0022-538X/80/01-0107/08$02.00/0
Temperature-Sensitive
Mutants of
Chandipura Virus
II.
Phenotypic
Characteristics of the Six
Complementation Groups
D. A. GADKARItAND C.R.PRINGLE*
Medical Research CouncilVirology Unit,Instituteof Virology. GlasgowGIl 5JR,Scotland
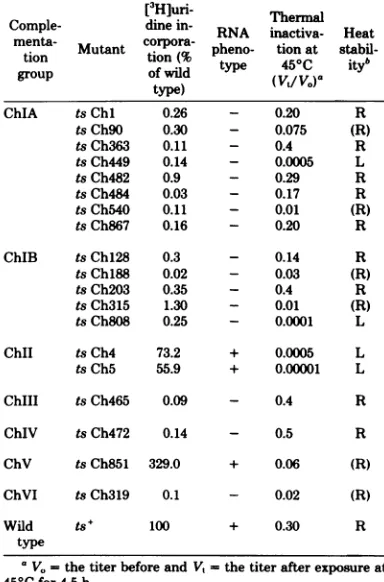
Fifty temperature-sensitive (ts) mutantsof the rhabdovirusChandipuravirus have been classified into six complementation groups designated ChIto ChVI.
GroupChI contains 44 mutants,groupChII contains 2 mutants, and the remaining groupshave 1mutanteach. Mutants ingroupsChI, ChIII,ChIV, and ChVI had
RNA-negative phenotypesinexperiments measuring amplificationofRNA
syn-thesis at restrictive temperature. The two mutants in group ChII had
RNA-positivephenotypes, and the virionswerethermolabile. MutanttsCh851 ofgroup
ChVwasalso RNApositive,and the Mpolypeptideof thismutantappearedto
be unstable in cellsincubated atrestrictive temperature. It is likely, therefore,
thatcomplementationgroups ChIl and ChV represent thegenescodingforthe
twoviralproteinsof the virionenvelope.Noprecise assignment canbe made in
thecaseof the fourRNA-negativegroups,since allthemutantsexaminedshowed
somepolymerase activityinvitroatrestrictivetemperature.Anattempt toobtain
polymerase mutantsby screening for sensitivitytorifampinwasnotsuccessful. Sixtemperature-dependent hostrangemutants (the tdCEphenotype) of
Chan-dipuravirus failedtomultiplyinchickenembryocellsatrestrictivetemperature,
but otherwisetheydifferedintheir hostrangepropertiesfromsimilarmutantsof vesicular stomatitis virus.
Fifty
temperature-sensitive
mutantsof Chan-dipuravirus,
a human rhabdovirus isolated inIndia, have been classified into six
complemen-tation groups.
Group
ChI contains 44mutants, group ChII contains two mutants, and groupsChIII,
ChIV, ChV,
and ChVI have onemutant each. Weakcomplementation
wasobservedbe-tweencertain mutantsin group
ChI,
and itwaspossible
todistinguish
twosubgroups
ChIA and ChIB bytheir interaction with mutant tsChl,
an
arbitrarily
chosen prototype for groupChI
(1).
Wenowdescribe characterization of the
phe-notype of the mutants of groups
ChII, ChIII,
ChIV,
ChV,
and ChVI andarepresentative
sam-ple ofthe mutants ofsubgroups
ChIAandChIB.Theproperties examined includedRNA
pheno-typein
vivo,
RNApolymerase
activity
invitro,
thermal
lability,
and intracellularprotein
syn-thesis. Evidence ispresented
which associatescomplementation
groupChV with theMprotein
transcriptionalunit of thegenome.
The isolationand hostrange of six
tempera-ture-dependent
host range(tdCE)
mutants ofChandipura
virusarealso described.Thesemu-tants of
Chandipura
virus failed tomultiply
in chickenembryo
(CE)
cellsatrestrictivetemper-tPresent address: National Institute ofVirology,Pune 411 001,India.
ature, butotherwise were distinct in their host rangeproperties from previously described tdCE
mutants of the Indiana and New Jersey
sero-types ofvesicular stomatitis virus (VSV) (10).
MATERIALS AND METHODS
Cells. The BS-C-1 line of African green monkey kidneycellswaspropagated inrollerbottlecultures in Eagle minimum essential medium (Glasgow modifi-cation) supplemented with 10% calf serum. BHK-21 clone13cellsweresupplied routinely by the Cytology Department of the Institute of Virology.
Bovine embryo kidney (BEK) cells and hamster embryo (HE)cellswereobtained fromGibco Biocult,
Glasgow. Feline embryo (FEA) cells were obtained from 0. Jarrett, Department of
Veterinary
Pathology, University of Glasgow, and rat embryo (RE) cells weresuppliedby J. Macnab of the Institute of Virology. Mouse embryo (NIH/3T3) cells were obtained from S. A. Aaronson, National Institutes of Health, Be-thesda, U.S.A.
