THESES, SIS/LIBRARY A.G. MENZIES BUILDING N02 Australian National University Canberra ACT 0200 Australia
USE OF THESES
This copy is supplied for purposes
of private study and research only.
Passages from the thesis may not be
copied or closely paraphrased without the
written consent of the author.
THE AUSTRALIAN NATIONAL UNMRSITY
Telephone: +61 2 6125 4631 Facsimile: +61 2 6125 4063
()
' (
,,J
COMPETITIVE INTERACTIONS BETWEEN THE LARVAE OF HELIOTHIS ARMIGERA (HUBNER) AND
HELIOTHIS PUNCTIGERA WALLENGREN (LEPIDOPTERA: NOCTUIDAE)
by
Suzanne Maree Stanley B.Sc.(Hons.) W.A.
Department of Population Biology, Research School Biological Sciences, Austrqlian National University.
A thesis submitted for the degree of Doctor of Philosophy of the Australian National Univ~rsi~y.
DECLARATION
I declare that the investigations described in this thesis are wholly the original work of the Author, except where specifically acknowledged in the text.
Signed
)
FRONTISPIECE A. Fifth instar larvae Top: Heliothis armigera
Bottom: Heliothis punctigera
---~·---illiiili--B.
c.
FRONTISPIECE B. The arrangement of cotton plants in the experimental cell used to analyse larval feeding preferences (Chapter 5). C. The arrangement of cotton plants in the experimental cell
'J'J\BU: OF CONTENTS
ACKNOWLEDGEMENTS ABSTRACT
CHAPTER 1
1.1
1. 2 1. 3 CHAPTER 2
2.1 2.2
2.3 2.4
CHAPTER 3
3.1 3.2 3.3 3.4 3.5 3.6 3.7 3.8 3.9 INTRODUCTION
Review of the approaches used in the study of competition
Background to the study of competition between
.!!:
armigera and.!!.-
punctigeraOt\tline of the approach used in the present study
GENERAL CULTURE TECHNIQUES The animals used
Methods for culturing laboratory stocks of Heliothis 2.2.1 The environment
2.2.2 Rearing procedure
Identification of
!!.·
armigera and H. punctigera The culture of cotton plants2.4.1 The environment 2.4.2 The cotton plants 2.4.3 Pest control
A LABORATORY STUDY OF INTERFERENCE BEHAVIOUR IN H. ARMIGER.A AND H. PUNCTIGERA
Introduction Methods
3.2.1 Experimental design
3.2.2 The analysis and treatment of data Section A: Intrnspecific Interference Behaviour Comparison between
!!.·
armigera and.!:!.·
punctigera in the standard system3.3.1 Results 3.3..2 Discussion
The effect of larval density 3.4.1 Methods
3 . 4 . 2 Results 3.4.3 Discussion
The effect of dietary differences 1.5.1 Interference behaviour
3.5.l.l Methods
3. 5. L ~1 Resu1.ts
3.5,2 Davel{)pment
3.5.2.1 The effect of diet on development 3.5.2.2 The developmental consequences
of
cannibalism 3.5.3 Discussion
The effect of diet quality on the response to density
3.6.1 Hethods
3.6.2 Results and t:liscussion Summary 0£ intraspecific behaviour Section B: Interspecific Competition
The role of interference behaviour in interspecif.ic competition
3.8.1 Methods
3.8.2 Results and discussion
'.l
CHAPTER 4
4.1 4.2 4.3
4.4
4.5
CHAPTER 5
5.1 5.2
5.3
5.4
CHAPTER 6
6.1 6.2
6.3
6.4
DEVELOPHENT OF H. ARMIGERA AND H, Pl'NCTJ:GERA ON COTTON PLANTS
Introduction General techniques Development
4.3.1 Methods 4. 3. 2 Analysis 4.3.3 Results
4.3.3.1 Larval development 4.3.3.2 Pupal development 4.3.3.3 Pupal size
4.3.4 Discussion Instar duration
4.4.1 Methods
4.4.2 Results and discussion
Consumption of fruiting forms by larvae
4.5.l Methods
4.5.2 Results and discussion
Discussion and conclusions
FEEDING BEHAVIOUR OF H. ARHIGERA AND H. PUNCTIGERA LARVAE
Introduction Methods
5.2.1 Feeding behaviour of second to fifth instar larvae
5.2.2 Feeding behaviour of first instar larvae Results
5.3.1 Feeding preferences
5.3.1.1 Comparison between form classes for each instar
5.3.1.2 Comparison between instars and species with regard to preferences for each form class
5.3.2 Degree of overlap between the feeding niches of each species
Discussion
COMPETITION BEt\\iEEN R. AR.NIGERA AND .!:!. • PUNCTIGERA ON COTTON PLANTS
61 62 62 63 65 67 69 71 71 74 76 76 76 73 78 79 81 84 86 86 87 88 88 88 99 100 105
Introducti.m 108
~~tbods 109
6.2.l. The e:xperimental system 109
6.2.2 Experimental treatments 110
6.2.3 Experimental procedure 113
Relationship between survival and food
availability 114
6. 3 .1 Analysis 114
6.3.2 Results and aiscussion 119
6.3.2.1 Survival from hatching to pupation 119 6.3.2.2 Survival from hatching to 100 day°C 119 6.3.2.3 Survival from 100 day°C to pupation 122 Relationship between the outcome of interspecif ic
fj
u
.
