Genome of Sindbis Virus
B. M. ARIF AND P. FAULKNER
Department ofMicrobiologyandImmunology, Queen'sUniversity, Kingston, Onltario, Canada
Received for publication 13 September 1971
32P-labeled ribonucleic acid (RNA) from purified Sindbis virus was examined for thepresenceofhidden breaks. Viral RNA was treated with acid at pH 2.9 or withformaldehyde andwasanalyzedonsucrose gradients or bypolyacrylamidegel electrophoresis. Thesedimentationpattern and mobility onpolyacrylamidegels of the 42SRNA was unaffected by heatingand quickcoolingand was not altered by denaturing agents such as dimethyl sulfoxide and urea. No evidence that Sindbis RNAis apolyaggregateoffragments was obtained. It is concluded that the genome consists ofacontinuouslength of single-stranded polynucleotide.
Several group A arbovirusesare consideredto contain a genome which consists of a single-stranded polynucleotide. In the case of Sindbis virus, it exists as42S infectious ribonucleic acid (RNA) which may be extracted either from purified virionsorfrom the140Snucleoidpresent in infected cells (6). The ribonuclease-sensitive
infectious
RNAof
these viruses appears homo-geneous on sucrosegradients (7, 15)
andmoves as a single fraction onpolyacrylamide
gels (3, 7). RNA from the virion is converted to a species having alowered sedimentation coefficient upon heating followed by rapid cooling (7, 15). This change in sedimentation coefficient is accom-panied by fragmentation and increased mobility ongels (3, 7). Both Sindbisand Westernequineencephalomyelitis (WEE)
40SRNAarebelieved to possess a hydrogen-bonded secondary struc-ture sincethey undergo hypochromic shift upon heating and appear to be almost completely reannealed upon slow cooling (14, 15). Treat-mentof 40SWEERNAwithdimethyl sulfoxide(DMSO)
orwith ureadidnotresultin achange inthe
sedimentation pattern insucrosegradient
after removal of the reagents
(15).
However, virion RNA fromWEEandSindbis
appeared
to cosedimentwith 28Scellribosomal RNA(rRNA) orwithviral-specific
26S RNAwhencentrifuged
insucrose gradients preparedinDMSO (5, 15). Sindbis RNA treated with 12 M urea showed a
similarelectrophoretic mobilityto28SrRNAon gelscontaining8M urea(8).Thesechangesinthe biophysical properties of group A arbovirus RNAinduced bytreatmentswhichareknown to break hydrogen bonds have led to
speculations
aboutthe structure of the viral genome (3, 5, 7, 15).It wassuggestedthat viral RNA may exist in twoconformationswhich sedimentdifferentlyon
sucrose
gradients (42
and 26Sspecies)
andwhichmay migrate differently on gels; or the genome may beconsidered as apolyaggregate oftwo or more roughly equal-size fragments which sedi-ment at the same rate and migrate at roughly similarmobilityongels.
Inthis communicationweshowthatifsufficient precautions are taken to extract RNA free of hidden breaks, no fragmentation ofthe genome is observed after heating or treatment with denaturing agents and that Sindbis virus RNA behavesas anunbrokenpolynucleotide.
MATERIALS AND METHODS
Chemicals. Acrylamide,
N,N'-methylenebisacryl-amide and N,N,
N',
N'-tetramethylenebisacrylamidewere purchased from Eastman Organic Chemicals.
Acrylamide and N',N'-methylenebisacrylamide were
recrystallized from chloroform and acetone,
respec-tively. Carrier-free 32po4 was obtained from the
Atomic Energy of Canada Ltd. DMSO and urea wereboughtfromFisherScientificCo.
Tissue culture andvirus growth.Theprocedurefor thepreparationofBHK/21 cell monolayersonFalcon
plastic petri dishes and the growth ofSindbis virus
havebeendescribed (1).
Preparation of 32P-labeled cells and extraction of
RNA.Cellmonolayers ofBHK/21cellswerewashed
twice with phosphate-free minimum essential
me-dium (MEM, Grand Island Biological Co.) and
were overlaid with medium A: MEM containing 2.5% BHK/21 medium (Grand Island Biological Co.), 1.5%dialyzedcalf serum, and 10,ACi Of
32PO4
perml. Cell monolayerswereincubated at 37 Cfor 18 hr.RNA wasextractedaspreviouslydescribed(6).
