0022-538X/86/070142-12$02.00/0
Copyright C) 1986, American Society for Microbiology
Antigenic
Variation (mar Mutations) in Herpes Simplex Virus
Glycoprotein B Can Induce
Temperature-Dependent
Alterations in
gB Processing and Virus
Production
S. D. MARLIN,'t S. L. HIGHLANDER,' T. C. HOLLAND,2t M. LEVINE,3 ANDJ. C.
GLORIOSO2*
The GraduateProgram in Cellular and Molecular Biology,' Department of Microbiology and Immunology and Unitfor LaboratoryAnimal Medicine,2 andDepartmentofHuman Genetics,3 University ofMichigan Medical School,Ann Arbor,
Michigan48109-0010
Received 27 December1985/Accepted 25 March 1986
Monoclonal antibody-resistant (mar) mutants altered in the antigenicstructure of glycoproteinB (gB) of herpes simplex virustype 1, strain KOS-321,were selected byneutralization witheachof six independently
derived gB-specific monoclonal antibodies. Analysis ofthereactivitypatternsofthesemarmutantswithapanel
of 16virus-neutralizingmonoclonal antibodies identifiedatleastfivenonoverlapping epitopesonthisantigen,
designatedgroupsIthrough V. Multiplemarmutationswerealso introducedintothe gBstructural geneby
recombination and sequential antibody selection to produce a set ofmarmutants with double, triple, and quadruple epitope alterations. Group II (B2) and group III (B4) antibodies were used to select the corresponding mutants, mar B2.1 and mar B4.1, which in addition to carrying the mar phenotype were
temperature sensitive (ts) for processing of the major partially glycosylated precursor of gB, pgB (Mr =
107,000),tomaturegB (Mr = 126,000)andshowed reducedlevelsofgBonthe cellsurfaceathightemperature (39°C). These mutantswerenot, however,tsforproductionofinfectious progeny. Arecombinantvirus,mar
B2/4.1, carryingboth of thesealterations wastsfor virusproductionand failed toproduceand transportany detectablematuregBtothecell surfaceat39°C. Rather, pgB accumulatedintheinfected cell. Revertants of the ts phenotype, isolated from virus plaques at 39°C, regained the B2 but not the B4 epitope and were
phenotypically indistinguishablefrom themarB4.1 parent. Finally,itwasshownthatgroupII(B5)andgroup
III(B4) antibodiesfailedtoimmunoprecipitate pgB (39°C)produced bytsgBmutantsofherpes simplex virus type 1whichwerenotselected withmonoclonal antibodies.Takentogether, ourfindingsindicate that(i)mar
mutations can alter antigenicas well as otherfunctional domains ofgB, namely, the domain(s) involved in processingandinfectivity,and(ii) groupIIandgroupIIIepitopeslie withinanessential functional domain of
gB which isa targetfortsgBmutations.
Herpes simplex virus (HSV) encodes at least six
glyco-proteins designatedgB,gC, gD, gE, gH, andgG (3, 34, 36).
Although many of the HSV glycoproteins were first
de-scribedover 10 years ago(34), littleinformation is available concerning their functionin virusreplicationandinfectivity.
It is clear that they are incorporated into virion envelopes and are transported to the cell surface membrane during infection (36). Because of their exposed position on these membranes they play a crucial role in virus infectivity, pathogenesis, and the immunobiology ofinfection (12, 21,
24, 25, 35).
Thusfar, gB is the only glycoproteinfor which tempera-ture-sensitive(ts)mutantshavebeendescribed (7,13, 22,30) identifying gBas anessentialvirus component. Although ts
gB mutants grow normally and produce phenotypically
normal gB at permissive temperature (34°C), they produce very little fully glycosylated gB atthe nonpermissive tem-perature(39°C). Rather,alower-molecular-weight form(Mr
= 107,000 [107K])accumulates in thecytoplasm, aproduct which is not exposed on the cell surface as is the mature
species(Mr = 126,000) (30). In contrast tothematureform,
the 107K species is endoglycosidase H sensitive and is
* Correspondingauthor.
t Presentaddress: Dana Farber CancerInstitute, Harvard Uni-versity Medical School, Boston, MA 02115.
t Present address: Department ofMicrobiology and Immunol-ogy, Wayne State MedicalSchool, Detroit, MI 48201.
similar in sizeto themostabundantgB precursor, pgB(18,
37). It appears that ts gB mutant virus produced at39°C is
enveloped and will attach to host cells (22, 30). However,
these mutantsfailtopenetrate host cell surface membranes
unless the virion envelopes are artificially fused to the cellular plasma membranes with polyethylene glycol (22, 30). These
findings
wereinterpreted
to mean that gB is essentialtothe processof viruspenetration (22, 30). Relatedtoitsfunctioninviruspenetration, gBhas been showntobe involved infusingcellmembranes, therebyformingsyncytia
(7, 20). One of thefoursynlociin HSVtype1 (HSV-1)has been physically mapped to the coding sequence for the
cytoplasmic domain of gB (7, 20).
The HSV glycoproteins also have been shown to be the
principal antigensresponsiblefor the induction ofimmunity
to HSV infection (6, 35). They induce both humoral and cell-mediated protective immune responses and
provide
targetantigens for nonimmune defense mechanisms suchas natural killer cells (1, 2, 4, 5, 9, 13, 24-27, 32, 33, 39). The
characterization of the antigenic structure of the HSV
gly-coproteinsis essentialto ourunderstandingof the
immuno-biology of HSV infection. Studies along these lines have beenaidedbyusingHSVglycoprotein-directed monoclonal
antibodies(8, 15, 16,20, 23,28).Thesehighlyspecific probes
have been used to catalog distinct antibody-binding sites
(epitopes) andorganizethem into antigenic sites composed
ofoverlapping epitopes (10, 23).
142
on November 10, 2019 by guest
http://jvi.asm.org/
Our laboratory has used virus-neutralizing monoclonal
antibodiesto select aseries of antigenic variants in each of
the HSV-1 glycoproteins gC, gB, and gD; the resulting
strains are referred to as monoclonal antibody-resistant
(mar) mutants (15). The identification of unique reactivity patternsofmar mutantswithpanels ofmonoclonal antibod-ies operationally defines distinct epitopes (23). mar
muta-tions alterthe sequence orconfiguration of aminoacids with which an antibody reacts. Thus far, all mar mutations studied have been shown by marker rescue to map physi-cally to their respective glycoprotein structural genes (16,
28). The identification ofthesemutationsby DNA
sequenc-ing indicatesthey resultfrom point mutations (28;B. Wu, F. Homa, L. Holland, D. J. Dorney, J. C. Glorioso, and M.
Levine, manuscript in preparation). In this report, we
de-scribe the useof monoclonal antibodiesand mar mutants to extend our understanding of the antigenic structure of
HSV-1 gB. In addition, we provide evidence that mar
mutations in gB appear toalter this antigen athigh temper-ature,which results in defectsingBprocessingandfunction
in virus replication.
MATERIALS ANDMETHODS
Virus strains. Wild-type HSV andantigenic variantswere grown andtiterswere determinedon Africangreen monkey kidney (Vero) cells at 37°Cby infection at low
multiplicity
(15). Plaque-purified isolates of wild-type HSV-1
(strain
KOS) and HSV-2 (strain 186), designated KOS-321 and
186.111, respectively, were used in all
experiments.
The KOS mutant, tsF13, wasisolatedin M. Levine'slaboratory
by in vitro mutagenesis ofthe EcoRI F
fragment
and hasbeen described previously (13). Also
employed
were ts B5(HSV-1, strain HFEM) and ts J12
(HSV-1,
strainKOS),
kindly provided by Patricia Spear
(University
ofChicago)
andPriscilla Schaffer(Harvard
University), respectively.
All three tsgB mutants were tsforvirusgrowth andprocessing
of pgB to gB (13, 22, 30). The ts mutants were grown and
titerswere determinedat 34°C (13).
Cell cultures and media. Human
embryonic lung (HEL)
and Vero cells were grown and maintained in
Eagle
mini-mumessential medium(GIBCOLaboratories,
GrandIsland,
N.Y.) supplemented with nonessential amino
acids,
10 mMHEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid), and 10% fetal calfserum
(GIBCO Laboratories).
TheBALB/c myeloma cell line, P3-X63-Ag8.653, and the
my-eloma-spleen cell hybridomas were grown in
supplemented
Dulbecco modified Eagle medium (GIBCO
Laboratories)
containing 20% fetal calf serum
(Sterile
Systems, Inc.,Logan, Utah),asdescribed previously (15).
Production of hybridomas and polyclonal antisera. The
procedures used fortheproduction of hybridomas
secreting
neutralizingmonoclonal antibodies specificfor HSV-1 have been described in detail elsewhere (15).
Briefly,
BALB/cmicewereinfected
intraperitoneally
with 2 x 107PFUofliveKOS-321 virusonday1and boostedon
day
13with108
PFUof UV-inactivated virus. On
day 16,
immunespleen
cells andP3-X63-Ag8.653 myeloma cells were fused using
polyethyl-eneglycol.
