Copyright 0 1974 AmericanSociety for Microbiology Printed in U.S.A.
Sendai Virion
Transcriptase Complex: Polypeptide
Composition
and
Inhibition
by
Virion
Envelope
Proteins
PRESTON A. MARX, ALLEN PORTNER, ANDD. W. KINGSBURY
Laboratoriesof Virology and Immunology,St.Jude Children's ResearchHospital,and The University of Tennessee MedicalUnits, Memphis, Tennessee 38101
Received forpublication 3 August 1973
Sendaivirions, disrupted in 2%Triton X-100 in 1 M KCl,wereseparatedinto nucleocapsids and envelope proteins by centrifugation. The nucleocapsids, representing 46% of the virion proteins, hadabuoyant density of 1.29 gm/cm3in
D2Osucrose. RNA-dependent transcriptase activity associated with them hada ninefold greater specific activity than transcriptase assayed in unfractionated detergent-disrupted virions. These enzyme-active nucleocapsids contained only two polypeptides, thelargest virion polypeptide (molecular weight 75,000) and the nucleocapsid structure unit (molecular weight 60,000). Virion envelope proteins, either glycoproteins or nonglycosylated matrix protein, inhibited nucleocapsid-associated polymerase activity; brief heat denaturation abolished their inhibitory activity. Yeast RNA stimulated nucleocapsid-associated enzyme, suggestingthat stimulatory polyanions act atthe enzyme-template level.
RNA-dependent RNA polymerase (tran-scriptase) has been described in Sendaivirions (8, 13) and in nucleocapsid-like structures
(transcriptive complexes) isolated from Sendai virus-infected cells late in infection (12). The enzyme-active structures from infected cells contained two polypeptides. One of these mi-grated in sodium dodecyl
sulfate-polyacryla-mide gels like the largest virion polypeptide
(molecular weight 75,000). The other migrated like thenucleocapsidstructure unit (molecular
weight 60,000). These findings suggested that the largest virion polypeptide is involved in
transcriptase function. Althoughstructures iso-lated from virions with properties of nucleocap-sids were shown to contain some transcriptase activity (8), theirpolypeptide composition was notdetermined. If thelargest virion polypeptide
is involved in primary transcription of virion
RNA, as wellasinsecondary transcription late
ininfection,thenit mustbepresentin
enzyme-active nucleocapsids isolated from virions. In this report, we show that the largest virion polypeptideisindeedpresent andtightlybound to virion nucleocapsids active in transcription
and that thenucleocapsid structureunit isthe only other polypeptide resolved in these struc-tures. Inhibitoryeffects of virion envelope pro-teins onthe nucleocapsid transcriptase and its
stimulation by yeast RNA (11) are also de-scribed.
MATERIALS AND METHODS
Virus. A Sendai virus clone free of incomplete virionswasgrown in chickenembryolungcell cultures
at 30C (13). Radioisotopically labeled virions were
preparedasdescribed (12), except labeled precursors were added 16 h after infection and virions were
collected 32 h later. Culture medium containing released viruswascentrifugedfor10minat3,600 x g
to removecells and debris. Viruswaspelleted(78,000
x g at 5C for 30 min) and suspended in 0.01 M Tris-hydrochloride, 0.03 M NaCl (pH 8.0) for the standardpolymeraseassay (12) orin 0.01 M sodium phosphate (pH7.2) for fractionation experiments.
Virusfractionation. The high salt-detergent
treat-ment of Scheid et al. (9, 10) was used with the followingmodifications.Unlabeledorlabeled virions (0.1to0.2mgofprotein)werefractionated in 25 ml of 2%Triton X-100and1M KCl.After 20 minat25C, the mixturewasclarifiedbycentrifugation at 5 C for 20 min at 10,000 x g, which removed clumped, incompletely disrupted virions (5% of total protein). Thesupernatantwas centrifuged at 78,000 x g for 4 h
at 5C, and the pellet was resuspended in 0.01 M Tris-hydrochloride, 0.03 M NaCl (pH 8.0). The mate-rial in the 78,000 x g supernatant was further separated into glycopolypeptides and the non-glycosylated envelope polypeptide by dialysis and centrifugation (9, 10).