Preparation of 32P-labeled viral RNA. BHK/21 cells were prelabeled with 32po4 as described above and wereinfectedwith Sindbis virusatamultiplicity of 40:1. After adsorption, they were overlaid with medium A, and the
32P-labeled
virus was harvested 18 hr later. The medium was clarified at 15,000 rev/min for 15 min at 4 C (Sorvall SS34), and thevirus was pelleted from the supernatant fraction at
102
on November 11, 2019 by guest
http://jvi.asm.org/
GENOME OF SINDBIS VIRUS
25,000 rev/min for 1 hr in a Spinco SW25 rotor at
4C. Theviruspellet wassuspended in boratesaline (BS; 50mmborate and 120 mMNaCl,pH9)
contain-ing 0.4% bovine serum albumin (BS/BSA) and was
clarified at 10,000 rev/min for 10 min in a Spinco
SW50rotor at 4C. Concentrated virus wasfurther
purified by centrifugation on a preformed 5to 50%
potassium tartrate gradient in BS/BSA (11). The virus was detected by radioactivity assay and by a
hemagglutination test with goose erythrocytes (4). Fractions containing the virus were pooled, diluted with BS, and pelleted by centrifugation at 25,000 rev/min for 2 hr in a Spinco SW25 rotor at 4 C. Theviruspelletwasdigested with4%sodiumdodecyl
sulfate (SDS) and 1% 2-mercaptoethanol in phos-phate-buffered saline A (140 mmNaCl, 27 mM KCI,
CPM
A
8mM Na2HPO4, 1.5 mM KH2PO4, pH 7.2) at 37 C
for1hr. Thedigested material was then centrifuged
on a10to20%sucrosegradient(38,000 rev/minfor 3
hrin anSW50rotorat21C) prepared in
electrophore-sis buffer [EPB;4 M
tris(hydroxymethyl)aminometh-ane (Tris), 2mmsodium acetate, 1 mn ethylenedia-minetetraacetic acid (EDTA), 0.1% SDS, pH 7.2]. The viral RNA sedimenting at 42S (calculated with
respecttocell RNA markersincludedinthegradient)
was precipitated in thecoldwith 2.5volumes of
ab-soluteethanoland storedat -20C.
Acid treatment. 32P-labeled viralRNAwasexposed
to 0.2 ml of 5 mm hydrochloric acid to lower the pH to2.9for 30sec (10), followed by0.15 mlof 10
mMEDTAcontaining0.2%SDS. Thefinal pH of the
mixtule was5.6. Thematerial wasthenlayered onto
B
x
io
3CPM 28S1SS4S
Xio-2
1
l1
C
O D
260
4 8 12 16 20 4 8 12 16 20
Fraction Number
FIG. 1. Test for hidden breaks by the acidtreatment.32P-labeled viral RNA (0) wascentrifugedon10to20%
sucrosegradients preparedin EPB(see text)at45,000 rev/min for2.25hr inanS W50rotor at21C. Mammalian
cellRNA markers (0) were includedin thegradients. A, Viral RNAwasdissolved in EPB and centrifugedon
on asucrosegradient. B, Viral RNAwastreated with 0.2 mlof5mm HCIat21Cfor30sec (pH 2.9), followed by0.15 mlof10mMEDTAcontaining0.2%SDS(final pHwas5.6).Itwasthencentrifugedon asucrosegradient. C, Viral RNA wastreated with2jugofribonucleaseperml inEPBat0CforSmin. SDS (0.1%) and cellRNA
markerswereadded andanalyzedonasucrosegradient. D,Ribonuclease-treated viral RNA wasexposedtoacid
asdescribedfor panel B, followed by analysison asucrosegradient. 2
VOL. 9, 1972
on November 11, 2019 by guest
http://jvi.asm.org/
[image:2.498.106.395.231.581.2]a 10 to 20% sucrose gradient prepared in EPB and centrifuged at 38,000 rev/min for 3 hr in a Spinco SW50 rotor at21C.Eachfraction (0.2 ml) was added to 10 ml ofp-dioxane-based scintillator and counted in a liquidscintillation spectrometer.
