pka2A⌬::URA5 ura5/ This study
MATapka1D⌬::NEO pka2D⌬::NAT ura5 a
FOAr
, 5-fluoroorotic acid resistant.
on September 8, 2020 by guest
http://ec.asm.org/
obtainpka1⌬pka2⌬double mutants of serotypes B and D, thepka1B⌬::NAT strain (JKH290) and thepka1D⌬::NEOstrain (JKH313) were transformed with pka2B⌬::NEOandpka2D⌬::NATdisruption alleles to create strains JKH317 and JKH314, respectively. Thepka1⌬andpka2⌬strains were screened by diagnostic PCR for the 5⬘junction and confirmed by Southern blot analysis using specific probes generated by PCR (data not shown).
Complementation experiments.Complementation of thepka2D⌬mutant with the serotype A PKA genes was tested by transforming strain JKH19 (serotype D pka2D⌬) (11) with plasmid pJH80 (containing the wild-typePKA1A gene) and pJH167 (containing the wild-typePKA2Agene) by using a biolistic apparatus (27) to create strains JKH287 and JKH247, respectively. Construction details of pJH80 and pJH167, as well as the other plasmids in the complementation analysis, are provided in the supplemental material.
Complementation of thepka1A⌬mutant with the serotype D PKA genes was tested by transforming strain JKH165 with pJH26 (PKA2D) and pJH166 (PKA1D) to create strains JKH176 and JKH242, respectively.
Complementation of thepka1A⌬andpka2D⌬mutants with the serotype B PKA1BandPKA2Bgenes was tested by transforming strain JKH7 (serotype A pka1A⌬) with pJH235 (PKA1B) and pJH240 (PKA2B) to create strains JKH308 and JKH297, respectively, and strain CDC85 (serotype Dpka2D⌬) was trans-formed with pJH235 and pJH240 to create JKH315 and JKH299, respectively. The serotype Bpka2B⌬mutant (JKH293) was also transformed with pJH240 to create strain JKH311. RNA analysis was performed on all transformants to confirm the expression of the heterologous gene.
Serotype A and D hybrid assays.Hybrid assays were accomplished by mixing 1⫻107
cells/ml of each of the mating partners and then plating 20l of the mixed cell suspension onto V8 medium (pH 7.0). After 2 days of growth in the dark, the cells were scraped off the V8 medium (pH 7.0) and replated on selective proline medium containing either G418 (JKH96⫻JKH313 and JKH4⫻JKH313) or G418 and nourseothricin (JKH96⫻JKH314 and JKH4⫻JKH314). The plates were then incubated at 37°C for 4 days. Fusion products were then examined for melanin and capsule production. The test crosses were as follows: JKH324, MAT␣PKA1A pka2A⌬::URA5(JKH4)⫻MATapka1D⌬::NEO pka2D⌬::NAT ura5(JKH314); JKH321,MAT␣ pka1A⌬::URA5 pka2A⌬::URA5 (JKH96)⫻ MATa pka1D⌬::NEO PKA2D ura5 (JKH313); JKH323, MAT␣ PKA1A pka2A⌬::URA5 (JKH4) ⫻ MATa pka1D⌬::NEO PKA2D ura5 (JKH313); JKH322,MAT␣ pka1A⌬::URA5 pka2A⌬::URA5(JKH96)⫻MATa pka1D⌬:: NEO pka2D⌬::NAT ura5(JKH314). PCRs with serotype AMATa- andMAT␣ -specific primers (7270/7271 and 7264/7266, respectively) and serotype DMATa -andMAT␣-specific primers (7273/7274 and 7267/7269, respectively) were utilized to confirm the hybrid diploid status of the fusion products.
Mating assays.␣andastrains were cocultured on V8 medium (pH 7) and incubated at room temperature for 4 days in the dark prior to photography (⫻100 magnification). These strains included wild-type VGII ␣ ⫻ wild-type VGIIIa(R265⫻B4546);pka1B⌬VGII␣ ⫻VGIIIa(JKH290⫻B4546); pka2B⌬VGII␣ ⫻VGIIIa(JKH293⫻B4546); andpka1B⌬pka2B⌬VGII␣ ⫻ VGIIIa(JKH317⫻B4546).
Expression analysis.Fungal strains were inoculated into 5 ml YPD medium and grown overnight at 30°C. Fifty-milliliter amounts of YPD medium in 125-ml flasks were inoculated with 500l of the overnight cultures and grown at 250 rpm and 30°C for 5 h prior to harvesting. RNA was isolated from the harvested cultures with TRIzol (Gibco-BRL) following the manufacturer’s instructions. Fifteen micrograms (based on spectrophotometric measurement) of RNA was separated on a denaturing gel and transferred to nylon membrane. The resulting blots were probed withPKA1B,PKA1D,PKA2B, orPKA2Dgene-specific probes and with actin as a loading control.