Isopycniccentrifugationofnucleocapsids. About
2 mg ofunlabeled virions, or 0.1 to 0.2 mg of labeled virions, disrupted as described above but in 2.0 ml of Triton X-100 andKCl,were centrifuged at 10,000 x g for20minat5C. Thesupernatantwaslayered onto a
sucrosegradient column prepared asfollows: 2 ml of
107
on November 10, 2019 by guest
http://jvi.asm.org/
MARX, PORTNER, AND KINGSBURY
±.36 gm/cm3 of D,O-sucrose was placed in a 1- by 3.5-inch centrifuge tube; a 27-ml linearD,O-sucrose gradient (1.15 to 1.33gm/cm3) was formed above it; and a top layer of 2% Triton X-100 and 1 M KCl in
D,O was then added. This preparation was cen-trifuged at 26,000 rpm for 16 h at 12 C in a Spinco SW 27 rotor. Fractions werecollected, anddensitieswere determined(12).
RNAtranscriptase assay. Reactions were run as before with
[3H]guanosine
5'-triphosphate asthe la-beled precursor (13), except Triton X-100 was substi-tuted for Triton N-101 when nucleocapsids were tested. Protein determinations were doneaccording to Lowry etal. (6).Acrylamide gel electrophoresis and radioactiv-ity measurements. These methods have been de-scribed in previous reports (12, 13).
Materials.Rabbithemoglobinwasprepared in our laboratory from a lysate of reticulocytes. Bovine serum albumin (BSA) was purchased from Sigma Chemical Co.Fetuinwasobtained fromGrandIsland BiologicalCompany,and ovalbuminwasfrom Worth-ington BiochemicalCorp.
RESULTS
Isolation ofenzyme-containing structures
from virions. Enzyme-active structures were isolated by centrifugation of Sendai virions disrupted in 1 M KCl containing 2% Triton
X-100.Enzyme activityof different virion prep-arations and 78,000 x g pellets derived from
them varied, but the latter were consistently
fourfold more activethan unfractionated virions
(Table 1). Incorporation by unfractionated
vi-rionsand the 78,000 x gpelletwaslinear for 5 h
(data notshown). About 46% ofvirion protein
wasrecovered inthe 78,000 x g
pellet,
sothere [image:2.495.262.452.387.593.2]wasaboutaninefoldincrease inspecific activity ofthe enzyme (Table 1).Thesupernatantfrom
TABLE 1. RNApolymeraseactivityinunfractionated virionsand the 78,000 xgpellet from high
salt-detergent treatedvirionsa
Experi-ment Counts! Counts per
ri Sample min min per mg
ofprotein
1 Virions 1,700 45,000
78,000 x gpellet 7,160 400,000
2 Virions 635 28,000
78,000 x gpellet 2,506 245,000
aReactions (13) of 0.1 ml of final volume were
incubated for 5h at 24C.Acomplete sampleofeach wasincubated at 4C for5handcountsper minute fromthesereactions, whichrangedfrom 35to60,were
subtracted fromexperimentalvalues.
b Each experimentwasdoniewithadifferent prepa-ration of virus.
the 78,000 x g centrifugation contained no
polymerase activity (datanotshown).
Virions disrupted by detergent-high ionic
strength buffer were banded isopycnically in
D,O-sucrose gradients to characterize the
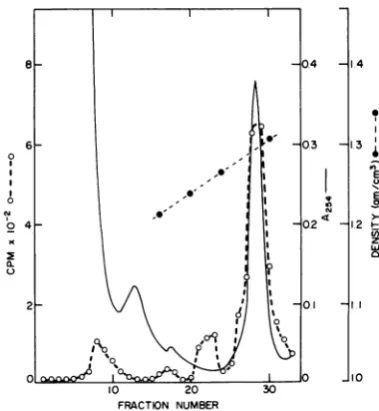
rap-idly sedimenting material with transcriptase activity. A peak of RNA polymerase activity was found at 1.29 gm/cm3 (Fig. 1), where
nucleocapsids band (5, 8), confirming
Robin-son's (8) observations. Electron microscopy of
this material revealed nucleocapsids (not
shown).