Formaldehyde treatment. "P-labeled viral RNA was treated with 1% formaldehyde in EPB, and then
centrifuged on a 10 to
20%10
sucrosegradientcontain-ing 1% formaldehyde at 21 C. The fractions were countedasdescribed for theacidtreatment.
Treatment of RNA with DMSO. 32P-labeledRNA,
in 10Mliters of EPB, was incubated with 0.1 ml of DMSO atroomtemperature for15min. TheDMSO was removed by dialysis against 500 ml of EPB at room temperature, and the RNA was examined by
polyacrylamide gel electrophoresis.
Thetechniqueforsedimentationof RNA in sucrose
gradients preparedin DMSO-1 mm EDTA has been described (5).
Treatmentof RNA withurea. 32P-labeled RNA was
incubated with 10 M urea prepared in saline A at 37 C for 30 min. The RNA was analyzed by gel electrophoresis.
Polyacrylamide gel electrophoresis. Composite gels (6cm in length) of 2.5% acrylamideand 0.5%
agarose were prepared in EPB by the method of Peacock andDingman(13). Thegelswereprerun by
electrophoresis in EPB at 50v for 30 min at 4C
before the RNA sample was added. After RNA
analysis thegelsweresliced transversely on a plastic fractionating device.Eachslice wasdriedon aplanchet
andcountedinagas flow counter. RESULTS
Absenceof hidden breaks in thepreparation.To investigate the effect ofdenaturing reagents such as DMSO and urea on viralRNA, it was
neces-sary toshow thatalltheRNApreparationswere free of hidden breaks which may have been in-duced during the extraction procedures. Treat-ment of viral preparations with acid at pH 2.9 (10) or withformaldehyde (2, 9) followed by re-examination of the RNA preparation on
gradi-entsmay beusedtoindicate the absence ofhidden breaks. These criteria were
applied
to Sindbis viral RNA(Fig. 1 and 3).TheprofileinFig. IA showsthat Sindbis RNA,prepared
asdescribed
above,sedimentsatabout42Swhencomparedto
hamster cell RNA.InFig. lB it isseenthatviral RNA continues to sediment as a single
species
after theacidtreatment.
Viral RNAwasthennicked
by
alimiteddiges-tion with pancreatic ribonuclease
(2
jg/ml
for 5min at 0C). Theenzyme actionwas
stopped
by
theadditionofSDSandcellRNAmarkers. The samplewasdividedintotwoparts. One partwas
treated with acid as described
above,
and the otherpartwasleftuntreated. The data inFig.
1C show that mild ribonuclease treatment did notalter the sedimentation coefficient of the viral RNA. However, a heterogeneous
profile
wasobservedwhen the ribonuclease-treated material was exposed to acid (Fig. 1D) which indicated that noneoftheoriginal42Sviral RNAremained intact.
Further evidence of the integrity of the viral RNAwasobtainedwhen the mobility of prepara-tions onacrylamidegels was examined after acid treatment at pH 2.9. Theprofiles in Fig. 2 show that themobilities of28S rRNA and Sindbis 42S RNAwereunalteredby thetreatment.There is no evidenceof increased heterogeneitywhich would have resultedin the productionoffaster-moving (smaller)RNAfragments.
A similar series ofresults was obtained when the viral RNA was reacted with 1 % formalde-hyde (Fig. 3). The 42S viral RNA (Fig. 3A) re-tained its sedimentation characteristicafter treat-mentwith formaldehyde. In this experiment the effect of ribonuclease was more severe than that seenin Fig. 1B, and even before treatmentwith formaldehyde itwas possible todemonstratethe heterogeneity of the viral RNA (Fig. 3C). When the ribonuclease-treated viral RNA was reacted withformaldehydeandre-examinedon asucrose gradient containing 1 %formaldehyde (Fig.
3D),
themajor species of RNA sedimented in therange 18 to 25S.
