JOURNAL OFVIROLOGY,Oct. 1967,p.956-962 Copyright © 1967 American Society for Microbiology
Temperature-sensitive
Mutants
of Sindbis
Virus:
Biochemical
Correlates of
Complementation
BOYCE W. BURGE' AND E. R. PFEFFERKORN2
Department of Bacteriology andImmunology, Harvard Medical School,Boston, Massachusetts 02115
Receivedfor publication5June1967
Temperature-sensitive mutants of Sindbis virus fail to grow at a temperature
that permits growth of the wild type, but when certain pairs of these mutants,
mixed together, infect cells at that temperature, viral growth (i.e., complementa-tion) occurs. Theyield from this complementation, however, is of thesameorder ofmagnitudeastheinfectivity in the inoculum. Since in animal virus infections the proteincomponentsof the virionprobably enterthecell with the viral nucleic acid, itwas necessary to demonstrate that the observed complementation required syn-thesis ofnewviral protein and nucleic acid rather thansomesortofrearrangement ofthe structuralcomponentsofthe inoculum. To demonstrate thatcomplementation does require new biosynthesis, three biochemical events of normal virus growth have been observed during complementation and correlated with the efficiency of viral growthseenincomplementation. Theseeventsinclude: (i) entranceof parental viral ribonucleic acid (RNA) intoa double-stranded form; (ii) subsequent synthesis ofviral RNA; and (iii) synthesis and subsequent incorporation of viral protein(s) intocell membranes where theyweredetected byhemadsorption. Although the in-fecting single-stranded RNA genome ofthe wild type was converted to a ribo-nuclease-resistant form, thegenomeofamutant(ts-11) incapable ofRNAsynthesis
at a nonpermissive temperature was not so converted. However, during comple-mentation with anothermutantalso defective in viral RNAsynthesis, someof the RNA ofmutant ts-1 was converted to a ribonuclease-resistant form, and total synthesis of virus-specific RNA wasmarkedly enhanced. The virus-specific altera-tion ofthe cell surface, detected byhemadsorption, was also extensively increased duringcomplementation. These observations support theview that complementa-tion between temperature-sensitive mutants and replication of wild-type virusare similarprocesses.
We have previously described complementa-tion between certain pairs of temperature-sensitive (ts) mutants ofSindbis virus (1, 2). In thosereports, complementationwasmeasured by comparing the yield produced by cells infected
by two mutants together, at a nonpermissive
temperature,withthesumoftheyields produced
by thesame twomutants grown separately. Under ideal circumstances, a complementing pair ofmutantsmightproduce 100 timesasmuch virus as either mutant grown separately, a complementation yield representing 1 to 10% of
the titer produced by wild-type virus grown under similar conditions. However, since a high multiplicity of infection was required for
suc-1Present address: Department of Biochemistry,
Albert EinsteinCollegeofMedicine,Bronx, N.Y. 2Present address: Department of Microbiology,
DartmouthMedicalSchool,Hanover,N.H. 03755.
cessful complementation (2), the inoculum used to produce the mixed infection was usually of the same order of magnitude as the comple-mentation yield.
Thus, it could be argued thatcomplementation between ts mutants occurred wholly or partly through a reshuffling of viral components
introduced into the cell by the inoculum. This explanation could theoretically be tested by determining whether labeled precursors were incorporated into the virus released as a result ofcomplementation. However, the lowyields of virus in complementation experiments would complicateitspurification.
We have therefore used several biochemical methods to examine the biosynthetic conse-quences of complementation. (As in the usual complementationtest, these biochemical methods were used to compare cells infected at a non-permissivetemperature by single ts mutants and
956
Vol. 1, No. 5 Printed in U.S.A.
on November 11, 2019 by guest
http://jvi.asm.org/
COMPLEMENTATION IN MUTANTS OF SINDBIS VIRUS
by pairs of ts mutants.) Thefollowing important events in viral replication were studied: (i) entrance of the parental viral ribonucleic acid (RNA) into a double-stranded form; (ii) subse-quentsynthesis of viral RNA; and (iii) synthesis andsubsequent incorporationof viral protein(s) into the cellular membrane where they were detected by hemadsorption. In each case, the magnitude of these biochemical changes was compared withtheefficiencyofcomplementation asmeasured bytheusual viraltitrations.