SecondaryCE cell cultures were prepared from eggs obtained fromaleukosis-free flock maintained by the Poultry Research Institute, Edinburgh, Scotland.
Virus.Chandipuraviruswasobtained from F. Mur-phy, Center for Disease Control, Atlanta, Ga., and cloned by three sequential isolations from single plaques on BHK-21 cell monolayers. Isolation and classification of the ts mutants ofChandipura virus have beendescribedinthe preceding paper(1).
The rifampin-sensitive mutant (VSV-29) of VSV Indianaand itswild type (VSV-VJ) were kindly
pro-videdby Ch.Chany, INSERM U-43,H6pitalSt.
Vin-107
on November 10, 2019 by guest
http://jvi.asm.org/
108 GADKARI AND PRINGLE
cent de Paul, Paris, France. The Evans vaccine strain of vaccinia virus was obtained from H. Pennington, Institute of Virology.
Determination of RNAphenotype. BS-C-1 cell monolayers in 30-ml bottles were incubated at 37°C for2hin the presence of 10
fig
of actinomycin D per ml (Cosmegen; Merck Sharp and Dohme, Rahway, N.J.). The cultures were then infected induplicate at a multiplicity of 10 PFU/cell, and the virus was al-lowed to adsorb at +4°C for 30 min. Eagle medium containing10,ugofactinomycinDperml and 10,uCi of [3H]uridine per ml (26 Ci/mmol; Radiochemical Centre, Amersham) was added to each culture bottle, and one set ofcultures was immersed in a water bath at39.5°Cwhile the other set was incubated at 31°C. At 9hafterinfection, 2 ml of a solution of200-yg/ml proteinase K (Boehringen-Mannheim) in 0.01 M Tris-hydrochloride (pH 8.0) containing 0.01 M EDTA, 0.01MNaCl, and 0.2% (wt/vol) sodium dodecyl sulfate was addedtoeachmonolayer, and the cellsweredigested
at 20°C for 18 h. Trichloroacetic acid-precipitable radioactivity was measured for each sample in an Intertechnique SL30 liquid scintillation counter.
Temperature shift experiments. The effect of temperatureshift onthesynthesis of RNA in ts mu-tant-infectedcellswasdeterminedasfollows. BS-C-1 cells in30-mlbottleswereinfectedat amultiplicity of
10approximately, and the cultureswereincubatedat
31 or39.5°C in 2ml of medium containing 10
jg
ofactinomycin D per ml. After3 h of incubation the culture mediumwasreplaced with2mlofprewarmed (31 or39.5°C) medium containing 10,uCi of[3H]uridine
perml and 10
jig
ofactinomycinD perml.Culturespreviouslyincubated at31°Cwere shiftedto 39.5°C
andincubated for6 h atthat temperature. Cultures
previouslyincubatedat39.5°Cwereshiftedto31°C.
Other cultures were maintained at 31 or 39.5°C throughout the experiment. Allcultures were inocu-lated in duplicate and harvested at9h asdescribed above.
Invitropolymeraseassay. Thepolymerase
ac-tivity ofpurified preparationsofwild-type Chandipura
virusand itsts mutantswasassayedat31 and39°C
under theconditionsestablished for VSV Indiana(11).
Itwasdetermined by experiment, however, that the optimum concentration of Triton N-101 in the reaction mixturewas0.08% forChandipuravirus. The reaction mixtureswerewarmed for1minateither31or390C priortoaddition of220 mM MgCl2. Samplesof20
Al
werewithdrawnimmediatelybefore addition ofMgC12(thezero time sample) andafter 30, 60,90, and 120
min at 31 or 39°C, transferred to 2.5-cm2 Whatman
DE81(ionic) paper disks, andairdried. The diskswere
then washedfive times in 5% (wt/vol) sodium pyro-phosphateat20°C, followed bytwowashesin deion-izedwater,ethanol,anddiethylether.Residual ether
wasremovedin a stream ofair, andthe diskswere
immersedovernight intoluene-basedscintillantprior
tocounting (12).
Viruspurification.Purified virionswereobtained by inoculating BHK-21 clone 13 cells inrotating
2-literbottlesat amultiplicityof 0.1. The contents of 10
bottleswerepooledand clarifiedby centrifugationat
1,500 rpmfor 20 min at 40Cin aMistralcentrifuge
(M.S.E. Ltd.,London). Virus in thesupernatantwas
sedimented through a 30% (vol/vol) glycerol layer
using a rotor (10 by 100 ml) at 40,000 x g in a
Superspeed65ultracentrifuge (M.S.E. Ltd.,London)
at4°C. The resuspendedpelletwaslayeredon a 15 to
45%(wt/vol) linearsucrosegradient and centrifuged in aBeckman SW27rotor at50,000xgfor60minat
4°C. Theopalescent bandwascollectedby pipette and pelleted by centrifugationat150,000xgfor30minat
4°C in a Beckman SW40 Ti rotor. The pellet was
resuspendedin asmall volume of20mM
Tris-hydro-chloride (pH 8.0).