"6.5
6.6 CHAPTER 7 REFERENCES APPENDIX 1 APPENDIX 2 APPENDIX 3 APPENDIX 4
APPENDIX 5
Comparison of the performance of each species during intraspecif ic and interspecif ic
competition for food 6.5.1 Analysis
6.5.2 Results and discussion 6.5.2.1 'Young' cotton plants 6.5.2.2 'Old' cotton plants General discussion
DISCUSSION AND CONCLUSIONS
"""""
ACKNOWLEDGMENTS
I wish to thank the members of my supetvisory committee,
Prof. B. John, Dr. J,C. Cullen, Dr. R.D. Hughes and Pr,of, R.O. Slatyer for their guidance and criticisms during this study.
Mr. M.A. Adena and Dr. M.J. Dallwitz provided valuable and much
appreciated advice with regard to the statistical analysis of data and the development of the simulation model, respectively.
Mrs. Betty Lee kindly advised me on pest control measures· in the glasshouse and provided biological control agents whenever necessary.
Many people supplied me with field caught populations of
H. armigera and
.!!·
punctigera to use in the establishment of laboratory stocks; they are acknowledged in detail in the text. In particular, I am grateful to Dr. K.G. Wardhaugh for his help and hospitality during my visits to Narrabri and for permitting me to quote many of his unpublishedobservations.
Finally, special thanks are due to Prof. John and my husband, John Dearn, for their interest, encouragement and helpfulness throughout
=
c!)
\\
i i .
/\.BS TRACT
Experiments were conducted in the laboratory and glass.house to
'•
investigate the role of competitive interactions. between the larvae of Heliothis armigera and !!_. punctlgera (Lepidoptera:Noctuidae) in regulating
the relative abundance of the two species on cotton plants.
Under uniform laboratory conditions !!.· armigera and
!:!.·
punctigera larvae differed in their cannibalistic behaviour; the frequency ofcannibalism in H. armigera was primarily encounter-dependent while in
!!_. punctigera it appeared to be independent of density. When larvae of both species were reared together on artificial diet the survivorship of
H. punctigera was depressed ~ompared to that observed when reared by itself at the same overall densit:y. By contrast, the survivorship of U. armigera was not affected.
Both species required a similar absolute tim~ for development on cotton plants and thus developmental characteristics appear unlikely to affect the outcome of interspecific interference competition in one generation. Nevertheles~, the interference advantage possessed by
!!_. armigera may be enhanced over a number of generations because this species passes through the pupal stage more rapi.dly than!!.· punctigera.
Although the resource utilization patterns of !!.· armigera and H. punctigera larvae on cotton plants overlapped broadly there were
quanlitl!-tive .J:I.fferen~I;'~ in feeding prefereni (.l:S. Leaves were mc:ri.: "H'.:ce::pt.:ltble
to
!!.·
.E..~igera than H. a!7migera and the development of a pref .. :.a forlarge bolls occurred later during the larval period for H. _Eunctigera
than 1f. !1rmi_gera.
Mixed species trials on cotton plants showed that the greater
iii.
0
preferences. Under these conditions aggressive interference behaviour is
directed primarily .:-t conspecific.s. Thus, although.!:!_. amigera is potentall~
able to limit the population density of .!!_. punctigera in a uniform
environment, intraspecifc interference encounters may be more important
0
in regulating the population density on plants. The relevence of these
findings to the regulation of field populations is dis:cussed,
__ ,)
'\)
c-CHAPTER 1 INTRODUCTION
1.
A central problem in ecology concerns the elucidation of the role that biological interactions, such as competition, facilitation and predation, play in limiting the distribution and relative abundance of natural populations. The work described in this thesis involves a study of the significance of one class of these interactions, namely competition, in regulating the dynamics of two closely related species, Heliothis
armigera (Hubner) and H. punctigera Wallengren (Lepidoptera: Noctuidae), which overlap broadly in many aspscts of their ecology.
1.1 Review of the approaches used in the sturly of competition
Competition occurs either when a number of individuals of the same or different species utilise a common resource which is in short supply or, if the resource is not in short supply, when individuals nevertheless harm one another in the process of acquiring that resource (Birch 1957). Evidence of competition is provided by an observed decrease in the relative abundance of one or more species (or genotypes of the same species) due to their mutual presence on a resource. The theoretical mod-::ls of Lotka and Volterra (as discussed in many text books e.g. Krebs 1972, Emlen 1973, Pianka 1974), together with experiments using simple laboratory systems (e.g. Gause 193!1, Park 1948, Ayala 1972.) predict that c:ompetition w:i.11 usually result in the exclusion of one of the spec;ies
(or genotypes) from that particular resource which is limiting. Ir1 the absence of external influences, physical or biotic, these theoretical and
2.