Effectof heating
viral
RNA."2P-labeled
Sindbis2- A
CPM X10-3
4 8 12 16 20 24 28 32 36 40 44
Fraction Number
FIG. 2.Polyacrylamidegel electrophoresis of
acid-treatedRNA. 32P-labeled viral RNA was treatedwith acidasdescribedinlegendto Fig. I andwasanalyzed byelectrophoresisat4 Con2.5 to0.5%
acrylamide-agarose composite gels for 50minat125v. Symbols:
(0),
acid-treated viral RNA;(O),
acid-treated 28SrRNA; (-), viral RNA control;
U,
28S rRNAcon-trol.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.498.266.455.376.585.2]GENOME OF SINDBIS VIRUS
CPM -3 X 10
2
1
O
D260
FRACTION NUMBER
FIG. 3. Treatmenit ofviral RNA withformaldehyde.32P-labeled viralRNA (a) wascentrifugedoni 10to20%o
sucrosegradientspreparedinEPB(see text)at45,000 rev/minfor2.25hr in anSW50rotor at21C.
Mammalian-cell RNA markers (0) wereincludedinthegradients. A, Viral RNAwasdissolvedin EPBandcentrifugedona
sucrose gradient. B, Viral RNA was dissolvedin EPBconitaininlg
1%Cc
formaldehyde andanalzyedon agradient colntaining 1%formaldehyde. C, Viral RNA wastreatedwithl 2 sg of ribonuclease per mlinEPB at0 Cfor5mitt. SDS (0.1%0) antdcell RNA markers were addedanidthe mixture was anialyzedon a sucrosegradielnt. D,
Ribonuclease-treated viral RNA wasmade up to 1% with respect toformaldehyde anidanalyzedoniagradient conttaining1%Oformaldehyde.
RNA was prepared and dissolved in buffers of either low molarity (EPB) or high molarity (standard buffer, STB; 500mm Tris, 100mm NaCl, 1 mm EDTA, pH 7.2). The solution was held at 85 Cfor 3 minin a waterbath and then quickly cooled in melting ice. Analysis of the RNA on polyacrylamide gels (Fig. 4) showed that therewas no lossof the principal42S RNA componentwhenbreak-freeRNAwasused in the experiment. Previous studies with Sindbis and Semliki Forest RNAtreated in this manner (3, 7) indicated that fragmentation of the genome had occurred,and, in the case ofSemlikiForestvirus RNA, it was thought that several distinct sub-units were separated (3). However, it must be concludedfrom the data inFig.4that the genome doesnotcleave uponheating.
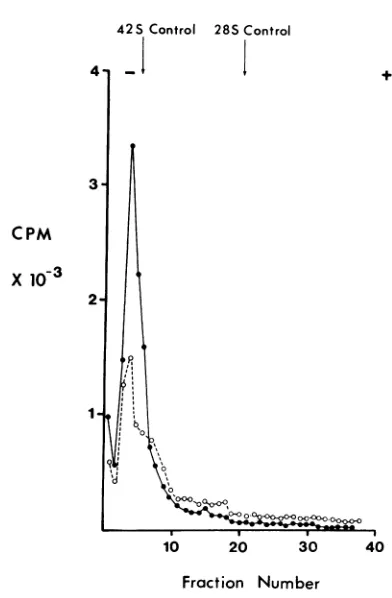
Effects of DMSO and urea on viral RNA. Sindbis RNA was obtained from 32P-labeled
virionsbydigestion with SDS andwasseparated on a sucrose
gradient
andprecipitated.
The RNAwas subsequently dissolved in EPB and divided into two portions. One wastreated with DMSO (finalconcentration, 91 %) andthe other wasleft untreated. The 28S rRNA was similarlytreated, and the four samples were examined on 2.5 to 0.5%7c polyacrylamide-agarose gels. These gels containedaplug oflowporosityatthebottomto
trap anyfast-moving low-molecular-weight prod-uct. The profile in Fig. 5A indicates that the mobility of285rRNAis virtually unaltered upon
treatment with DMSO. The mobility ofSindbis RNA was fractionally increased upon DMSO
treatment as aresultof the denaturation process, but therewas noevidenceoffragmentation.
