0095-1137/07/$08.00⫹0 doi:10.1128/JCM.02451-06
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Multiple-Locus Variable-Number Tandem-Repeat Assay Analysis of
Methicillin-Resistant
Staphylococcus aureus
Strains
䌤
Fred C. Tenover,
1* Rebekah R. Vaughn,
2Linda K. McDougal,
1Gregory E. Fosheim,
1and John E. McGowan, Jr.
2Division of Healthcare Quality Promotion, Centers for Disease Control and Prevention, Atlanta, Georgia 30333,1and Rollins School of Public Health, Emory University, Atlanta, Georgia 303222
Received 6 December 2006/Returned for modification 10 March 2007/Accepted 2 May 2007
Our objective was to determine if a multiple-locus variable-number tandem-repeat assay (MLVA) for
Staphylococcus aureus could predict pulsed-field gel electrophoresis (PFGE) types (i.e., USA types), thus allowing us to replace PFGE with a simpler and more rapid typing method. One hundred three well-characterized isolates representing 13 major lineages ofS. aureuswere tested by both PFGE and MLVA. MLVA was performed using a rapid DNA extraction technique and PCR primers forsdrCDE,clfA,clfB,sspA, andspa.
PFGE was performed with genomic DNA fragments generated using SmaI, as per CDC protocols. Banding patterns were analyzed both visually and with BioNumerics software. All isolates were typeable with MLVA and PFGE. MLVA patterns were highly reproducible. PFGE separated the isolates into 13 types with 42 subtypes. Using any band difference to designate a novel MLVA type, MLVA produced 45 types, including 9 clusters containing multiple isolates. Using BioNumerics and a cutoff of >75% relatedness, MLVA produced 28 types, 11 of which contained >1 isolate. Epidemiologically related outbreak isolates of USA300-0114 from five states clustered in one MLVA pattern. USA100 isolates were present in several unrelated (<40%) MLVA types. A cutoff of >80% separated outbreak strains of USA300-0114 into three distinct MLVA types. MLVA did not differentiate community methicillin-resistant S. aureus (MRSA) lineages (USA300, USA400, USA1000, and USA1100) from health care MRSA lineages (USA100, USA200, or USA500). The ability of MLVA to differen-tiate among strains that are indistinguishable by PFGE may be of epidemiologic value and warrants further study.
Infections caused by methicillin-resistantStaphylococcus au-reus(MRSA) have traditionally been a problem in health care settings (8, 15). More recently, MRSA has been recognized as a cause of significant infections in community settings world-wide (2, 7, 14). Having access to an accurate strain typing method is critical for understanding the changing epidemiol-ogy of MRSA infections (12, 18) and for evaluating the efficacy of outbreak intervention and prevention strategies (20). The ability to type MRSA strains rapidly in clinical microbiology laboratories and to differentiate between the multiresistant MRSA strains associated with health care facilities and the more susceptible community-associated MRSA strains could improve public health and infection control efforts to reduce MRSA spread in hospitals and communities and to guide em-pirical therapy in community settings (14, 19).
A system of nomenclature for S. aureus pulsed-field gel electrophoresis (PFGE) results, known as USA types, has been adopted nationally and provides scientists with a common lan-guage for describing typing results (10, 11). The USA types have been validated using multilocus sequence typing (MLST) andspatyping (11). USA types determined by PFGE further differentiate the strains classified by MLST as ST5 or ST8 into epidemiologically and biologically distinct subgroups. For
ex-ample, ST8 strains can be divided by PFGE into USA300, the most commonly isolated community-associated MRSA strain in the United States (7, 13, 17, 18), and USA500, a classic multidrug resistant health care-associated MRSA strain (11). These PFGE types have different antibiograms and toxin pro-files. Similarly, PFGE types USA100 and USA800 are biolog-ically distinct subgroups of ST5. Nonetheless, PFGE requires 72 h to perform, is expensive compared to PCR-based typing, requires specialized training for performance and interpreta-tion, and is not readily available in most clinical microbiology laboratories.