It is necessary to distinguish between two forms of competition
which have different implications for population regulation. The terms
exploitation or scramble competition refer to situations in which the
resource is unavailable to one group of consumers because it has been
utilised by another group. For example, the study of intraspecific
competition in Lucilia cuprina by Nicholson (1954) indicated that the
proportion of larvae which reached a sufficiently large size to result
in a viable pupa declined progressively with increased larval density.
A further increase above a critical density resulted not only in a
greater percent mortality, but also a decrease in the actual number of
adults produc.,..J per food unit. Thus, " ... success is colillnonly incomplete,
so that some, and at times all, .:.if the requisite secured by the competing
animals takes no part in sustaining the population, being dissipated by
individuals which obtain insufficient for survival'' (Nicholson 1954, p20).
Since exploitation competition only occurs during periods of acute
resource shortage, relatively violent and potentially unstable population
oscillations may therefore be expected in systems in which this form of
competition is prevalent, especially if the availability of the resource
to the next generation is depleted. The reproductive potential of
individuals may also be expected to vary between generations, due to
differences in size and development rate.
Int.erference or contest competj tion, on the other hand, results
in individuals being " ... either fully successful or unsuccessful; and
the whole <'.i.mount of the requisite obtained ~ollect.ively by the animals
is used effectively and without wastage in maintaining the population"
(Nicholson 1954, p20). This form of competition is usually behaviourally
or chemically based (e.g. dominance, territoriality, cannibalism,
allelopathy) and may occur in conditions of both actual and potential
resource limitation; in general it acts to reduce the population
3.
Relatively stable and constant populations of both the competitors and
the resources may therefore be expected, with the reproductive potential
of the surviving individuals remaining similar from generation to generation.
Two forms of interference competition relevant to this thesis are
cannibalism, defined as intraspecific predation, and mutual predation
be.tween species belonging to the same trophic level. The most detailed
studies of these interactions have been conducted on flour beetles
(Tribolium) by Park and his colleagues (see review by King and Dawson
1972). Cannibalism and interspecies predation exert major influences on
the equilibrium size of populations in these species, although fecundity
responses to density are also important. Ayala (1970) considers these
interactions as predation rather than competition, where predation is
defined as an animal using another individual as its source of food.
This viewpoint is justifiable in normally carnivorous species, where in a
given species, different ase-classes of a population utilise different size
spectra of food and where juvenile individuals are of a similar size to
the food range of the adult (Bulkley 1970, Thibault 1974, Fox 1975b). In
these cases, adults and juveniles do not compete for the same food,
although the demographic consequences of predation 9n younger
age-classes may pre-empt later competition for limited food supplies (Fox
1975a,b). With herbivores, gramnivores and detritivores, however, all
size classes (at least of the predatory stage) utilise a similar food
spectrum. Here, therefore., the outcome of the pr".datory behaviour
against individuals of the same or different speci• is to reduce
competition for the primary food resource., similar to other forms of
interference behaviour.
There is conflicting evidence with respect to th~ significance of
competition in nature (see review by Connell 1975). Some fi.e.ld
observations suggest that it is a significant factor in popu1ation
4.
Waloff 1968, Lock and Reynoldson 1976), Other obs1;\rvations suggest that ecological homologues are able to co-exist without competition because other factors, such as w,eather and predation, mainta~.n the popul,ation size below the carrying capacity of the resource (Andrewat'tha and Birch 1954, Ross 1957, Connell 1971, Springett 1968, Dayton 1971). In fact, except ior examples of aggressive displacement of one species by another (e.g. Brian 1952, Pontin 1957, Brown 1971), it has been difficult to obtain direct evidence of competition ln nature, largely because of (1) the potential complexity and (2) the evolutionary impli~ations of this form of interaction. 'J:hus;
(1) Most detailed field studies have indicated that, species, which superficially appear to compe~e for limiting resources, actually avoid intense competition because they differ in their utilisation pattern of another resource dimension of the niche1. For example, food resources may be partitionetl according to foraging area (Townes and Townes 1960
[cited in Hiller 1967], Edington and Edington 1972, Cody 1973), foraging time (Fraser 1976), f~od taxon (Reynoldson nnd Davies 1970, Otte 1975, heinrich 1976), food size (Schoener 1970, Pianka 1975, Cody 1974) or a
co1;~t1ination of these characteristics (MacArthur 1958, ·--~fab 1971, Brown and Lieberman 1973). Numerous other examples are detailed in a review by Schoener (1974).
(2) Nstu~al selection may be expected to act on differential
exploitation efficiences to favour the evolution of mor.phological, physiological or behaviou~al traits ~hich differentiate the niches of
1
In th:I.s thesis, the term 'niche' is used in the. sense of Hutchinson
(1958), but with modifications to include the effects of predation
{Connell 1975, Pianka 1976). The niche of a species is perceived as an n-dimensional bypervolume in space, the axes 0£ which define the total anay of var.fables (e.g. food, hnbitat, temperature toletance) which limit the sun~ival of individuals of that species. Because of
the practical difficulties of measuring the probability of persistence, niche measurements are usuall; b~sed on patterns of resource utilisation. The 1fundamental1 niche is that measured in the absence of competitors
and predatt;;.-~, while the 1
11
5.
