JouRNAL OFVIROLOGY, Nov. 1974,p.1220-1228 Copyrighti 1974 American Society forMicrobiology
Vol.14,No. 5 PrintedinU.SA.
Envelope Proteins of Vesicular
Stomatitis
Virus: Effect of
Temperature-Sensitive
Mutations in
Complementation Groups
III and
V
FLORENCE LAFAY
Laboratoire de Biologie Experim'entale, Bat. 400; Institut deMicrobiologie, Bat. 409, UniversiteParis-Sud Centre d'Orsay, 91405 Orsay, France
Received forpublication 8 July 1974
All fivemajor viral proteins weresynthesized inchickenembryocellsinfected
with vesicular stomatitis virustemperature-sensitive (ts)mutantsof
complemen-tation groups III and V and maintained at thenonpermissive temperature. The
distribution of theseproteins among cytoplasmic cellular fractions separated on
discontinuous sucrose gradients was identical for wild-type and tsllI-infected
cells. Strikingly different patternswere observed for the Gprotein in gradients
fromcells infectedbytsV mutants; verylittle,if any, Gproteinwasfound inthe
lightest fraction. Pulse and chase experiments with wild-type, virus-infected cells showed that protein G moves from the heaviest to thelightest fraction before being incorporated into the virion. After shift down tothe permissive
tempera-ture (30 C), G protein synthesized at 39.6C in tsV-infected cells became
associated with the lightestcellular fraction and later with the released virions.
In contrast, M protein, synthesized at 39.6C in tsIII-infected cells, was not
incorporated into the virions after shift down. These datastronglysuggest,first,
that M protein isencodedbythe vesicular stomatitisgenem, andsecond, that
incorporation ofG proteininthe lightest cellular fraction isanecessary step of
vesicular stomatitis maturation. This step isimpairedby tsV mutations.
The vesicular stomatitis virion contains at
least fiveproteinsdesignatedaccording to
Wag-ner et al. (23): L, G, N, NS, and M (12, 24).
These proteins constitute the major proteins
found in cytoplasmic extracts ofinfected cells
(13), thoughadditional viralminorcomponents
maybe present (1, 19, 20, 25).
These extracts can be fractionated by
high-speed centrifugation into soluble andinsoluble
components.NS and half of the Narerecovered
in the soluble fraction, and the majorityofM
and G and halfof the N are recovered in the
insoluble one (25). Association ofviralproteins
with cellular insoluble components has been
investigated by several methods (4-6,8).
Wag-ner et al. (22) observed an unequal distribution
ofthe viral proteins among four cellular
frac-tions which can beseparated on discontinuous
sucrose gradients. The lightest one contains
mainly protein G, whereas protein N is the
major viral component of the third fraction.
Inthispaper we show that
temperature-sensi-tive (ts) mutations classified in
complementa-tion group V (10, 11) specifically affect the
association of the G protein with the lightest
cellularfraction atthenonpermissive
tempera-ture.
(This work is part of a thesis [Doctorat
d'Etat]).
MATERIALS AND METHODS
Virus,cells,and media.Wild-typets+strainand the following spontaneous ts mutants of vesicular stomatitisvirus, serotype Indiana,describedby Lafay (15-17) and Flamand (10), have been used: tsV 44,
tsV45, tsV 57, tsV110,and
tsMf
89.Chickenembryocellsweregrown inEagleminimal essential medium buffered with Tris and
supple-mented with 6% calf serum and 8% tryptose phos-phate.
Chemicals. Calf serum, Eagle minimal essential
medium, and special Eagle minimal essential me-diumwithout leucinewerepurchasedfrom Laborato-ries Eurobio. Ultrapure Tris was obtained from
Schwartz/Mann. [8H]leucine (30 Ci/mM) and
[14C]amino
acid mixture(50mCi/mg)wereobtainfedfrom Commissariat a l'Energie Atomique (CEA-France). Actinomycin D was a generous gift of Merck, Sharp and Dohme (French branch).
Labeling of chicken embryo cells orvirus. Mono-layers of chicken embryo cells in 9-cm petri dishes were infected at a multiplicity of 40 to 150PFU/cell. Virus was adsorbed for 45 min at 17 C, and the cells were incubated at 30 C in Eagle medium. Ac-tinomycin D (1
lAg/ml)
was used when themultiplicity ofinfectionwas less than 100PFU/cell. Three hours 1220on November 10, 2019 by guest
http://jvi.asm.org/
after infection, the medium was removed, and
pre-warmed special minimal essential medium without unlabeled leucine and containing 10 uCiof ['Hlleu-cine perml wasadded. Plates wereimmediately set either at 39.6 or at 30Cforthe labelingperiod.