Microscopy.All images of mating and confrontation assays were captured with a Nikon Eclipse E400 microscope equipped with a Nikon DXM1200F camera. Images of melanized colonies were captured with a Nikon CoolPix digital cam-era. Differential interference microscopy images were taken with the⫻1,000 objective of a Zeiss Axioskop 2 Plus fluorescence microscope equipped with an AxioCam MRM digital camera.
RESULTS
Identification of theC. gattiiserotype B Pka1 and Pka2.We have previously shown that, despite sharing only 35% identity at the amino acid level, the serotype A Pka1 and serotype D Pka2 catalytic subunits share similar roles in regulating mela-nin and capsule production and mating in their respective serotypes (11). To better understand the functional divergence
of Pka1 and Pka2, we identified the Pka1 and Pka2 catalytic subunits from the genomic sequence of the R265 strain (mo-lecular group VGII) (6) ofC. gattii, a species closely related to
C. neoformans(32) and the cause of an outbreak on Vancouver
Island (6, 16). At the amino acid level, the Pka1 protein from
C. gattii(Pka1B) shares 86% identity with the serotype A Pka1
subunit (Pka1A) and 84% identity with the serotype D Pka1 subunit (Pka1D); Pka1A and Pka1D are 93% identical. TheC.
gattiiPka2 subunit (Pka2B) shares 87% identity with the
type A Pka2 subunit (Pka2A) and 88% identity with the sero-type D Pka2 subunit (Pka2D), similar to the 93% identity shared between Pka2A and Pka2D. In contrast, the Pka1B subunit shares only 34% identity with the Pka2B subunit. This is similar to the identity comparison of Pka1A and Pka2A (35%) and that of Pka1D and Pka2D (35%).
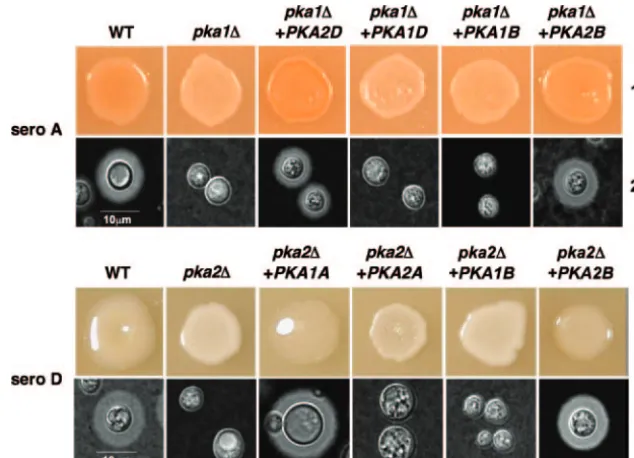
Complementation of pka1A⌬ andpka2D⌬mutations with thePKA1andPKA2genes.We were interested in determining whether the wild-typePKA1A,PKA2B, orPKA2Dgene could complement thepka2D⌬andpka1A⌬mutations in serotypes D and A, respectively. To test this, wild-typePKA1A,PKA2A,
PKA1B,PKA2B,PKA1D, andPKA2Dgenes were ligated into
transformation vectors (plasmids pJH80, pJH167, pJH235, pJH240, pJH166, and pJH26, respectively) and biolistically transformed into pka2D⌬ and pka1A⌬ mutant strains. The resulting transformants were examined for melanin and cap-sule production. The wild-typePKA1Agene, but notPKA2A, complemented the melanin and capsule defects of apka2D⌬
mutant strain (Fig. 1, lanes 3 and 4), while the wild-type
PKA2D gene, but not the PKA1D gene, complemented the
melanin and capsule defects of apka1A⌬mutant strain (Fig. 1, lanes 1 and 2). The PKA2B gene, but not thePKA1B gene, complemented the capsule and melanin defect in bothpka1A⌬
and pka2D⌬mutant strains (Fig. 1). In control experiments,
when thepka1A⌬,pka2D⌬, andpka2B⌬mutant strains were transformed with the plasmids containing the wild-typePKA1A
(pJH80),PKA2D (pJH26), and PKA2B (pJH240) genes, re-spectively, the melanin and capsule defects of the resulting transformants were complemented (data not shown). Expres-sion analysis by Northern blot analysis confirmed that all het-erologous genes were expressed (data not shown).