closely related species and reduce the intensity of competition between theu1. This does not mean that competition is not ~ significant factor; on the contrary, competition between species of different exploitation efficiency probably provides the basis on which resource partitioning evolves. We usually see only the end-point of such interactions, however, and it is invalid to make the reverse assumption that observed differences have evolved as a direct result of competition in the past. Unfortunately, much of the evidence. of regulation through differential efficiences of resource exploitation is necessarily based on this untestable assumption. Examples include observations of two kinds: (i) Niche shifts on islands in the absence of competitors, but possibly also in the absence of predators and with different resource classes (Beauchamp and Ullyot i932 [cited in }filler 1967], Diamond 1970, Terbough and Weske 1975). (ii) Character displacement between overlapping populations, also possibly attributable
to resource and habitat differences (Schoener 1965, 1970, Huey~ al. 1974, Fenchel 1975).
A further complicating factor in the study of competitic;n is that species may broadly overlap in their utilisation patterns over several niche dimensions without being identical in any. Competition may still
be L~portant in these communities but the logistic problems involved in
studying them experimentally are considerable. A broad but incomplete overlap in many dimensions suggests the possibility of complex interactions between the response of individuals to different niche parameters.
McClure and Price (1975) and Rathcke (1976) have attempted to resolve the question of the significance of competition in these s:F .. tuations by adaptir:ig the concept of the limiting simi 1arity of coexisting species (Levins 1968, MacArthur and Levins 1967, May and MacArthur 1972.). They measured the similarity of several species with
'i"\
? 6.
level of similarity at which competition will occur and second, to
determine the degree of dependence between niche dimensions. McClure
and Price (1975) assigned a limiting similarity value of 70% in each
niche dimension, based on observed morphological differences between
coexisting species (e.g. Hutchinson 1959, Sch~ner 1965, Fenchel 1975).
'.
Although these observed differences accord with the theoretical predictionsof the models, the implied assumption remains that such differences are a
direct result of competition. Furthermore, Hespenheide (1973) and
Schoener (1974) showed that, for feeding structures, the minimum
necessary morphological differemce varied with the relative sizes of the
predator and prey species. The rroblem of the degree of independence
between niche dimensions arises because the overall similarity of two
species is obtained by multiplying together the similarities in each
dimension (May 1976, McClure and Price 1975). If, therefore, dimensions
are partially dependent on each other (e.g. temperature and humidity),
the degree of similarity will be ove;-estimated (Pianka 1976). Yet another
problem with this approach has been discussed by several authors (Colwell
and Futuyma 1971, Sale 1974, Schroeder and Rosenzweig 1975). Since
estimates of s:tmilarity are made on the 'realised' niche, it is not
possible to distinguish between the contradictory interpretations that
'~ broad ov.erlaps imply either intense competition or the complete absence
of competition. Similarly, narrow overlaps may either result from
competition or be completely unrelated to it.
An unambiguous method of estimating the intensity and
outcome of competitive interactions involves an experimental comparison
ot the pre-competitive and post-co~petitiva niches of coexisting species.
The approach developed by Connell (196la,b) is one of experimental
perturhation involving the removal from or addition to natural
0
7.
with that of control trials. This provid,es the best measure of the impact of competition on population regulation since all factors, including predation, are considered simultaneously.
Natural communities which have clearly defined boundaries (e.g. marine intertidal zone, freshwater ponds) or in which one of the
interactions uncl~~ investigation occurs between sessile organisms (e.g. plants, barnacles) are amenable to this type of analysis (e.g. Connell
ab
196H; 1970, Pa.lne 1966, 1969, Pu twain and Harper 1970, Reyn0ldson and Bellamy 1971, Dayton 1971). It is difficult, however, to conduct these experiments in communities with ill-defined boundaries and where the
spec~es involved are very mobile (but see De Long 1966, Davis 1973).
I
This is especially the case in territorial animals where interspecif ic interactions may be masked by very strong intraspecif !c interactions (Schroeder and Rosenzweig 1975). The logistic problems of arranging experiments, with replicates in which " ••• all environmental factors, except the one being tested, vary in the same way and to the same degree on experimental and control sites" (Connell 1975, p463), also makes this form of investigation difficult.
It is often advantageous, therefore, to study competition tn the laboratory or in field enclosures where certain variables can be
controlled, despite the problem that potentially i~portant factors, such as predation and migration, are often excluded. While direct
extrapolation to natural conununities is difficult, such studies serve to identify the nature of the competitive interactions, the significance of which can then be te$ted in the field. These typea of studies have been performed on a wide r&~ge of community types, including protozoans (Gill and Hairston 1972), freshwater microcrustacea (Neill 1975), amphibians
(Wilbur 1972) and rodents (Grant 1971, review 1972). A further advantage of laboratory/enclosure experiments is that they permit an examination of the effect of individ~al factors on the outcome of
0
8.
An experimental approach was therefore adopted in the present study, w:: ',:i the aim of first identifying the nature of the competitive interactions between H. armigera and B_. punctigera in the laboratory.
A :;>econd aim of the st:udy was to determine how the competitive
interactions were modified in the more complex environment provided by one of their preferred host plants, cotton (Gossypium hirsutum Linnaeus).