Cell fractionation. Whereas four to eight petri dishes ofcellsaresufficient to give enough radioactive
material, in order to obtain visible light-scattering
bands, carrier unlabeled cellswere added. For this reason, 15 monolayers were used foreach gradient.
The procedure of fractionation was basically that of
CaliguiriandTamm (3), already appliedtovesicular stomatitis virusby Wagneretal. (22).Afterwashing
with cold Tris-buffered saline, cells were scraped,
centrifuged, washedagain, and recentrifuged at 1,500 x g for 5 min.The pelletwassuspended in 5ml of reticulocyte standard buffer, consisting of 0.01 M
Tris, 0.01 M NaCl, and 0.0015 M MgCl, (pH 7.4). Afterallowingthe cells to swell for 20 min, thecells weredisrupted with20 strokes of a Dounce
homoge-nizer. Nuclei and large celldebriswere removedby centrifugationat800 x gfor 5 min. Thecytoplasmic suspension, containing membrane fragments, was
fractionated by centrifugation in a discontinuous sucrosegradientofthefollowingcomposition: 3 ml of 60% sucrose in reticulocyte standard buffer, 7 ml of 45%, 7mlof 40%, 10mlofthecytoplasmicsuspension in30%sucrose, 7 ml of 25%,and3 ml ofreticulocyte standard buffer. Gradientswerecentrifugedat86,000
xgfor 18 h inaSpincoSW27rotor. Four light-scat-tering bands appeared at the interfaces of the suc-cessive layers, except between the 0 to 25% layers. The bands were collected from the side of the tube with a syringe. Thepelletfraction was also recovered. Each fraction diluted withreticulocytestandardbuffer
wasrepelleted by centrifugationat80,000 x g for 90 minin aSpincotype 30 rotor.Solubleproteinsdid not migrate in the gradient and were recovered by
tri-chloracetic acid precipitation from the 30% sucrose
layer andfromthesupernatantsoftherepelleted frac-tions 1and2.
Extractionandanalysisofproteins.Cytoplasmic extracts and virions were treated by the Maizel
method (18). After adding0.1 vol of glacial acetic
acid, thesuspensionwasmade 0.5M with respectto urea and1%withrespecttosodiumdodecylsulfate.
After incubation for 1 h at 37 C, samples were
dialyzedovernightatroomtemperatureagainst 1,000
vol of 0.01 M phosphate buffer (pH 7.2) containing
0.1%sodiumdodecylsulfate,0.1%2-mercaptoethanol,
and 0.5 M urea.Cytoplasmicsolubilizedproteinswere
mixedbeforeelectrophoresis with10ilitersofmarker
"ICproteins extracted frompurifiedvesicular stoma-titis virions (21). Electrophoresis was performed as
previously described (16), except that a higher gel concentration (7.5% acrylamide) was used. Frozen
gels, sliced into 1-mm disks, weredepolymerized in 0.2 ml of 60 vol ofH,O,at60C for4hpriorto the addition of Bray scintillation fluid (2). Vials were counted in an Intertechnique SL40 assisted by a double-labelingcomputer program.
RESULTS
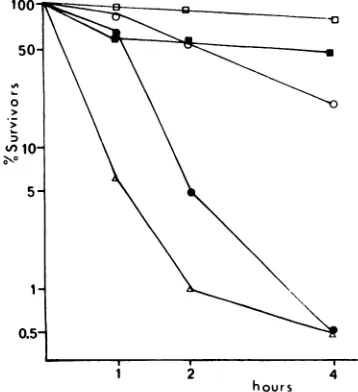
Thermolability.
Amongall thets mutantsofthe Orsay collection, the virions of tsV 45 and
tsV 110arethemorethermolabileinvitro. Both
belong to complementation group V. Thermal
inactivation of tsV 45 has already been
de-scribed by Deutsch and Berkaloff (9). Thetwo
othermutantsclassified ingroupV, tsV44and
tsV 57, show the same thermolability as the
reference wild-type strain (Fig. 1).
Viral proteins in whole cytoplasmic
sus-pensionof infected chicken embryo cells. The
distribution in asodiumdodecyl
sulfate-polya-crylamide gel of the viral proteins synthesized
at 39.6C in cells infected either with the
wild-typeorthe tsV 45 strains are shown in Fig.
2. As already noted by Printz and Wagner (21)
and by Wunner and Pringle (26),nosignificant
differences appear between the two profiles.
