Vol. 42, No. 3 JOURNALOFVIROLOGY, June 1982,p.1118-1122
0022-538X/82/061118-05$02.00/0
Complementation for Replicative
Form
DNA
Replication
of
a
Deletion
Mutant
of
H-1
by Various
Parvoviruses
SOLON L. RHODE III
Institutefor Medical Research of Bennington, Bennington, Vermont05201 Received7December1981/Accepted8February 1982
A defective interfering genome of the parvovirus H-1, called dll, has been isolated and characterized. The only alteration in dll that has been detected isa
300-base-pair deletionatmapposition 38. Replicative form DNA replication ofdll
requires the RF repfunction of a helper virus and the parvoviruses H-1, H-3,
MVM, or MVM(i), but not LuII complement dll for replicative form DNA
replication.
Defective interfering(DI) genomesof the
par-vovirus H-1 were found to be heterogeneous
genomes modestly deleted by up to 10 to 12%
andtocontain 58-base-pair (bp) tandemrepeats
originatingin themapregion91 to 95 (5, 8). DI particles required a helper virus to replicate replicative form (RF) DNA (6). In this study I
have explored this problem further by using a
cloneddeletionmutantof H-1, dll, that has the advantage of being a deleted DI genome that
appears tobe homogeneous in size and lacksany
additions at map positions 91 to 95. The
dele-tion, which is approximately300bp, allowsthe
quantitation of dllRFDNAreplication in mixed infections with various helper viruses because dll RF DNA can be separated from standard
length genomes by agarose gel electrophoresis. dllwas originally detectedas adeletedgenome
inastock of virus generated fromasingle plaque ofan H-1 tsl4revertant atthe restrictive
tem-perature39.5°C. ts14 isanH-1mutantdefective
in RF DNA replication (7). Anumberof
sponta-neousrevertantsoftsl4with normalratesofRF
DNA replication were isolated by plaqueassay at the restrictive temperature after three serial
passages at39.5°C. Examination of theRF DNA
replication of these revertants by agarose gel electrophoresis revealed that one of them
con-tained dll as wellas astandard lengthgenome.
Continued serialpassageof this virus stockwas
carriedoutby inoculating100-mmpetridishes of
NB cells that were tipped up atabout
300
with0.2 ml of undiluted virus suspension and ad-sorbedfor 0.5h.The disheswerereturnedtothe horizontalposition, 10 mlof mediumwasadded,
andthe dishes wereincubated at37°C until the cytopathic effect was complete. At the second
passage the cell lysate contained only 4 x
105
PFU/ml. The RF DNAproduced by this stock
wasalmostentirely dll, and the yield ofdll RF
DNA wasincreased bytheaddition ofwild-type
(wt) H-1 helper virus(datanotshown).
To test whether dll was defective, the dll
stock was serially passaged two times, with a
multiplicity of infection (MOI)of 0.01 PFU per
cellateachpassage. Thepresence ofdll in the
secondpassage stockwasthenassayed by
gen-eratingRF DNAandanalyzingitbyagarosegel electrophoresis. Surprisingly, this preparation
wasalmost entirelydll. Becausedll might have been in large excess in the initial stock this experimentwas repeated, exceptthatthe serial
passage was repeated three times at an MOI of
0.001 at each passage. Examination of the RF DNAproduced by eachpassageshowedonlywt
RF DNA in the second andthirdpassages.This indicated that dll was probably defective and requiredahelpervirus. Asanadditionaltestfor defectiveness, plaques were generated from a
dll stock, and 20 well-isolated plaques were
picked, propagated, and then their RF DNAs
werecharacterized by gelelectrophoresis. In15
of 20clones, dll RF DNAwaspresentinvarious proportions with helper virus DNA, and the remaining 5 were standard length RF DNA. Because H-1 dll was in excess, yet no plaque isolates were dll only, dll probably does not
form plaques and thus is defective. The virus stockproduced by oneof theseplaques, which
generatedmostlydeleted RF DNA, was usedfor
theremaining experiments in this study.