To independently confirm these findings by a complemen-tary approach, we isolated a series of serotype AD hybrid diploid strains that contained combinations of the pka1A⌬,
pka2A⌬, pka1D⌬, and pka2D⌬ mutations and determined
which PKA catalytic subunit was functional in the hybrid by examining melanin and capsule production. AD hybrids were isolated following cell-cell fusions, and PCR analysis using both serotype A- and serotype D-specific primers confirmed the presence of both the serotype AMAT␣mating type and the serotype D MATamating type in each hybrid. Melanin and capsule were produced bypka2A⌬/pka1D⌬pka2D⌬(JKH324),
pka1A⌬ pka2A⌬/pka1D⌬ (JKH321), and pka2A⌬/pka1D⌬
(JKH323) hybrids, all of which express PKA1A, PKA2D, or both. Only the hybrid with no functional PKA catalytic sub-units (pka1A⌬pka2A⌬/pka1D⌬pka2D⌬; JKH322) was unable to produce melanin and capsule (Fig. 2). These results confirm that Pka1A and Pka2D are able to cross-complement the
pka2D⌬andpka1A⌬mutations in serotypes D and A,
respec-tively.
on September 8, 2020 by guest
http://ec.asm.org/
The serotype B Pka2B subunit is responsible for melanin production, while both Pka1B and Pka2B play roles in capsule production.To determine which of theC. gattiiPKA catalytic subunits, Pka1B or Pka2B, was responsible for melanin pro-duction inC. gattii, we deleted the entire open reading frames of both thePKA1Band thePKA2Bgenes. The deletions were confirmed by Southern blot and Northern analyses. The dele-tion strains were then grown onL-DOPA (L-, 4-dihydroxyphe-nylalanine) and Niger seed medium and incubated at 37°C for
12 to 14 h. The pka2B⌬ strain exhibited a severe defect in melanin production, whereas melanin production by the
pka1B⌬strain was indistinguishable from that of the serotype
B wild-type strain (Fig. 3, lane 1). These findings indicate that the Pka2 catalytic subunit is responsible for positively regulat-ing melanin production in serotype B.
In contrast, neither the pka1B⌬ mutant strain nor the
pka2B⌬mutant strain exhibited a demonstrable capsule defect
(Fig. 3, lane 2). We hypothesized that Pka1B and Pka2B might FIG. 1.PKA1A,PKA2D, andPKA2Bgenes complement defects conferred bypka1A⌬andpka2D⌬mutations. The ability of wild-type (WT) PKA catalytic subunits from serotypes (sero) A, D, and B to complement melanin and capsule production defects in strains bearing mutations in their counterparts from other serotypes was tested. (Lane 1) A wild-type serotype A strain (KN99␣), apka1A⌬mutant strain (JKH7), andpka1A⌬
mutant strains carrying wild-type alleles ofPKA1andPKA2from serotypes D and B (strains JKH242, JKH308, JKH179, and JKH297, respectively) were grown on Niger seed agar and incubated for 2 days at 37°C prior to being photographed. (Lane 2) The strains described for lane 1 were inoculated onto agar-based DME medium and incubated at 37°C for 2 days prior to being examined for capsule production by exclusion of India ink (⫻1,000 magnification). (Lane 3) A wild-type serotype D strain (JEC21), a pka2D⌬mutant strain (CDC85), andpka2D⌬mutant strains carrying wild-type alleles ofPKA1andPKA2from serotypes A and B (strains JKH287, JKH315, JKH247, and JKH299, respectively) were grown on Niger seed agar and incubated for 2 days at 30°C prior to being photographed. (Lane 4) The strains described for lane 3 were inoculated onto agar-based DME medium and incubated at 37°C for 2 days prior to being examined by exclusion of India ink (⫻1,000 magnification).
FIG. 2. Pka1A and Pka2D are functional in serotype AD hybrids. To test ifPKA1AandPKA2Dcan complement mutations in serotypes D and A, respectively, hybrid diploid serotype AD strains bearing various combinations ofpka1A⌬andpka2D⌬ mutations were isolated. (Lane 1) Wild-type (WT) serotype A and D strains (KN99␣and JEC20), as well aspka2A⌬(JKH4),pka1A⌬pka2A⌬(JKH96),pka1D⌬(JKH313), and
pka1D⌬pka2D⌬(JKH314) mutant strains and serotype AD diploids with the various combinations ofpka1⌬andpka2⌬mutations (JKH321 to JKH324), were grown on Niger seed medium and incubated for 2 days at 30°C prior to photographing. (Lane 2) The strains described for lane 1 were inoculated onto agar-based DME medium and incubated at 37°C for 2 days prior to being examined for capsule production by exclusion of India ink (⫻1,000 magnification). Three independent diploid strains with each genotype were examined in two independent experiments, and representative data are shown.
on September 8, 2020 by guest
http://ec.asm.org/
play a shared role in regulating capsule production. To test this, apka1B⌬pka2B⌬double-mutant strain was created via transformation of thepka1B⌬mutant with apka2B⌬ disrup-tion allele. As with the pka2B⌬ single mutant, the double mutant had a severe defect in melanin production (Fig. 3, lane 1). In addition, several independent double-mutant strains also exhibited a profound capsule defect that was not observed in either single-mutant strain, indicating that Pka1B and Pka2B play a redundant role in capsule production (Fig. 3, lane 2).