High-titer ascites fluid
preparations containing
monoclo-nal antibodies were produced aspreviously
described(23).Antibodies fromclarified ascites fluid were concentratedby
ammonium sulfate precipitation and characterized for
monoclonality byisoelectricfocusingasdetailed earlier(23).
Monoclonalantibody isotypes were determined
by
enzymeimmunoassay with
subclass-specific
antiserausing
aMonoAb-ID EIA Kit
(Zymed Laboratories,
SanFrancisco,
Calif.).
In someexperiments,
apool
ofequal
volumes ofantibodies
B2, B3,
B4,
and B6 was used. The monoclonal antibodies3S,24S, 25S, 30S, 33S, 40S,
and 61Swerekindly
provided
by MartinZweig (National
CancerInstitute,
Frederick, Md.).Theseantibodieswere
produced
from miceimmunized with HSV-1
(strain 14012)
and have beenchar-acterized
by
Showalteret al.(33).
To produce
HSV-1-specific antisera,
groups of threeBALB/c mice were immunized
by
intraperitoneal
infection with various dosesofvirusranging
from102
to106
PFU per mouse.Themicewerebled fromtheretroorbitalcavity
at3, 5, 7,and 10days afterimmunization,
andserafromeachday
were pooled.
Virusneutralization assays.
Wild-type
virus andantigenic
variantsweretestedfor
reactivity
withmonoclonal antibod-ies in 50%plaque
reduction microneutralization assays asreported
previously
(23). Neutralization titers ofconcen-trated ascites fluid stockswere
expressed
asthereciprocal
ofthe
highest
dilution which reducedastandardized virusinput
(20 to 30 PFU per
well) by
at least 50% relative tonon-neutralizedcontrols.
Mouse antiserawere
assayed
forvirus-neutralizing
activ-ity
inlarge
scale assays.Equal
volumes(0.5
ml)
ofserially
diluted
antibody
and virus stock(2,000
PFU/ml)
weremixed andincubated asdescribed above. Titers ofsurviving
virus were then determined on Vero cellmonolayers
in 60-mmdishes,
in whichapproximately
200plaques
could becounted in the unneutralized controls. Results were then
expressed
as percent neutralization at eachantibody
dilu-tion.Enzyme immunoassays. The
binding
of monoclonal anti-bodiestoglutaraldehyde-fixed,
virus-infectedHELcellswasdetected
using
horseradishperoxidase-conjugated
goatanti-mouse
immunoglobulin
G(IgG)
inanenzymeimmunoassay.
This assay has beendescribedindetail
by
Marlinetal.(23).
A405valuesweredetermined withaTitertekMultiscan
(Flow
Laboratories,
Rockville, Md.).
Theimmunoperoxidase
"black
plaque"
assay wasperformed
aspreviously
de-scribed
(16).
Briefly,
Vero cellmonolayers
in 60-mmdishes were infected with 20 to 200 PFU ofvirus,
glutaraldehyde
fixed after
plaque
formation,
and incubatedwithgB-specific
monoclonal antibodies.
Next,
monolayers
were incubatedwith horseradish
peroxidase-conjugated
goat anti-mouseIgG.
Subsequent
treatment with4-chloro-1-napthol
pro-ducedablackprecipitate
onplaques
towhichgB
antibodieswerebound.
Radiolabeling,
immunoprecipitation,
andelectrophoresis.
HEL cells were infected with
wild-type
KOS-321 virus orantigenic
variants at amultiplicity
of 10 and radiolabeledwith 40
,Ci
of[35S]methionine
per ml from 4 to 18 hpostinfection
at either 34 or39°C.
Nonidet P-40extracts ofradiolabeled infected cells were
immunoprecipitated
with apool
ofgB-specific
monoclonalantibodies, electrophoresed
on 7%
polyacrylamide
slabgels
cross-linked withdiallyl-tartardiamide,
andfluorographed
asdetailedpreviously (16).
Isolation of
single
andmultiple antigenic
variants.Anti-genic
variants ofwild-type
HSV-1(KOS-321)
resistant to neutralizationby single
gB-specific
monoclonal antibodies in the presenceof rabbitcomplement
were isolatedessentially
as described earlier
(15).
Isolates of the virussurviving
neutralizationwere
plaque
purified
threetimes and tested for neutralization in the 50%plaque
reduction assay described above. Thoseantigenic
variants whichqualified
were desig-natedmonoclonalantibody-resistant (mar)
mutantsaccord-ing
topreviously
definedcriteria(15).
Multiple
marmutantson November 10, 2019 by guest
http://jvi.asm.org/
wereselected sequentially byusingtwoormoremonoclonal
antibodies from virus produced during mixed infections with singlemar mutants(see Results).
Some antigenic variants were isolated from stocks of
KOS-321 mutagenized with 5-bromo-2-deoxyuridine as
re-ported earlier (15). Briefly, viruswas grownin Vero cellsat
amultiplicity of infection (MOI) of 5 for 24 h in thepresence
of2.5 ,ug of5-bromo-2-deoxyuridine perml. Before
mono-clonal antibody selection, the mutagenized virus was
pas-sagedonVerocells atan MOI of 0.01 for 24 h toeliminate phenotype mixing ofmutant and wild-typeglycoproteins in thesame virion envelope.
Isolation of ts+ revertants from ts double mar mutants. Revertantvirusesnolongertsforplatingwereisolated from
thedoublemutant marB2/4.1 by passageatthe nonpermis-sivetemperature. Vero cell monolayerswere infectedatan
MOI of 0.01 and incubated at 39°C until cytopathic effect
wasgeneralized. For isolation of therevertant,R3marB4.1, two rounds of low-MOI passage at 39°C were done. The appearance of putative revertants was identified by
deter-mining titers of the resulting virusat34 and39°C. Individual plaqueswere picked from the plates incubatedat 39°C, and small stocks were grown in Vero cells at 34°C. The virus isolateswerethenplaquepurified three timesandtested for
plating efficiency and virus yield at permissive and nonpermissive temperatures.
RESULTS
Characteristics of monoclonal antibodies. Apanel of mono-clonal antibodies was produced for use as probes of the
antigenic structureofgB. Nine hybridomas secreting virus-neutralizing monoclonal antibodies were derived from six
independent fusions using spleen cells from BALB/c mice immunized by infection with wild-type HSV-1 (KOS-321)as
describedpreviously (23). As demonstrated earlier (15),the antibodies were determined tobe specific forgB by
immu-noprecipitationof[35S]methionine-labeled gB from Nonidet P-40lysates of HSV-1-infected cells (datanot shown). The derivation andcharacteristics of the monoclonal antibodies arelisted inTable 1. All oftheantibodies neutralized HSV-1 tohigh titers in thepresenceof rabbitcomplement,butonly antibody B4 produced significant neutralization in the ab-senceofcomplement. Six oftheantibodies(B2, B4, B5, B7, B8, and B9) were cross-reactive with HSV-2 and thus
[image:3.612.320.560.86.187.2]recognized epitopes shared by thetwo strains tested. TABLE 1. Characteristics ofgB-specificmonoclonal antibodies
Neutralization titer Monoclonal Fusion Isotype HSV-1(KOS-321) HSV-2
antibody no. HSV_____2_
+a _b (186.111)c
B1 1 IgG3 2,560 <10 <10
B2 3 IgG2b 163,840 <40 40,960
B3 4 IgG3 5,120 <40 <40
B4 4 IgG3 2,560 1,280 640
B5 5 IgG3 81,920 <160 5,120
B6 7 IgG2b 81,920 <160 <160
B7 6 IgG2a 655,360 <160 5,120
B8 7 IgG2a 655,360 <160 2,560
B9 8 IgG2a 655,360 <160 40,960
a+, Neutralization mixture contained normalrabbit serum as a comple-mentsourceatafinalconcentration of 10%.
b-,No normal rabbitserum waspresent.
cAll neutralizationmixturescontained normalrabbit serumasa
[image:3.612.66.305.559.687.2]comple-mentsourceatafinalconcentrationofl1o.
TABLE 2. Isolation ofmarmutants with gB-specific antibodies
Selection Selecting Frequencyof marmutant Mutagena isolation of
expt antibody marmutantb designation
1 B1 BUdR 1 x 10-3 B1.1, B1.2
2 B2 1 x 10-7 B2.1, B2.2
3 B3 1 x 10-2 B3.1,B3.2
4 B4 8 x 10-7 B4.1,B4.2
5 B5 3 x 1O-7 B5.1
6 B5 BUdR 5 x 10-7 B5.2
7 B6 1 x 10-6 B6.1, B6.2
aBUdR,5-Bromo-2-deoxyuridine.
bFrequency = fraction of virus surviving neutralization x fraction of isolateswiththemarphenotype.