Under these conditions, proteins N and NS
migrate very closely and are not generally
resolved as twowell-separated peaks. In Fig. 2
the NSpeak is separatedby only one point from
tsV 45 and is notdistinct from the N peak for
the wild type. Such a difference is not
signifi-cant. As noted by Kang and Prevec (13), the
virion marker G protein migrates slower than
the intracellular G. Compared to results
ob-tained at 37 C with L cells (13, 25) or BHK-21
cells (26), we regularly find a smaller relative
amount of M protein. Cells infected at 39.6 C
with tsV 57, tsV 110, and tsllI 89 give patterns
identical to those ofFig. 2.
Distribution of viral proteins in cytoplas-mic fractions from infected chickenembryo
-0
1 2 4
hours
FIG. 1. Thermolability of tsV mutant strains.
Samples (1ml) of fresh virus stocks wereimmersed in awater bath at 40 C 4 0.1 andperiodically assayed
for survival of plaque-forming ability on chicken
embryo cells at 30C. Symbols: U, ts+; 0, tsV44; A.tsV45;0,tsV57;0,tsVlIO.
1221
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.498.259.438.408.604.2]LAFAY
peratr nciknebyel
infce
ihrwtL G
the ts+ wild-type or the tsV 45 strains. Chicken
embryocellsareinfectedatamultiplicity ofinfection
of80PFUforthe wildtype(a_0)andof120PFUfor
tsV45(O--O).Afteranad-sorption period of45min
at 17C, cells were incubated at 30C for 3 h in
medium without (tsV 45) or with (ts+) I jug of
actinomycinD per mi. [3H]leucine was then added
andtheinfectedcellswerefurtherincubatedat39.6C
for3h.Cellswerethenwashed,scraped,pelleted,and
disruptedinreticulocytestandardbuffer.Cytoplasmic
extractswerepreparedasdescribed inMaterials and
Methods. Electroptwre8i8 wasperformed for5 h on 7.5%6sodiumdodecyl -sulfatepolyacrylamidegels.The arrows show thepeakg position ofthe
"4C-coelectro-phoresedvesicularstomatitisstructuralproteirs.
cells.Thedistributionof viral
proteins L, G, N,
and
M,
synthesized
at39.6C in fivecytoplas-mic fractions of infected cells
separated
on adiscontinuoussucrose
gradient,
isshown inFig.
3. In the
wild-type
infection, Gprotein
is themajor
viral component of band 1, whichcon-tains 20% of theradioactive-insolubleG
protein.
N
protein
isregularly
presentinsmallamounts,whereas M
protein
isnotalways
found.G is alsopredominant
inbands 2 and3, wherethe threeother
proteins
arealways
found. N ispredomi-nant in band 4 and in the
pellet, which,
together,
containtwo-thirds of theradioactivity
associated with sedimentable material. We do
not find M in the
pellet.
The distribution of eachprotein
inthe different fractionsand of thedifferent
proteins
in each fraction isgiven
inTable 1, which includes results for soluble
material.
Extracts of cells infected either with ts+ or
with tsV45, tsV57,ortsV 110 andincubatedat
30Cgivesimilar distribution.
Strikingly
differ-entpattemsareobtained for G
protein
synthe-sized at 39.6C in cells infected
by
the tsVmutantstrains, tsV110
(Fig.
3), tsV45(Fig.
6),and tsV57 (not shown).
Very
little Gprotein
isfound in band 1, andthe proportion of G protein
in band 2 is often reduced. Results obtained
from cells infected at 39.6 C by the
tsllI
89strain aresimilar tothose observed with the ts+
wildtype (Table 2). Thus, it would appear that
the abnormal distribution of G protein is a characteristic of group V mutants.
Distribution of viralpulse-labeledproteins
in cytoplasmic fractions. In order to provide
information on the virus biogenesis, we have
studied the evolution of ts+ wild-type, viral pulse-labeled proteins in the cytoplasmic
frac-tions of adiscontinuous sucrose gradient.