Pka1B and Pka2B have degenerate roles in mating.To ex-amine the roles of Pka1B and Pka2B in mating, thepka1B⌬
and pka2B⌬ single-mutant strains and the pka1B⌬ pka2B⌬
double-mutant strain were tested as mating partners with the
C. gattii MATastrain B4546 (molecular group VGIII) (6, 7).
After 4 days of growth on V8 medium (pH 7.0), profuse fila-mentation and basidial formation was observed in the mating between thepka2B⌬mutant and wild-type strain B4546. This was in comparison to the mating between the wild-type strains (R265⫻B4546) and the mating between thepka1B⌬mutant and wild-type strain B4546, in which only minimal filamenta-tion and basidial formafilamenta-tion were observed. With thepka1B⌬
pka2B⌬double mutant crossed to the wild-type strain, no
fila-mentation or basidial formation was observed (Fig. 4). These data provide evidence that the PKA catalytic subunits inC.
gattiihave overlapping roles and at least one functional PKA
catalytic subunit is required for mating. In addition, the Pka2 catalytic subunit appears to play an additional role in repress-ing matrepress-ing inC. gattii.
Protein kinase A catalytic and regulatory subunits interact in the two-hybrid system.To begin to address the mechanistic basis for the functional differences between the Pka1 and Pka2 catalytic subunits in divergent varieties and species, the yeast two-hybrid system was employed to assess interactions be-tween the subunits. First, we found that the protein kinase A regulatory subunit Pkr1 was capable of interacting with itself when fused to both the Gal4 DNA binding domain and the activation domain, and the magnitude of this interaction was reduced by the provision of 10 mM exogenous cyclic AMP (cAMP) (data not shown). Additionally, we found that the serotype A Pka1 catalytic subunit interacted much more ro-bustly with the Pkr1 regulatory subunit than did the Pka2 catalytic subunit, and the magnitudes of both the Pka1-Pkr1 and the Pka2-Pkr1 interaction were reduced by 10 mM cAMP, as assessed by monitoring Gal4-driven-galactosidase expres-sion (Fig. 5). Finally, the serotype D Pka1 and Pka2 subunits FIG. 3. Pka2B functions in regulating melanin, and both Pka1B
and Pka2B regulate capsule production. Thepka1B⌬,pka2B⌬, and
pka1B⌬pka2B⌬ mutants were examined for the ability to produce melanin and capsule. (Lane 1) A wild-type (WT) strain (R265) and strains containingpka1B⌬(JKH290),pka2B⌬(JKH293), andpka1B⌬
pka2B⌬(JKH317) mutations were grown onL-DOPA agar medium
and incubated for 12 to 14 h prior to being photographed. (Lane 2) The strains described for lane 1 were inoculated onto agar-based DME medium and incubated at 37°C for 2 days prior to being examined for capsule production by exclusion of India ink (⫻1,000 magnification). Three independent transformants with each genotype were examined in two independent experiments, and representative data are shown.
FIG. 4. Pka1B and Pka2B share a role in promotingC. gattiimating, and Pka2B has an additional role in repressing mating. The abilities of the␣wild type andpka1B⌬,pka2B⌬, andpka1B⌬pka2B⌬mutant strains to mate with theastrain B4546 were tested. Both the wild-type mating (R265) (panel 1) and thepka1B⌬mating (JKH290) (panel 2) produced only a few basidiospores. Thepka2B⌬mating (JKH293) (panel 3) was more robust, producing many more basidiospores. Thepka1B⌬pka2B⌬mating (JKH317) (panel 4) was completely devoid of basidiospore formation. Matings were performed as described in Materials and Methods. Three independent transformants from each genotype were tested in three independent experiments. Representative data are shown. Arrows point to the filaments, basidia, and basidiospores.
FIG. 5. Protein kinase A regulatory and catalytic subunit interac-tions in the yeast two-hybrid assay. Yeast two-hybrid reporter strains expressing Gal4 domains fused to Pkr1 and Pka1 or Pka2 from sero-type A or D as indicated were tested for-galatactosidase expression in the presence (⫹) and absence (⫺) of 10 mM exogenous cAMP. Samples were assayed in triplicate, and the standard errors of the means are presented as error bars.
on September 8, 2020 by guest
http://ec.asm.org/
interacted to a more modest extent with the Pkr1 regulatory subunit, and the provision of cAMP reduced the Pkr1-Pka2 interaction but not the Pkr1-Pka1 interaction (Fig. 5). Taken together, these findings suggest that differences in the interac-tion of the catalytic and regulatory subunits of protein kinase A or in the cAMP responsiveness of the complex could contrib-ute to functional differences between subunits.
DISCUSSION
We have taken advantage of the availability of the total genome sequences from three Cryptococcus serotypes (sero-type A,C. neoformansvar.grubii; serotype D,C. neoformans
var. neoformans; and serotype B, C. gattii) to examine the
evolution of two PKA catalytic subunits, Pka1 and Pka2.C.
gattii diverged from C. neoformans approximately 38.5 mya,
while the twoC. neoformansvarieties (C. neoformansvar.
gru-bii and C. neoformans var. neoformans) diverged from each
other around 18.5 mya (32).