Reports by Sabat et al. (16), Malachowa et al. (9), and Gilbert et al. (5) suggest that multiple-locus variable-number tandem-repeat assays (MLVA) could discriminate among MRSA strain types while providing results that paralleled the results of PFGE. The MLVA assay used by Sabat et al. tar-geted variable-number tandem repeats in the clfA, clfB,
sdrCDE, spa, and sspA loci and used gel electrophoresis to analyze the banding patterns (16). The individual numbers of repeats in the fragments generated were not determined, which some investigators consider a drawback to the method (19). Francois et al. expanded the number of loci investigated and used a BioAnalyzer to improve the analysis of the PCR products (3). Others investigators have evaluated the staphy-lococcal interspersed repeat units for typing; however, this analysis required multiple agarose gels for comparisons (6).
The goal of our study was to determine whether MLVA was simple enough to perform in a routine clinical microbiology laboratory and whether MLVA patterns could predict PFGE
* Corresponding author. Mailing address: Division of Healthcare Quality Promotion (G-08), Centers for Disease Control and Preven-tion, 1600 Clifton Rd. NE, Atlanta, GA 30333. Phone: (404) 639-3375. Fax: (404) 639-1381. E-mail: [email protected].
䌤Published ahead of print on 9 May 2007.
2215
on May 16, 2020 by guest
http://jcm.asm.org/
USA types. Thus, we assessed the technical performance of MLVA and compared the results of MLVA to the results of PFGE for a diverse set of MRSA and methicillin-susceptibleS. aureusisolates, including subtypes from known outbreaks de-fined by epidemiologic and microbiologic data.
MATERIALS AND METHODS
Bacterial strains.A group of 103 community- and health care-associated MRSA isolates and two methicillin-susceptible isolates (USA900 and USA1200) were selected from the strain collections of the Centers for Disease Control and
Prevention (CDC) and Project ICARE (4).S. aureusisolates were selected to
achieve a diversity of PFGE types. The types included USA100 (n⫽ 24),
USA200 (n⫽5), USA300 (n⫽45), USA400 (n⫽1), USA500 (n⫽3), USA600
(n⫽1), USA700 (n⫽3), USA800 (n⫽12), USA900 (n⫽1), USA1000 (n⫽
2), USA1100 (n⫽1), USA1200 (n⫽1), and Iberian clone (n⫽2) (1). Two
additional isolates did not belong to any specific USA type (pattern 0441). Included in the collection were epidemiologically related sets of isolates of subtype USA300-0114 from outbreaks in the following states: California (four isolates), Colorado (four isolates), Georgia (five isolates), Illinois (five isolates), Louisiana (five isolates), and Pennsylvania (four isolates).
Preparation of DNA.Bacterial isolates were subcultured in Trypticase soy agar II containing 5% sheep blood (BDDS, Franklin Lakes, NJ) for 24 h at 37°C.
Total genomic DNA was extracted fromS. aureusisolates using a GenElute
bacterial genomic DNA kit (Sigma-Aldrich, St. Louis, MO), following the
man-ufacturer’s protocol. Purified DNA was stored at⫺20°C.
MLVA typing.The multiplex PCR assay containingclfA,clfB,sdrCDE,spa,
andsspAprimers was performed initially as described by Sabat et al. (16). PCR
assay mixtures contained 10l ofTaqPCR Master Mix, 2.5 units ofTaqDNA
polymerase, 1.5 mM MgCl2, 200 M of each deoxynucleoside triphosphate
(QIAGEN, Valencia, CA), 0.5M of SspA-F (forward) and SspA-R (reverse),
1.0 M of ClfA-F and ClfA-R, 1.0M of ClfB-F and ClfA-R, 1.5 l of
sdrCDE-F and sdrCDE-R, 0.5M of Spa-F and Spa-R, 1l of DNA template
(⬃50 ng), and 1l PCR-grade water (Fisher Biotech, Fair Lawn, NJ) to make a
final volume of 20l. In later reactions, the amplification ofspausing Spa-F and
Spa-R was performed separately since these primers generated a large amount of
product. ThespaPCR assay mixture included 10l ofTaqPCR Master Mix as
used above, 0.5M Spa-F and Spa-R, 1l of DNA template (⬃50 ng) and 8l
of PCR-grade water. Amplification of DNA fragments occurred with an initial denaturation at 94°C for 5 min followed by 25 cycles of 30 s at 94°C, 30 s at 55°C, and 1 min at 72°C, with a final extension at 72°C for 5 min on a Bio-Rad
MyCycler 1500 (Hercules, CA). Five microliters of thespaPCR product was
added to 20l of the multiplex PCR for analysis in 2% agarose gels.