Determination of heat stability. Virus stocks were diluted 10-fold into phosphate-buffered saline and distributed into thin-walled glassvials. Heat
ex-posurewascarriedoutby totallyimmersing these vials
in a waterbath at 45± 0.1°C. Sampleswere
trans-ferredto anice bathbeforeheat exposure andat
45-minintervals after immersionat45°C.Residual infec-tivitywasassayedonBS-C-1monolayers.
Analysisof intracellularproteins.BHK-21 cells in 30-mm petri disheswereinfectedat amultiplicity of20PFU/cell. The infected cellswerelabeledat 4.5
h after infection in Eagle medium containing
one-tenth the normal concentration of methionine and30
,uCi of [nS]methionine per ml (5 to 7 mCi/mmol;
Radiochemical Centre, Amersham). The cells were
eitherpulse-labeledfor15minorcontinuouslylabeled for3h.
At 7.5 h after infection the medium from each cultureplatewasremoved,0.2ml ofasolubilization mixture (2%[wt/vol] sodium dodecyl sulfate and 5% [vol/vol] 2-mercaptoethanol in0.14M Tris-hydrochlo-ride [pH 6.7]plus 10%[vol/vol]glycerol and bromo-phenol blue)wasaddeddirectlytothemonolayer, and theplateswereleftat20°C for15min.Before electro-phoresis, the samples were heated at 100°C for 5 min. Trichloroaceticacid-insolubleradioactivitywas meas-ured, and the concentration of the samples was ad-justed sothat each track in anysingle gel slab
con-tainedapproximately equalcounts.
Polyacrylamide gel electrophoresis.
Polyacryl-amidegel electrophoresiswascarriedoutin5 to12.5% gradient slab gels accordingto themethod described by Marsdenetal(5).
RESULTS
RNA phenotype. The RNA phenotype of
the mutants was established as described in Materials and Methods. Eight mutants from subgroup ChIA and five mutants from subgroup ChIBwerechosenatrandomtorepresent group
ChI.
Incorporation
of[3H]uridine
was c1.3%of the wild-type value (Table 1). The group ChI mutants, therefore, were all considered to beRNA-negativemutants.Thethreemutants
rep-resenting groups ChIII, ChIV, and ChVI also exhibited RNA-negative phenotypes, having levels ofincorporation 0.15% ofwildtype. On the other hand, the two mutants belonging to
group ChII showed goodincorporation, 56 and 73% of wild type, and were designated
RNA-positivemutants.MutanttsCh851of groupChV
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
wasalsoanRNA-positivemutantandappeared
to incorporate more [3H]uridine (329%) than
wildtype.
No significant incorporation of [3H]uridine
was observed when mostof the RNA-negative
ts mutant-infected cultures were shifted from
permissivetorestrictive temperatureat3 h after infection. These observations suggested that the input virion proteins,or thosesynthesized
dur-ing the first3hatthe permissivetemperature,
remained nonfunctional atthe restrictive
tem-perature. Cellsinfected with twoRNA-negative
mutants,tsCh2O3 (ChIB) andtsCh472(ChIV), did showsignificant incorporationof[3H]uridine after temperatureup-shift (Table 2).This
incor-poration was not due to presence ofwild-type
virus in the inoculum, nor to reversion during multiplication, since the culturesmaintained at
restrictive temperature throughout the entire
experiment showed no increase in [3H]uridine incorporationbetween 3 and 9 h afterinfection.
These resultssuggestedthat viralpolypeptides synthesized during the first3 h at thepermissive
temperature remained functional after the
in-creaseinincubationtemperature.