An examination of theC. gattiiserotype B strain R265 ge-nome revealed thatC. gattiialso has two catalytic subunits that share 84% (Pka1) and 87% (Pka2) identity with their serotype A and D counterparts. The presence of two catalytic subunits
inC. gattiisuggests that a gene duplication event occurred over
38.5 mya, prior to the divergence ofC. gattiiandC. neoformans, resulting in two catalytic subunits in both species. We were interested in characterizing theC. gattiiPka1 and Pka2 sub-units to determine how the roles in regulating melanin and capsule production and mating were allocated between the two subunits in this more divergent species. The deletion of both
the PKA1B and PKA2B genes revealed that solely Pka2B is
responsible for regulating melanin production (Fig. 3, lane 1). However, a defect in capsule production was observed only when both thePKA1BandPKA2Bgenes were deleted (Fig. 3, lane 2), indicating that Pka1B and Pka2B share redundant roles in regulating capsule production.
Similar to the opposing roles played by Tpk1/3 and Tpk2 in regulating pseudohyphal growth in Saccharomyces cerevisiae
(22, 24), the diverse roles of the PKA catalytic subunits in mating inCryptococcusserotypes A, D, and B are consummate examples of the complexity of the PKA signaling pathway. In serotype A, the deletion of thePKA1gene (but notPKA2) in either parental strain results in loss of mating. The situation is more complex in serotype D, wherein a mating defect is de-tected only when both parental strains harbor apka2D⌬ mu-tation. Furthermore, unlike serotype A pka1A⌬ mutants,
pka2D⌬mutants (but notpka1D⌬mutants) have defects in cell
and nuclear fusion, leading to production of aberrant filaments (11). In C. gattii, the situation is different than that in the lineage of either serotype A or D because both the Pka1B and Pka2B subunits are required for mating, as indicated by the observation that mating still occurs when either thePKA1Bor
PKA2Bgene was individually deleted but no mating was
ob-served when both thePKA1BandPKA2Bgenes were deleted (Fig. 4). In addition to playing a redundant role that it shares with Pka1B in activating mating, Pka2B may play a role in repressing mating, as suggested by the observation that the mating of thepka2B⌬mutant with a tester strain resulted in more robust filamentation than was seen with the wild type (Fig. 4).
Our heterologous complementation studies, as well as our serotype AD hybrid diploid studies, showed that the wild-type
PKA2Dgene complemented the melanin and capsule defects
resulting from a pka1A⌬ mutation. Similarly, a wild-type
PKA1Agene complemented the melanin and capsule defects
of a pka2D⌬ mutation. The wild-type PKA2B gene, but not
PKA1B, was able to complement the melanin defect of the
pka1A⌬andpka2D⌬mutants. Interestingly, even though our
data show that both theC. gattii Pka1B and Pka2B subunits have roles in capsule production, only the wild-type PKA2B
gene, and not PKA1B, was able to complement the capsule defect of thepka1A⌬andpka2D⌬mutants. The failure of the
PKA1Bgene to complement was not due to a defect in
expres-sion or attributable to a mutation in the complementation allele, based on sequence analysis. One remaining possibility is that the lack of complementation of the capsule defect of the
pka1A⌬ and the pka2D⌬ mutant by the PKA1Bgene is the
result of the Pka1B subunit having, over time, lost its ability to promote capsule production in the divergent C. neoformans
serotype A and D lineages.
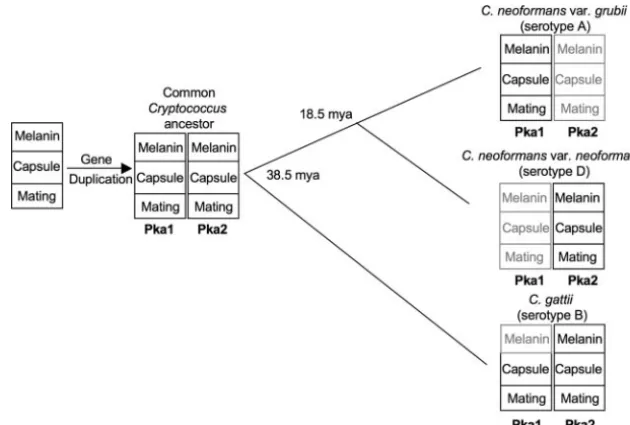
Based on the data presented here and our previous data showing that the serotype A Pka1A subunit and the serotype D Pka2D subunit have the same function despite having only 35% identity at the amino acid level (11), we propose a model in which theC. gattiiPka1B and Pka2B subunits are represen-tative of the PKA catalytic subunits shortly after the gene duplication event, when the two subunits both still retained ancestral shared functions (Fig. 5). Over time, the serotype A Pka2A subunit lost many of its original functions, with the exception of a minor role in melanin production, whereas the Pka1A subunit retained all of its original functions. On the contrary, in serotype D, the Pka1D subunit lost most of its original functions, whereas the Pka2D subunit retained its original functions (Fig. 6).