The routine method of extracting viral RNA was by digestion of the virions with SDS and subsequent separation of the viral RNA on a
105
VOL. 9, 1972
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.498.104.393.71.386.2]CPM
x
10-3
1-42S Control
I1
28SControl
1 20
Fraction Num FIG.4. Effect of heatingon viral RN viral RNA, dissolved in eitherEPB (a)
was heatedat 85 Cfor 3 miii andquic
melting ice. It was analyzed by eleci
4 C on 2.5 to 0.5% polyacrylamide A
for I hr.
sucrose gradient. When the RNA
thistechnique was further extracted
to remove traces ofprotein, similar
obtained upon DMSO treatment (F mobility of the DMSO-treated vira only fractionally increased by the
sedimentation pattern of arbovirL
sucrose gradients prepared in DMS
shownto be similar tothat of 28Sr
viral-specific 26S RNA (15). These
ternswerethoughttohave beendue
down ofapolyaggregate 42SRNA in
DMSOgradientortoaconformatioi
theviral 42S RNA. Anexperiment w;
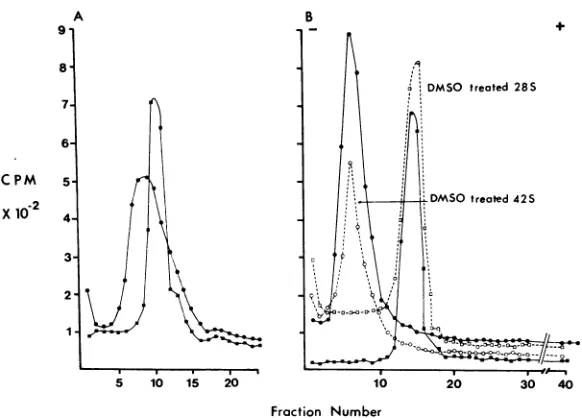
toseewhethercleavage occurswhen
RNA is fractionated on 99% DI
gradients. The results (Fig. 6A) con
sedimentationpattern of SindbisR1S tothat of28S rRNAasdescribed pr
RNAwastaken from the peak frac
gradient, precipitated with unlab
RNA, and applied to polyacrylami
data (Fig. 6B) show that the 42S RNA derived from the gradient prepared in 99%0 DMSO
co-+ migrated with untreated 42S RNA and was not
cleaved by the treatment.
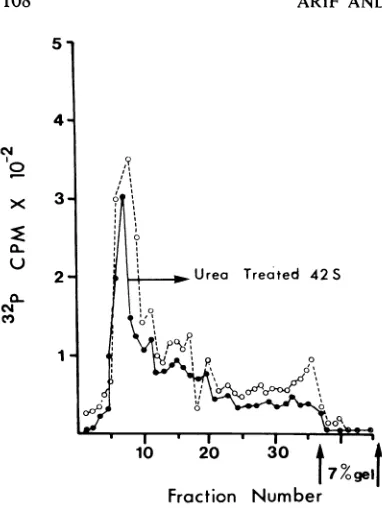
Anessentially similarresult wasobtained when the electrophoretic pattern of urea-treated viral RNA was compared with that of untreated 42S RNA. Inthis experiment, viral RNA was incu-bated with 10 M ureafor 30minat 37C.Carrier RNA was added, and the mixture was precipi-tated with ethanol. The RNA was dissolved in EPB and appliedto gels (Fig. 7). The profileof theuntreated RNAindicates some heterogeneity inthe preparation, butthe principal component was not eliminated by urea treatment as would have occurred if fragmentation hadbeen brought aboutby theurea treatment.
Absence of low-molecular-weight RNA in the virion. Amodel that has beenconsidered for the structure of arbovirus RNA is that the genome consists of two approximately equal-sized frag-mentsofRNAlinkedtogetherbycomplementary base pairing withashortpiece of oligonucleotide. We investigated this possibility by subjecting Sindbis RNA of
high specific
activity
togel
* * electrophoresis in a
10%
gel with cell 4S RNA 30 40 used as a marker (Fig. 8). As expected,viral RNAiber
did not penetrate the gel, whereas 4S RNAber moved about42%
along
itslength.