TABLE 1. RNAphenotypeandheatstability of19ts mutantsof Chandipuravirus
[3H]uri- Thermal
Comple- dine in- RNA inactiva- Heat
menta- Mutant corpora- pheno- tion
stabil-tion tion(% type 450C ityb
group ofwild (Vt/V0)0
type) a
ChIA tsChi 0.26 - 0.20 R
tsCh9O 0.30 - 0.075 (R)
tsCh363 0.11 - 0.4 R
tsCh449 0.14 - 0.0005 L
tsCh482 0.9 - 0.29 R
tsCh484 0.03 - 0.17 R
tsCh540 0.11 - 0.01 (R)
tsCh867 0.16 - 0.20 R
ChIB tsChl28 0.3 - 0.14 R
tsCh188 0.02 - 0.03 (R)
tsCh203 0.35 - 0.4 R
tsCh315 1.30 - 0.01 (R)
tsCh808 0.25 - 0.0001 L
ChII tsCh4 73.2 + 0.0005 L
tsCh5 55.9 + 0.00001 L
ChIII tsCh465 0.09 - 0.4 R
ChIV tsCh472 0.14 - 0.5 R
ChV tsCh851 329.0 + 0.06 (R) ChVI tsCh319 0.1 - 0.02 (R)
Wild ts+ 100 + 0.30 R
type
aV. the titerbefore andVt the titer after exposureat
450Cfor 4.5 h.
[image:3.504.248.441.79.334.2]b R, Resistant; (R),partially resistant; L,labile.
TABLE 2. Effectof temperature shifton
intracellular RNAsynthesis
[3H]uridineincorporation (%Y' Group Mutant 3 C 31°C a 39.5°C
39.5 39.5°C 31°C
ChIA tsCh363 0.15 0.09 62.0
tsCh449 0.04 NDb 82.0
tsCh540 0.1 0.16 57.7
tsCh867 0.16 0.12 139.7
ChIB tsChi 0.1 0.1 79.6
tsCh128 0.14 3.5 134.0
tsCh203 0.07 13.1 99.0
tsCh315 0.8 0.16 39.2
tsCh808 0.08 0.08 38.0
ChIl tsCh4 35.4 32.1 221.9
tsCh5 10.5 39.4 96.0
ChIII tsCh465 0.1 0.1 59.5
ChIV tsCh472 0.15 17.7 83.8
ChV tsCh851 37.0 39.6 38.5
ChVI tsCh319 0.1 0.08 39.6
Wild ts+ 12.3 12.5 12.6
type
a[3H]uridineincorporation between3and9hpostinfection aspercentageof the310Clevel.
'ND,
No data.Inthereciprocal experiment, cultures infected with any of the
RNA-negative
ts mutantsshowed
significant
[3H]uridine
incorporation
when incubatedfor 3 hatrestrictive tempera-tureand then labeled after transfertopermissive
temperature for 6 h
(Table
2). These results suggested that the mutant genomes remainedfunctional
during
the 3-h sequestration atre-strictivetemperature.
As
expected,
the RNA-positive ts mutantsinduced
significant
incorporation
of[3H]uridine
inboth theup-shift anddown-shiftexperiments
(Table
2).Heat
stability.
Preliminary experiments showed that themutantscould be differentiatedbestby exposureat45°Cfor4.5h. Table 1
lists
thereduction in
infectivity
for eachmutantand wildtype.Bothmutantsbelonging
togroupChII and mutants ts Ch449(ChIA)
and tsCh8O8
(ChIB) appeared to be thermolabile (>2 logio
unitsinactivation). Theremainingmutantswere
fully
orpartiallyheat resistant.Invitro
polymerase
activity.
The invitro polymerase activity ofpurified preparations ofChandipuravirusand itsts mutants was deter-mined as described in Materials and Methods.
Wild-type virus and RNA-positive mutants ts
Ch4 (group ChII), ts Ch5 (group ChII), and ts Ch851 (group ChV) showed considerablymore
VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.44.236.350.641.2]110 GADKARI AND PRINGLE
in vitropolymerase activity than the remaining mutants atboth 31 and39°C; nevertheless, an
unequivocallyRNA-negative phenotype was not
detected among the remaining mutants (data notshown).
Sincenoneof thets mutantsshowedanyclear
defectinpolymeraseactivity in vitro,anattempt was made to obtain polymerase mutants by
screening clones ofmutagenized Chandipura
vi-rusfor sensitivity to rifampin. Previously, Mo-reau(6)had reportedthe isolation of a
rifampin-sensitive mutant of VSV Indiana which
dis-playedatypical polymerase activityin vitro (7).
Rifampinwasobtained from bothSigma Chem-icals (London) and, as aged andfresh prepara-tions, from G. Tocchini-Valentini, Lancini-Le-petit, Milan, Italy. The antibioticwas dissolved
by adding200mg to 2ml ofdimethylsulfoxide anddilutingto 10 mlwithsterile distilledwater. Thisstock solutionwasdiluted1:100 in solid or liquidincubationmedium to give afinal
concen--M *
*MIt
-.M
.Ch Ch
I I:I
3 4 5,6
£Ch
78
J. VIROL.
tration of 200 ,ug/ml. A total of 593 clones of
Chandipuravirusgrown in the presence of
100-,ug/ml
5-fluorouracil were tested for ability tomultiply in the presence of 200 ,ug ofrifampin
perml, butnone was sensitive to the drug. All the preparations of rifampin reduced vaccinia virus yields by more than 4log10 units at this
concentration. VSVwas slightly morerifampin
sensitive than Chandipuravirus, but it was not
possibleto clearlydifferentiate the rifampin-sen-sitive mutant (VSV-29) isolated by Moreau from
its wild type
(VSV-VJ)
in BS-C-I cells under theseconditions,
and the attempt to obtain polymerase mutants by this procedure wasabandoned.