MATERIALS AND METHODS
Tissueculture and viruses. Methods for thegrowth
andtitration of Sindbis virus inprimarychickembryo
fibroblast cultureshave beenalready described (10).
The ts mutants were isolatedand characterized as
described by Burge and Pfefferkorn (1, 2). These ts
mutants growwellat30C,apermissivetemperature,
butfailtogrowat40C,anonpermissivetemperature.
All ts mutants fall into one of twomajor categories:
(i) those defective in viral RNAsynthesisat
nonper-missive temperatures (RNA-), and (ii) those ableto makeviralRNA butdefectiveinsomeother matura-tional step at nonpermissive temperatures (RNA+). All experiments describedhere usedRNA- mutants.
When a comparison with the wild-type virus was
required, weusedthe HR strain ofSindbis virus (1) from which the ts mutants had been derived. This strain has notsmutations and willbe simplycalled
thewildtypeinthisreport.
Complementation.Thiswascarriedout asdescribed previously (2). Briefly, to test two mutants for the ability to complement, stocks of both mutants were
used to infect doubly a monolayer with about 20 plaque-forming units (PFU) of each per cell. This monolayer wasincubated at 40 C, and the yield of virus producedbetween the 4th andthe6thhrof in-fectionwascomparedwiththe sumoftheyields pro-duced by the two mutants in single infections. The ratios ofthesetwomeasurementsis calledthe comple-mentationlevel; forapair ofcomplementarymutants this level may be from three to 100, depending on several variables, including multiplicity ofinfection, incubation temperature, and the mutant pair under study (2).
Actinomycin D-resistant 3H-uridine incorporation.
Measurementwasin aconventionalway. Cellswere
exposed to 1 gg ofactinomycin D per ml (Merck, Sharp and Dohme Research Laboratories, Rahway, N.J.) throughout theinfection. At a specified time,
freshprewarmedmediumcontaining actinomycinand
3H-uridine (20 c/mmole, 0.03 mc/ml; Schwarz Bio
Research Inc., Orangeburg, N.Y.) was added; 2 hr
later the monolayers werechilled, rinsed twice with
phosphate-buffered saline (4) lacking calcium and
magnesiumandcontaining0.001 M
ethylenediamine-tetraacetic acid(EDTA),anddissolved at room
tem-perature in 2 mlof thesame solutioncontaining 1%
sodiumdodecyl sulfate (SDS).Asampleof this
solu-tion was precipitated with 0.3 Ntrichloroacetic acid
with 200 Ag of yeast RNA as carrier. The rest was
stored frozen at -60 C for later sedimentation analysis.
Sedimentationanalysis. It wascarriedout on15 to
30% lineargradientsof glyceroldissolved in a buffer
containing 0.1 M NaCl, 0.01 M tris(hydroxymethyl)-aminomethane, pH 7.5, 0.001 M EDTA, and 0.2%
SDS. A 0.24-ml sample was layered onto a 4.5-mi
gradientandcentrifugedin a Spinco SW-39 rotor at
38,000 rev/min for 2.75 hr at 25 C. Fractions were collected bypuncturingthetubeandcollecting drops.
All fractions were precipitated with 0.3 N
trichloro-aceticacidwith200 Ag ofyeast RNA ascarrier,
col-lected on membrane filters (Millipore Corp., Bed-ford,Mass.),rinsed withethyl alcohol,and dissolved in 2 ml ofpolyether scintillation solution for
deter-mination of radioactivity. When a sedimentation
marker wasrequired,asmallamount ofchick
fibro-blast monolayer previously labeled for 3 days with 14C-uridine and dissolved in SDS was added to the
sample. Acorrectionwas made for 14C counts in the
3Hchannel.
Fate of theinfecting viral RNA. This was traced by
use of virus labeled with 3H-uridine (20 c/mmole;
Schwarz Bio Research). To prepare this virus,
cul-tures wereinfected at 28 C in a medium containing
0.25 ugof actinomycin D per mland 125 Muc of 3H-uridine per ml. The low temperature allowed ts
mutantsand the wild type to be labeled under
identi-cal conditions. Thepresence ofactinomycin assured
that most of the 3H-uridine would be incorporated
intovirus-specificRNA.After 16 hr ofincubation,the
virus in the medium was partially purified by one
cycle of differential centrifugation. The resulting preparation was radiochemically homogeneous by several criteriaincluding sucrosegradient centrifuga-tion, resistance of the radioactivity to ribonuclease, andultracentrifugal homogeneity ofthe labeled RNA extracted from thevirus. Thevirus (about 3 X 10-8
countsper min perPFU) wassuspendedin medium
containing 100 ,ugofuridine per ml toeliminate in-corporation of any residual 3H-uridine from the growthmedium.