1. 2 Background to the study of competition between H. armisera and H. punctigera
H. Eunctiger~ is endemic to Australia and has been found ,.,, throughout the continent wherever suitable food is available (Common
1953). H. armigera has an Old World dj,stribution and is found
throughout Africa, Europe and Asia, south of the 40° latitude in the northern hemisphere, and also in the Pacific islands of 'Fiji and Torga
(Hardwick 1965). Within Australia, its distribution broadly overlaps that of
B.·
Eunctigera in northern and eastern Australia. Common (1953) regardedl!·
armigera as being mainly limited to within one hundred miles of the coast, but with the subseq11ent introduction of irrigation and cropping, it has become more common in some inland areas (Twine 1974).Both species are broad generalists with respect to their food plants and, again, their host ranges overlap broadly. They have been collected from at least fourteen plant families within Australia, mostly herbaceous and low woody plants;·-b'ut including _fruit trees and vines
9.
The close ecological .similarity of the species is further
indicated by their life history characteristics. After overwintering in a facultative pupal diapause, the adults emerge in spring and pass
through several generations before the population re-enters pupal
,,
diapause in the autumn (Kirkpatrick 1962, Cullen 1969). The number of generations varies geographically between thr.ee and seven, depending on temperature, as does the time of emergence and diapause. Peak egg-laying of each species within different crops coincides with the flowering period
() of plants, although in cotton, which has extra floral nectaries, laying
may connnence soon after seedling emergsnce (Cullen 1969, K.G. Wardhaugh pers. comm.) . Mixed species populations are of ten found (Kirkpatrick 1961
r-,
personal observations). Cotton plants usually start flowering
approximately nine weeks after planting and continue for a further twelve weeks (Reynolds et al. 1975). Oviposition by Heliothis species continues throughout this period (K.G. Wardhaugh pers.comm.). Both species usually lay their eggs about the top 20 cm of the cotton plants, on young leaves and growing tips. The developmental threshold temp'Bratures and the duration of each development stage of each species is similar, with the larvae usually passing through six instars, although this varies with diet quality (Hardwick 1965, Twine 1974).
Cannibalism has been observed in both
.!!.·
armigera andH. punct1gera under laboratory conditions (Cullen 1969, Twine 1971) and it has been suggested that this interference behaviour may be a major factor in intraspecific population regulotion, at least i.n H. armigera
(Twine 1974).
!.!.·
armigera and!.!.·
Eunctigera also share natural enemies, with noevidence that any of the pr~dator species prefer one over the other.
\.
10.
found in natural populations. The intensity of pa~asitism and of viral
and bacteria .. l infection varies geographically but generally increases during the season. The overall impa,· t on Heliothis populations generally appears to be low (Twine 1974, K.G. Wardhaugh pers.comm.), although
Cullen (1969) reporteci that viral infection was a major mortality agent among
.!!.·
punctigera populations in South Australia, especially late inthe season. During the 1975-76 season in Queensland, there was an average parasitism rate of 15.7% in eggs and 4% in larvae (I.P.M.u. 1 Report 1976). No data is available with regard to the intensity of predation in natural populations but it is also believed to be low
(LP.M.U. Report 1976).
Despite the apparently low impact of natural enemies on Heliothis populations, mortality from the egg to the larval stage may be as high as 98% (K.G. Wardhaugh pers.connn.). Much of this mortality occurs between the egg and small larva stages, possibly due to physical factors such as being washed off by rain (Cullen 1969). Even so, the population density of large larvae on cot ton crops in the Ord Valley, W. A. (Wilson et al. 1972) and in the Namoi Valley, N.s.w. (A.G.L. Wilson pers.comm. 1972) may reach 12-15 larvae per metre of row. At these densities, Wilson
!:!.
al. (1972~ noted heavy competition for the diminishing supply offruiting forms and frequently observed cannibalism. Thus, the broad overlaps in geographic and host range and life history characteristics,
tc.gether with the incidence of interference behaviour and the low levels of predation, suggest that interspecific competition may be a significant agent in determining the abundance
of].. armigera and
!!·
punctigera. A regulatory influence is further suggested by the regular observation of a ch:mge in the relativeabundance of each species during the growing season. Although both species are caught in light traps throughout the season,
!!·
punctigera1
--.
...
~~;...
~.. ..
0
~. 11 (
is more common during the first half while H. armig~ra is more common
1
during the second half of the season.