Our model is consistent with the neofunctionalization model proposed by Ohno (21). In this model, after gene duplication, one gene retains the ancestral function while selection pressure on the other gene is reduced, allowing it to lose the original functions and possibly gain new ones. Although neither Pka2A nor Pka1D showed any obvious new functions, as is predicted by Ohno’s model, this could be either because not enough time has elapsed since the gene duplication for these functions to have evolved or because we have not tested enough conditions to reveal novel functions. Alternatively, it is possible that in-stead of the subunits gaining functions, the Pka2A and Pka1D subunit genes may be in the process of becoming pseudogenes and will eventually be completely lost, leaving only one recog-nizable PKA catalytic subunit, Pka1A and Pka2D, in the re-spective serotypes.
The mechanism for the differential actions of Pka1 and Pka2 within each of the three serotypes is unclear. One possibility is that the Pka1 and Pka2 proteins interact differently with the regulatory subunit, Pkr1. Pkr1 is highly conserved among the serotypes (Pkr1B shares 89% amino acid identity with both Pkr1A and Pkr1D, and Pkr1A shares 95% amino acid identity with Pkr1D). Our yeast two-hybrid system data (Fig. 5) indi-cate that Pka1A, but not Pka2A, interacts strongly with Pkr1A. One explanation for this may lie in the amino acid residues in Pka1 and Pka2 that occur at positions in the catalytic subunit critical for binding to the regulatory subunit. Kim et al. (17)
on September 8, 2020 by guest
http://ec.asm.org/
have identified several residues in the PKA catalytic subunit that are necessary for interactions with the regulatory subunit. A comparison of the PKA catalytic subunit protein sequence analyzed by Kim et al. and the protein sequences of the Pka1 and Pka2 catalytic subunits from serotypes A, B, and D reveals that at least four of the critical amino acids (at amino acid positions 379, 381, 396, and 398) are substantially different between the Pka1 subunit (isoleucine, tryptophan, glutamine, and lysine at the respective positions) and the Pka2 subunit (arginine, phenylalanine, leucine, and glutamine at the respec-tive positions), although the residues are conserved within the Pka1 and Pka2 proteins of the respective serotypes. Another possibility is that Pka1 and Pka2 interact with different sub-strates or interact differentially with the same subsub-strates. Fi-nally, previous studies have indicated that the catalytic subunits of PKA have targets both in the cytoplasm and in the nucleus (9). Thus, it is possible that the Pka1 and Pka2 proteins are localized differently within the cell and may either act on dif-ferent targets depending on their location or, alternatively, be functional only if they are localized in one organelle or an-other. Further studies will be necessary to address these and other models.
Studies have utilized comparative approaches for closely related species to examine gene duplication and to test models of gene duplication. This is especially true for the well-char-acterized yeast Saccharomyces cerevisiae, in which a whole-genome duplication occurred and was subsequently followed by extensive gene loss and gene specialization (15, 25, 31). The majority of these studies have focused on gene and protein structures (e.g., references 10 and 15), although some studies have advanced a functional approach (e.g., reference 28). In our study, we implemented a gene function approach rather
than a gene structure approach by examining the roles of duplicated proteins in divergent, related species (C. gattii) to understand the origin of function in a more recently diverged pair of varieties (C. neoformansvar.grubiiandC. neoformans
var.neoformans). Similarly, we utilized inter- and intraspecific
complementation approaches, including the isolation and analysis of hybrid diploid strains, to decipher the functions of these signaling cascade genes. Both of these techniques may, in combination with the more commonly used gene structure comparison approach, have applications for deter-mining gene function and exadeter-mining the results of gene duplication events in other fungi, such as the sensu stricto strains ofSaccharomyces.
ACKNOWLEDGMENTS
We thank Alex Idnurm and Andrew Alspaugh for helpful sug-gestions in the preparation of the manuscript. We thank the Broad Institute for the serotype A and B genome sequences (http://www .broad.mit.edu/annotation/genome/cryptococcus_neoformans/Home.html and http://www.broad.mit.edu/annotation/genome/cryptococcus_neoformans _b/Home.html, respectively) and The Institute for Genomic Research for the serotype D genome sequence information (http://www.tigr.org/tdb/e2k1 /cna1/).
This research was funded in part by NIH Interdisciplinary AIDS Training grant AI07392-14 (J.K.H.) and RO1 grant AI39115 (J.H.).
REFERENCES
1.Alspaugh, J. A., J. R. Perfect, and J. Heitman.1997.Cryptococcus neofor-mansmating and virulence are regulated by the G-protein␣subunit GPA1 and cAMP. Genes Dev.11:3206–3217.