The DI particles of H-1 generated by serial
high multiplicity passage expressed their inter-ference by inhibition ofwtvirionprotein synthe-sis,whichwaseasilymonitoredbya hemaggluti-nationassay.dll wassimilarly tested,and when
NBcells wereinfected withwtH-1 atanMOI of
3 in the presence or absence of the dll virus stock(MOI of0.6PFUpercell),the
hemafluti-nation assays at48hpostinfectionwere2 and 224 U/ml, respectively. Thus, coinfection with dll reduced thehemagglutinationassayyieldof
wt virusby
2-7
U/ml. Inconjunction withthis, polyacrylamide gel electrophoresis ofthepro-1118
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 42, 1982
teins synthesized during infection with dll
re-vealed only adecrease in yield of viral proteins
VP1 and VP2, with no evidence for truncated
viralproteins.
The location ofthe deletion indll was
exam-ined by restriction enzymedigestion of dll RF
DNA. Restriction of dll RF DNA withHindIlI
(cleaves wtH-1 at mapposition 51) and EcoRI
(cleaves at map position 21) indicated that the
deletion mapped between mapunits21 and51.
Mapunitsare on ascaleof0 to 100,with0 atthe
3' terminus of the virion strand. RF DNAs of wt
H-1, dll, and DI-wt passage 30, labeled with
[32P]orthophosphate, were prepared and digest-edwithHhaI (5). The restrictionmap of wt H-1 to the restriction endonuclease HhaI is
de-scribed elsewhere (8). The digests were
ana-lyzed by electrophoresis in a 1.8% agarose gel
1 2 3 4 5 6 7
did-1RF- 1
I
I3.=
ji4
I VJd!-lRF- 1 Wi6i
[image:2.487.253.444.59.320.2]wt DV-wt di-]
FIG. 2. Fluorogramof agarosegelelectrophoresis of dll RF DNA at various MOIs of helper virus.
Replicate NB cell cultures were infected with dll
(endogenousH-1helpervirusat anMOIof1)andwith
variousamountsof exogenous H-1 as inTable 1. The
MOIsof helper virus shownare:(1) 1,(2) 10,(3) 30,(4)
300;and,in lanes 5to7,culturesreceivedhelper virus
onlyatMOIsof:(5)10,(6) 30,and(7)100.
Electropho--l4oobp resis was carried out in a 1.8% agarose gel at 100 Vfor 16 h.Fluorographywas asdescribed in(1).ThedIl RF DNA has arelative mobility of 1.05 comparedwith thatof H-1 RF DNA.
FIG. 1. Electropherogram of HhaI digests of the RF DNAofwt,DI-wt (passage 30)and dIl. Electro-phoresiswascarriedoutinaslab gelof 1.8%agarose
at400V/h.Portionsof this experimentwere
previous-lyreported (5). The Hhal digests for DI-wt and dllare
not complete, accounting for the high-molecular-weight fragments. Thenumbersintheleft margin refer
to themap positions of the fragments;0isatthe 3'-terminus of the virion strand, and 100 is atthe 5'-terminus.
(Fig. 1). dll RF DNA was missing the Hhal
20.5/38.0 (930 bp) and 38.0/52.9 (790 bp)
frag-ments aspreviously reported for DI-wt (5). dll
hasanewfragment, withamobility of 1,400 bp,
which indicates that the deletion of 300 bp
encompassestheHhaI siteat38.0. Examination
oftheremaining HhaIfragments of dll in
acryla-mide gels showed them to be the same size as
thoseofwtH-1 (datanotshown).
The effect ofhelper virusesat various MOIs
on the RF DNA replication of dll was tested.