There are, of course, other factors which may also affect the
11.
relative abundance of each species. Insecticide pressure is an obvious possibility, since H. armigeru populations have developed resistance to DDT in cotton growing areas while
!!.·
punctigera populations remain susceptible (Wilson i974, Goodyer et al. 1975). This cannot totally explain the species change in relative abundance, however, since it also occurred in an area used by the I. P .M. U. to test an integrated pest-management study. Chlordimefor.m was the only chemical insecticide used against Heliothis in this area during the 1975-76 season; neither species was resistant to this compound (I.P.M.U. Report 1976). Another possible factor leading to the change in the relative abundance of each species is occasional long distance migrations, but no direct evidence has been reported.1.3 Outline of the approach used in the present stuc,!y
The change in the relative abundance, from
!!.·
punctigera toa
H. ~rmigera being more C"'.)mmon, occurs when both species are most,;
frequently found within cotton crops. It coincides with a change in the food types available on the developing cotton plants, from mainly squares
(buds) to maturing bolls, suggesting that the altered ~attern of resource availability may be a factor causing the change in the relative abundance of the two species of Heliothis. In order to investigate this hypothesis and to study the nature of c0mpetitive interactions between
!!.·
armigera and H. punctigera, experiments 'Were conducted at two levels o.f complexity.1
Details of the light crap catches at Narrabri, N.S.w., wer~ kindly provided for the 1972-73, 1974-75 and 1975-76 seasons by Mr. L.R. Greenup and Mr. L. Tuart of the N.S.W. Department of Agriculture. Information on the light trap catches in the {fk ~nsland cotton growing
I
12.(1) Initially the aggressive behaviour, development characteristics
and feeding behaviour of the larvae were studied, independently of each
other in order to determine:
\)
(i) the relative intensity of aggressive interference
behaviour among the larvae of H .. armigera. and
.!!·
punctigera and the outcomeof interspecific interference competition between them in a simple
laboratory system.
(ii) ~hether there are any differences in the developmental
characteristics of each species when reared on cotton plants which may
modify the outcome of interspecific aggressive encounters.
(iii) to what extent the two species utilise the same food Q
resources whe-..1. feeding on cotton plants and hen.ce are likely to compete
0 when these resources are limited.
(2) Experiments were then conducted on cotton plants to investigate
' 0
how interactions between the factors mentioned above modified the intensity
of interspecific competition. This study w-as made under glasshouse.
conditions.
-,--13.
l
C9APTER 2
GENERAL CULTURE TECHNIQUES
2.1 The animals used
Although detailed studies of the behaviour an0 development of animal populations are most conveniently conducted in controlled
laboratory envi·ronments, the possible existence of differences between laboratory and field populations limits the validity of extrapolating from laboratory data to the field situation. The differences
cncountereu in laboratory cultures include reduced mating frequency, especially during the initial stages of colony establishment (Baumhover et al. 1966, Gahan 1966, Callahan 1962), changes j.n size and development period (Evens and Niemegiers 1954 cited in Lumsden and Saunders 1966, Guthrie et al. 1965 - cited in Raun 1966, Twine 1974) and behavioural changes (Shorey and Hale 1965, Bush et al. 1976). These altered
characteristics may be partly attri0uted to physiological responses to the environmental ~egime, for example the laboratory diet (Vanderzant 1969). Genetic changes may also occur, especially if the laboratory population size becomes very small at some stage (De Meillon and Thomas 1966, Gahan 1966, Bush et al. 1976). In order to ensure that laboratory observations are appl:!.cable to natural populations, therefore, studies should ideally be limited to animals which have as brief a laboratory history as
possible.
\
col] ected fr1dividuals because of the inci.dence of parasitism and disease However, it is often impracticable to conduct experiments on field or because a large number of animals of a standard age and nutritional status may be required. Even studies on the first couple of laboratory generations are often impossible because major difficulties are often involved in establishing a viable laboratory culture. This is true for)
----... .,
14.
establishing and maintaining stocks, although they have been reared in the laboratory for many years (Callahan 1962, Cullen 1969, Raulston 1972, Twine 1974). The main problems include disease and Jnreliable mating and oviposition.
In the present study, the frequent incidence of bacterial and granulosis virus infections (identified by Dr. A. Gibbs) among both field collected larvae and the first laboratory generation derived from field collected pupae often caused the ·abandonment of experiments on these groups. Furthermore, the markedly reduced fertility of th~ adults· d.uring t'ne first and second lab.oratory generations often resulted in larval populations which were too small for experimentation. Consequently, it was not possible to limit the study to animals which were recently derived
from the field. Finally, it became necessary to use populations of each species which were derived from widely separated iocalities in studies of interspecific interactions, since because uf low fertility, only.four of the twenty-four stocks initially established persisted for more. than two generations (Appendtx 1). The extent of interpopulation diff~rences is not kno•m, although unconfirmed indications of long distance movement by Heliothis moths (French and Hurst 1969, Sparks 1972) should minimise these. The only known study of geographic variation in Heliothis indicated that
i t was insignificant in
.!i·
~ with respect to electrophoretic variants of an esterase enzyme (Sell ~ al. 19711), although the ecological relevance of this is unclear. In any case, while interpretation of the. results may be complicated by the different sources of the two species and their laboratory history, it was thought: better to use these large, reliable and healthy stocks rather than risk the repeated introduction of disease and'
f) Q t
15.
had been reared routinely or had been subjected to different experimental treatments.