Thislabeled viruswasallowedtoadsorbto mono-layer culturesfor 1 hr at 4 C. Thecultureswere then rinsed andincubated at the desired temperature. At intervals,thesecultureswereexaminedforconversion
of the infecting viral RNA to a
ribonuclease-resistant form byuseof amodification ofthe method describedby Lodishand Zinder (8). Rinsed cultures weredissolvedin 3.5 ml of asolution containing0.07 MNaCl,0.001 MKCI,0.001 MEDTA,and0.1%SDS.
TheresultingSDS extract wascentrifugedat2,000X
g, and samples of the supernatant fluid, which
con-tainednearlyall of theradioactivity, were diluted
10-fold into water and buffer (8) containing 25 ,ug of
pancreatic ribonuclease perml. Thesesolutions were incubated at room temperature for 1 hr; then the ribonuclease-resistant material was precipitated with
0.3 N trichloroacetic acid, and its radioactivity was
determined inascintillation counter. In each case, the
ribonuclease-resistant radioactivity from the sample
diluted into water was subtracted from the value
ob-tained inbuffer as recommended by Lodish and Zinder
(5). Only a negligible fraction of the infecting viral 957
VOL. 1)1967
on November 11, 2019 by guest
http://jvi.asm.org/
BURGE AND PFEFFERKORN
RNA was ribonuclease-resistant immediately after
adsorption. Under conditions that permitted viral
replication, the infecting viral RNAwaspartially
con-vertedtoaform thatwas ribonuclease-resistant only
athigh salt concentrations and had the sedimentation
properties reported for double-stranded arbovirus
RNA (6, 11). [The characterization of the labeled
virus and details of the intracellular conversion ofits
RNA to a double-stranded form will be presented
elsewhere (Pfefferkorn, Burge, and Coady, Virology,
inpress).]
Hemadsorption. The capacity of monolayer cultures
toadsorb goosered blood cellswasdetermined bya
modification of the hemagglutinationtestfor Sindbis
virus (3). To allow quantitative estimation of
hemad-sorption,the red cells werelabeled with 51Cr asina
modification of the usual hematological procedure
(5). A 7.5% suspension of gander red cells in
bar-biturate buffer containing glucose and gelatin (3)
wasincubated with Na251CrO4 (0.125 Ac/ml) for 1 hr
atroomtemperature.The labeled red cellswerethen
washed twicewith the same buffer and once with a
1:1 mixture oftheborate andpyrophosphate buffers
(final pH 5.8) used in hemagglutination titration (3,
10).The cellswerefinally suspendedata3% concen-tration in the borate-pyrophosphate mixture and used within the next few hours.
Monolayerstobeassayedforhemadsorptionwere
chilledin anice bath,drained and rinsed twicewith
borate-pyrophosphatebuffer. A0.3-mlvolumeof the
6'Cr-labeled red cell suspension (about 7 X 10-4
counts permin per cell) was thenpipetted ontothe
monolayer, which was held at 4 C with occasional
rockingtoredistribute the red cells. After 45min,the
monolayersweredrained and rinsedgently four times
with 2.5-ml volumes of cold borate-pyrophosphate buffer.
The monolayer, together with any adsorbed red
cells,wasdissolved in 1 ml of 0.3NNH40H,added to
10 ml of polyether scintillation solution (7), and
counted inascintillation counter. Quenching bythe
dissolved monolayer was a constant factor in all
samples. Hemoglobin from cultures positive for
hemadsorption did not quench significantly. The
hemadsorption appeared to be a virus-specific
reac-tion, for itwasreducedtothecontrol levelbyprior
treatment of the infected monolayer with rabbit
anti-Sindbis virus serum that had been exhaustively
adsorbedwith uninfected cells.
RESULTS
Complementation between RNA- mutants.