NBcells wereinfected witha constant amount
ofdll andcoinfected withwtH-1atMOIs of1,
10, 30, 100, and 300 PFUpercell. Viral DNAs
werelabeled with[3H]thymidine, extracted, and
subjected to electrophoresis in a 1.8% agarose
gel. Arepresentative fluorogram ofoneof these
experimentsisshown inFig.2. Theyieldsofdll
and the H-1 helper virus were quantitated by
excising the regions containing the appropriate
DNAandmeasuring theamountof 3Hby liquid
scintillation spectrometry(Table 1).
-dI hvRF
hvRF
72.5/97.4
-20.5/38.0
-53.4 /69.2 -38.0 /52.9
-9.4/19.0
-A
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.487.47.240.276.578.2]1120 NOTES
TABLE 1. Yield of RF DNA (counts perminute)a
wtMOI dll wt (+dll) wt
1 500 200
10 2,100 600 3,400
30 3,400 1,500 7,000
100 3,800 3,400 15,000
300 3,500 3,800
a Replicate NB cell cultures (3 x
106
cells per 60-mmpetri dish) werepartially synchronized by treat-mentwith methotrexate for 12 hbefore infection. The cultures were infected with a constant amount ofdll, with its endogenous helper virus (MOI of 1), and with various amounts of wtH-1 to provide MOIs of 1, 10, 30, 100, and 300. Three dishes received only the helper virus at anMOI of 10, 30, or 100. The viral DNAs were labeled with[3H]thymidineat5 uCi/mland10-6Min the presence of 5-fluorodeoxyuridine at 0.5 ,ug/ml from 12 to 17 h postinfection. The dll RF DNA was separated from wt RF DNA by gel electrophoresis (Fig. 2). Afterfluorography, the regions of gel contain-ing each type of RF DNA were excised, and the3H wasquantitated by liquidscintillation spectrometry.The yields of 3H-labeled dll RF DNAs were increased inadose-dependent fashion by
mod-est levels ofhelper virus (MOIs, 1 to 100), but
they began to decrease at the highest level of helper virus (MOI 300). The yields of wt RF
DNA or even the sum ofwt + dll RF DNAs
weredepressed in the mixed infectioncompared with thatofwtalone.
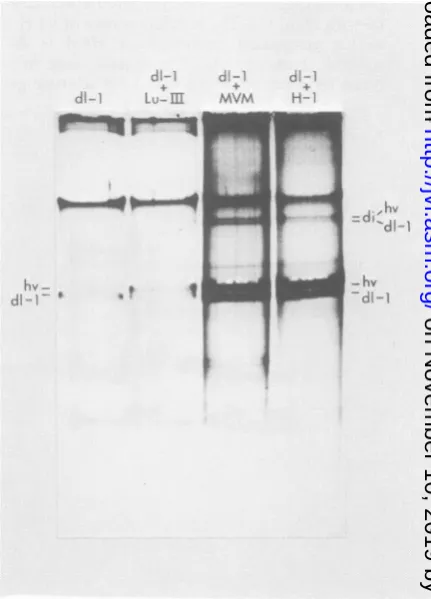
In asimilarexperiment, theheterologous
par-vovirus MVM was tested and found to be an efficienthelper virus for H-1 dll RFreplication, whereasLuIl wasnot(Fig. 3). H-3 andMVM(i) (3, obtained fromP.Tattersall, Yale University, New Haven,Conn.)werealso foundto comple-mentdll forRFreplication (data not shown).