Intracellular protein synthesis. BHK-21
cell monolayers were used forradiolabeling of
intracellular polypeptides in preference to
BS-C-1cells, becausethere was less loss of
cells
bydetachmentduringthe labeling period. Figure 1 shows the[3S]methionine-labeledpolypeptides
cb
tIV
9 10
*.. :.. .... :... :.:
" .s.- .
1h1
11 1.2
1*
WT
13 14
3IC 39C
15 16
1N
=rN
NS/A
aM
_ _
4NVP1I
FIG. 1. Polypeptide synthesisin BHK-21 cellsinfectedwithtsmutants tsChl,tsCh4,tsCh465,tsCh472,
andtsCh851,fromcomplementationgroupsChI,ChII, ChIII,ChIV,andChV, respectively.The odd-numbered
tracksarethepolypeptide profiles of samples fromcultures labeledat31°C,and the even-numbered tracks
aresamplesfromcultures labeledat39°C. Monolayers ofBHK-21 cells(orBS-C-1 cellsfortracks 15 and16)
wereinfected at20PFU/cell and labeled between 4 and 7 h with 30,iCi of [35S]methionineper ml. The
cultureswere harvested andproteins were extracted asdescribed in the text. Samples containing equal
amountsofradioactivitywereelectrophoresedon a5to12.5%polyacrylamide gel for3to4hat40mA. The
viralpolypeptidesare labeledL,G,N,NS, andM;Aisactin, NVP1is nonviralpolypeptide 1,and MI isa
mockinfection.Tracks15and 16weretakenfromanothergeltoshow thatNVP1wasalso resolved in
BS-C-1 cellsinfectedwithtsCh851 (groupChV)at39°C.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.124.411.292.567.2]CHANDIPURA VIRUS MUTANT PHENOTYPES 111
synthesized in cells infected withwild-typevirus
and mutants ts
Chl,
tsCh4,
tsCh465,
tsCh472,
and ts Ch851, representing complementation groups ChI, Chll, ChIII, ChIV, and ChV,
re-spectively.All the tsmutants,includingtsCh319 of group ChVI (not shown), synthesized less
viral protein at 39°C (even-numbered tracks).
The Lpolypeptide was notresolved in the cul-tures incubated at 390C, except in ts Ch851
(group ChV)-infected cells. The G protein was
resolved intotwobandsin most of the mutant-infected cells at310C. Thesetwobandsprobably correspond to the Gl and G2 polypeptides of
VSV andrepresent two stages in glycosylation
(2; R. M. L.Buller, Ph.D. thesis, University of
Glasgow, Glasgow, Scotland, 1975). The pulse-chaseexperiments illustrated in Fig. 2, 3,and 4
confirmed thatashift inelectrophoretic mobility
1L
4N
NS/A
ofthe G protein occurred duringincubation, as with VSV Indiana (2). The N polypeptide of Chandipura virus migrated just above the cel-lular actin band and was clearly resolved in all the mutant-infected cells. The NS polypeptide
was notresolved, since it migrated in the region of cellular actin. The Mpolypeptidewas present in allthevirus-infectedcell extracts.
A[35S]methionine-labeled polypeptide desig-nated NVP1 (nonviral polypeptide 1) was ob-served in both BHK-21 (track 12) and BS-C-1
(track 16) cells infected with mutant ts Ch851 andincubatedat390C.NVP1 was not observed in tsCh851-infectedcells at310C(tracks 11 and 13), or incells infectedwithwild-type virusand other ts mutants ateither 31 or390C. NVP1was also not resolved incellsinfected witheitherof two spontaneous revertantsof tsCh851 (Fig. 4).
*
;
... ... . N.C
-'*
''r;}: :i, :: .: .. .. : .:.:
*s.
z z..X
v o!:.. .'
::'
....:a
e'.:t
*w,:,,i
...:.
...
..
.::e:#i,r}
;w--
__44M
_ __
_;w
..._..