Complementation between the RNA-ts mutant
ts-6 and other RNA- mutants is quite efficient,
in terms of both the complementation level achieved and the absolute titer of virus
pro-duced (9a). Hence, if complementation did
involve an increase in viral RNA and protein synthesis, these increases should be easily
detectable. Table 1 records the results ofa
com-plementationtestinvolvingmutants ts-6, -11 and
-24.In eachcase,thesingleinfectionsbymutants
produced only about 0.01 % the infectivity
TABLE 1. Complementation as measured by infectivitya
Virusproduced Complemen- Percent of Inoculum between 4 and 6 tation Pe eld
hr(PFU/ml) levelb wild-typeyield
Wild-type 1.9 X 1O9 100.0
ts-6 32.0 X 104 0.016
ts-11 2.0 X 104 0.001
ts-24 10.0 X 104 0.005
ts-6 X ts-11 12.0 X 107 353 6.3
ts-6 X ts-24 8.0 X 107 190 4.2
aComplementationwas carried out as described
in Materials and Methods. Data in Tables 1 and
2 were obtained in the same experiment.
bThe complementation level is the titer
pro-ducedby the double infection divided by the sum
ofthe titers produced by the two mutants in single
infections.
produced by the wild type. The complementing
pairs, however, produced about 5% of that yield. These results are similar to those
pre-sented elsewhere (2). With the exception of
the study of the fate ofmutantviral RNA, which
required the use of labeled parental virus, all
of the biochemical assays of complementation
were done with replicate cultures infected and
incubated with those described in Table 1.
Thus, the biochemical results can be directly
comparedwith this bioassay of complementation.
Fate of parental viral RNA during comple-mentation. The replication of a single-stranded
RNA virus probably requires the formation
of a virus-determined enzyme that directs
synthesis of new "minus" strands,
comple-mentary to the viral RNA "plus" strand (8, 9,
12). Newly synthesized "minus" strands are
found together with parental "plus" strands in
a double-stranded structure that can be
iden-tified by its resistance to ribonuclese.
The RNA of wild-type Sindbis virus labeled
with 3H-uridine was partially converted to a
ribonuclease-resistant form very
early
in thecourse ofinfectionsat 30 orat 40 C. This form
had a sedimentation constant of about 20S and
several of the other properties expected for
double-stranded RNA (Pfefferkorn, Burge, and
Coady, in
press).
We attempted to correlate
complementation
with thefate of labeled viral RNA in
single
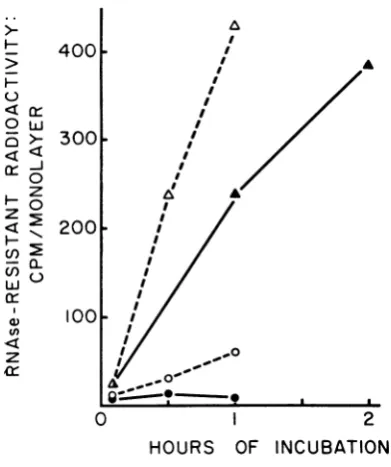
andmixed infections. Figure 1 records the
con-version of labeled
input
RNA of mutant ts-11to aribonuclease-resistantforminsingleinfection
at 30 C and in
single
and mixed infection withmutant ts-6 at 40 C. In
single infections,
con-version of the RNA of mutant ts-11 to a
ribonuclease-resistant form was normal at 30 C
958 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
COMPLEMENTATION IN MUTANTS OF SINDBIS VIRUS
> A
400. /
Ii
>- 300
-<~~~~~~~~~
0
z~~~~~~~~
Z A'
H0\ 200
-a~~~~~~~
oxJ
I
OI 2
a) 100 1/
2
HOURS OF INCUBATION
FIG. 1. Intracellular conversion oflabeledinfecting
viralRNA to a ribonuclease-resistant form early in
thecourseof inifection.Labeledviruswaspreparedand
thefate ofits RNA was traced as described in the
Materials and Methods.Symbols: A,3H-labeled
wild-type infection at 40 C; 0, 3H-labeled mutant ts-1l
infectionat40C;A,3H-labeledmutantts-1i infection
.at 30 C; 0, 3H-labeled mutant ts-11 and unlabeled
mutantts-6 mixedinfectionat40 C.
but completely defectiveat 40 C. Since the wild
typeshowednormal conversion of its RNA toa
ribonuclease-resistant form at 40 C, mutant
ts-11 appears to be defective in this initial step
of viral replication at the nonpermissive
tem-perature. This defect was partially relieved in
the mixed infection with unlabeled mutant ts-6,
another RNA- mutant. The efficiency of
con-version ofmutant viral RNAto a ribonuclease-resistant form, about 5% ofthe value obtained
forthemutant at 30 Cor the wild typeat40C, wascomparabletothe efficiencyof
complementa-tionbetween thesetwomutants(Table 1).