Isolation of mar mutants. The gB-specific monoclonal
antibodies were used to select aseriesof mar mutantswhich
wereresistant toneutralizationyetexpressed gB in infected
cells and incorporated this antigen into virion envelopes. The mar mutants were obtained by neutralizing plaque-purified wild-type strain KOS-321 virus with single
mono-clonalantibodiesin the presence ofcomplement. The virus
survivingneutralization (typically lessthan0.1%) was
pas-saged at low MOI and subjected to a second round of neutralization. Plaque-purified isolates ofthesurviving virus
were thentestedfor resistancetoneutralization in
quantita-tive
50%
endpoint plaque reduction assays. Anisolate wasdesignatedas amarmutantifthe monoclonalantibody titer against the mutant was at least 32-fold less than the titer when tested against the wild-type parent virus, KOS-321 (15). In most cases, even undiluted selecting monoclonal
antibodies failed to neutralize the corresponding mar mu-tants. marmutant-infected cellsexpressed gB as shownby specific radioimmunoprecipitation with a pool of the gB-reactive monoclonal antibodies followed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (datanotshown).
Insevenindependentexperiments, sixgB-specific
mono-clonalantibodieswere usedto isolate12 mar mutantsfrom mutagenized or nonmutagenized virus stocks. The deriva-tion and frequency of isolation of these mutants is
summa-rized in Table 2. The frequency of isolation of the mar mutants varied from
10-2
to10-7.
Operational antigenic map ofgB. The availability ofmar mutants and gB-specific monoclonal antibodies provided a means of analyzing the antigenic structure of this viral
glycoprotein. Each mar mutantshould express an antigeni-cally variant gB with a structural alteration that affects
reactivity with one or moremonoclonalantibodies. Thus, at least one epitope ongB has been alteredineach mutant. A
panel ofmar mutantswith different mutationscanbe usedto
discriminate between monoclonal antibodies reactive with different epitopes on the glycoprotein. As we have previ-ously shown with
gC-specific
monoclonal antibodies and mar C mutants, an operational antigenic map can be con-structed from the reactivity patterns ofmar mutants with monoclonal antibodies (23).Alterations indifferentepitopescan be identified by unique patterns of reactivity, while
overlappingpatterns ofreactivitycanbe
grouped
togetherto defineantigenicsites composed ofmultipleepitopes (23).Wetested thepanel ofgB-specific antibodiesforreactivity
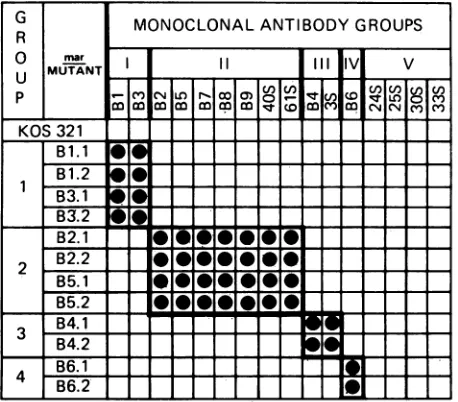
with the mar mutantsin virusneutralization assays(Fig. 1).
Inaddition to the nine antibodiesagainstKOS-321produced
in this laboratory, we also tested seven produced by
Showalteret al. (33) againstHSV-1 strain 14012
(antibodies
3Sthrough61S). Eachantibody orgroupof antibodieswith
on November 10, 2019 by guest
http://jvi.asm.org/
G
MONOCLONAL ANTIBODY GROUPS
0 nr
IIi
IIIIIV
vu MUTANT
C.0() (tJc1)n 1
P
C") CN L_O
r- 00o
racT-
(_
(nCT
(LO C)ma m co m co m vtC mVM coC1 C1 m cn
KOS321 _ -_
B1.1 *-_
B1.2 *
-B3.11
f
B3.2 * _-_
B2.1
2
B2.2
-
* * * * _ _B5.1 __ * *
0
* * ___B5.2
_ * * _ _ _3 B4.1 __ ___ _ __ L _- - -_
B4.2 --
-B6.1
_
_
B6.2_FIG. 1. Operationally defined epitopes of HSV-1 (KOS-321) gB. Individual monoclonal antibodiesweretested for neutralization of wild-type virus (KOS-321) and mar mutants in50% plaque reduction neutralization assays. The virusesweredesignated as resistant(0) orsensitive
(L)
toneutralization with eachantibody. The criterion for resistance was that thetiter of the antibody, when tested against the mutant, was at least 32-fold less than the titer against the wild-type virus.aunique reactivity patterndefined adifferentepitope on gB.
Accordingly, a minimum of five nonoverlapping epitopes (designated I through V) were identified by the distinct verticalpatterns. With the exception of antibodies
represent-ing epitope V, for which mar mutants have not been
se-lected,alloftheantibodies produced against KOS-321could
be placed into groups with
corresponding
mar mutants,designatedgroup IthroughgroupIV. Ofthe seven antibod-ies produced against strain14012, three(antibodies 3S,40S,
and 61S) had reactivity patterns similar to antibodies
pro-ducedagainst KOS-321. The remaining four antibodies
neu-tralized all of the mar mutants as
effectively
as wild-type virus and constitute antibody group V. Although the anti-bodies in this group have the same reactivity pattern anddefineatleast oneepitope, theyarenotnecessarily reactive with the same epitope and could define as many as four differentepitopes.
Identification of distinct antibody clonotypes byIsoelectric Focusing. To identify the number of different monoclonal
antibody
clonotypes in the above analysis, the gB-specific antibodies produced against KOS-321 were examined for their isoelectric focusing banding patterns (Fig. 2). Thisanalysis showed that the nine antibodies produced against KOS-321represent sevendistinctclonotypes. AntibodiesBi
andB3, whichcomprisegroupI,hadsimilarelectrophoretic patterns andprobably represent the sameclonotype. How-ever, these two antibodies were not derived from sister hybridoma clones since they wereisolated in two separate fusion experiments (Table 1). Of the five antibodies which
recognized thegroup II epitope, antibodies B7and B8 had
similar banding patterns (although derived from separate
fusions), while antibodies B2, B5, and B9 appeared to be
distinct and again were isolated from separate hybridoma fusion experiments (Table 1). Antibodies B4 and B6, in groupsIIIandIV,respectively,haduniquebanding
patterns
andweredifferentfrom the othergB-specific antibodies. Selection of mar mutants with multiple epitopechanges in
gB. Mutants withmultiple antigenic changes mightbealtered inotherphenotypes, suchastheirreactivitywith
polyclonal
antisera produced against wild-type HSV-1 (KOS-321) or suchas virus production athigh temperature. To examine these possibilities, mar mutants with multiple epitope changes in gB were isolated and studied.
A series of multiple mar mutants were constructed with two, three, and four epitope changes in gB. Since the
frequency of mutants with simultaneous changes in more than one epitope could be expected to be very low, the
epitope changes in gB were introduced sequentially. Two double mutants, mar B3/4.1 and mar B2/4.1, were first isolated by recombination after mixed infections with mu-tants mar B3.1 and mar B4.1, or mar B2.1 and mar B4.1.
B1
B3 B2 B5 B7 B8
B9
B4 B6
1+I
A.
JR
-I
-m.i.
l-I
FIG. 2. Isoelectric focusing ofgB-specific monoclonal antibod-ies. Antibodies were electrophoresed in a5% polyacrylamide gel (pH 4.0 to 9.0) at 2 mA for 18 h as described in Materials and Methods. The electrophoretically separated proteins were visual-izedbystainingwithCoomassiebrilliant blue. The anode(+)and cathode(-)areindicated.
4S
adM
;W
44mom
'Aqmvml.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.58.287.70.271.2] [image:4.612.314.555.231.660.2]Because HSV recombines actively, it was expected that somemultiplemarmutantrecombinantswouldbe produced. Progeny virus of the mixed infections was subjected to
neutralization with a mixture of the relevant monoclonal antibodies (B3 + B4 or B2 + B4) to enrich for recombinants with the double mar mutant phenotype. These antibody
mixtures neutralized the two parentalvirus mutants as well as wild-type virus recombinants, allowing only double mu-tants tosurvive selectionby neutralization. marB3/4.1 and mar B2/4.1 had the expected patterns of resistance to neutralization (Fig. 3); these mutants were resistant to
neutralizationby antibodies of the two appropriate antibody groups. Thereactivitiesofrepresentativesingle mutants are
also shown in Fig. 3 for comparison.
We then introduced mutations affecting epitopes
recog-nized by monoclonal antibodies B6 and B3 by means of sequential immunoselection against mar B2/4.1 progeny
virus. In this manner, we isolated the triple mutants mar
B2/416.3 and mar B2/4/6.4, which have alterations in the three epitopes definedby antibodies B2, B4, and B6. In an
independent selection, antibody B3 was used to isolate the
quadruple mutants mar B2141613.3 and marB2141613.4 from the triple mutant mar
B214/6.3.