Todetect complementationfor later steps in
virus replication suchasprogeny DNA synthe-sis and encapsidation by aheterologous helper virus, dll was propagated in THK22A cells (Simian virus 40-transformed hamster kidney obtained from Joan Kaplan, Harvard
Universi-ty,Boston,Mass.), withMVM as ahelper virus fortwo serial passages. If dll iscomplemented by MVMfor all of its defective functions, phe-notypicmixing willoccur,and dll willbe
encap-sidated by the capsid proteins of the MVM
helper virus.In thatevent, the yields of dll RF DNA will be sensitive to neutralization ofthe
virus inoculum with anti-MVM antisera. For this
assay,H-3wasaddedtotheculturesatthetime
ofinfection to ensurethepresence of sufficient helper virus (Fig.4Aand4B). Theresults show
that the MVM-dll passage 1 virus contained
largeamountsof dll neutralized byMVM
anti-serum, but not by H-1 antiserum or normal
hamster serum.ForMVM-dllpassage2,asmall
quantity of deleted RF DNA was generated by
the MVM + dll virus, and itwas decreasedby
the MVM antiserum. As a control, H-1 + dll
virusshowedelimination of dll RFDNAbyH-1 antiserum, but not with anti-MVM or normal
serum(Fig. 4A). Thus,MVM notonlysupports
RF DNA replication of H-1 dll, but it also
complements for progeny DNA synthesis and
encapsidation. The reason that the dll in the
MVM-dll passage 2 produced a reduced and
broadened RF DNA band is unknown, unless
MVM complements at reduced efficiency in
d-l
di- LKu--m
r II
hv
-di
-da-1 di--l,
*vm H~+
MVM H-.
hv
d1-1
9
iI
FIG. 3. Fluorogram of agarosegelelectrophoresis of dllDNAwithLulIlI,MVM,orH-1 ashelper virus. THK22A cellswerecultured in 100-mmpetridishesat 5 x 10' cells per dish. The cultures were partially synchronized bytreatmentwith methotrexate for 10 h before infection. Replicate plates were infected with
dll(H-1 helper virus [hv]at anMOI of1)and, for lane 1,noadditional helper virus; 2,LulIl (MOI of 10); 3, MVM(MOI of 6); and 4, H-1 (MOI of 40). The cultures were labeled with [3H]thymidine (5 ,uCi/ml, 10-6M
plus 5-fluorodeoxyuridineat0.5,ug/mlfrom 12to17 h
postinfection). Viral DNAs were extracted as
previ-ously described(4), and theelectrophoresiswas
car-riedoutina1.4% agarose gelat50Vfor 16 h. The
positionsofmonomerRF DNAofhelper virus and dll
areindicatedbyhvanddll,respectively;diindicates dimerRFDNA.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.487.254.470.209.509.2]NOTES 1121
rmvrn H-1 - d6-l
+
arnti uti-
antil-A.
mvml nvmI,] .S. H-1mvm - ad -i
calti
tllit-N.S. H-1 N.S. TqvmTl
: I
I
s;-I F
t'
i
i
9 i
hv REF
di-1 RFE-
[image:4.487.79.413.99.503.2]H-1 2 3 4 5 6 7
...
_~~~
_mvm-dI-I H-3-di-1
anti-B). N.S. mvm H-i H-3 N.S.
di
RF
-hv
RF-dl-I
RF-h4t~~~~
vi
L-8
hvRF
-61- RF
1 2 3 4 5
FIG. 4. Fluorograms ofagarosegelelectrophoresis of dll RF DNA producedaftertreatmentwithspecific antisera.dllwaspropagated serially fortwopassagesin THK22A cells with MVMorH-3addedashelpervirus
(hv)atanMOI of 10 (MVM-dll orH-3-dll). Thedli virus stockswereincubated withhamsterantiserabefore infectionandlabeling of THK22A culturesasinFig. 3. Virus stock (0.4 ml)wasincubated with50,ulofantiserum
for 1 hat37°C. The hemagglutinationinhibition titer of the anti-MVMwas2-l" toMVM,theanti-H-1titerwas
2-16toH-1,theanti-H-3 titerwas2-14toH-3,and the nonimmuneserum(N.S.)was0toallof the viruses. In (A),MVMwastreated with anti-MVM (lane 1)orN.S.(lane 5)as acontrol. Toensureadequate helpervirusfor
anydllwhichsurvived neutralization, H-3wasaddedatanMOI of30toallofthedllinfections. TheH-1-dll stockafterneutralization is in lanes2 to4, and theMVM-dll (passage 2)is in lanes6to8. In (B), MVM-dll (passage 1)wassimilarly treated (lanes1to3)andH-3-dll (passage 2) is shownaftertreatmentwithanti-H-3
antiserumornomimmuneserum(lanes 4 and 5). TheMVM-dllandH-3-dllwereanalyzedonseparategels,and onlythe bottomportion ofthe gels areshown. The DNA species areidentified onthe margins: (A) hvRF,
monomerhelpervirusRFDNA;dllRF,monomerdllRFDNA; (B)di RF,dimerRF DNA;*indicatesa
heat-labileconfiguration ofviral DNAnotfully characterized thatis prominent in this experiment. Electrophoresis
wasina1.6%agarosegelat100Vfor16h.