Viral RNA synthesis during complementation. We next sought to correlate complementation with net synthesis of viral RNA. Since actino-mycin D was used to suppress cellular RNA synthesis, incorporation of 3H-uridine was largely into viral-specific RNA. Table 2 shows
thatcultures infected byany of thethree
RNA-mutants at a nonpermissive temperature incor-porated no more 3H-uridine than did the
unin-fected control. This, ofcourse, is the definition
of an RNA- mutant. However, the cultures infected by the mutants together showed a
substantial increase in incorporation of 3H-uridine to values 8 to 10V of the wild-type level. These values are only slightly greater
[image:4.462.26.222.78.312.2]than the complementation measured by virus production (Table 1).
TABLE 2. Complementation as measured by
hemad-sorptionand3H-uridinieincorporationa
Inoculum Hemadsorptionb 3Hiuridineb
Uninfected 103 4.9 X 103
Wild-type 12,481 325.0 X 103
ts-6 112 5.2 X 103
ts-11 99 5.1 X 103
ts-24 125 6.8 X 103
ts-6 X ts-1 1 5,419 25.5 X 103
ts-6 X ts-24 4,680 32.5X 103
aData in Tables 1 and 2 were obtained in the
samecomplementation experiment.
IMeasurement of hemadsorption and 3H-uri-dine incorporation is described inMaterials and Methods.Both areexpressed incountsperminute permonolayer.
Sedimentation properties of the RNA labeled
during complementation. Virus-specific RNA
produced during arbovirus infections is known
to sediment intwo major peaks, at 42S and 26S
(6, 11). Both RNA species are ribonuclease
sensitive. A third, ribonuclease-resistant peak,
sedimentingat20S, may be resolved by digesting
away the ribonuclease-sensitive species (6, 11).
Since the physiological significance of these
RNA's, particularly the 26S species, isnotcertain,
it was of interest to examine the sedimentation
properties of viral RNA produced during
com-plementation between ts RNA-mutants.
Samples of SDS-dissolved monolayers from
the
complementation
described in Tables 1 and 2 were analyzed on linear gradients of glycerol-SDS. Figure 2 shows the pattern seen in the wild-type infection at 40 C. No attemptwasmadeto resolve theminor20S
ribonuclease-resistant fraction.
In Fig. 3, the 3H-uridine incorporated by the
RNA- mutants in single infections (in the presence of actinomycin D) can be compared
to the 3H-uridine incorporated by an uninfected
monolayer. In all samples, most of the labeled
RNA appeared in a slowly sedimenting band of
about 4S. A small fraction of the radioactivity sedimented more rapidly than the 16S marker, but there was no indication of any preferential synthesis of the virus-specific 42 and 265 RNA. Thus, the RNA- mutants in single infections at 40 C stimulate a negligible amount of
virus-specific RNAsynthesis.
959
VOL. 1, 1967
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.462.231.423.161.290.2]BURGE AND PFEFFERKORN
I
-r 111
w
z
w
a-C,
z I:2
0
Z,w
z
300 LiJ
C,)
z
100
o
0
1 5
9
13 17FRACTION -NUMBER
FIG. 2. Sedimentation analysis of 3H-uridine
in-corporated into a monolayer infected with wild-type
virus.A monolayerwasinfected withahigh multiplicity
ofvirus inthepresence of I jugof actinomycinD/ml.
At 4 hr after infection, 3H-uridine (0.03 mc/ml, 20
clmmole)
wasadded and aftera 2-hrlabeling periodthemonolayerwasdissolvedinSDSbuffer. A 0.24-ml
sample with added 14C-labeled chick cell RNA for
sedimentation markers was centrifuged in a Spinco
SW-39 rotor at38,000 rev/min for 2.75 hr at 25 C.
Symbols: 0, 3H-labeled virus-specific RNA; 0,
cellularRNA. Thissample andthoseanalyzedinFig.
3 and 4 were obtainedfrom the complementation
ex-periment recorded in Tables I and 2.
Sedimentation patterns forlabeled RNA from cells infected by complementing pairs of
RNA-mutants appear in Fig. 4. These patterns were
similar to those seen in the wild-type infection; the additional radioactivity incorporated in mixed infections (Table 2) all appeared in the 26S and 42S regions. Proportionately more 42S RNAwas seeninthe mixedinfection ts-6 X
ts-24than inthewild-typeinfection. The signifi-canceof this observation is unknown.