The reactivity patterns of thesemutants toneutralization with thegB-specificantibod-ies arealsoshown in Fig. 3. As expected, thetriplemutants wereresistanttoneutralization with thegB-specific antibod-ies representing three of the epitopes ongB, andthe
quad-ruplemutants were resistant to neutralization by all of the antibodies tested, representingthefourantibodygroups.
MONOCLONAL ANTIBODY
mar _____
MUTANT IV
-. CV) CN LD) cm(
B1.1
B3.1 @0
B2.1 _
B5.1 @0
B4.1 B6.1
B3/4.1 * *
B2/4.1 * * *
B2/4/6.3
B2/4/6.4 * * * *
B2/4/6/3.3 _ _
B2/4/6/3.4 * * _
R3B14.1
FIG. 3. Neutralizationpatternsofmutantswithmultipleepitope changesin gB. Mutantsweretested forsensitivitytoneutralization with the gB-specific antibodies
Bi
through B6 in 50% plaque reduction neutralization assays. The mutants were designated as resistant(0)orsensitive(OI)
toneutralization as described inFig.1.100
80-z 0
$ 60
N
I
40-L1
z
20-0
9 10 11 12 13 14 15
LOG2 ANTIBODY DILUTION
FIG. 4. Neutralization ofmultiple mar mutants with antiserum. Wild-type KOS-321 (0) and mar B2/41613.1 (A) were neutralized with twofold serial dilutions of mouse antiserum. The antiserum was produced by infecting mice with 103 PFU of live KOS-321 virus intraperitoneally. The mice were bled at 7 dayspostinfection, and sera were pooled from three mice for virus neutralization assays. Thepercent neutralization wascalculated from triplicate assays of thesurvivingvirus.
Reactivity ofmultiplemar mutants with antisera produced against wild-type HSV-1 (KOS-321). The effect of antigenic alterations present in multiple mar mutants on reactivity withapolyclonal antiserumproducedagainstwild-type virus
was tested. This would examine whether the multiple epitope alterations in gB significantly affected the overall antigenic structure of the virus envelope. Antisera against KOS-321 were produced by infecting groups ofmice with various doses of live virusranging from
102
to106
PFU permouse. The mice were then bled at 3, 5, 7, and 10 days postinfection, and the
neutralizing
activity titers of thesera were determined. Sera frommice immunized with103
PFU hadneutralizing titers of 1,024at7days postinfection. Mice bledatearlier timesorimmunized with lower doses ofvirusproduced verylow titers (<64). The day 7 antiserum from mice immunized with
103
PFUwasstrongly reactive with gBas detectedby
radioimmunoprecipitation
(datanot shown). Weak reactivities with gC and gD were also noted. The observation that gB is themajor
inducer of immuno-precipitating antibodies during HSV infection agrees withourearlier
findings
(13). This antiserum was tested for its abilitytoneutralizewild-type
KOS-321 andmarB214/613.3. Despite thefactthat themar mutantlacked fourepitopes
ingB, and gB is a major target antigen for precipitating antibody, the quadruplemutantappearedtobeassensitive
toneutralization with this antiserumas
wild-type
virus(Fig. 4). Thus, thecombination offour differentepitope
changesingB didnot
significantly
affectreactivity
withapolyclonal
antiserumproduced againstwild-type
virus,
suggesting
thatthese epitopes were not strongly immunodominant in the
totalantiviral antibody response.
marmutants withmultiplemutations exhibita tsphenotype.
Viruses with point mutations in the structural gene of
gB
havebeen identifiedwhichshow
temperature-dependent (ts)
defects in
processing
ofpgBtogB,inplaque
formation,
andon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.321.558.71.282.2] [image:5.612.93.269.372.670.2]TABLE 3. Effects of temperature on plating efficiency and production of mar Bmutantsa
Platingefficiency Virus yield Virusmutant Temp Fraction of Fraction of
M~) PFU/ml 34Cvle PFU/ml 34'Cvalue
KOS-321 34 9.2 x 107 3.1 x 108
39 4.2 x 107 0.443 1.4 x 107 0.044
mar B1.1 34 9.0 x 107 1.2 x 108
39 2.2 x 107 0.250 1.5 x 106 0.012
marB2.1 34 1.7 x 108 9.5 x 107
39 7.9 x 107 0.465 1.3 x 106 0.014
marB3.1 34 6.4 x 107 3.5 x 108
39 3.8 x 107 0.568 3.4 x 107 0.097
marB4.1 34 3.9 x 108 2.9 x 108
39 1.4 x 108 0.367 5.6 x 106 0.019
marB5.1 34 2.0 x 107 2.9 x 107
39 8.5 x 106 0.436 5.3 x 105 0.018
marB6.1 34 4.4 x 107 2.3 x 108
39 2.2 x 107 0.506 7.2 x 106 0.031 marB2/4.1 34 8.8 x 107 5.6 x 107
39 2.5 x 104 <0.001 5.0 x 104 <0.001 marB3/4.1 34 2.0 x 107 2.1 x 108
39 8.5 x 106 0.425 4.6 x 107 0.139 marB2/4/6.3 34 7.4 x 107 1.1 x 108
39 4.5 x 104 <0.001 4.6x 104 <0.001 marB2/4/6/3.3 34 3.8 x 107 7.0 x 107
39 1.8 x 104 <0.001 6.0 x 104 <0.001
ts F13 34 9.0 x 106 3.0 x 108
39 1.5 x 102 <0.001 1.6 x 104 <0.001 R3marB4.1 34 1.4 x 108 2.3 x 107
39 5.0 x 107 0.357 1.7 x 106 0.074
aViruses were grown and titers were determined on Vero cells at37°C.
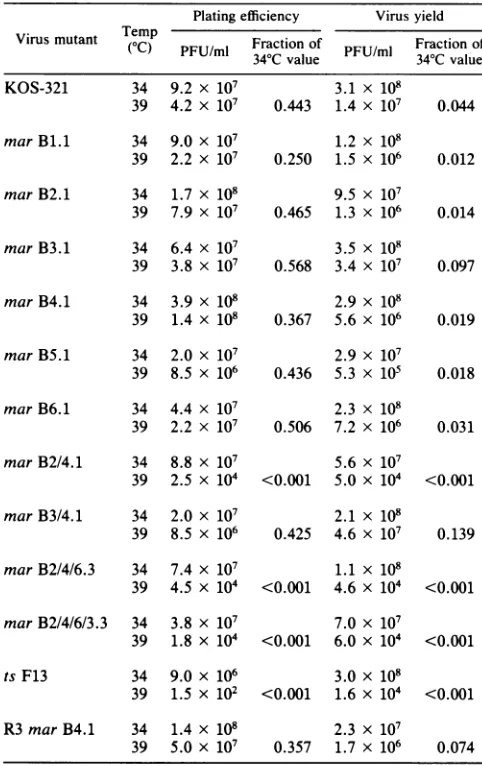
Plating efficiency is presented as the fraction ofplaques formed at 39°C compared with34°C. Virusyieldisdescribed asthe fraction of virus PFU producedat39°C compared with 34°C when titersaredeterminedat37°C.
invirus production (7). DNAsequencing ofmarmutationsin the gB gene has shown that antigenic variation can be conferred by single-base-pair substitutions (28; D. J.
Dorney, S. L. Highlander, M. Levine, and J. Glorioso, unpublished data). This finding raised the question of
whether antigenic alterations in gB can simultaneously
im-pair the ability of gB to function in virus replication in a
temperature-dependent fashion.To examine thispossibility,
marmutantsweretested forefficiency of plaque formationat 34 and 39°C, temperatures which are permissive and
nonpermissive
for virusproduction
of othertsgBmutantsofHSV-1 (KOS) (13, 22, 30). At the nonpermissive tempera-ture, wild-type KOS-321 showed an approximate 2-fold reduction in plating efficiency whereas the previously iso-lated KOS mutant, ts F13, showed a60,000-fold reduction
(Table3). tsF13carriesamutation in the structural gene for gB and is defective in the processing of pgB to the mature
form ofgB at 39°C (13; S. Highlander, unpublished data).
Noneofthesinglemar mutants showedts defects inplating efficiency. However, the double mutant marB2/4.1 and the
multi-mar mutants derived from it showed plating
effi-ciencies reducedto anextentcomparablewith the mutant,ts F13. In contrast, another double mar mutant, mar B3/4.1, was not ts for plating efficiency, demonstrating that the presenceof two marmutations was not in itself sufficient to produce tseffects.
Since these multiple mar mutants were ts for plating efficiency, we determined whether these mutants were also ts forvirus production. Virus production by the single mar mutants was notsignificantly different from that ofwild-type
virus at high temperature (Table 3). However, the multiple marmutants,which wereresistant to both antibodies B2 and B4, exhibited reduced virus production at 39°C, similar to the reduction observed for ts F13 (Table 3). Again, mar B3/4.1 did not show atemperature-dependent defect in virus production.