Treatment ofA
oP-labeled
Sindbis RNA with DMSO did not release a)lor
STB(0)
fragment that could be detected under thesetrophoresis
at conditions. We estimate thesensitivity
of ourgels at 100 v
technique
was such thatwewould have detectedan oligonucleotide 100 nucleotides inlength by thistechnique.
obtained by DISCUSSION
withphenol
results were Thefindings reported here indicate that Sindbis ig. 5B). The RNA which is demonstrably free of hidden d RNA was breaks does not change its sedimentation be-agent. The havior on sucrose gradients or its
mobility
on as RNA in acrylamidegelswhenheatedandquick-cooledor '0 has been when treated with agents which denature sec-rRNA (5) or ondary structure. Thereappeared
to be ample similar pat- evidence that the genome of the group A arbo-e toa break- viruswascleaved whenRNAextracted from the X the sucrose- virions washeated in aqueous neutral solutions. nalchange in This treatment was reported to result in a de-asperformed creaseinsedimentation coefficientof WEE RNA Sindbis 42S (15),accompanied by
an increasedmobility
on ISO-sucrose gels in the cases ofSemliki Forest RNA (3) and firm that the Sindbis RNA(7). Pronounced increasedmobilityIA
is similar ofheatedRNAwasascribedtothemigrationof reviously (5). fragments of the viral RNAformedby
chemical ,tions onthe cleavage orby meltingofhydrogen
bondswhich )eled carrier hold subunits of the genome in apolyaggregate [de gels. The structure (3, 7). Conformationalchanges
haveon November 11, 2019 by guest
http://jvi.asm.org/
[image:5.498.52.248.71.370.2]5-A
B+ -4
4-28S 28S
CPM DMSOtreated425
X 102 DMSO treated 28S
2-8 16 24 32 40.48 8 16 24 324048
o*Oo'0
gel gel
[image:6.498.113.401.71.256.2]Fraction Number
FIG. 5. EffectofDMSOonviral and cellular RNA.A, 32P-labeled viral and cellular 28S rRNAweredissolved in 10,lAitersof EPB(see text)followedby 0.1 mlofDMSOand incubatedatroomtemperaturefor JOmin.DMSO wasremovedbydialysis against EPB, and the RNAwasanalyzed byelectrophoresisat4 Con2.5 to0.5%
acryla-mide-agarose compositegelsat125vfor50min.Symbols: (0) DMSO-treated viral RNA; (A)DMSO-treated 28SrRNA; (0) untreatedviral RNAcontrol. Arrow indicates the positionofthe untreated 28S rRNAcontrol. B,32P-labeled viralRNA,preparedasoutlinedintext,wasextractedtwicewithredistilledwater-saturatedphenol.
Itwasprecipitated with 2.5 volumes of absolute ethanol and storedat -20 C. Aportion ofthe precipitate was
dissolvedin10,ulitersofEPB, treatedwithl DMSO, andanalyzed by gelelectrophoresisasdescribedabove. Symbols
are asfor A.
A B
9 4.
8
DMSO treated 28S
7-
6-CPM 5
-2
~~~~~~~~~~~~~~DMSO
treated42S3-2
-5 10 15 20 10 20 30 40
Fraction Number
FIG. 6. A, Anialysisof viral RNA and 28S rRNA on 99% DMSO gradients. 32P-labeledviral RNAand28S
rRNA weredissolvedin 10,ulitersofEPB(see text),followed by0.1 mlof DMSO. It was centrifuged at22C
on apreformed5 to20%sucrose gradient preparedin99% DMSO andIm.i EDTAat 55,000rev/minfor19
hrin an SW56rotor.TheviralRNA (0) infractions 8, 9, and 10 and the 28S rRNA (D) in fractions 10 and 11
wereprecipitated with2.5volumes ofabsolute ethanoland anexcessofcarrieryeastRNA. B, The RNA
precipi-tatesfrom theDMSO gradient were dissolvedinEPB andanalyzed at 4Con 2.5 to 0.5% acrylamide-agarose
compositegelsat 125 vfor50min.Symbols: (0) DMSO-treated 42S; (O)DMSO-treated 28S; (0) untreated 42S; (-) untreated 28S.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.498.104.395.368.577.2]cM
0 y
3-10
000
10 20 30
7
%gel
FractionNumber
FIG. 7. Effect of area oln viral RNA. 32P-labeled viralRNA wasdissolvedin EPB (see text) containzing
10 m1 urea an2d in2cubatedat 37 C for 30 miti. It was
analyzed on 2.5 to 0.5%'; acrylamide-agarose
com-posite gels at 100 v for I hr. Symbols: (a)
urea-treated viral RNA; (0) unztreated viralRNA conitrol.