2.2 Methods for culturing laboratory stocks of Heliothil:j_
2:2.1 The environment
Heliothis species require long photoperiods and warm, humid conditions for optimal survival, development and reproduction in the laboratory (Callahan 1962, Reach and Adkis.son 1970). Although responses may vary between species, the moths apparently require light intensities of less than 10 lux, temperatures below 30°C and relative humidities greater than 65% (although this varies with temperature) in order to mate
(Callahan 1962, Cullen 1969). For continuous non-diapause development, minimum day lengths of 11 hours for the moths and 13 hours for the larvae are necessary (Cullen 1969, Roach and Adkisson 1970). Twine (1974) found
that the survival of H. armigera larvae was greatest when they were reared at temperatures between 24-27°C. In the present study, howev~r, the larvae developed too sl~wly to 'work' the artificial diet satisfactorily below 25°C. This led to a marked increase in the incidence of microbial and fungal
infections.
Because only one room was available for the maintenance of laboratory stocks and the performance of laboratory experiments, it was necessary to compromise between the environmental optima of different developmental stages. Satisfactory results were obtained when all stages of both
Ji·
armigera andJi·
punctigera were reared under constant conditions in a windowless room which was maintained at a temperature of 27°±
l.5°C and a relative humidity c;f 65+
5%. Artificial illumination was provided between 0830-1700 hrs each day by eight overhead 40 watt 'natural' type fluorescent lights, at a vertical distance of 1.5 metres above the0
16.
the front of the bceeding cages. These liahts remained on between 0600-2000 hrs each day, giving a total photoperiod of 14 hours and a simulated
'dawn' and 'dusk' between 0600-0830 hrs and 1700-2000 hrs respectively. The intensity of light inside the breeding cages varied from 100 lux during
'dawn' and 'dusk' to 900 lux during full illumination.
2.2.2 Rearing procedure
a) The pupae
Heliothis samples were usually collected from the field as pupae. The species were separated then placed in 2 litre capacity, opaque plastic containers which contained a 4 cm layer of vermiculite until the adults emerged. An observation window was cut into the lid of the container and covered with organdy cl0th.
b) Adults
Newly emerged moths were collected daily and placed in breeding cages made of clear perspex and meas~ring 30 cm square by 60 cm in length
(Fig. 2.1). A 30 cm square piece of cheesecloth was placed over the open front end of the cage to provide an egg laying surface. Tr.is end was oriented towards th~ light source. Two cctton wool wads soaked in a 10% honey solution were placed in each cage for the moths to feed on. The floor of the cage was perforated and placed over a metal tray which
contained a saturated solution of sodium chloride to ensure a high humidity (Solomon 1951). The ~umidity inside the cage was increased at dusk each day by ~praying a fine mist of water into it.
c) Eggs
Because Heliothis moths prefer to ovlposit on rough textured surfaces (Cullen 1969) and are attracted to light, most eggs were laid on the cheesecloth. The eggs were harvested every two days and the cloth containing eggs was transferred to a plastic container until hatching was complete.
0
0
()
0
[image:28.618.60.578.16.756.2]17.
d) Larvae
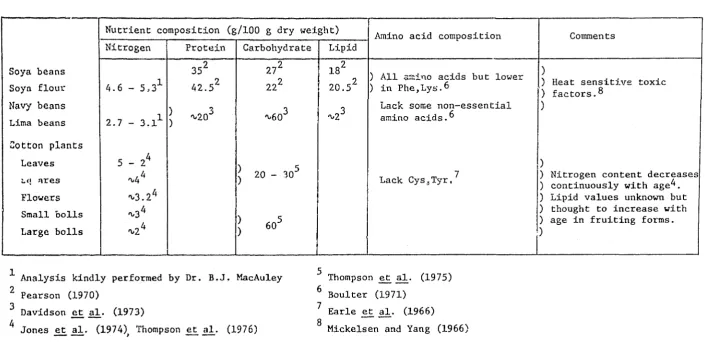
The larvae were reared on an artificial bean-based diet (Appendix 2) similar to that developed by Shorey and Hale (1965). Dried navy beans, lima beans, soya. beans and soya bDan flour were each tried in tl~.a diet and the larvae survived and developed satisfactorily on all of them. Navy beans were used routinely because they were readily available, inexpenslve and had been used successfully by other workers (Reid pers.comm., Greenup pe:rs. comm.).
Newly hatched larvae were transferred from the egg-laying cloth to a plastic box containing several strips of fresh artificial diet using a fine sable brush. The larvae were reared together for seven days, after which they were separated into individual 30 ml disposable plastic cups which were half filled ~ith fresh food. The containers were inspected daily and those with dead larvae were discarded in an attempt to reduce the
possibility vf disease spreading through a culture. The larvae pupated in
the pla~tic cups and were then transferred to pupal containers.
e) _Sanitation
The larval rearing cups were discarded after use. All other
containers and equipment were washed first with detergent, then soaked for 48 hrs i~ a 1% sodium hypochlorite solution to kill bacterial' spores.and virus particles (Ignoffo and Dutky 1963) and finally rinsed in water. The breeding cages were washed regularly ~ith deter~ent.