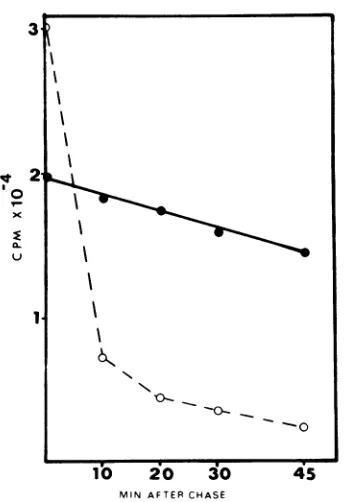
At the end of a 10-min labeling period with
['H]leucine (10
,uCi/ml),
the majority of theradioactivity found inthe cells is not
trichloro-acetic acid-precipitable. This soluble
radioac-tivity decreases veryrapidlyduring the first10
minofincubation in Eaglemedium containing
100
lAg
ofnon-radioactive leucine per ml. It isclear from Fig. 4 that the bulk of the soluble radioactivity in the cells at the end of a pulse is
notultimately incorporated in thetrichloracetic
acid-insoluble material, attesting to the chase efficiency.
Fractionation in sucrose gradients of
cyto-plasmic extracts was performed immediately
afterpulse-labelingfor 20 min(to) and after 20
(t,0)
and 45 min(t4,)
ofincubation at 37 C inchase medium. The virions presentin the
cul-ture supematant and the soluble proteins
re-covered from the gradient were also analyzed.
The results are summarized in Fig. 5. The virion-associated radioactivity increases from 5%ofthe totallabelattoto15%att20 and 33%
at
t4,,
but the kinetics of incorporation ofradioactivity into the virions is very different
from oneprotein to another. Half of thelabeled
Mprotein isalreadyincorporatedinto virions at
to, 80% at t2o, and almost 100% at
t45.
Nodetectable labeled G protein is found in the
virions attoandt2oandonly12%isfoundat
t45.
The proportion oflabeled Gprotein increasesin
band 1from2%attoto 4%at
t,o
and10% att43.
Nosignificant variation in the proportion ofGis
observed inthepelletand infractions 2, 3,and
4.Solubleproteins, whichareessentiallyN and
NS,decreasefrom 20% attoto5 to 6%att2oand
t,4.
Concomitantly, the proportion of N labelincreases first in the pellet and later in the
virions.
Distribution among cytoplasmic fractions
of the tsV viralproteinssynthesizedat 39.6 C
after shift down to 30 C. Cells infected with
tsVmutants wereincubatedat39.6 Cfrom3 to
6 h after infection inmediumcontaining 10 uCi
of [3H]leucine
per
ml.The distribution of viral1222 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
lb 20 3b 40 1b 2'0 30
FRACTIONS 40
ib 20 306 4'0
2
2
x
10,
FIG. 3. Distribution of viral proteins in cytoplasmic fractions ofinfected cells separatedondiscontinuous
sucrosegradient. Aftera45-minperiod of adsorptionat17C, infected cellswereincubatedfor 3 hat30 C in thepresence of actinomycin D(1 pg/ml). ['HJLeucine (10/Ci/ml) was then added, and cells were further
incubatedfor 3hat39.6C. The cellswere thendisrupted andcentrifugedat800xgtoremovenuclei. The supernatantwascentrifugedfor 18hat86,000xginadiscontinuous0to60%sucrosegradient. Light-scattering
bands, locatedattheinterfaces of the successivesucroselayers, werecollected andpelleted by centrifugationat 80,000 xgfor 90 min. Fraction1 is locatedatthe25to30%interface; fraction 2atthe 30to40%interface; fraction 3atthe 40to45%interface, and fraction4atthe 45to60%interface.Left panel: wildtype(multiplicity
of infection [MOI]- 60); middlepanel: tsIII89(MOI - 30);rightpanel: tsV 110(MOI = 40).
proteins labeledat39.6Cwasanalyzedafter0,
20, and 40 min ofincubation at 30 C in
non-radioactive chase medium (Fig. 6). Duringthe
labeling period, therewas noreleaseofPFUto
the supernatant. After the temperature shift
down, virion releasebegan within 30 min, and
extracellular particles accumulated rapidly (Fig. 7).
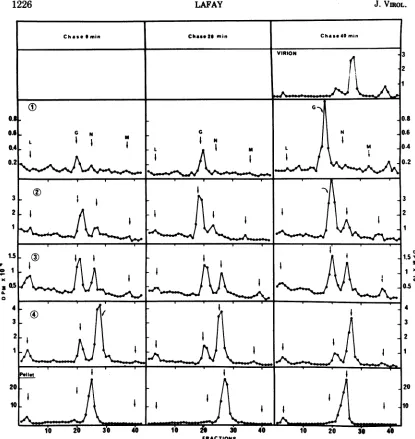
The maturation of G protein synthesized at
nonpermissive temperature during the chase
period at 30C is summarized in Fig. 6. As
alreadyseen,verylittlyradioactiveG ispresent
in band 1atthe end of the 3-hlabeling periodat
39.6 C. The amount of labeled G in band 1
increasesprogressively during postincubationat
30C andappearsinvirions.Thus,the Gprotein
synthesized at the nonpermissive temperature
in cellsinfected bytsV mutantsisable, aftera
temperatureshiftdown, tobeincorporatedfirst into thecellularfraction 1 and later into virions.