Effect of temperature on production of mature gB by mar mutants. All previously reported ts mutants mapping to gB
have an associated defect in the processing of the major precursorform of gB, carrying the mannose core sugars, to the mature fullyglycosylated form (13, 22, 30). We examined whether a similar defect in processing of pgB to gB also occurred in cells infected with the mar mutants. Radiola-beled viral proteins from cells infected at 34 or 39°C were
immunoprecipitated with a pool of gB-specific antibodies andanalyzedby sodiumdodecylsulfate-polyacrylamidegel electrophoresis (Fig.SA). Both the mature form of gB and its partiallyglycosylatedprecursor,pgB,were
immunoprecipit-ated from cells infected with wild-type KOS-321 at 34 or
39°C. However, mar B2.1 and mar B4.1 both exhibited
partialbutsignificant decreases in theexpression of mature
gB at 39°C; gB was produced by both mutants, but at
substantially lower amounts than by wild-type virus. In
addition, pgB overaccumulated relative towild-type virus. mar B5.1 had a similar phenotype (data not shown). This mutant was selected with monoclonal antibody B5, which
recognized the same epitope as antibody B2 (Fig. 1). Of theseviruses, marB2.1 appearedtobemoreaffected inthe
production ofmature gB. These findings were unexpected
since none of these mutants was ts for platingefficiency or
virus production. These observations suggest that apartial
defect in gB processing athigh temperature is not in itself sufficient to inactivate the glycoprotein function in virus
infectivity. In contrast, mar B3.1 was not affected in gB processing at 39°C, indicating that mar mutations do not
always affecttheintracellularprocessingofthe glycoprotein. However,therecombinantmarB3/4.1wasaffected in
proc-essing,and thus the defect in this double mutant waslikely
derived from the mar B4.1 parent and not the mar B3.1 parent. Insteadof the partial processing defect seen inthe
singlemutants marB4.1and mar B2. 1,the recombinantmar B2/4.1 (Fig. 5) and all ofthe multiple mar mutants derived fromthisrecombinant (datanotshown) producedno detect-able mature gB at 39°C. In addition, pgB was
immunopre-cipitated
inanoverabundance. Apparently, theintroduction of two mar mutations, each of which partially affected processing ofgB athigh temperature, greatly inhibited the processing of this molecule.Previously,weshowed that mutant ts F13failed to present any form ofgB in infected cell surface membranes at the
nonpermissivetemperature(13).Accordingly,wecompared
the single mutants mar B2.1 and mar B4.1 with the double mutant marB2/4.1 for the ability to present gB on infected cells. Monoclonalantibody B3,whichrecognizesaseparate, unaltered epitope in these mutants, was used to detect the presence of gB on glutaraldehyde-fixed infected cells in
on November 10, 2019 by guest
http://jvi.asm.org/
As
KOS B2.1 B3.1 B4.1 B3:4.1 B2 4.1
34 39 34 39 34 39 34 39 34 39 34 39
pgB'
UUII
UIIUIII
B.
KOS
B 2 /4.1 R3 B4.134 39 34 39 34 39
_O
.."
~
~
~~~~~~~
--gB
w-
w
gB
Sib 1I~~'I'
FIG. 5. Effect oftemperature on processing ofgB marmutants. HEL cells were infected at an MOI of 10 at either 34 or 39°C and radiolabeled with[35S]methioninefor4 to18 hpostinfection.gB wasimmunoprecipitatedfrominfected-cell lysates with a pool ofmonoclonal antibodies (B3, B4, B5, andB6). The immunoprecipitates were electrophoresedin a 7% sodium dodecyl sulfate-polyacrylamide gel and visualized byfluorography. The gel wasintentionallyoverexposed to detect small amounts of theimmunoprecipitatedprotein. The relative positions of thematuregB species and theprecursor, pgB, areindicated.
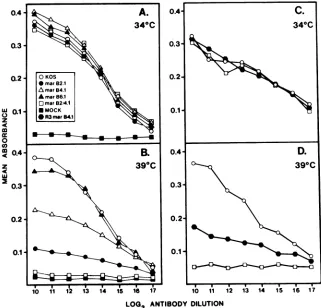
enzyme immunoassays (Fig. 6AandB). At34WC (Fig. 6A),
antibody B3 reacted equally well with cells infected with
wild-type
virus and with mutants mar B2.1, mar B4.1, and marB2/4.1,aswellaswithmar B6.1,whichhas nodefect inw
al.1
0 z
m 0 co 2 0.4. z
w
<
gB processing at 39°C. No reactivity was detected against
mock-infected
cells. Cell surface expression of gBin
cellsinfectedat39°C(Fig. 6B)closely paralleledthe immunopre-cipitation data.Thatis,cellsinfectedwith marB2/4.1had no
0.4
0.3
0.2-0.1
-
0.4-0.3
0.2'
0.1-10 11 12 13 14 15 16 17
[image:7.612.86.542.77.262.2]LOG2 ANTIBODY DILUTION
FIG. 6. Effect oftemperatureonexpression of gBonsurface membranes of cells infected withmarmutants.HELcellswereinfectedat
anMOI of10,ormock-infected, andglutaraldehyde fixed at18 hpostinfection. The fixed cells weretestedfor reactivity with monoclonal antibody B3 by enzyme immunoassay as detailed in Materials and Methods. Each antibody dilution was tested in triplicate. Standard
deviationswerelessthan 0.03.
C.
340C
D.
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.154.475.375.683.2]TABLE 4. Comparison of the ability of monoclonal antibodies reactive with epitope groupsIthroughIVtoimmunoprecipitategB produced by mar and ts gBmutantsof HSV-1
Immunoprecipitation by monoclonalantibodiesa
B3(groupI) B5(groupII) B4(group III) B6 (groupIV)
HSV-1
34°C 39°C 34°C 39°C 34°C 39°C 34°C 39°C
pgBb gBb pgB gB pgB gB pgB gB pgB gB pgB gB pgB gB pgB gB
KOS-321 + + + + + + + + + + + + + + + +
marB3.1 - - - - + + + + + + + + + + + +
marB6.1 + + + + + + + + + + + + + + +
marB5.1c + + + + - - - - + + + - + + + +
marB4.1c + + + ± + + - - + + - - + + + *
tsB5(HFEM) + + + NP + + - NP + + - NP - - - NP
tsF13 (KOS) + + + NP + + - NP + + - NP + + + NP
tsJ12 (KOS) + + + NP + + - NP + + - NP + + + NP
aVerocells were infected at 34 and 39°Catan MOI of 5, radiolabeled with[35S]methionine5hafterinfection, extracted after 15 h with 1% NonidetP-40, and
immunoprecipitated with monoclonal antibody. Monoclonal antibodies all react with gB and represent distinct epitope specificities. The precipitates were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. +, Immunoprecipitation; -, lack of immunoprecipitation; +, weak immunoprecipitation. NP, Mature gB not produced at 39°C.
bMaturegB(Mr = 126K) and its major precursorpgB (Mr = 107K).
cMature gB produced in reducedamounts at39°C.
detectablegB on the cellsurface,whereas cellsinfectedwith marB2.1ormarB4.1 presented gB onthecell surface,but
inamounts substantially lessthanwild-type virus.marB2.1
produced less surface membrane gB than mar B4.1, while the control mutant infection, with marB6.1, wassimilar to
the wild-type virusinfectionin thecell surfaceproduction of
gB.
Comparison of immunoreactivity of gB produced by mar mutants andtsgB mutantsof HSV-1. Asdescribedabove, the
double marmutantmarB2/4.1failed toproduce maturegB at high temperature. This abnormal processing phenotype
hasalsobeenreported forothertsgBmutantsselectedforts virus production, rather than by resistance to neutralizing antibodies (13, 22, 30). Thisobservation raised the possibil-itythat thesetsgBmutantsalsoproduceaform of gB which lacks the group II and group III epitopes. To examine this question, group II (B5) and group III (B4) monoclonal antibodiesweretestedfor theirabilitytoimmunoprecipitate
pgB and gB (at 34°C) and pgB (at 39°C) from lysates of infections with each ofthree HSV-1 ts gB mutants, ts B5 (HFEM), ts F13 (KOS), and ts J12 (KOS) (Table 4).
Im-munoprecipitations with group (B3) and group IV (B6)
antibodies werealsocarried outforcomparative purposes. EachofthetsgBmutantsreacted with group II and group IIIantibodies inasimilar fashion. At34°C, both pgBandgB
wereimmunoprecipitated,demonstratingthe presenceofthe groupIIand group IIIepitopesontheseproteins. Strikingly,
the pgB produced by these three mutants at 39°C was not
precipitated by these antibodies. That is, the group II and group III epitopes were not present at high temperature. This effect of temperature is limitedtothe group II and III
epitopes;the group Iand IVepitopesappeartobeunaffected
by high temperature. Monoclonal antibody B3 ofgroup I
immunoprecipitated the gB polypeptides produced at low
and high temperatures from all three ts mutant infections.