verylittle effect onthe mobility of large fragments of RNA inpolyacrylamide gels (12), so the sug-gestion that thechange in sedimentation behavior of WEE RNA from 42 to 26S upon heating is due to an extreme conformation change (15) is not consideredfeasible. It appearsthat hiddenbreaks were present in arboviral RNA previously used for studies ofsedimentation behavior and electro-phoretic mobility. The data in Fig. 1 demonstrate clearly that nicked RNA sediments ongradients andmigrates on gels in a manner indistinguishable fromthe nativemolecule, anddivergence fromthe pattern is observedonly when ribonuclease action is more extensive (Fig. 3). We show in Fig. 4 that break-free Sindbis RNA is not fragmented when heatedeither inhigh (STB)- or low (EPB)-ionic-strength buffers at 85 C. Sindbis RNA melts at 65.5 C (14), and a polyaggregate RNA would have dissociated under the heating conditions used in the experiment.
We found no evidence that denaturing agents such asDMSO and urea led to fragmentation of the genome, whichshould have beendetected by increased mobility onacrylamide gels. Similarly, Sreevalsan et al. (15) found no change in sedi-mentation coefficient in sucrose gradients when
WEE RNA was treated with 75, DMSO and
subsequently rerun ongradient intheabsence of DMSO. We conclude that RNA extracted from
IS Viroal RNA
4S
*S "s
S ^_-m--!1te-m-9_Les-e*!(__*F«--.--¢se-E.¢e*rMI-s#e3-- a
I I I I
4 8 12 16 20 24 28 32 36 40 44 48 52 56
Fraction Number
FIG. 8. Absence ofsmall-molecular-weight RNA fromthegeniomeo,Sinzdbis vurus. 0P-labeledviral RNAwas
dissolved in 10 jliters ofEPB (see text) followed by0.1 mlofDMSO an2dinicubatedat room temperature for
10 min. Itwastheiilayeredon 1O0%o polyacrylamide gels (0.2570bisacrylamide), an2d electrophoresis wascarried
out at50 vfor50 miti atroomtemperature. Symbols: (0) viralRNA; (0) 28SRNA; (0) 18S RNA; (U) 4S
RNA.
CPM x10-3
10
-8
6- \42
2
s'o I
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.498.61.252.57.312.2] [image:7.498.121.420.393.585.2]SINDBIS
virions is present as a single strand and not made upofsubunits joined by hydrogenbonding, since the subunit structure would have been detected inour experiments in which RNA was treated at pH2.9 or with formaldehyde, DMSO, or urea.
The data in Fig. 6 show that no cleavage oc-curred when Sindbis RNA was sedimented in sucrose gradients prepared in
99%
DMSO-1 mM EDTA since thepeak fractions taken from these gradients comigrated with untreated 42S RNA ongels. Inlight of the foregoing, it isdifficultto understand how Sindbis RNA as well as WEE RNA sedimented at rates similar to that of 28S rRNA or viral 26S RNA in sucrose gradients prepared in DMSO (5, 15), where there exists a linear relationship between S25 of RNA in 99%,0DMSOand log molecular weight(17).If this technique is used todetermine molecular weight ofRNA,extremelyprecisemeasurementshaveto be made since, using the formula given (17), the S25 value ofRNA havinga molecular weight of 1.7 x 106is4.44, and an RNA having molecular weight of 4 X 106 has anS25 of5.66. Thus, ap-parent similarities in sedimentation coefficients observed insucrose gradients preparedinDMSO between arbovirus RNA and 28S cell RNA (5) maybe questioned on technical grounds when the resolution of the method is not known. Factors such as these may help explainthediscrepancies noted in estimations of the molecular weight of Sindbis RNA madeby this technique (5, 16) as opposed to estimates made byusing polyacryla-midegelelectrophoresis (3, 8).Theworkreported here emphasizestheneedto check that RNA used in biophysical studies is break-free. We findnoevidencethatSindbisviral RNA exists other than as a continuous poly-nucleotide strand and consider that previously statedtheories aboutitspolyaggregate natureare unfounded.