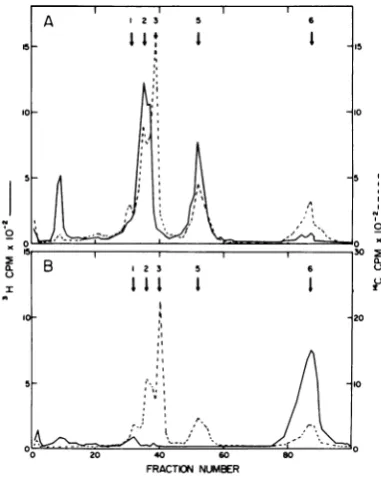
Polypeptides in the enzyme-containing structure. Enzyme-containing structures were analyzed by acrylamide gel electrophoresis to determine which polypeptides were present. In the following data, we number virion polypep-tides according to Mountcastle et al. (7); as before (12) we did not observe polypeptide 4, which is not marked in our figures. Acrylamide
gelelectrophoresisofthe1.29-gm/cm3structure
revealed three polypeptides (Fig. 2A), the larg-est virionpolypeptide (peak 1), the nucleocap-sid structure unit (peak 3), and relatively small amounts of one ofthepolypeptides in the region ofthesmaller virion glycopolypeptide (peak 5).
Toprevent adventitious bindingofirrelevant
polypeptides,virions weredisruptedina10-fold
larger volume ofbuffer (25ml), which resulted
0 0 0 l l l
a.3
L)
14
13
E C
12
z
0
1.1
ID
FRACTON NUMBER
FIG. 1. Isolationoftranscriptase-containing
struc-turefrom Sendai virions. Virionsdisruptedin deter-gent-high ionicstrength bufferwerecentrifugedina D,0-sucrosegradient as described in Materials and Methods. Radioactivity represents [3H]guanosine 5'-monophosphate incorporated in a5-h transcriptase
assay.
108 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.495.56.254.479.613.2]SENDAIVIRONTRANSCRIPTASECOMPLEX
25Z^tiU-tl
Co~~ ~~~~~~~.J
0.~~~~~~~~~~~~~~~~~~~~~~~0
' B 23 5 6 1)
I- III 1 l
15.2
6- 06i
0 20 40 so 8
[image:3.495.50.241.52.318.2]FRCTIONNLNBER
FIG. 2. Polyacrylamide gel electrophoresis of 3H-polypeptides in transcriptase-active structures. 14C-polypeptides from Sendai virionswereadded before electrophoresis. The numbering system follows Mountcastleetal.(7).Migrationisfromlefttoright. A, 3H-polypeptides from the 1.29-gm/cm8 structure; B, 3H-polypeptides from the 78,000 x gpellet.
in the 78,000 x g pellet containing only two
polypeptides, thelargestvirionpolypeptideand
thenucleocapsid structure unit(Fig. 2B). Tran-scriptase activity was recovered in full from unlabeled virionsdisruptedatthe same concen-tration. Thus, only two virion polypeptides
appear to be involved intranscriptase function. Inhibition of enzyme activity by viral enve-lope proteins. As shown above, enzyme-active
nucleocapsids contained about half of the total
protein of virions, but they had a ninefold greater specific activity than unfractionated virions. Thissuggests that virionenvelope com-ponents, perhaps the polypeptides, inhibit
po-lymeraseactivity. Before testingenvelope
com-ponents for inhibition of polymerase activity, the 78,000 x g supematant was dialyzed to remove KCland renderpolypeptide 6 insoluble (10). After centrifugation, the supematant con-tained mainly the two virion glycopolypeptides (Fig. 3A), and the precipitate contained mainly polypeptide 6 (Fig. 3B). The peak of radioactiv-ity at fraction 8 in Fig. 3A appears to be an aggregate of virion polypeptides formed during isolation ofthe envelope fraction, because it is
notusuallypresent inunfractionatedvirions (7,
12) (cf. Fig. 2and Fig. 3B).
Results in Fig. 4 show that either fraction
markedly decreased enzyme activity.
Polypep-tide 6 inhibited about 80% of polymerase activ-ity when 40
jug
was added to a reaction contain-ing 40gg
of nucleocapsids. At the same concen-tration, glycopolypeptides almost totally abol-ished enzyme activity. Several experiments summarized inTable2weredone to investigate whether inhibition by viral envelope proteins was a virus-specific phenomenon. Brief heatdenaturation of envelope proteins completely
destroyed their ability to inhibit polymerase
activity,indicating that protein conformation is important for inhibition. When labeled 50S
virion RNAwas added to a complete reaction containing 400 ug ofenvelope proteins per ml and the mixture was incubated at 24 C for 16 h, noradioactivitybecame acid soluble. This indi-cates that inhibition of transcriptase by enve-lope proteins is not due to a ribonuclease and confirms earlier evidence against a nuclease activity inSendaivirions (11).