*
* .'.*uNVP1
S
FIG. 2. Thepolypeptides oftsCh851-infectedcells inapulse-chase experiment. BHK-21cellcultureswere
infected withmutant tsCh851at20PFU/cellandincubatedfor4hat31or39°C. The cultureswerelabeled
for15 min with 30 uCiof [35S]methionineperml and then returnedto ordinary medium. Cultures were
harvestedat0, 1,2, and 3hafter labelingasdescribed in thetext.Samples containing equalcountswere
electrophoresed on a 5 to 12.5%polyacrylamide gel. Tracks 3, 4, 5, and6, Cultures incubated at 31°C
throughout and harvested at0, 1, 2, and 3 h, respectively, after labeling; tracks 7, 8, 9, and 10, cultures
maintainedat39°Cthroughout;tracks11, 12,and13, cultures labeledat39°C andsubsequently incubatedat
31°C for 1, 2, and 3h, respectively. Tracks I and 14show markerpolypeptides from partially purifiedts
Ch851,andtrack2showspartially purified wild-type virus. The lettersareidentified in the legendtoFig.1.
si
1£ *
SC
<*3tC4:
S 8: 4: S S ? S 0 1011 1213l14 VOL. 33,1980
.1:
op
A
C;
rN::
on November 10, 2019 by guest
http://jvi.asm.org/
112 GADKARI AND PRINGLE
-4 3C
i}%
3,a*NVP2
-
*~~~~~NVP
1
FIG. 3. Polypeptide synthesisintsCh851-infected
cells after transfertorestrictive temperatureat4 h
after infection.Theconditionsof infectionand
label-ingwere asdescribed inthelegendtoFig.2.Tracks
2, 3, and 4, Samples from cultures incubated and
labeledat31°Cand chasedfor 0, 1,and 2h,
respec-tively; tracks5, 6,and 7, correspondingcultures
in-cubated at 39°C throughout; tracks 8, 9, and 10,
cultures incubatedat31°Candsubsequentlylabeled
and chasedat39°C for 0, 1,and2 h. Track I shows
thepolypeptides fromamock-infectedculture at3lC.
The lettersareidentifiedin thecaptiontoFig.1.
Thestabilityoftheviralpolypeptides synthe-sized at39°C was examined inpulse-chase
ex-periments (3) asdescribedinthelegendtoFig. 2. Nodegradationof viralpolypeptideswas
de-tected in cells infected with mutants ts Chi (group ChI), ts Ch4 (group ChII), ts Ch465 (group ChIII), ts Ch472 (group ChIV), and ts
Ch319 (group ChVI).IntsCh851-infected cells, however, it was evident that the intensity of
labeling ofthe Mpolypeptidedecreasedduring incubationat390C(Fig. 2, tracks 7to10). This effectwasnotobserved when the cultureswere
incubatedat310Cafterlabelingat390C (Fig. 2, tracks 11 to 13). However, if the cells were
incubatedat310Cand thenlabeledand chased
at39°C,degradationof Mwasobserved(Fig. 3,
tracks8to10).NVP1wasapparent ints Ch851-infected cells,providedthe cultures were
incu-batedat390C duringorafterpulse-labeling (Fig.
2, tracks7to 13,andFig. 3,tracks 5 to10). An
additional polypeptide (NVP2) has been
re-solved in Fig. 3, but the nature of this
poly-peptideor the band between N and M in track 5 isunknown. Neither was observed consistently in other gels.
Aspontaneous revertant (Ch851/R2) of mu-tant ts Ch851 was isolated and compared with the parental mutant in the same experiment (Fig.4). Theinstabilityof the Mpolypeptidein tsCh851-infectedcellsat39°Cwas again appar-ent(tracks5 to7),but incells infectedwith the revertantthere wasless degradationof M (tracks 11 to 13). Densitometertracings of the tracks in
Fig. 4 showed thatduring the chase period
ra-dioactivity in theMpeak declinedto60%of the
initial level forrevertantCh851/R2 and33% for mutant ts Ch851, standardized in each case to
the Npeak. Similarresults were obtained with
anotherrevertantcloneof tsCh851.
Host rangeproperties of ts and tdCE mu-tantsof
Chandipura
virus. Thewild-typeandconventional ts mutants produced plaques on
CE cells at
310C,
andonly
the wild typepro-duced plaques at 39°C, i.e., the ts mutants
re-tained their phenotype inavian cells. Mutants
of both VSVIndiana andVSVNew Jersey have
been isolated, however, which were
condition-ally
temperature sensitive in CEcellsand weredesignated tdCEmutants(10). Therefore Chan-dipura
wild-type
virus was mutagenized by growth in thepresence of 100,g of 5-fluorouracil perml,
and cloneswerescreened for theirabilityto form plaques on BS-C-1 and CE cellsat 31
and
390C.
The six tdCEmutantslistedinTable3 were obtained from 509 isolates, i.e., a fre-quency of 1.2%. No mutants completely
re-strictedonCE cells
(the
hrCEphenotype)
were obtained. The six tdCE mutants were titrated alsoonmonolayers of hamster (HE),rat (RE),feline (FEA), bovine (BEK), andmouse
(NIH/
3T3)embryocells andonmouseL929cellsat 31
and
390C
(Table
3). Three ofthe six mutants were temperature sensitive on CE cellsonly,
whereasthe other three mutants showedsome
temperaturesensitivity onmammalianembryo
cells.