ACKNOWLEDGMENTS
WewishtothankManjulaChandullalforvaluabletechnical assistance.
This workwassupported by a grant from theMedical Research Council, Ottawa, Canada.
LITERATURECITED
1. Arif,B.M.,and P.Faulkner. 1971. Release of viral hemagglu-tinin and nucleocapsid components after disruption of Sindbis virus withether.Can.J. Microbiol. 17:161-169. 2. Boedtker, H. 1967. The reactions of RNA with
formalde-hyde. I.Optical absorbance studies. Biochemistry 6:2718-2727.
3. Cartwright,K.L., andD. C.Burke. 1970. Virus nucleicacids formed in chickembryocells infected withSemlikiForest virus. J.Gen.Virol. 6:231-248.
4. Clarke, D. H.,and J. Casals.1958.Techniquesfor hemaggluti-nation and hemagglutination-inhibition with arthropod-borneviruses.Amer. J.Trop.Med.Hyg.7:561-571. 5. Dobos, P.,B. M.Arif, andP.Faulkner. 1971. Denaturation of
Sindbis virusRNAwithdimethylsulphoxide.J. Gen.Virol. 10:103-106.
6. Dobos, P., and P. Faulkner. 1969.Characterizationofa cyto-plasmicfraction fromSindbis virus infected cultures.Can. J. Microbiol. 15:215-222.
7. Dobos, P., and P.Faulkner.1969. Propertiesof42S and 26S Sindbis viralribonucleic acid species. J. Virol. 4:429-438. 8. Dobos, P., and P. Faulkner. 1970. Molecular weight ofSindbis
virusribonucleic acidasmeasured bypolyacrylamidegel electrophoresis. J. Virol.6:145-147.
9. Fenwick, M. L. 1968.Theeffect ofreaction with formalde-hyde on thesedimentationratesofribonucleicacids. Bio-chem.J. 107:851-859.
10. Granboulan, N.,and K. Scherrer.1969.Visualization in the electron microscope and size of RNA from animalcells. Eur.J. Biochem. 9:1-20.
11. Kaariainen,L., andP.J. Gomatos.1969.Akinetic analysisof thesynthesis in BHK/21 cells of RNA'sspecific forSemliki Forestvirus.J. Gen.Virol.5:251-265.
12. Loening, U. E. 1969. The determination ofthe molecular weightofribonucleic acid by polyacrylamide gel electro-phoresis.Theeffects of changes in conformation.Biochem. J.113:131-138.
13. Peacock,A.C., andC.W.Dingman. 1968. Molecular weight estimation andseparation of ribonucleic acid by electro-phoresis inagarose-acrylamide compositegels. Biochemis-try7:668-674.
14. Sprecher-Goldberger, S. 1967.Differences between the struc-tures ofpoliovirus and Sindbis virus infectious RNA's. Arch.Gesamte Virusforsch. 20:225-230.
15. Sreevalsan,T., R. Z.Lockart, Jr., M.L.Dodson, Jr.,andK. A. Hartman. 1968. Replication of Western equine en-cephalomyelitis virus. I. Some chemical andphysical char-acteristics of viral ribonucleic acid. J.Virol. 2:558-566. 16. Strauss, J. H. 1969. In H. Levy (ed.), Thebiochemistry of
viruses,p.34. MarcelDekker, New York and London. 17. Strauss, J. H.,R. B.Kelly,and R. L.Sinsheimer. 1968.
De-naturation of RNA with dimethylsulfoxide. Biopolymers 6:793-807.
109
VOL. 9, 1972