Visualization of the PCR product.Three microliters of the combined multiplex
andspaPCR products was electrophoresed in 2% UltraPure agarose (Gibco
BRL, Grand Island, NJ) gels with 0.5⫻TBE buffer (Tris-borate-EDTA [pH 8])
for 50 min at 80 V/cm. As a size standard, 3l of 100-bp ladder (Invitrogen,
Carlsbad, CA) at a concentration of 0.5g/l was included in every fifth sample
well. Gels were run in 1 liter of 0.5⫻TBE buffer. MLVA band profiles were
visualized on a UV illuminator and photographed. MLVA profiles were analyzed visually first, using the criteria of Sabat et al. that any pattern differing by one or more bands was a distinct type (16). Then, the MLVA patterns were reanalyzed using BioNumerics software (version 4.1; Applied Maths, Belgium) using the following tolerance settings: optimization, 0.5%, and position tolerance, 1.25%. Patterns were clustered using Dice’s coefficients and the unweighted-pair group method using average linkages into MLVA types. The effect of grouping similar MLVA patterns together into a smaller number of MLVA types was examined
using cutoffs of⬎75% and⬎80% relatedness. We specifically evaluated the
ability of the MLVA type to predict a USA pulsed-field type and the ability of the MLVA to cluster epidemiologically related isolates into the same cluster.
Reproducibility studies.To confirm the reproducibility of the MLVA typing procedure, DNA was extracted from two isolates (one USA100 isolate and one USA300 isolate) four times each on four consecutive days (a total of 16 extrac-tions and PCR assays for each of the two isolates). MLVA reacextrac-tions were run separately on each DNA extract, and the products were compared on agarose gels.
RESULTS AND DISCUSSION
Technical assessment.All 103S. aureusisolates were type-able by MLVA and PFGE. MLVA was simple enough to be
performed in a clinical laboratory, although it did take several weeks to optimize the results. This included running thespa
primers separately from the other four primer sets to get more balanced gel patterns where thespaproducts did not obscure
sspA products. Fifteen isolates could readily be typed by MLVA in a single day if pure cultures were available.
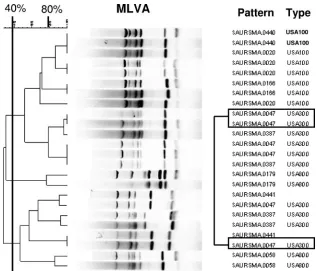
Analysis of USA type strains using PFGE and MLVA. Over-all, PFGE separated the 103S. aureusisolates into 13 major pulsed-field types (the 12 USA types plus an additional type referred to as “Iberian clone” [1]) based on BioNumerics anal-ysis using⬎80% relatedness to define the USA types, as pre-viously described (11). Among the 13 pulsed-field types, there were 42 subtypes (i.e., unique pulsed-field patterns). The PFGE subtypes (such as USA300-0114) typically represented changes in the PFGE banding patterns (up to six band differ-ences) that were still⬎80% related to their parent USA type strains (11). Given the band variability in the PFGE patterns, we expected to see some variation in the MLVA patterns for isolates within a given pulsed-field type. The key question of this study was how often the patterns for a group of isolates of the same PFGE USA type appeared to be unrelated by MLVA. Based on visual analysis of the MLVA patterns using the criteria established by Sabat et al. (where any band change was classified as a distinct MLVA type), MLVA produced 45 distinct patterns. Nine clusters of isolates shared the same MLVA patterns, while 36 organisms had unique MLVA pat-terns (data not shown). Using these stringent criteria for de-fining MLVA types (where each pattern is a separate MLVA type) epidemiologically defined outbreak strains of USA300-0114 were separated into four different clusters. MLVA also separated USA100 isolates into 9 distinct types, USA200 into 5 types, USA500 into 2 types, USA700 into 3 types, USA800 into 10 types, and the Iberian isolates into 2 types. Thus, visual analysis using any band difference to establish unique MLVA types was abandoned because it did not group together strains that were known to be epidemiologically related or highly related by PFGE.