Envelope proteins were isolated from
New-castle
N
disease virions (NDV) byusing the same0.
a.)tL I
0
I!
0.
FRACTONNUMBER
FIG. 3. Polyacrylamide gel electrophoresis of 3H-polypeptides from the 78,000 xg supernatant. 14C-polypeptides from virionswereaddedbefore electro-phoresis. A, 8H-polypeptides which remain soluble after dialysis of the 78,000 x g supematant, B, insoluble3H-polypeptidesafterdialysis.
109 VOL.13,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.255.446.357.596.2]MARX, PORTNER, AND KINGSBURY
100
-J
0
6
LL
0
e-a 4t
0 z
a.
0-50
10
5
16 10 20 30 40
PROTEIN ADDED (pg)
FIG. 4. Inhibition of transcriptase innucleocapsids by envelope polypeptides. Radioactivity represents
[image:4.495.82.226.61.244.2][3Hjguanosine 5'-monophosphate incorporated in a 5-h, 0.1-ml transcriptase assay. Counts/min in the control assay were 7,510. Symbols: 0, polypeptide6; 0,glycopolypeptides2and 5.
TABLE 2. Inhibition ofnucleocapsid-associated polymeraseactivitya
Additios.Controlcounts/min"
Additions (%)
Sendai virusglycoproteins ... 4
Sendai viruspolypeptide6 ... 23
Heat-inactivatedSendai virus glycoproteinc ... ... 100
Heat-inactivatedSendai virus polypeptide6 ... 100
NDVglycoproteins ... 30
NDVpolypeptide6... 59
Ovalbumin ... 100
Fetuin... 100
Hemoglobin ... 100
BSA ... 100
aReaction mixtures of 0.1 ml of final volume
containing40 Agofnucleocapsidplus40Agofadded protein were incubated at 24C for 5 h. Counts per minute fromreactionsincubated at5C for5 hwere
subtracted fromexperimentalvalues.
bCounts/minfrom reactions withnoadditionswere 5,775.
cVirionproteins were heatedat 100 Cfor 2min.
method as for Sendai virions. They inhibited the polymerase in Sendai viral nucleocapsids,
but less efficiently than Sendai viral envelope
components. Because any undenatured protein might becapableofinhibitingenzymeactivity, several nonviral proteins were tested: 40 sgof either BSA, hemoglobin, or the glycoproteins
ovalbumin or fetuin was added to
nucleocap-sids. None of these proteins inhibited enzyme activity (Table 2).
Stimulation of enzyme activity by yeast
RNA. YeastRNA andother polyanions
stimu-late Sendai virion transcriptase activity (11), buttheirmechanism ofaction is not clear. The
results presented abovesuggested that
polyan-ions mightact byreversinginhibitoryeffectsof
envelope proteins. If this were true, then yeast RNA should stimulate nucleocapsid-associated
enzyme only in the presence of envelope
pro-teins. Resultssummarized in Table 3 show that yeastRNAstimulates nucleocapsidseven in the
absenceofenvelope proteins.
Intheseexperiments, it was noted that some
highly active nucleocapsid preparations could notbe stimulatedatallbyyeast RNA,although
lessactive preparationsalwayswerestimulated.
This indicated that yeast RNA might be
reac-tivating heat-denatured transcriptase. There-fore,apreparation ofhighactivity waspartially
inactivated by mild heat treatment and tested
forstimulation. DatainTable3showthatyeast
RNA restored mostofthe activity destroyed by
heat treatment.