AntibodyB6of groupIValsoprecipitated the gB molecules produced in low- andhigh-temperature infections with mu-tants tsF13 and ts J12. The B6 antibody failed to
immuno-precipitate any form of gB produced in ts B5 infections,
indicatingthat thismutantlacks the B6epitopeinadditionto itstsphenotype. Theseobservationssuggest thatthegenetic alteration leadingtothe tsphenotype of these mutantsalso results in alterationof thegBpolypeptide formedatelevated
temperature,whichaffects recognition ofthe groupIIandIII epitopes bythe corresponding monoclonal antibodies.
Also presented in Table 4 are immunoprecipitation data from infectionswithrepresentativemarmutants.Thesedata
bear on thethermostability ofthegBepitopes just described.
That is, epitope thermostability is related to the altered
processing and epitope structure of gB forms produced at
39°Crather thantoepitope denaturationathightemperature.
AntibodyB3 (groupI)immunoprecipitated pgBandgB from
mar B5.1, mar B4.1, and mar B6.1 infections at
both
temperatures. ThemarB3.1mutationaltered theB3 epitope inthatantibodyB3 failedtoimmunoprecipitate anyform
of
gB produced by mar B3.1 infections. While the mar B6.1
mutation rendered themutantvirions resistantto neutraliza-tionbyantibody B6(groupIV),gBwasweakly
precipitated
at eithertemperature from mar B6.1-infected cell extracts. Asexpected, B6immunoprecipitated pgBand
gB
(at34and 39°C) frommarB3.1-,marB5.1-,andmarB4.1-infected cellextracts.These dataindicatethattheB3andB6epitopesare notthermolabile.
Both epitopes II and III are made thermolabile to immu-noprecipitation by the mar B4.1 mutation. Antibodies
of
group II (B5) and group III (B4) were unable to precipitate
pgB produced at 39°C in mar B4.1 infections. In contrast, pgBproduced at39°C inmarB5.1infections appears to be thermostable in that it is precipitated by antibody B4 of
group III. That is, epitope III of this protein can still be recognizedby the corresponding antibody athigh tempera-ture.ThemarB4.1gBphenotype mimicksthegBphenotype ofthetsmutantsdescribed above. Itshouldbe
remembered
that theseantigenicvariants behaveassinglemar mutantsin
virusneutralization assays. Takentogether, epitopes II and
III appeartodependonconformation for theirintegrity and
may reside in a domain susceptible to
temperature-depen-dent alteration.
Characterization ofrevertants of ts mar B2/4.1. If the ts phenotype isaconsequence of themutations producingthe marphenotype,itwould beexpectedthatreversionofthets defect would correlate with coreversion to reactivity with
monoclonalantibodyB2 orB4. Toexamine thispossibility, potentialrevertants of the ts phenotype were
generated
bytwo passages at 39°C. Twenty-four isolates were plaque purifiedand testedforreactivitywithmonoclonal
antibodies
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.53.553.94.227.2]B2 and B4 in animmunoperoxidase plaque assay (16). All 24 isolates regained reactivity with antibody B2 but remained unreactive withantibody B4, asshown with a representative isolate, R3 mar B4.1 (Fig. 7). Thedoublemutant mar B2/4.1 failed to react witheitherantibody (data notshown). When tested, R3 mar B4.1wasfound to have lostthe ts phenotype for both plating efficiency and virusproduction (Table 3).
R3 mar B4.1 was also tested for quantitative reactivity withmonoclonal antibodiesB2 and B4 in 50% plaque reduc-tion assays. R3 mar B4.1remainedresistant to antibody B4, but regained sensitivity to neutralization by antibody B2. The parental double-mar mutantremained resistant to
neu-tralization by both antibodies (Fig. 3). These data indicate that in revertantsrepresented byR3 mar B4.1, reversionof the tsphenotypewasassociatedwithre-establishmentofthe
epitopedefined by antibody B2.
Since the double mutantmar B2/4.1exhibitedtsdefectsin theproductionof gB,R3 mar B4.1 wasfurther characterized forprocessingof pgB to thematureform ofgB at39°C. As
detected by immunoprecipitation, R3 mar B4.1 partially
regainedthe ability toprocesspgB to gB at levelssimilarto the single mar mutant parent mar B4.1, but less than
wild-typevirus (Fig. SB).This result was expected since the
revertant retainedtheB4 mutation which confers apartialts defect in gBprocessing (Fig. 5A).
Inadditiontoprocessingof gB, therevertantR3 marB4.1 also partially regained cell surface expression of the glyco-proteinasdetermined byquantitative enzyme immunoassay
of fixed infected cells withantibody B3 (Fig. 6C and D). At
34°C, R3marB4.1-infectedcellsexpressedcellsurface gBin
amountsequal towild-type virus or mar B4.1. However,at 39°C, R3 mar B4.1 retained expression of cell surface gB, but inintermediateamounts similar to the amountexpressed
by the single mutant mar B4.1. Thus, concomitant with
reversion of the ts phenotype, R3 mar B4.1 regained reac-tivity withantibody B2 butnot B4 andregainedthe
process-ing and cell surface expression characteristics of the mar B4.1 parent.
DISCUSSION
The purposes of our studies of HSV-1 (KOS-321) gB were (i) to define the antigenic structure ofthis antigenand(ii) to
determine whetherantigenic variation in gB canaffect virus production in cell culture. To carry out these analyses, monoclonal antibodies were used to select a series of vari-ants, marmutants, altered intheantigenicstructureof gB.A panel ofgB-reactive monoclonal antibodies representing six distinct clonotypeswas used to selectvariants. Inthis way, we hoped to obtain mar mutants which sustained mutations
in different sites within theregion ofthe gB structural gene that encodes the external glycoprotein domain. Virus-neutralizing monoclonal antibodies were employed in mar mutant selectionsbecause oftherelative ease with which the mutants are isolated and because loss inepitope expression
can be quantitated by changes in antibody titer against the mutant compared with the titer against the wild-type virus.
Although our minimum criterion was a 32-fold reduction in
relative antibody titer (15), most selecting antibodies
com-pletely failed to neutralize the corresponding mar mutants. Thus, we were assured of analyzing mutants which were altered in an amino acid critical to recognition of gB by the
selecting monoclonal antibody.
Many of thegB-specificmonoclonal antibodiesrecognized epitopeswhich are present in both virusserotypes, suggest-ing that the antigenic makeup of HSV-1 and HSV-2 gB is
mAb B2
mAb 84
[image:9.612.329.564.71.324.2]IMMLUNOPEROXIDASE CRYSTAL VIOLET
FIG. 7. Reactivityof arevertantvirus with monoclonal antibod-ies in the immunoperoxidase black plaque assay. The ts revertant R3 mar B4.1 was tested for its ability to react with either of two monoclonal antibodies (B2 and B4) in an immunoperoxidase assay. Virus was plated on Vero cell monolayers and glutaraldehyde fixed when plaques developed. Duplicate plates were then incubated with monoclonal antibody B2 or B4, followed by incubation with goat anti-mouse horseradish peroxidase-conjugated antibody. After ad-dition of substrate (4-chloro-1-naphthol), plaques showing black precipitate were scored as positive, and plates were counterstained with crystal violet.
highly conserved. B4 was the only antibody that neutralized HSV without complement. These observations may be con-trasted with ourstudies of gC-reactive monoclonal antibod-ies. Of more than 30 hybridomas producinganti-gC-1 (23) or anti-gC-2 monoclonal antibodies (J. C. Glorioso and M. Levine, unpublished data), none cross-reacted with gC of the other virus serotype. Moreover, none of the gC antibod-ies neutralized virus in acomplement-independent manner. Based on the analysis of the reactivity patterns of the gB mar mutants with a panel of 16different gB-reactive mono-clonal antibodies, four distinct epitopes were identified. The remaining antibodies, which neutralized all single-mar mu-tants, comprised a separate grouprepresenting at least one additional epitope. One epitope, group II, was somewhat more immunodominant than the rest. Five antibodies were shown torecognize thisepitope, includingthoseproducedin adifferent laboratory and against a different strain of HSV-1 (33). In contrast to our findings with gC mar mutants (23), none of the epitopes in gB gave overlapping reactivity patterns, suggesting that the antibodies bind to different parts of the gBmolecule. This suggestion iscompatiblewith the finding that multiple mar mutations could be readily introduced, one at a time, into the gB structural gene by recombination and sequential antibody selection. The addi-tion of each new mutation had a simple additive effect, resulting in resistance to neutralization only by the corre-spondingselecting antibody. No unexpected multiple resist-ancephenotypes were found. Monoclonal antibody compe-tition studies are in progress to determine whether the
on November 10, 2019 by guest
http://jvi.asm.org/
antibodies indeed bind to topographically distinct antigenic
domains. Giventhe size of the external domain ofgB (-700 amino acids), it is likely that this molecule is more
antigen-ically complex than our analyses have shown. With the
isolation of additional mar mutants selected with new
anti-bodies,amorecompleteandpossiblymorecomplex picture oftheantigenic structureofgB may emerge.