Using BioNumerics software to facilitate analysis, two po-tential cutoff values of⬎80% relatedness and⬎75% related-ness were investigated in order to group the MLVA patterns into MLVA types that more closely matched the epidemiologic clustering. The former cutoff was selected because it is the gold standard for defining pulsed-field types (11). The latter cutoff was selected because on initial review it grouped epidemiolog-ically related strains of the same PFGE subtype (USA300-0114) from five of six outbreaks together in a single MLVA cluster. No matter which cutoff was used, the USA type strains for USA300 and USA500 were indistinguishable by MLVA (Fig. 1), suggesting that MLVA may have difficulty grouping ST8 isolates (which include USA300, USA500, and the Iberian clone) into patterns that parallel the PFGE types, which was one of the goals of the study. Using the⬎80% relatedness cutoff, the number of MLVA types was reduced from 45 to 38 and included 11 clusters of⬎1 organism and 27 unique MLVA patterns (Table 1). At 80% relatedness, strains of USA300-0114 from well-defined outbreaks in California, Georgia, and Louisiana were separated into unique MLVA types, although they were still closely related (at approximately 77%). Out-break strains from Colorado and Pennsylvania clustered in the same MLVA type with the isolates from Georgia. Three of the
on May 16, 2020 by guest
http://jcm.asm.org/
four California outbreak isolates had a distinctly lower spa
product (Fig. 2). Using the cutoff of⬎80% relatedness, only 17 of the 24 USA100 isolates had the same MLVA type; 5 other MLVA types, some at less than 40% relatedness, were also observed. Twenty-three USA300-0114 isolates grouped in one MLVA type, while 14 USA300 isolates of six highly related PFGE subtypes, including several USA300-0114 isolates, grouped in a second MLVA type. Interestingly, USA800 iso-lates, which, like USA100 isoiso-lates, are MLST ST5 (and part of MLST clonal complex 5), were separated into eight different MLVA types, six of which contained only a single organism. Thus, USA100 isolates were more homogeneous by MLVA typing than were USA800 isolates. Yet, one USA800 isolate and one USA1000 isolate were clustered among the USA100 isolates by MLVA, even at the more stringent 80% similarity level. It would be particularly difficult to predict that an MRSA isolate belonged to the USA800 lineage based on its MLVA type.
Using a cutoff value of ⬎75% relatedness increased the number of isolates in each MLVA type, thus reducing the number of MLVA types from 38 to 28. Using the⬎75% cutoff with BioNumerics software clustered several MLVA patterns together that would likely have been designated as unrelated by visual categorization, such as the USA200 isolates (Table 1). Forty of the 45 USA300 isolates (including 8 different PFGE USA300 subtypes) clustered into a single MLVA type at 75% relatedness, including the USA300-0114 outbreak strains from California, Colorado, Georgia, Louisiana, and Pennsylvania. Interestingly, the USA300-0114 isolates from the Illinois out-break still clustered separately in a different MLVA type, per-haps because of a smallersspband (Fig. 2, second band from the right). The fact that MLVA differentiated among PFGE subtypes is especially apparent for the USA300-0047 isolates, which produced MLVA patterns that were⬍40% related (Fig. 3). In this case, the USA300-0047 isolates, although indistin-guishable by PFGE, were all epidemiologically distinct. The ability of MLVA to differentiate among USA300-0114 out-breaks and among epidemiologically unrelated USA300-0047 isolates from different states (using the 80% cutoff value) may be of value in long-term epidemiologic studies, perhaps high-lighting biological differences among the isolates that are
masked by PFGE, while using the⬎75% cutoff may have more utility for local hospitals for screening isolates from potential hospital outbreaks.