TABLE 3. Stimulation of virion or
nucleocapsid-associated transcriptase by yeast RNAa
Enhance
-Enzyme Additions Counts/min mentover
control
Virions None 1,220
30 Mg of yeast 1,936 1.6 RNAper ml
300 gg of yeast 2,397 2.0 RNAperml
Nucleocapsids None 5,020
30 gg of yeast 5,937 1.2 RNAperml
300 gg of yeast 9,179 1.8 RNAperml
Heated nucle- None 1,384 ocapsids'
30 ug of yeast 2,521 1.8 RNA per ml
300 ug of yeast 7,591 5.5 RNAperml
aReactionmixtures of 0.1mlof final volumecontaining26
ugofnucleocapsid proteinor39 gg of virionproteinand the indicated additionswereincubatedat 24Cfor 5h.Areaction mixture with nucleocapsids, which was incubated at 4C,
contained 76 counts/min and was subtracted from each
experimentalvalue. Background subtractionfor virion-con-tainingmixturewas84counts/min.
bNucleocapsidswereincubatedat30Cfor 12 min in 0.01 MTris-hydrochloride,0.03MNaCl(pH8.0).Theywerethen brought to 24 C, mixed withthe remaining ingredients of the transcriptase assay (13) and yeast RNA as indicated
above,and incubatedat24Cfor 5h.
I
0
\I
I
I I*,
I
110 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.57.250.336.486.2] [image:4.495.258.456.352.559.2]DISCUSSION
Nucleocapsids isolated from Sendai virions containanRNA polymerase that is significantly moreactive thanreportedpreviouslyfor
unfrac-tionated virions (13). Removal of the virion envelope accounts for the increaseintwoways:
first, half of the viral proteinsareremoved, and
second, the RNA polymerase is freed from inhibition by envelope proteins. Inhibition by envelope proteins appeared to be specific,
re-quiringundenatured viral proteins.
The enzyme-containingnucleocapsids
resem-ble transcriptive complexes isolated from in-fected cells (12) in buoyant density,
morphol-ogy, and polypeptide composition. Thus,
simi-larstructuresand perhaps thesameenzyme are
responsible for transcription in virions and in infected cells. More detailsareneededto deter-mine whether virion polypeptide 1 is indeed identicalto"polypeptide1"detectedincellular transcriptive complexes (12).
The specificity and mechanismof the
inhibi-tion of transcriptase by envelope polypeptides
maybe revealed by further work. Although only
viral envelope proteins were inhibitory among
those tested, effects of envelope proteins from other viruses as well as additional nonviral
proteins need to be examined to learn if the phenomenon is limited to paramyxovirus com-ponents. Among possible mechanisms of inhibi-tion, the presence ofa proteaseintheenvelope
fraction must be considered. However, when
various amountsof BSA (uptoa 10-foldexcess
ofenvelope components) were added either to
glycopolypeptideorpolypeptide 6fractions, the
inhibition of transcriptase activity was
unaf-fected (data not shown), ruling out gross
con-tamination by a protease.
If the cellular transcriptive complexes are
destined for maturation into virions, our data
on transcriptase inhibition by envelope
poly-peptides suggest a mechanism forterminating
transcription intheenvelopmentprocess.
How-ever, if this were true, it is surprising that the virion glycopolypeptides, which are presumed to reside on the outside of the envelope (2),
inhibit transcriptase activity. Interaction
be-tween nucleocapsid-bound transcriptase and polypeptide 6, which is thought to be on the
inside of the envelope (2), seems more likely.
Although polypeptide6inhibitedless efficiently thantheglycopolypeptides,thismayreflect the poorsolubilityofpolypeptide6 atthe low ionic
strengthused in the transcriptase assay (13).
Other speculations about enzyme inhibition
by envelopepolypeptides might be entertained. Forexample, envelope glycopolypeptides might
regulate RNAsynthesis at a point in replication before virus maturation.
Recovery of only two virion polypeptides in the enzyme-containing nucleocapsids clearly rules out both glycopolypeptides and polypep-tide 6 as part of the transcriptase. Nucleocap-sids isolated from virions contained less poly-peptide 1 than virions or transcriptive com-plexes from infected cells. This could result from removal of part of polypeptide 1 by salt-detergent treatment, or it is possible that two virionpolypeptides comigrate at this position in the gel (7), one of which is removed during fractionation.