- hv RE
-dl-1 RE
VOL.42, 1982
t.
on November 10, 2019 by guest
http://jvi.asm.org/
1122 NOTES
comparison with H-1.Asimilarphenotypic mix-ing was obtained with H-3 and dll (Fig. 4B).
In summary, the DI deletion mutant of H-1, dll, requires the RF rep function of a helper virus to replicate its RF DNA. The homologous virus H-1 and the heterologous viruses H-3, MVM, MVM (i), but not LuIII, complement dll for RF replication. H-3 and MVM were also found to be capable ofcomplementing dll for the functions required for progeny DNA synthesis andencapsidation. dll is strongly interfering to its helper virus, yet it does not contain extra copiesof the tandem repeats near the replication origin characteristic of H-1 DI genomes (5, 8). Therefore, I conclude that these tandem repeats do not effect the interference. It should be noted that themajor promoter for H-1 has been report-ed tobe at map position 40 (2). My preliminary sequencing data show that the canonical pro-motersequence TATAAA (sense strand)is only 20nucleotides proximal to the HhaI site at 38 (S. L. Rhode, unpublished data). Thus, it is likely that the deletion in dll has disrupted this pro-moter and caused a defect in the synthesis of both of the capsid proteins, VP1 and VP2. This lesionalso seemed to be present in DI genomes generated by serial highmultiplicity passage and maybe an important property of DI particles (5).
IthankJessica Bratton and Jeanne Helft for expert techni-calassistance.
Thiswork was supported by Public Health Service grants CA25866 and CA26801 from the National Institutes of Health.
LITERATURECITED
1. Laskey, R. A., and A. D. Mlls. 1975. Quantitative film detection of 3H and"Cinpolyacrylamide gels by fluorog-raphy. Eur. J. Biochem. 56:335-341.
2. Lebovitz, R. M., and R. G. Roeder. 1981. Structure and DNA-proteininteractions ofreplication origins,p. 431. In Dan S. Ray and C. Fred Fox(ed.), ICN-UCLA Symposium onMolecular and Cellular Biology, vol. XXI. Academic Press, Inc., New York.
3. McMaster, G. K., P. Beard, H. D. Engers, and B. Hfrt. 1981.Characterization ofanimmunosuppressive
parvovi-rusrelated to minutevirusofmice. J.Virol.38:317-326. 4. Rhode, S. L. 1977.Replicationprocessof the parvovirus
H-1. VI.Characterization of areplicationterminus of H-1 replicative-form DNA. J. Virol. 21:694-712.
5. Rhode, S. L. 1978. Defectiveinterfering particlesof parvo-virusH-1. J. Virol. 27:347-356.
6. Rhode, S. L. 1978. H-1 DNAsynthesis,p. 279-296. In D. Ward and P. Tattersall (ed.), Replication of mammalian parvoviruses. Cold Spring Harbor Laboratory, Cold Spring
Harbor, N.Y.
7. Rhode, S. L. 1978.Replicationprocessoftheparvovirus H-1. X. Isolation of a mutantdefectiveinreplicative-form
DNAreplication.J.Virol. 25:215-223.
8. Rhode, S. L., and B. Klaassen. 1982. DNA sequence of the 5' terminuscontainingthereplicationorigin ofparvovirus replicative formDNA. J.Virol.41:990-999.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/