Appearance of proteins synthesized at
39.6C in taIII and tsV virions after shift
down at30 C. In order tosee ifradioactive G
protein synthesized atthe nonpermissive
tem-perature is normally incorporated into virions after a temperature shift down, we have
com-paredthe 'H-C140ratio for each structural
pro-tein ofgroupsmandVtsmutants. ['H
]leucine
0;
WIC
a.
I
2
5
I~~~~~
Pellet~~~~~~~~~~~~~~~~~~~~~~~~~~~~
20
____ . . * . . ~~~~~~~~~~~~~~~~~~.1
1223
I I
I I
-4k-I
4
0
I v I
(1)
I
I
I
A
on November 10, 2019 by guest
http://jvi.asm.org/
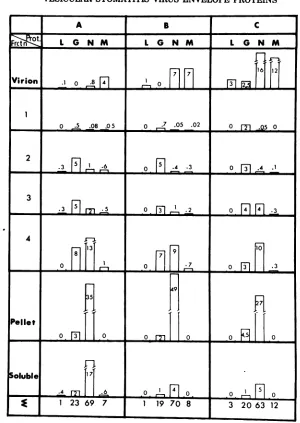
[image:4.498.39.452.48.406.2]TBmLE 1. Distributionoftheradioactivityassociated withG,N(+NS forthesolublefraction),andMproteins in thefractionsisolatedfromadiscontinuous sucrosegradienta
Protein"
G N M Labelineach
Band fato
Distribution Proteinin Distribution Proteinin Distribution Proteinin fraction (%) in sixfrac- eachfrac- in six frac- eachfrac- insix frac- each
frac-tions(%) tions(%) tions(%) tion(%) tions(%) tion(%)
1 19 95 0.2 2 5 3 6
2 27 58 6 33 36 9 13
3 7 61 1 28 10 11 3
4 22 31 17 60 50 8 20
Pellet 8 18 14 82 NDC ND 12
Soluble 16 9 61 91 ND ND 47
aChickenembryocells wereinfectedwith wild type. TheexperimentalprotocolisgiveninthelegendtoFig. 3.
bPercent oflabel in eachprotein: G,27;N, 70;M, 3.
cND, Notdetectable.
3
°t
20
:E~_
~
10 20 30 45
[image:5.498.72.246.324.575.2]MIN AFTER CHASE
FIG. 4. Efficiency ofthe chase medium. Cells
in-fectedwith ts+ wildtype (multiplicityofinfection = 100) were incubated at 37 C. ['HJleucinewasadded 4 hafterinfection. Ten minutes later, cells were washed severaltimeswith cold l7risbuffer andcoveredwith
Eaglemediumsupplementedwith 100 sgofunlabeled
TABLE 2. DistributionofGproteinsynthesizedat
nonpermissivetemperature indifferent cytoplasmic
fractionsa
Complementationgroup
Band ts+ tsV 45
ts1189 tsV110
1 2 1 2
1 18 23 18 2.5 2 3
2 38 32 31 17 22 40
3 11 9 20 12 9 14
4 17 26 17 31 33 30
Pellet 17 10 17 37 33 13
Fractionb 35 35 25 16 25 15
aExpressed as the percentage of G in each frac-tion. Experimental procedures were as described in
legendtoFig. 3.
b Fractionofthetotalradioactivity incorporated in the viral proteins associatedwith G.
leucine per ml. The zerotimesample was takenat this point, and subsequent samples were taken at 10-minintervals. Eachsamplewaswashedcarefully,
and cells were treated with 5% trichloroacetic acid. The amounts ofradioactivity in the trichloroacetic
acid-soluble (0- -0)andprecipitable (0-O)
frac-tions were determined andplotted.