Oneapproachtodetermine whetheraparticularviralgene product is essential for virus growth is to search for ts mutants. Such mutants altered in gB have now been
de-scribed byanumberoflaboratories(7, 13, 22, 30), and some
ofthese mutations have been shown to result from single amino acid substitutions in the external domain of this
antigen (7). As our laboratory (Dorney et al., unpublished data) and others (28) haveshown that marmutations ingB also can result from single amino acid changes, we were
intrigued by thepossibility that marmutations ingB might affect otheraspects of the biology ofthisprotein, including alterations in its processing and function at high tempera-ture. While none of the single marmutants wasaffected in
virusgrowth,adoublemarmutant, marB2/4.1, anditstriple and quadruple mar mutant derivatives all showed reduced plating efficiency and virusyieldat39°Cto adegree
compa-rabletothe other ts mutantsaffectedin gB whichwere not
selected with antibodies.
All ts mutants ofgB are defective in processing of the
major precursor ofgB to the mature gB form (13, 22, 30). Indeed, all of these mutants accumulate a pgB species of
similar molecularsize (13, 22, 30), which led us toexamine both single and multiplemar mutantsfordefectsin
process-ing of gB at high temperature. Several mar mutants, mar
B2.1, marB5.1, and marB4.1, were
partially
defective for processing of pgB to gB at 39°C, despite the finding that these mutants undergo productive infection at elevated temperature. Recombinantscarrying
two ofthese marmu-tationswerehighly defective forthis
processing
step,aswell asbeing defective for theproduction of activevirusparticles
(39°C). Furthermore, at hightemperature the HSV-1 ts gB
mutants, ts F13, ts B5, and ts J12, all accumulated pgB which wassimilar in size to that accumulated bythedouble mar mutant mar B2/4.1. Revertants ofthe ts phenotype of
marB2/4.1regainedthe B2 but not the B4epitope,
suggest-ing that the mar mutations, rather than a second-site ts
mutation, were responsible for the altered processing and temperature sensitivity. The combination oftwomutational changes in the doublemarB2/4.1mutantmoleculemay have
altered the
folding
ofthe precursormoleculemoreseverelyat the nonpermissive temperature, resulting in complete
arrest ofGolgi processing and transport to the cell surface, while atthe sametime producing apgBform which fails to supportvirusproduction.Thesefindingsshow thatantigenic variation in gB can result in changes not only in epitope
configuration, but perhaps moreimportantly, ininactivation ofthefunction of thisantigen invirus replication.
One of severalexplanations could account forthe failure
to produce active double mar B2/4.1 mutant virus at high
temperature. These include failure to produce enveloped particles, production of enveloped particles lacking gB or
pgB, or production of enveloped particles containing nonfunctional pgB. The last ofthese seems most likely for several reasons. First, the underglycosylated precursor
form, pgB, is found in virionenvelopes (34) and may be as
functionally active as the maturegB species. Second, sev-eral studies have shown that monensin or ammonium chlo-ride treatmentof HSV-infected cells results in thedisruption
of Golgi function and the addition of complex
oligosac-charides to the HSV glycoproteins. Although under these
conditions the yield of progeny virus is greatly reduced, these particles are nonetheless infectious (17, 19). Appar-ently, the failure to add terminal sugar residues tothe gB polypeptideisnotsufficienttoinactivate the function of this molecule. Third,revertants of themutant ts B5, affected in
gB, regain the ability to form plaques and produce active
virus at high temperature despite reduced formation of
mature gB (14). Revertants ofmarB2/4.1 showeda similar
phenotype. These data further suggest that processing of pgB togB is not coupledwith ts virus production.
Regard-less ofthe mechanism underlying the ts phenotype ofmar
B2/4.1, processing of pgB togBappearstobe linkedto,but notstrictly correlated with,the
expression
ofafunctionalgBmolecule.
Onequestion arising fromthese studies iswhetherts mar mutantscarrymutations whichmaptothe sameregionas ts
gBmutantsselectedonthebasis oftsgrowth. Althoughdata are not yet availableto answer this
question satisfactorily,
we have determinedbymarkerrescueexperimentsthatthe marmutationsresultingin thetsphenotype of gB
physically
maptothe samegeneral region ofthegB structural geneas the other ts gB mutations
(Highlander, unpublished data).
The pgB form produced by the ts gB mutants at 34°C is
immunoprecipitable by group I (B3), group II (B5), and groupIII(B4)
antibodies, demonstrating
thattheseepitopes
are present in these strains. The HSV-1 HFEM strain is
apparently a natural
antigenic
variant since the group IV(B6) antibody failed to immunoprecipitate the gB of this
virus. At 39°C, the B3 antibody immunoprecipitated pgB from all threetsgB mutants and B6reacted with pgB ofts F13and ts J12. In contrast,antibodies ofgroup II
(B5)
andgroup III (B4) failed to immunoprecipitate pgB (made at
39°C)
despite
thefact that thesets mutants were notselected with monoclonal antibodies, but rather on the basis of ts virus production. Therefore, group II and III antibodies recognizeadomain ofgBwhich is alteredat39°Cin several virus strains while other antigenic domains in the same molecule remain intact. Thiswasillustratedbythecontinued reactivityofpgBat39°Cwith group I and group IVantibod-ies. Theepitopes
recognized by
group II and III antibodiesappear to lie in a thermolabile
portion
ofthe gB moleculecritical forproper
gB
processing
and virusreplication.
Thefinding
that the group IIIantibody recognized
anepitope
conserved in thetwoserotypes and neutralizedvirus in the absence of
complement
also suggests that this domain is criticaltovirusinfectivity.
Thethermostability
ofthe group I and group IV epitopes suggests either that they may be linear determinantsorthat thesemarmutationsdonotaffectportions
of the molecule which lead totemperature-dependent alterations of theseepitopes.
Inaddition to the nature of the tsphenotype of
multiple
marB mutants, several other issuesarise fromourstudies of gB. Whilefurther
analysis
isrequiredtodetermine whether thefivegBepitopes
identifiedinthisstudy
arerepresentative
of the
major
antigenic site(s)
for thisantigen,
our data do allow us to assess whether thesechanges
aresignificant
interms of immune recognition of the variant viruses by polyclonal antibodies.
First, the
antigenic
complexity of the HSV envelope in itself suggests that smallchangesintheantigenicstructureof the virus will not dramatically affect immune control of infection. As demonstrated in the presentstudy,alteration of four distinct epitopes ofgB had no measurable effect onsusceptibility to neutralization by polyclonal antisera di-rected against the wild-type virus. Second, particular
on November 10, 2019 by guest
http://jvi.asm.org/
genic variants may be selected against in natural infections. Based on the results with mar B2/4.1, certain immune-selected mutations may result in an inactive form of this glycoprotein in the natural host. Thus, certain regions of the gB structural gene are likely to be conserved in natural infections. This hypothesis is supported by the following observations: (i) preliminary examination of 40 primary virus isolates from humans has revealed that the group III (B4) epitope is present in all strains, whereas other epitopes of gB are highly variant and double mar B mutants have not been detected (G. Toth and J. Glorioso, unpublished data); and (ii) the ability of mar B2/4.1 to cause encephalitis in mice after intracranial inoculation is greatly inhibited as a result of these mutations (21). It is nevertheless clear that antigenic variation in the HSV glycoproteins is common (11, 29). Studies of antigenic variation in influenza virus have identi-fied mutations responsible for widespreaddisease outbreaks, suggesting that antigenic variation driven by antibody selec-tion can be epidemiologically significant(38). It isunclear at this time whether antigenic variation of HSV can lead to changes in the immunobiology of natural infections.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants AI-16216, AI-17900, AI-18228, and RR00200 from the National Institutes of Health. S.D.M. was supported by a Horace Rackham Predoctoral Fellowship from the University of Michigan, and both S.D.M. and S.L.H. received support from the University of Mich-igan Graduate Program in Cellular and Molecular Biology (Public Health Service Training Grant 2T32GM07315). We also acknowl-edge the support of T.C.H. by the Dental Research Institute Associate Program from the Office of Vice President for Research. We thank Elizabeth Smileyfor herexcellent technical work with monoclonal antibodies.
LITERATURE CITED
1. Balachandran, N.,S. Bacchetti, and W. E. Rawls. 1982. Protec-tion against lethal challenge ofBALB/c mice by passive transfer of monoclonal antibodies to fiveglycoproteins ofherpes simplex virus type 2. Infect. Immun. 37:1132-1137.
2. Bishop, G. A., J. Glorioso, and S. A. Schwartz. 1983. Relation-ship between expression ofherpessimplex virus glycoproteins and susceptibility of target cells to humannaturalkiller activity. J. Exp. Med. 157:1544-1561.
3. Buckmaster, E. A., U. Gompels, and A. Minson. 1984. Charac-terization and physical mapping of an HSV-1 glycoprotein of approximately 115 x 103 molecular weight. Virology 139: 408-413.