2.Bahn, Y.-S., G. M. Cox, J. R. Perfect, and J. Heitman. 2005. Carbonic anhydrase and CO2sensing duringCryptococcus neoformansgrowth,
differ-entiation, and virulence. Curr. Biol.15:2013–2020.
3.Bahn, Y.-S., J. K. Hicks, S. S. Giles, G. M. Cox, and J. Heitman.2004. Adenylyl cyclase-associated protein Aca1 regulates virulence and differenti-ation ofCryptococcus neoformansvia the cyclic AMP-protein kinase A cas-cade. Eukaryot. Cell3:1476–1491.
FIG. 6. Model for functional loss and speciation following duplication of the PKA catalytic subunits inCryptococcus. Prior to the divergence ofC. gattiifromC. neoformansaround 38.5 mya, a gene duplication event that resulted in two PKA catalytic subunits, Pka1 and Pka2, occurred. The catalytic subunits are involved in the regulation of at least three virulence factors that include melanin production, capsule production, and mating. InC. gattii, Pka1 and Pka2 both retain the majority of their original functions, with the exception of Pka1 having lost its ability to regulate melanin (indicated by gray shading). With the exception of a minor role for Pka2A in melanin production, serotype A Pka2A and serotype D Pka1D have lost their original functions, whereas serotype A Pka1A and serotype D Pka2D retain the original functions. Solid lines indicate branch points in evolution, and the numbers at the branch points indicate when the serotypes diverged.
on September 8, 2020 by guest
http://ec.asm.org/
4.Bahn, Y.-S., K. Kojima, G. M. Cox, and J. Heitman.2005. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryp-tococcus neoformans. Mol. Biol. Cell16:2285–2300.
5.Davidson, R. C., J. R. Blankenship, P. R. Kraus, M. de Jesus Berrios, C. M. Hull, C. D’Souza, P. Wang, and J. Heitman.2002. A PCR-based strategy to generate integrative targeting alleles with large regions of homology. Micro-biology148:2607–2615.
6.Fraser, J. A., S. S. Giles, E. C. Wenink, S. G. Geunes-Boyer, J. R. Wright, S. Diezmann, A. Allen, J. E. Stajich, F. S. Dietrich, J. R. Perfect, and J. Heitman.2005. Same-sex mating and the origin of the Vancouver Island Cryptococcus gattiioutbreak. Nature437:1360–1364.
7.Fraser, J. A., R. L. Subaran, C. B. Nichols, and J. Heitman.2003. Recapit-ulation of the sexual cycle of the primary fungal pathogenCryptococcus neoformansvar.gattii: implications for an outbreak on Vancouver Island. Eukaryot. Cell2:1036–1045.
8.Granger, D. L., J. R. Perfect, and D. T. Durack.1985. Virulence of Crypto-coccus neoformans: regulation of capsule synthesis by carbon dioxide. J. Clin. Investig.76:508–516.
9.Harootunian, A. T., S. R. Adams, W. Wen, J. L. Meinkoth, S. S. Taylor, and R. Y. Tsien.1993. Movement of the free catalytic subunit of cAMP-depen-dent protein kinase into and out of the nucleus can be explained by diffusion. Mol. Biol. Cell4:993–1002.
10.He, X., and J. Zhang.2005. Rapid subfunctionalization accompanied by prolonged and substantial neofunctionalization in duplicate gene evolution. Genetics169:1157–1164.
11.Hicks, J. K., C. A. D’Souza, G. M. Cox, and J. Heitman.2004. Cyclic-AMP dependent protein kinase catalytic subunits have divergent roles in virulence factor production in two varieties of the fungal pathogenCryptococcus neo-formans. Eukaryot. Cell3:14–26.
12.Hull, C. M., M.-J. Boily, and J. Heitman.2005. Sex-specific homeodomain proteins Sxi1␣and Sxi2a coordinately regulate sexual development in Cryp-tococcus neoformans. Eukaryot. Cell4:526–535.
13.Idnurm, A., Y.-S. Bahn, K. Nielsen, X. Lin, J. Fraser, and J. Heitman.2005. Deciphering the model pathoegenic fungusCryptococcus neoformans. Nat. Rev. Microbiol.3:753–764.
14.Idnurm, A., J. L. Reedy, J. C. Nussbaum, and J. Heitman.2004.Cryptococcus neoformansvirulence gene discovery through insertional mutagenesis. Eu-karyot. Cell3:420–429.
15.Kellis, M., B. W. Birren, and E. S. Lander.2004. Proof and evolutionary analysis of ancient genome duplication in the yeastSaccharomyces cerevisiae. Nature428:617–624.