1224 LAFAY J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.498.264.456.345.553.2]A B C
~~0t L G N M L G NM L GN M
VIrin
.1
0_
.8
[1 1
jjJJj
0
3J
jfJj
0
L..08
.05 0 7 .05 .02 0 M.05
02
3
-~
~~~
]mM3n4 .2 0F4]
Fl34
1171
Pelt
12369
71
19708
3
206312FIG. 5. Distributionof theradioactivityassociated withL,G,N(+ NS forthe solublefraction),and M in the
fractions (frctn) ofa sucrosegradient and in the virions. Eachfigurerepresents thepercentage ofthe total
radioactivity recovered in eachgradient. Chickenembryocellswereinfected with wildtypeandpulse-labeled for20min. at 4hpostinfection. Incubationtemperature was 37C.(A)Chase0min;(B)chase20min;(C) chase
45min.
waspresentduring incubation at39.6 C (3to6
h postinfection), washed outjust before shift
down at 30C, and replaced by a
[14C]amino
acid mixture. Extracellular virions were
col-lected after 1.5h of additional incubation.The
tsM mutant was abettercontrol than ts+ wild
type, because maturationwas blockedforboth
groups at 39.6C and began within the same
time after shift down to 30C. The relative
incorporation into virions of proteinsG, N, and
M synthesized at the nonpermissive (3H la-beled) and at the permissive ("IC labeled)
temperature is shown inFig. 8.The 3H-14C
ra-tiosinproteinsG andN areidenticalforthetwo
mutants. Thus, the G protein synthesized at
39.6C in cells infected by tsV mutants is
therefore incorporated into virions in normal
amounts after shift down at 30C. The M
protein is synthesized at 39.6 C by tsII
89-infected cells andeasily detectedincytoplasmic
14, 1225
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.498.90.390.61.484.2]Chase*min0 ChaseZs min Chase 40min VIRION
______ L
3 2 1
G
I N
L I m1
I
I
N L M
I i
-
s
w I I*I||, t
I
I~
II I I $
I.III
-I~~~~~~~~~
I
I-3
2
[image:7.498.48.463.52.491.2]FRACTIONS
FiG. 6. Evolution afterashift downat30 Cof the distributionamongcytoplasmic fractions of the tsV45 viralproteins synthesizedat39.6 C. Cellswereinfected with tsV45 (multiplicity of infection =120) in thesame
wayasdescribed in Fig.2.After the 3-h labelingperiodat39.6C, the cellswerefurther incubated in unlabeled
chasemediumat30Cfor20or40minbefore collection. Labeled virus, released into thesupernatantonly after 40 min of incubationat30C, were also collected. Fractionation was carried outon sucrose discontinuous
gradientsasdescribed in Fig.3.
extracts (Fig. 3), but it is not recovered into
virions produced after the temperature shift
down,as already reportedfor tsM 23(16). The
high valueofthe 5H-14C ratio ofM protein in
the tsV 45virions is also regularly observed in wild-type infection for such labeling periods. This maybe duetothe fact that protein M is
incorporated into virions very quickly after
synthesis, as already shown in Fig. 5 and by
Kangand Prevec (13).
DISCUSSION
The viralproteins L, G, N, and M arefound
inthesameproportion in the whole cytoplasmic
extracts from cells infected at permissive or
nonpermissive temperature either by the
wild-3
2
0.8 0.6 0.4 0.2
1.5
I1
cx30.5
a.
3 2
lx
0,
0.5 '
4 3 2
1
20
10
el
1226 LAFAY J.-VItOL.
I
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
VESICULAR STOMATIS VIRUS ENVELOPE PROTEINS
1op
I10.6
a.
105
3.1&
103
AP- ---_. . . .-A
1 2 3 4 5 6 7 8
[image:8.498.49.236.52.253.2]HOURS
FIG. 7. Growthcurvesat30, 39.6,and 30-39.6-30C shift of mutant tsV 45. After a 45-min period of
adsorptionat17C, cells infected with tsV45 (multi-plicity of infection=5)werewashed with Tris buffer.
Fresh medium wasadded and petri disheswereheld
at 30 or 39.6C. One hour later the medium was
discarded andreplaced by prewarmed medium; 3 h after infection oneplateat 30Cwas transferred to 39.6 Cfor 3 h andfinally set back at30C. At the indicated times samples were taken for titration.
Symbols: 0,30Cthroughout; A,39.6Cthroughout; 0, 30-39.6-30C; -, 30C incubation; - -, 39.6C
incubation.
typeorby tsmutantsofgroupsHI and V. This
is not unexpected, since thermosensitivity is
usually theconsequenceofmis-sense mutations
which havenoeffectonthegenetranslationand
whose expressionatthepolypeptide level isthe
substitutionofoneaminoacidbyanother. Such
a substitution, which could affect the protein
configuration and function at nonpermissive
temperature, is not expected to modify the
migration properties of the polypeptide in a
sodiumdodecylsulfate-polyacrylamide gel.