To further assess the reproducibility of the MLVA method, DNA was extracted from two isolates, one USA100 and one USA300, on four consecutive days. Each lysate was used as DNA template for four separate MLVA typing experiments. Thus, there were 16 MLVA patterns generated for each or-ganism. Although some variation in band intensity was noted, the MLVA patterns for each isolate were indistinguishable (data not shown). Thus, as previously noted by Sabat and coworkers, the intralaboratory reproducibility of the MLVA patterns was very high. This is reassuring for laboratories that would consider using MLVA as a rapid screening method for hospital outbreak studies.
[image:3.585.53.272.67.226.2]Clearly, the utility of the MLVA procedure depends upon how the test is used. Using a cutoff of ⬎75% relatedness, MLVA could have utility as a screening test to determine whether isolates collected over a relatively short period of time in a single hospital were likely to be part of an outbreak cluster. Among outbreak isolates from five states that clustered to-gether, only one isolate, from the California outbreak, had a slightly different MLVA pattern (although, using⬎75% relat-edness, it would still have been classified as the same MLVA type). Thus, the reproducibility of the method is high for known outbreak strains. The fact that many of the USA100 isolates clustered together in the same MLVA type is also an encouraging result. However, the fact that the USA800 isolates were split among six quite different MLVA types using a 75% relatedness cutoff suggests that the variability of the patterns observed may be strain dependent. Grouping of the MLVA patterns into MLVA types can be done visually, although using any band difference in a pattern to designate MLVA types
FIG. 1. Comparison of PFGE and MLVA results for previously described USA type strains (11).
TABLE 1. Number of unique strain types defined by PFGE and MLVA
USA
type (n)
No. of PFGE
subtypesa
No. of MLVA types by
⬎80% cutoff
No. of MLVA types by
⬎75% cutoff
Total
types Clusters
b
Unique Total
types Clusters
b
Unique
100 (24) 10 6 3 3 4 2 2
200 (5) 4 3 1 2 1 1 0
300 (45) 7 6 4 2 3 2 1
400 (1) 1 1 1 1 1
500 (3) 3 2 1 1 2 1 1
600 (1) 1 1 1 1 1
700 (3) 3 3 3 2 1 1
800 (12) 6 8 2 6 7 3 4
900 (1) 1 1 1 1 1
1000 (2) 1 1 1 1 1
1100 (1) 1 1 1 1 1
1200 (1) 1 1 1 1 1
Iberian (2) 2 2 2 1 1 0
Pattern 0441 (2)
1 2 2 2 2
Total (103) 42 38 11 27 28 11 17
aSubtypes all show⬎80% relatedness by Dice’s coefficients using the
un-weighted-pair group method using average linkage clustering as defined by BioNumerics software.
bClusters are groups of⬎1 organism that show related MLVA patterns as
defined by BioNumerics software at the given cutoff value.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.299.540.475.683.2]overdiscriminates among strains that are related epidemiolog-ically by PFGE. Using strain typing software aids in clustering related isolates over time and for archiving and long-term storage of data.
[image:4.585.136.451.68.351.2]In summary, MLVA would likely be helpful for identifying outbreaks in a hospital setting, where the major goal is to determine if isolates that are epidemiologically linked and collected over a relatively short period of time are related or
FIG. 2. MLVA typing ofS. aureusisolates showing the separation of USA300-0114 isolates from a California outbreak into two distinct clusters.
FIG. 3. MLVA typing ofS. aureusisolates showing separation of USA300-0047 isolates into two divergent clusters.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.133.450.439.710.2]unrelated. Fifteen isolates can be typed in a single working day, the equipment required is relatively inexpensive compared to that required for PFGE, and only modest training is required to set up the assay. The results can be interpreted visually without the need for additional software; however, we found that for longitudinal studies, a software package greatly facil-itated data management. MLVA would be particularly useful for laboratories already using PCR-based assays, since they already have the expertise for performing amplification assays. However, our data, using a set of geographically diverse MRSA strains isolated from across the United States, showed that MLVA clustered isolates collected over a period of several years in a manner different from PFGE. Thus, MLVA cannot be used to predict PFGE types The biological reasons for the differences in the patterns, such as those seen in the Illinois and California outbreaks, are intriguing and merit further study.