Synthesis ofviralprotein during
complementa-tion. The synthesis ofthe viral protein active in
hemagglutination was measured by determining thecapacity ofthe infected cellstoadsorbgoose
red cells. To obtainquantitative results, weused 51Cr-labeled red cells and measured the radio-activity that could not be washed from the monolayer.
The ability of virus-infected monolayers to adsorb 65Cr-labeledred cellswas a verysensitive indicator ofviral growth. As Fig. 5 shows, this
alteration in cellsurface was detectable at least
1 hr before a linear rate of virus release was
w
z
w
(n z
0
100
50
100 50
100
50
100
50
ts-24
28s 16s
/..
/
..O.
_/
K
Is-lI
.0 .*S ^
ts-6
,~~~
UNINFECTEDI
,.'
1 5 9 13 17 21
FRACTION NUMBER
FIG. 3. Sedimentation analysis of 3H-uridine
in-corporated into an uninfected monolayer and into
monolayers infected with single RNA- ts mutants.
Actinomycin D, I ,ug/ml, was present in all cultures.
Seelegendof Fig. 2 forconditions of sedimentation.
achieved. The number ofred
cells
adsorbed permonolayercontinued to increase until the 4th hr
ofinfection. At this point, the ratio of labeled
red cells to monolayer cells was about 3, and
saturation of the available surface probably
prevented theadsorption of more red cells onto
monolayers incubated for longer times.
On the basis of this kinetic experiment, we
measured hemadsorption at 6 hr after infection
in complementation experiments. Table 2 shows
that cultures infected singly with the mutants
at the nonpermissive temperature had only the
minimal hemadsorbing capacity of uninfected
cultures. In contrast, mixed infection by
com-plementing
pairs of mutants induced ahemad-sorbing capacity that was about
50%
thatinduced by wild-type infection, indicating
ex-tensive synthesis of at least one viral protein
during
complementation.DISCussIoN
In
complementation
betweenbacteriophage
conditional-lethal mutants, the viral coat of the
infecting phage particle remains outside the cell
and could not
possibly
be used in the formation960
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.462.46.248.65.289.2] [image:5.462.235.444.68.340.2]COMPLEMENTATION IN MUTANTS OF SINDBIS VIRUS
40(
w
204
z
L6
a
0
ts-6
x
fs-24
1
5
9
13
17
21
FRACTION
NUMBER
FiG. 4. Sedimentation analysis of 3H-uridine
in-corporated into monolayers treated with actinomycin
D,I ,ug/ml, and infectedinamixedmannerwith pairs
ofts mutants. Seelegend of Fig. 2for conditions of sedimentation.
of new infectious particles. In animal virus
infections, however, all viral components
prob-ably enter the cell, and might conceivably be used in theassemblyofnewparticles. Itis
there-fore important to demonstrate that
comple-mentation with ts mutants of an animal virus
involves new synthetic events and that these
events are similar to, or reconcilable with,
theevents observedin awild-type infection.
In this report, we have shown that
comple-mentation between ts mutants is accompanied
by three biochemical events: (i) an increase in the amount of mutant viral RNA that is con-verted to a ribonuclease-resistant form; (ii) an increase in the synthesis of virus-specific
RNA;and (iii) anincrease in theamountof the
viral hemagglutinating protein in the cell
mem-brane,reflectedbyanincrease inhemadsorption.
Each of these increases is commensurate with,
thoughnotpreciselyproportional to,theincrease
intiter of viralinfectivity.
0Qr
I>-LLOc
m
-Jz
LL O- 7000 00
Z
-'
50000 1.
L-o 3000
0 0
000
<
I0
I,
/
,,/~~~~~~~~
<0S-S * .~~~~~~~
7xlO8
0..
w 5x18>
cn
cr->
u-108
DD
cJ
0 2 4 6
HOURS AFTER INFECTION
FIG. 5. Kinetics ofappearance of hemadsorption
capacity and viral infectivity. A series ofmonolayers
was infected with wild-type Sindbis virus (about 20
PFU/cell) and incubated at 36 C. At intervals, mono-layers were withdrawn; the infectivity in the medium
wasassayed(0) and thehemadsorbingcapacity of the
monolayer(0) was measured asdescribed in Materials
andMethods.