We interpret the results of our pulse-chase
experiment with wild-type-infected cells as
meaningthat the vesicular stomatitis Gprotein
migratesfrom the heaviestfractionstothe band
1 before being incorporated into the virions.
Migration for influenza glycoproteins through
thecellular fractionsseemsquitesimilar(7, 14).
As already notedby KangandPrevec (13), we
observe that newly synthesized M protein is
very quickly incorporated into the virions. In
accordwithWagneretal.(25), wefound thatN
protein migratesfrom the soluble tothe
heavi-estfractions, probablyas aresultof
nucleocap-sid formation.
Thiswork demonstrates that association ofG
protein with the lightest insoluble fraction
which can be resolved by discontinuous sucrose gradients is blocked atthenonpermissive
tem-perature by all four mutations classified in
complementation group V by Flamand (10). It
isimportant to notice that identical results are
obtainedeither withgroup V mutants whichare thermolabile in vitro or with those which are
not.The simplest interpretation of these
obser-vations is to assume that incorporation of G
protein in cellularfraction 1 is a necessarystep
ofvesicular stomatitis virus maturation.
After transfer tothe permissive temperature,
the G protein synthesized at the nonpermissive
temperature becomes associated with the light
cellularfraction and released virions. This result does not allow a choice between two hypotheses:
either gene V is the structural gene for the G
protein or gene V encodes a minor protein
necessary for G incorporation in cellular
frac-tion 1. If gene V isthe structural gene of protein
20 '5 5-~~ ~~~~~I 1
a.9
o U
a.
?: 0
5 I
16 20 30 410
FRACTIONS
FIG. 8. Polyacrylamide gel electrophoresis oftsIII 89and tsV 45virionsreleased afterashift down at 30C. Cells infected with tsV 45 (multiplicity of infection [MOIJ = 20) or tsffI89(MOI = 15) were
incubatedat 39.6C from 3 to 6 h afterinfection in
Eaglemedium containing2.5MCiof[3H]leucineper
ml. Cells were washed, and minimal essential
me-diumdiluted10-foldinTris-saline containing2.5uCi
of ["'CJamino acid mixture per ml and 100 gg of
unlabeled leucine per ml was added. Virions were collectedafter1.5hofincubationat 30C andpelleted
at80,000 x gfor60min,andproteinswereextracted as described in Materials and Methods. Top panel:
tsV45; lowerpaneltsHI89.Symbols:*, 3Hlabel;0; "4C label.
14, 1227
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.498.257.439.293.513.2]J. VIROL.
G, we mustconclude that alterations produced by the four tsV mutations in the G protein
synthesized at 39.6C are reversibleupon shift
downor, alternatively, that the Gprotein
syn-thesized after shift down allows G protein
synthesized at 39.6 C to be incorporated into
fraction 1.
Protein M, synthesized at39.6 C bytsIII 89,
isreadily detectedincytoplasmicextracts, but,
in contrast to group V mutants, it is not
incorporated into virions after transfer to the
permissivetemperature. These results strongly
confirmourprevious hypothesis thatgene III is
thestructural geneofthe M protein (16, 17).
ACKNOWLEDGMENTS
Thisresearchwassupported by the Centre National dela
Recherche Scientifique through the L. A. 86and 136, the
Fondation pourla Recherche M6dicale Francaise, and the Commissariat al'Energie Atomique, Saclay,France.
I thank Ph.Vigier and A. Berkalofffortheirinterest inthis
work and formanyhelpful discussions. I thank L. Prevec for
criticism ofthe manuscript. I am greatly indebted to B.
Jaillard, D. Pese, and A. Friedman for technical assistance. LITERATURE CITED
1. Bishop, D. H. L., and P. Roy. 1972. Dissociation of vesicular stomatitis virus and relation of the virion proteinstothe viral transcriptase. J. Virol. 10:234-243.
2. Bray, G. A. 1960. A simple efficient scintillator for counting aqueous solution in a liquid scintillation
counter.Anal.Biochem. 1:279-285.
3. Caliguiri, L. A., and I. Tamm. 1970.The roleof cytoplas-mic membranes in poliovirus biosynthesis. Virology 42:100-122.
4. Cartwright, B. 1973. The distribution of Virus proteinsin
BHK21cells infected with Vesicular Stomatitis Virus (Indiana C). J. Gen. Virol. 21:407-411.
5 Cohen, G. H., P. H. Atkinson, and D. F. Summers. 1971. Interaction of Vesicular Stomatitis Virus structural proteins with HeLa plasma membranes. Nature N. Biol. 231:121-123.