4. Carter, V. C., P. A. Schaffer, and S. S. Tevethia. 1981. The involvement of herpes simplex virus glycoproteins in cell-mediated immunity. J. Immunol. 126:1655-1660.
5. Chan, W. L. 1983. Protective immunization of mice with spe-cific HSV-1glycoproteins. Immunology 49:343-352.
6. Courtney, R. J. 1984. Virus-specific components of herpes simplex virus involved in the immune response, p. 34 44. In B. T. Rouse and C. Lopez (ed.), Immunobiology of herpes simplex virus infection. CRC Press, Inc. Boca Raton, Fla. 7. DeLuca, N., D. J. Bzik, V. C. Bond, S.Person, and W. Snipes.
1982. Nucleotide sequences of herpes simplex type 1 (HSV-1) affecting virusentry, cell fusion, andproduction of glycoprotein gB (VP7). Virology 122:411-432.
8. Dietzschald, B., R. Eisenberg, M. Ponce de Leon, E. Golub, F. Hudeca, A. Varrichio, and G. Cohen. 1984. Fine structure analysis of type-specific and type-common antigenic sites of herpes simplex virusglycoprotein D. J. Virol. 52:431-435. 9. Dix, R. D., L. Pereira, and J. R. Baringer. 1981. Use of
monoclonal antibody directed against herpes simplex virus glycoproteins to protect mice against acute virus-induced neu-rological disease. Infect. Immun. 34:192-199.
10. Eisenberg,R.J., D.Long, M. Ponce de Leon, J. T.Matthews, P.G.Spear,M. G. Gibson, L. A. Lasky, P. Berman, E.Golub, and G. H. Cohen. 1985. Localization of epitopes of herpes simplexvirus type1glycoprotein D. J. Virol. 53:634-644. 11. Frankel, M.E., W. Gerhard, Z. Wroblewska, N. W. Fraser, and
H. Koprowski. 1985. Possible relationship between antigenic properties and isolation history of HSV-1 stains. Virus Res. 3:323-337.
12. Glorioso, J.C., and M. Levine. 1985. Monoclonalantibodiesand herpes simplex virus infections, p. 235-260. In S. Feroneand M. P. Dierich (ed.), Handbook on the use of monoclonal antibodies in biology and medicine. Noyes Publications, Park Ridge, N.J.
13. Glorioso, J. C., C. H.Schroder,G.Kumel, M. Szczesiul, and M. Levine.1984.Immunogenicity of herpes simplex virus glycopro-teins gC and gB andtheir role in protective immunity. J.Virol. 50:805-812.
14. Haffey,M.L.,and P.G. Spear. 1980. Alterations inglycoprotein gB specified by mutants and their partial revertants in herpes simplex virus type 1 and relationship to other mutant pheno-types. J. Virol. 35:114-128.
15. Holland, T. C.,S. D. Marlin, M. Levine, and J. Glorioso. 1983. Antigenic variants of herpes simplex virus selected with glyco-protein-specific monoclonal antibodies. J. Virol. 45:672-682. 16. Holland, T. C., R. M. Sandri-Goldin, L. E. Holland, S. D.
Marlin, M. Levine, and J. C. Glorioso. 1983. Physicalmapping of the mutation inanantigenic variant of herpes simplex virus type 1 by use of an immunoreactive plaque assay. J. Virol. 46:649-652.
17. Johnson, D.C., and P. G. Spear. 1982. Monensin inhibits the processing of herpes simplex virus glycoproteins, their trans-porttothe cell surface, and the egress of virions frominfected cells.J. Virol.43:1102-1112.
18. Johnson, D. C., and P. G. Spear. 1983. 0-linked oligosac-charides are acquired by herpes simplex virus glycoproteins in theGolgi apparatus. Cell 32:987-997.
19. Kousoulas, K. G., D. J. Bzik, N. DeLuca, and S. Person. 1983. The effect of ammonium chloride and tunicamycin on the glycoprotein content and infectivity of herpes simplex virus type 1. Virology 125:468-474.
20. Kousoulas, K. G., P. E. Pellett, L. Pereira, and B. Roizman. 1984. Mutations affecting conformationorsequence of neutral-izing epitopes identified by reactivity of viable plaques segre-gatedfrom syn and tsdomainsofHSV-1(F) gB gene.Virology 135:379-394.
21. Kumel, G., H. C. Kaerner, M. Levine, C. H. Schroder, and J. C. Glorioso. 1985. Passive immune protection by herpes simplex virus-specific monoclonal antibodiesandmonoclonal antibody-resistant mutants altered in pathogenicity. J.Virol. 56:930-937. 22. Little, S. P., J. T. Jofre, R. J. Courtney, and P. A. Schaffer. 1981. A virion-associated glycoprotein essential for infectivity of herpes simplex virus type 1. Virology 115:149-160. 23. Marlin, S. D., T. C. Holland, M. Levine, and J. C. Glorioso.
1985. Epitopes of herpes simplex virus 1 glycoprotein gC are clustered intwodistinct antigenic sites. J. Virol. 53:128-136. 24. Nash, A. A., K.-N. Leung, and P. Wildy. 1985. The
T-cell-mediated immune responseofmice toherpes simplexvirus,p. 87-100. In B. Roizman (ed.), The herpesvirus, vol. 4. Plenum Publishing Corp., NewYork.
25. Norrild, B. 1985. Humoral response to herpes simplex virus infections, p. 69-82. In B.Roizman (ed.), Theherpesvirus, vol. 4. Plenum Publishing Corp., New York.
26. Oakes, J. E., W. B. Davis, J. A. Taylor, and W. A. Weppner. 1980. Lymphocyte reactivity contributes to protection con-ferred by specific antibody passively transferred to herpes simplex virus-infected mice. Infect. Immun. 29:642-649. 27. Oakes, J. E., and H. Rosemond-Hornbeak. 1978.
Antibody-mediated recovery from subcutaneous herpes simplex virus type 2infection. Infect. Immun. 21:489-495.
28. Pellett, P. E., K. G. Kousoulas, L. Pereira, and B. Roizman. 1985. Anatomy of the herpes simplex virus 1strainF glycopro-tein B gene: primary sequence and predicted protein structure of the wild type and ofmonoclonal antibody-resistant mutants.
on November 10, 2019 by guest
http://jvi.asm.org/
J. Virol.52:243-253.
29. Pereira, L., D. V. Dondero, D. Gallpo, V. Devlin, and J. D. Woodie. 1982. Serological analysis of herpes simplexvirustypes
1and 2withmonoclonalantibodies. Infect.Immun. 35:363-367. 30. Sarmiento, M., M. Haffey, and P. G. Spear. 1979. Membrane proteins specified by herpes simplex viruses. III. Role of
glycoprotein VP7(B2) in virion infectivity. J. Virol. 29:1149-1158.
31. Schrier,R. D., L. I.Pizer, andJ. W. Moorhead. 1983.
Type-specific delayed hypersensitivity andprotective immunity in-ducedbyisolatedherpes simplexvirusglycoproteins. J. Immu-nol. 130:1413-1420.
32. Sethi, K. K., Y. Omata, and E. E. Schneweis. 1983.Protection of mice from fatal herpes simplex virustype 1infection by adop-tive transfer of cloned virus-specific and H-2-restricted
cyto-toxicTlymphocytes. J. Gen.Virol. 64:443-447.
33. Showalter,S. D., M.Zweig,and B.Hampar. 1981. Monoclonal antibodiestoherpes simplexvirustype1proteins, includingthe immediate early protein ICP4.Infect. Immun. 34:684-692. 34. Spear, P. G. 1976. Membrane proteins specified by herpes
simplex viruses.I.Identification of four glycoproteinprecursors andtheir products intype1-infected cells.J.Virol. 17:991-1008. 35. Spear, P. G. 1985. Antigenic structure of herpes simplex
vi-ruses, p. 435-445. In M.H. V. van Regenmortal and A. R.
Neurath (ed.), Immunochemistry of viruses. The basis for serodiagnosis andvaccines. Elsevier, Amsterdam.
36. Spear, P. G. 1985. Theherpesvirus glycoproteins,p.315-347. In B. Roizman(ed.),The herpesvirus, vol. 3. Plenum Publishing Corp., NewYork.
37. Wenski, E. A., M. W. Bratton, and R. J. Courtney. 1982.
Endo-o-N-acetylglucosaminidase
Hsensitivity ofprecursorstoherpes simplexvirustype1glycoproteins gB and gC. J. Virol. 44:241-248.
38. Yewdell, J. W., and W. Gerhard. 1981.Antigenic characteriza-tion of viruses bymonoclonal antibodies. Annu. Rev. Micro-biol. 35:185-206.
39. Zweerink, H. J., D.Martinez, R. J. Lynch, and L. W. Stanton. 1981.Immune responsesin miceagainst herpes simplexvirus: mechanisms ofprotection against facial and ganglionic infec-tions. Infect. Immun. 31:267-275.