Either the largest virion polypeptide or the nucleocapsid structure unit remains a candi-date for the enzyme molecule. Results with vesicular stomatitis virus showed that
disrup-tion of virions with high salt and detergent released transcriptase-containing cores that contained the largest virion polypeptide L, a
minor component, NS 1, and the major
struc-tural polypeptide N (1, 14). Emerson and
Wag-ner (4) showed that L and NS 1, as well as
transcriptase activity, could be removed from the core structure bytreatment with detergent
at sufficiently high ionic strength. When
poly-peptidesL andNS 1wereaddedback tocores,
transcriptaseactivitywasrestored, implicating
either or both of these polypeptides in
tran-scriptase function. Thelargest virion
polypep-tide of Sendai virions isevidently moretightly
associated with the nucleocapsid than the L
polypeptide of vesicular stomatitis virus,
be-cause we have not been able to dislodge it at
very high ionic strength (P. Marx and D. W. Kingsbury, unpublished data).
The situation with influenza virusseemsless complex. Compans and Caliguiri (3) reported
transcriptase-active nucleocapsid structures isolated from influenzavirus-infected cells
con-tained only one polypeptide, the nucleocapsid
structure unit.
Stimulation ofnucleocapsid-associated
tran-scriptaseprovides further evidence that
polyan-ions act directly on the enzyme. The results suggested that polyanions stimulate by
revers-ing thermal inactivation of the transcriptase.
ACKNOWLEDGMENTS
Andrew W. Moseley and Ruth Ann Scroggs provided experttechnical assistance.
This research was supported by Public Health Service research grantAI-05343 from the National InstituteofAllergy and InfectiousDiseases,bytraining grantTO1-CA-05176 and Childhood Cancer Center research grant CA-08480fromthe NationalCancer Institute, and by ALSAC. D. W. K. received Career Development Award HD-14,491 from the National Institute ofChild Healthand HumanDevelopment.
13,1974
on November 10, 2019 by guest
http://jvi.asm.org/
MARX, PORTNER, AND KINGSBURY
LITERATURE CITED
1. Bishop, D. H. L., and P. Roy. 1972. Dissociation of vesicular stomatitis virus and relation of the virion proteinstothe viral transcriptase. J. Virol. 10:234-243. 2. Chen, C., R. W. Compans, and P. W. Choppin. 1971. Parainfluenza virus surface projections: glycoproteins with haemagglutinin and neuraminidase activities. J. Gen. Virol. 11:53-58.
3. Compans, R.W., and L. A. Caliguiri.1973. Isolationand properties ofanRNA polymerase from influenza
virus-infected cells. J. Virol.11:441-448.
4. Emerson, S. U., and R. R. Wagner. 1972. Dissociation and reconstitution of the transcriptase andtemplate activities ofvesicularstomatitis virusBand T virions. J. Virol. 10:297-309.
5. Hosaka, Y.1968.Isolation andstructureofthe nucleocap-sidofHVJ.Virology 35:445-457.
6. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, andR. J.
Randall. 1951. Protein measurement with the Folin phenolreagent.J.Biol. Chem. 193:265-275. 7. Mountcastle, W.E., R. W. Compans, and P. W. Choppin.
1971.Proteinsand glycoproteinsofparamyxoviruses:a
comparisonof simianvirus5,Newcastle diseasevirus, andSendai virus. J. Virol.7:47-52.
8. Robinson, W. S. 1971. Ribonucleic acid polymerase activityin Sendaivirions andnucleocapsid. J.Virol. 8:81-86.
9. Scheid, A., L. A. Caliguiri, R. W. Compans, and P. W. Choppin. 1972. Isolation of paramyxovirus glyco-proteins. Association of both hemagglutinating and neuraminidase activities with the larger SV5 glycopro-tein.Virology 50:640-652.
10. Scheid, A., and P. W. Choppin. 1973. Isolation and purification of the envelope proteins of Newcastle disease virus. J. Virol. 11:263-271.
11. Stone, H.O., and D. W. Kingsbury.1973.Stimulation of Sendai virion transcriptase by polyanions. J. Virol. 11:243-249.
12. Stone, H. O., D. W. Kingsbury, and R. W. Darlington. 1972.Sendai virus-induced transcriptase frominfected cells: polypeptides in the transcriptive complex. J. Virol. 10:1037-1043.
13. Stone, H. O., A. Portner, andD. W. Kingsbury. 1971. Ribonucleic acid transcriptases in Sendaivirionsand infected cells. J. Virol. 8:174-180.
14. Szilagyi, J. F., and L. Uryvayev. 1973. Isolation ofan
infectious ribonucleoprotein from vesicular stomatitis viruscontaininganactive RNAtranscriptase.J.Virol. 11:279-286.
112 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/