ACKNOWLEDGMENTS
We thank Robert Skov and Richard Goering for helpful discussions. Phase 5 of Project ICARE is supported in part by unrestricted research grants to the Rollins School of Public Health of Emory University from Astra-Zeneca Pharmaceuticals, Wilmington, DE; Elan Pharmaceuticals, San Diego, CA; Johnson & Johnson Pharma-ceutical Research & Development, LLC, Raritan, NJ; Pfizer Inc., New York, NY; and 3 M Health Care Products, St. Paul, MN.
The findings and conclusions in this publication are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention.
REFERENCES
1.Aires de Sousa, M., I. S. Sanches, M. L. Ferro, M. J. Vaz, Z. Saraiva, T. Tendeiro, J. Serra, and H. de Lencastre.1998. Intercontinental spread of a
multidrug-resistant methicillin-resistantStaphylococcus aureusclone. J. Clin.
Microbiol.36:2590–2596.
2.Deresinski, S.2005. Methicillin-resistantStaphylococcus aureus: an
evolu-tionary, epidemiologic, and therapeutic odyssey. Clin. Infect. Dis.40:562–
573.
3.Francois, P., A. Huyghe, Y. Charbonnier, M. Bento, S. Herzig, I. Topolski, B. Fleury, D. Lew, P. Vaudaux, S. Harbarth, W. van Leeuwen, A. van Belkum, D. S. Blanc, D. Pittet, and J. Schrenzel.2005. Use of an automated multiple-locus, variable-number tandem repeat-based method for rapid and
high-throughput genotyping ofStaphylococcus aureusisolates. J. Clin. Microbiol.
43:3346–3355.
4.Fridkin, S. K., C. D. Steward, J. R. Edwards, E. R. Pryor, J. E. McGowan, Jr., L. K. Archibald, R. P. Gaynes, F. C. Tenover, and Project Intensive Care Antimicrobial Resistance Epidemiology (ICARE) Hospitals.1999. Surveil-lance of antimicrobial use and antimicrobial resistance in United States
hospitals: Project ICARE phase 2. Clin. Infect. Dis.29:245–252.
5.Gilbert, F. B., A. Fromageau, L. Gelineau, and B. Poutrel.2006.
Differen-tiation of bovineStaphylococcus aureusisolates by use of polymorphic
tan-dem repeat typing. Vet. Microbiol.117:297–303.
6.Hardy, K. J., D. W. Ussery, B. A. Oppenheim, and P. M. Hawkey.2004.
Distribution and characterization of staphylococcal interspersed repeat units
(SIRUs) and potential use for strain differentiation. Microbiology150:4045–
4052.
7.Kazakova, S. V., J. C. Hageman, M. Matava, A. Srinivasan, L. Phelan, B. Garfinkel, T. Boo, S. McAllister, J. Anderson, B. Jensen, D. Dodson, D. Lonsway, L. K. McDougal, M. Arduino, V. J. Fraser, G. Killgore, F. C. Tenover, S. Cody, and D. B. Jernigan.2005. A clone of methicillin-resistant
Staphylococcus aureusamong professional football players. N. Engl. J. Med.
352:468–475.
8.Lowy, F.1998.Staphylococcus aureusinfections. N. Engl. J. Med.339:520– 532.
9.Malachowa, N., A. Sabat, M. Gniadkowski, J. Krzyszton-Russjan, J. Empel, J. Miedzobrodzki, K. Kosowska-Shick, P. C. Appelbaum, and W. Hryniewicz.
2005. Comparison of multiple-locus variable-number tandem-repeat analysis with pulsed-field gel electrophoresis, spa typing, and multilocus sequence typing
for clonal characterization ofStaphylococcus aureusisolates. J. Clin. Microbiol.