6. Cohen, G. H., and D. F. Summers. 1974. In vitro association of Vesicular Stomatitis Virusproteins with purified HeLa and erythrocyte plasma membranes. Virology 57:566-569.
7. Compans, R. W. 1973. Influenza virus proteins. II.
Association withcomponentsof thecytoplasm.
Virol-ogy51:56-70.
8. David, A. E.1973.Assembly of the vesicularstomatitis virusenvelope:incorporationofviralpolypeptides into the hostplasma membrane. J. Mol. Biol. 76:135-148.
9. Deutsch, V., and A. Berkaloff.1971.Analyse d'unmutant
thermolabile du Virus de la Stomatite V6siculaire. Ann. Inst.Pasteur(Paris) 121:101-106.
10. Flamand, A. 1970. Etude genetique du virus de la stomatitev6siculaire: classement de mutants thermo-sensiblesspontanes engroupe de complementation. J. Gen. Virol. 8:187-195.
11. Flamand, A., andC. R. Pringle. 1971. The homologies of spontaneous and induced temperature-sensitive mu-tants of vesicular stomatitis virus isolated in chick embryo andBHK21cells. J. Gen. Virol. 11:81-85. 12. Kang,C. Y., and L. Prevec. 1969. Proteins of vesicular
stomatitis virus. I.Polyacrylamidegel analysis of viral antigens. J. Virol. 3:404-413.
13. Kang, C. Y., and L. Prevec. 1971. Proteins of vesicular stomatitisvirus.Ill. Intracellular synthesisand extra-cellularappearance of virus-specific proteins. Virology 46:678-690.
14. Klenk, H. D., W. Wollert, R. Rott, and C. Scholtissek. 1974. Association of influenza virus proteins with cytoplasmic fractions. Virology 57:28-41.
15.,Lafay, F. 1969. Etude des mutants thermosensibles du virus de la stomatite v6siculaire: classification de quelques mutantsd'apres descriteresde fonctionne-ment.C.R.Acad. Sci.Ser.D 268:2385-2389. 16. Lafay, F. 1971. Etude des fonctions du virus de la
stomatite vesiculairealter6espar une mutation ther-mosensible: mise enevidencede laprot6inestructurale affecteepar la mutation ts23. J.Gen.Virol.13:449-453. 17. Lafay, F., and A. Berkaloff. 1969. Etude des mutants thermosensibles du virusde la stomatite vesiculaire: mutants de maturation. C. R. Acad. Sci. Ser. D 269:1031-1035.
18. Maizel, J. V. 1966. Acrylamide gels electrophoresis by mechanical fractionation radioactive adenovirus pro-teins.Science151:988-990.
19. Mudd, J. A., and D. F.Summers. 1970. Protein synthesis invesicular stomatitis virus-infected HeLa cells. Virol-ogy42:328-339.
20. Obijeski, J. F., and R. W. Simpson. 1974. Conditional lethalmutants of vesicular stomatitis virus.II. Synthe-sis of virusspecificpolypeptidesinnon-permissivecells infectedwith"RNA-"host-restrictedmutants. Virol-ogy 57Q69-377.
21. Printz, P., and R. R. Wagner. 1971. Temperature-sensi-tive mutants of vesicularstomatitis virus: synthesis of virus-specificproteins. J. Virol. 7:651-662.
22. Wagner, R. R., M. P. Kiley, R. M. Snyder, and C. A. Schnaitman. 1972. Cytoplasmic compartimentsliza-tion of the protein and ribonucleic acid species of vesicular stomatitis virus. J. Virol. 9:672-683. 23. Wagner, R. R., L. Prevec, F. Brown, D. F. Summers, F.
Sokol, andR.MacLeod. 1972. Classificationof rhab-dovirusproteins: aproposal.J. Virol. 10:1228-1230. 24. Wagner, R. R., T.C. Schnaitman, andR. M. Snyder.
1969. Structural proteins of vesicular stomatitis vi-ruses.J. Virol. 3:395-403.
25. Wagner, R. R., R. M. Snyder, and S. Yamazaki. 1970. Proteins of vesicuar stomatitis virus: kinetics and cellular sites ofsynthesis. J. Virol. 5:548-558. 26. Wunner, W. M., and C. R. Pringle. 1972. Protein
synthe-sis inBHK21 cells infected with vesicularstomatitis virus. I.tsmutants of theIndiana serotype. Virology 50:829-840.
1228 LAFAY
on November 10, 2019 by guest
http://jvi.asm.org/