Indeed, mutanttdCE Ch530 was temper-aturesensitiveinalltypesofcellsexceptBS-C-1 cells. None of the tdCE mutants
comple-mentedany of the six groups in CE cellsat
390C,
andconsequently
it was notpossible
toassign
themto
complementation
groups.However,
for reasons which arestillunclear,little significant complementation was observed between theconventionaltsmutantsof
Chandipura
mutantsinCE cells (datanotshown).
DISCUSSION
Thephenotypic properties of the ts mutants
ofChandipuravirussuggest that
complementa-J. VIROL.
'm .31
I ')'. ;, 4 .,
P-41-M!
I',' ::"..:..
i.,...:: ....
J--.w
t'.4'e"
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.73.262.67.326.2]CHANDIPURA VIRUS MUTANT PHENOTYPES 113
tsaC 851
M * 31C 4
39C
1 2 3 4 5 6 7
Ch 8S1
R/2
m31C 4 *
39C
48 9 10 11 12 13 14M
L
f
'...::...:.:
~~~i6
s v / . > ,.X. M j :*.N
'..:.}.2.§.:N_ A
_. 0 ....
_-
4 M4
NVP1
FIG. 4. Polypeptidesynthesisin cellsinfectedwith cloneCh851/R2,anon-temperature-sensitiverevertant
oftsCh851. The conditionsof infectionandlabelingwere asdescribed in the
legend
toFig.
2,exceptthatthe cultureswereharvestedat0, 1.5,and3hafter labeling. Tracks2, 3,and4, Samples fromculturesinfected
with tsCh851 and incubated throughoutat
31°C;
tracks5, 6, and 7,corresponding
cultures incubatedat39°C; tracks 8, 9, and10,culturesinfectedwith therevertantcloneCh851/R2and incubatedthroughoutat
310C;tracks11,12,and13,correspondingcultures incubatedat39°C. Tracks 1 and 14 showpolypeptidesin mock-infectedcellsat31°C.Thelettersareidentifiedin thecaptiontoFig. 1.
tion groups ChII and ChV
correspond
to thegenes
coding
for the twoproteins
ofthe viralenvelope.
ThevirionsofbothmutantsingroupChIIwerethermolabile
(Table 1),
andtheintra-cellularM
polypeptide
oftsCh851(group
ChV)
wasunstable atrestrictive temperature (Fig. 2,
3, and4).Reversionof the temperature
sensitiv-ity oftsCh851was
accompanied by
anincreasein stability of the intracellular M
polypeptide
(Fig.4). Mutant ts Ch851 wasalso an overpro-ducer of RNA at restrictive temperature, like the group IIImutantsof VSV Indiana whichare
also considered to be Mprotein mutants (3, 4, 9). It is notpossibletosuggest any gene
assign-ment in the case of the four
RNA-negative
groups.Group
ChI,
which contains88% of thetsmutants ofChandipura virus, may correspond
to group I of VSV Indiana
(containing
77% of mutants), which isconsidered torepresent thegenecodingfor the Lprotein (9). However,none
of thets mutants ofChandipura virus, including thethermolabiletsCh449 ofgroupChIA andts
Ch808
ofgroupChIB,
had defective RNApo-lymeraseactivityinvitroatthe restrictive
tem-perature.
TheapparentmutantspecificityofNVP1
sug-gested that it might be a minor virus-coded
polypeptide which accumulated in conditions wherematurationwasblocked buttranscription
continued unrestricted. However, NVP1 could not be chased into virions by
labeling
at39°Cand incubation at
31°C (data
notshown).
Inaddition, NVP1didnotappeartobederived
by
proteolytic cleavage from any of the five viral
VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.98.386.74.407.2]TABLE 3. Titers of six tdCE mutantsof Chandipura virus in various cell linesa
Temp of tdCE tdCE tdCE tdCE tdCE tdCE
Cels incubation Ch514 Ch516 Ch525 Ch530 Ch565 Ch859
(OC)
BS-C-1 31 5x 108 5x 108 1.5x 105 1.5x108 1.5x108 2x 108
39 2x 108 108 2.5x 107 107 107 5x 107
CE 31 1.5x107 107 io7 107 5X 106 3x 107
39 104 2X 103 1o2 102 5x103 2X 104
HE 31 1.5x 107 3X106 1.5x 107 107 5X106 1.5x 107
39 3X106 2.5X106 3x 106 4X104 106 106
RE 31 2x 106 3x 106 2x106 5 x105 3x 105 5x 106
39 5x 105 4x 105 5x 105 103 1.5x104 5 x104
FEA 31 2.5x107 2x 107 2.5x107 2x 107 107 3x 107
39 7.5x106 107 106 5 x103 106 106
BEK 31 2.5 X106 106 2x 106 2x106 1.5X106 2.5x 106
39 7x 105 5x 105 1.5x105 2X103 3.5x 104 5X 104
L929 31 5x 106 5 X 106 5x 106 2 X 106 3.5X 106 3x106
39 5x 105 3.5x105 2x 105 102 1.5x105 5x102
NIH/3T3 31 106 2x 106 2.5x 106 2x 106 5X 105 5x 106
39 106 5x 105 5x 105 102 2x 105 1.5 x104
a Thetitersthatshowsignificant
differences
areindicatedinitalics.