43:3095–3100.
10.McDougal, L. K., J. B. Patel, and F. C. Tenover.2005. Characterization of
two new community-associated oxacillin resistant Staphylococcus aureus
pulsed field types consisting of U.S. isolates that carry SCCmecIV and the Panton-Valentine leukocidin (PVL) gene, abstr. C-220, p. 144. Abstr. 105th Gen. Meet. Am. Soc. Microbiol., Atlanta, GA.
11.McDougal, L. K., C. D. Steward, G. E. Killgore, J. M. Chaitram, S. K. McAllister, and F. C. Tenover.2003. Pulsed-field gel electrophoresis typing
of oxacillin-resistantStaphylococcus aureusisolates from the United States:
establishing a national database. J. Clin. Microbiol.41:5113–5120.
12.Melles, D. C., R. F. Gorkink, H. A. Boelens, S. V. Snijders, J. K. Peeters, M. J. Moorhouse, P. J. van der Spek, W. B. van Leeuwen, G. Simons, H. A. Verbrugh, and A. van Belkum.2004. Natural population dynamics and
ex-pansion of pathogenic clones ofStaphylococcus aureus. J. Clin. Investig.
114:1732–1740.
13.Moran, G. J., A. Krishnadasan, R. J. Gorwitz, G. E. Fosheim, L. K. McDougal, R. B. Carey, and D. A. Talan.2006. Methicillin-resistantS. aureusinfections
among patients in the emergency department. N. Engl. J. Med.355:666–674.
14.Naimi, T. S., K. H. LeDell, K. Como-Sabetti, S. M. Borchardt, D. J. Boxrud, J. Etienne, S. K. Johnson, F. Vandenesch, S. Fridkin, C. O’Boyle, R. N. Danila, and R. Lynfield.2003. Comparison of community- and health
care-associated methicillin-resistantStaphylococcus aureusinfection. JAMA290:
2976–2984.
15.Panlilio, A. L., D. H. Culver, R. P. Gaynes, S. Banerjee, T. S. Henderson, J. S. Tolson, and W. J. Martone.1992. Methicillin-resistantStaphylococcus aureus
in U.S. hospitals, 1975–1991. Infect. Control Hosp. Epidemiol.13:582–586.
16.Sabat, A., J. Krzyszton-Russjan, W. Strzalka, R. Filipek, K. Kosowska, W. Hryniewicz, J. Travis, and J. Potempa.2003. New method for typing Staph-ylococcus aureusstrains: multiple-locus variable-number tandem repeat anal-ysis of polymorphism and genetic relationships of clinical isolates. J. Clin.
Microbiol.41:1801–1804.
17.Seybold, U., E. V. Kourbatova, J. G. Johnson, S. J. Halvosa, Y. F. Wang, M. D. King, S. M. Ray, and H. M. Blumberg.2006. Emergence of
commu-nity-associated methicillin-resistantStaphylococcus aureusUSA300 genotype
as a major cause of health care-associated bloodstream infections. Clin.
Infect. Dis.42:647–656.
18.Tenover, F. C., L. K. McDougal, R. V. Goering, G. Killgore, S. J. Projan, J. B. Patel, and P. M. Dunman.2006. Characterization of a strain of
community-associated methicillin-resistantStaphylococcus aureuswidely disseminated in
the United States. J. Clin. Microbiol.44:108–118.
19.van Belkum, A.2007. Tracing isolates of bacterial species by multilocus variable number of tandem repeat analysis (MLVA). FEMS Immunol. Med.
Microbiol.49:22–27.
20.Wertheim, H. F., W. B. van Leeuwen, S. Snijders, M. C. Vos, A. Voss, C. M. Vandenbroucke-Grauls, J. A. Kluytmans, H. A. Verbrugh, and A. van Belkum.
2005. Associations betweenStaphylococcus aureusgenotype, infection, and
in-hospital mortality: a nested case-control study. J. Infect. Dis.192:1196–1200.

![(2E) 2 [(3E) 4 Phenylbut 3 en 2 ylidene]hydrazinecarboxamide](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)