16.Kidd, S. E., F. Hagen, R. L. Tscharke, M. Huynh, K. H. Bartlett, M. Fyfe, L. MacDougall, T. Boekhout, K. J. Kwon-Chung, and W. Meyer.2004. A rare genotype ofCryptococcus gattiicaused the cryptococcosis outbreak on Van-couver Island (British Columbia, Canada). Proc. Natl. Acad. Sci. USA101:
17258–17263.
17.Kim, C., X. Nguyen-Huu, and S. S. Taylor.2005. Crystal structure of a complex between the catalytic and regulatory (RI␣) subunits of PKA. Sci-ence307:690–696.
18.Loftus, B. J., E. Fung, P. Roncaglia, D. Rowley, P. Amedeo, D. Bruno, J. Vamathevan, M. Miranda, I. J. Anderson, J. A. Fraser, J. E. Allen, I. E.
Bosdet, M. R. Brent, R. Chiu, T. L. Doering, M. J. Donlin, C. A. D’Souza, D. S. Fox, V. Grinberg, J. Fu, M. Fukushima, B. J. Haas, J. C. Huang, G. Janbon, S. J. M. Jones, H. L. Koo, M. I. Krzywinski, J. K. Kwon-Chung, K. B. Lengeler, R. Maiti, M. A. Marra, R. E. Marra, C. A. Mathewson, T. G. Mitchell, M. Pertea, F. R. Riggs, S. L. Salzberg, J. E. Schein, A. Shvartsbeyn, H. Shin, M. Shumway, C. A. Specht, B. B. Suh, A. Tenney, T. R. Utterback, B. L. Wickes, J. R. Wortman, N. H. Wye, J. W. Kronstad, J. K. Lodge, J. Heitman, R. W. Davis, C. M. Fraser, and R. W. Hyman.2005. The genome of the basidiomycetous yeast and human pathogenCryptococcus neoformans. Science307:1321–1324.
19.Moore, T. D. E., and J. C. Edman.1993. The␣-mating type locus of Cryp-tococcus neoformanscontains a peptide pheromone gene. Mol. Cell. Biol.
13:1962–1970.
20.Nielsen, K., G. M. Cox, P. Wang, D. L. Toffaletti, J. R. Perfect, and J. Heitman. 2003. Sexual cycle ofCryptococcus neoformansvar.grubiiand virulence of congenicaand␣isolates. Infect. Immun.71:4831–4841. 21.Ohno, S. 1970. Evolution by gene duplication. Springer-Verlag, New
York, NY.
22.Pan, X., and J. Heitman.2002. Protein kinase A operates a molecular switch that governs yeast pseudohyphal growth. Mol. Cell. Biol.22:3981–3993. 23.Pukkila-Worley, R., Q. D. Gerrald, P. R. Kraus, M.-J. Boily, M. J. Davis,
S. S. Giles, G. M. Cox, J. Heitman, and A. Alspaugh.2005. Transcriptional network of multiple capsule and melanin genes governed by theCryptococcus neoformanscyclic AMP cascade. Eukaryot. Cell4:190–201.
24.Robertson, L. S., and G. R. Fink.1998. The three yeast A kinases have specific signaling functions in pseudohyphal growth. Proc. Natl. Acad. Sci. USA95:13783–13787.
25.Scannell, D. R., K. P. Byrne, J. L. Gordon, S. Wong, and K. H. Wolfe.2006. Multiple rounds of speciation associated with reciprocal gene loss in polyploid yeasts. Nature440:341–345.
26.Sherman, F.1991. Getting started with yeast, p. 3–21.InC. Guthrie and G. R. Fink (ed.), Methods in enzymology, vol. 194. Academic Press, Inc., San Diego, CA.
27.Toffaletti, D. L., T. H. Rude, S. A. Johnston, D. T. Durack, and J. R. Perfect.
1993. Gene transfer inCryptococcus neoformansby use of biolistic delivery of DNA. J. Bacteriol.175:1405–1411.
28.van Hoof, A.2005. Conserved functions of yeast genes support the duplica-tion, degeneration and complementation model for gene duplication. Ge-netics171:1455–1461.
29.Wang, P., M. E. Cardenas, G. M. Cox, J. R. Perfect, and J. Heitman.2001. Two cyclophilin A homologs with shared and distinct functions important for growth and virulence ofCryptococcus neoformans. EMBO Rep.2:511–518. 30.Waugh, M. S., C. B. Nichols, C. M. DeCesare, G. M. Cox, J. Heitman, and
J. A. Alspaugh.2002. Ras1 and Ras2 contribute shared and unique roles in physiology and virulence ofCryptococcus neoformans. Microbiology148:191– 201.
31.Wolfe, K. H., and D. C. Shields.1997. Molecular evidence for an ancient duplication of the entire yeast genome. Nature387:708–713.
32.Xu, J., R. J. Vilgalys, and T. G. Mitchell.2000. Multiple gene genealogies reveal dispersion and hybridization in the human pathogenic fungus Cryp-tococcus neoformans. Mol. Ecol.9:1471–1481.