proteinsinexperiments (not shown)in which ts Ch851-infected cells were labeled andharvested
inthe presenceofthe proteaseinhibitors tolyl-sulfonyllysyl chloromethylketoneand tolylsul-fonylphenylalanyl chloromethylketone. There-fore itwasconcluded that NVP1 wasprobably
ahostproteinwhich accumulated at39°C asa consequenceof the defectivematuration of mu-tant ts Ch851. Occasionally a faint band was
observedin the NVP1positioninmock-infected cells(e.g.,Fig. 3,tracks 1to4). Stimulation ofa specific host protein by a rhabdovirus has not
beenreportedpreviously,butenhanced
synthe-sis of two host proteins has been observed in
simianvirus40-infectedCV1 cells (8).
Finally, the isolation of tdCE mutants of
Chandipuravirussuggests that host factorsare
involved in thereplication ofallrhabdoviruses,
notonlyVSV(10).
ACKNOWLEDGMENT
D.A.G.wastherecipient ofaFellowshipfrom the Com-monwealthCommissionduringthecourseof thiswork.
LITERATURE CITED
1. Gadkari,D.A.,and C. R.Pringle.1980.
Temperature-sensitive mutants ofChandipuravirus. I. Inter- and intragroupcomplementation.J.Virol. 33:100-106.
2. Knipe, D.,H. F.Lodish,andD.Baltimore.1977.
Lo-calization oftwocellularformsof the vesicular stoma-titisviralglycoprotein.J. Virol. 21:1121-1127. 3. Knipe, D.,H.F.Lodish,andD.Baltimore.1977.
Anal-ysis of the defects oftemperature-sensitivemutantsof vesicular stomatitis virus: intracellulardegradationof specific viral proteins.J. Virol. 21:1140-1148. 4. Lafay, F.1974.Envelope proteinsof vesicularstomatitis
virus: effect oftemperature-sensitive mutation in com-plementationgroupsIIIand V. J. Virol. 14:1220-1228.
5.Marsden, H. S., I. K. Crombie, and J. H.
Subak-Sharpe. 1976.Control ofprotein synthesis in
herpes-virus-infected cells: analysis of the polypeptidesinduced by wildtypeandsixteen temperature-sensitivemutants of HSV strain17.J.Gen. Virol. 31:347-372.
6.Moreau,M. C. 1974.Inhibition ofavesicular stomatitis
virusmutantbyrifampin. J. Virol. 14:517-521. 7. Moreau, M. C., B.Sanzey, and Ch. Chany.1975. In
vitrotranscriptionofavesicularstomatitis virus mu-tant, p. 103-108.In Invitro transcription and translation of viral genomes, INSERM Colloquium, vol.47.
IN-SERM,Paris.
8.O'Farrell,P.Z., and H. M. Goodman.1976.Resolution
of simian virus40proteins in whole cellextractsby two-dimensionalelectrophoresis: heterogeneity of the major capsid protein. Cell9:289-298.
9. Pringle,C. R. 1977. Genetics of rhabdoviruses, p. 239-289. In H. Fraenkel-Conrat and R. R.Wagner (ed.), Comprehensive virology, vol. 9. Plenum Publishing Corp.,New York.
10. Pringle, C.R. 1978. The tdCE andthehrCEphenotypes: host range mutants ofvesicular stomatitis virus in whichpolymerase functionis affected.Cell16:597-606.
11. Szilagyi,J.F.,andC.R.Pringle.1972.Effect of
tem-perature-sensitive mutations on thevirion-associated RNAtranscriptaseof vesicular stomatitis virus. J. Mol. Biol.71:281-291.
12.Szilaigyi,J.F.,andC. R.Pringle.1979.Effect of
tem-perature-sensitive mutation on activity of the RNA transcriptase of vesicularstomatitisvirusNewJersey. J.Virol.30:692-700.
114 GADKARI AND PRINGLE J. VIROL.