The only serious discrepancy lies in the measureofhemadsorption. Herethecellinfected by the mixed mutants showed about 50% of
the hemadsorbing capacity ofthe cells infected
by the wild-type virus, although the virus yield from mixed infections was only 5% of the
wild-type yield. It must be remembered, however,
that only a finite number of red cells can be adsorbed to a monolayer of fixed surface area, regardless of the concentration of hemadsorbing
protein in cell membranes. Once this saturation
point is reached, the number of red cells
ad-sorbed to the monolayer is no longer propor-tional to the amount of hemadsorbing protein
in the membrane (Fig. 1). In the wild-type
infection here compared to
complementation,
this point has been reached, and thus it is not
possible to determine the relative amounts of
hemadsorbing protein produced in the two
situations.
Using the three criteria listed
above,
wecon-clude that complementation requires new
bio-syntheticactivityand involves thesamesequence
of events thatcharacterizethegrowth of wild-type Sindbis virus.
ACKNOWLEDGMENTS
We aregratefultoJoanDaniels forsupplyinggoose
red blood cells andto ShelbyKashketfor advice on
labelingthemwith Na25CrO4. Helen M. Coady
con-tributed experttechnical assistance.
Thisinvestigationwassupported byPublic Health
Service research grantAI-04531-06from theNational
Institute ofAllergyandInfectious Diseases. Boyce W.
Burge is a Harold C. Ernst Fellow.
VOL. 1, 1 967 961
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.462.27.217.50.391.2] [image:6.462.229.422.73.239.2]BURGE AND PFEFFERKORN
LITERATURE CITED
1. BURGE, B. W., AND E. R. PFEFFERKORN. 1966.
Isolation and characterization of conditional
lethal mutants of Sindbis virus. Virology
30:204-213.
2. BURGE, B. W., AND E. R. PFEFFERKORN. 1966.
Complementation between
temperature-sensi-tive mutants of Sindbis virus.Virology
30:214-223.
3. CLARKE, D. A., ANDJ. CASALS. 1958.Techniques
for hemagglutination and
hemagglutination-inhibition with arthropod borne viruses. Am. J.
Trop. Med. Hygiene 7:561-573.
4. DULBECCO, R., AND M. VOGT. 1954. One step
growth curve of western encephalomyelitis
virus onchick embryocells grown in vitro and
analysis of virus yields from single cells. J. Exptl. Med. 99:183-189.
5. EBAUGH, F. G.,C. P. EMERSON, AND J. F. Ross.
1953. Use of radioactive chromium 51 as an
erythrocytetagging agent for the determination
of red cell survival in vivo. J. Clin. Invest.
32:1260-1276.
6. FRIEDMAN,R.M., H. B.LEVY,ANDW. B. CARTER.
1966. Replication of Semliki Forest virus:
three forms of viral RNAproduced during
in-fection.Proc. Nati. Acad. Sci. U.S. 56:440-446.
7. LAW, J. H., H. ZALKIN, AND T. KANESHIRO.
1963. Transmethylation reactions in bacterial
lipids. Biochim. Biophys. Acta 70:143-151.
8. LODISH,H. F.,ANDN.D. ZINDER. 1966.
Replica-tion of the RNA ofbacteriophage f2. Science
152:372-377.
9. MILLS, D. R., N. R. PACE, AND S. SPIEGELMAN.
1966. The in vitrosynthesis of a noninfectious
complex containing biologically active viral
RNA. Proc. Natl. Acad. Sci. U.S.
56:1778-1785.
9a. PFEFFERKORN, E. R., AND B. W. BURGE. 1967.
Genetics and biochemistry of arbovirus
tem-perature-sensitive mutants. In J. Colter [ed.],
The molecular biology of viruses, Academic
Press, Inc., NewYork. In press.
10. PFEFFERKORN, E. R., AND H. S. HUNTER. 1963.
Purification and partial chemical analysis of Sindbis virus. Virology 20:433-445.
11. SONNABEND, J. A.,E. M. MARTIN,AND E. MECS.
1967. Viral specific RNAs in infected cells.
Nature 213:365-367.
12. WEISSMAN,C.,AND G. FEIX. 1966. Replication of
viral RNA. XI. Synthesis of viral "minus"
strands in vitro. Proc. Natl. Acad. Sci. U.S.
55:1264-1268.
962 J. VIROL.