2. 3 Identification of lt. ar.m.;lzer_a and .H .• ~tigera
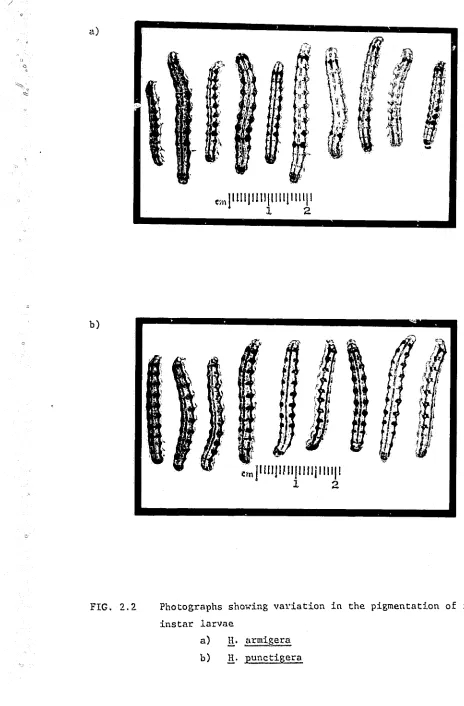
In studies of lnterspecific interactions, :.l.t is convenient to be. able to discriminate bet'We.en the species on obvious morphological
characters in order to minimise disturbance to the animals. ~his was previously thought to be impossible. for the larvae of
!!:
~gera andi!.
punctigera (Kirkpatrick 1961.'l) but reliable pigmentation differenceso'
18. ' '
at the first abdominal segment which is bounded by setae Dl, D2 and SDl (nomenclature after Hinton 1946, from Kirkpatrick 196~ is darkly pigmented.,,,. in .!!_. armigera whereas this area lacks pigmentation in
.!!.·
punctigera(frontispiece A). This character varies somewhat between individuals and instars (Figs. 2.2, 2.3), but the species can be easily distinguished during the third, fourth and fifth instars at least. The darkly pigmented patches sometimes fade during the sixth instar as the larva nears pupation and different coloured
fluori..~scent
powders (Day-Glo@) were used to help identification during this stage. The spi~acles and legs of .!:!_. armigera were also usually more darkly pigmented than those of H. punctigera but these characters were less reliable and more difficult to observe than the dorsal pigmented patches.This method for disciminating between the larvae of each species was originally developed for use in the laboratory when they were reared on artificial diets. However, it also held true for laxvae reared on cotton inside the glasshouse although the tlark pigmentation was more likely to fade during the sixth instar. A field test at Narrabri indicated that field
collected larvae were generally darker than those reared in the laboratory or glasshouse, but the species could still be distinguished in material collected from cotton, maize, linseed and sunflower crops. The technique was not reliable for larvae collected f~om lucerne, which were darkly coloured overall.
The pupae and adults of each species can be readily :i.dentified by the arrangement of the cremaster spines in the case of the pupae
(Kirkpatrick 1961) and by markings on the hind wings of the adult moths (Common 1953).
2.4
The culture of cotton plants2.4.1 The Environment
1)
;
\'
""'
a)
b)
FIG. 2.2
em
J
lll l} ll II! l Ill }11111 l
1
2
ein
JI I I I
J111Jf1111
i
I 111
j
I
l
2
Photographs showing vaYiation in the pigmentation of fifth instar larvae
a) b)
.!!:
armigera1!.·
punctigera [image:31.618.64.528.63.769.2]/
j
,,,j~~\--,\
/~:'
FIG. 2.3
~~~~~~~---..-.
rJ ~
Photographs comparing the pigmentation patterns in second and third instar larvae.
a) H. armigera II
b) H. armigera III c) H. J2UnctiBera II
[image:32.642.61.602.54.802.2]0 Q ·----,....-'·----=~
-
\~
-.->
-
•
..
...
...
..,
-
t-1
_,
-...
0--....
....
•
E
U,
_,
:.., •oN
. - J
l!ll!)
..
m
illlD
i5IQ
BlillE)
&Ill)
.., or-ti
~ Ell!!!r)..,
ill9ii)e
~.
=·N
9illD ~)
...,,
i.m ~m
..,
il!limD 8lmit) ililiillO•
!fia9
'Di-I
!IED..,
lililO ¥9me
lilmt)....e
...
u Ii!
E
~.
-.. ·N
--..->
-
m
~ imO...,
_,
IE
•Pl
11111119 !mm)
..,
~ D ~ li'8e-
....
':Ji a 118E
,-1··
,,
0 '
[
19.
varied to some extent according to the prevailing ambient conditions. The temperature varied in a regular diurnal cycle within limits set by the thermostatically controll~~ heating and evaporative cooling system. During winter the temperature cycle fluctuated between 16-27°C (occasional extremes of 14 °C, 31°C) while the S!•.mmer temperatures varied between 20-35°C
(occasional extremes of l7°C, 44°C). The relative humidity varied between 65-80% throughout the year. The photoperiod varied with ambient conditions but was increased during winter to a minimum of 13.5 hrs by four 60 watt incandescent lights which remained on between 1600-2030 hrs each day.
2.4.2 The cotton plants
The Namoi Seed Distributors provided 50 kgm of Gossypium hirsutum L. (Malvaceae), variety Deltapine 16, seed for use in this study. The seeds were germinated in 'jiffy' pots and transplanted to 4 litre pots at the
two-leaf stage. Fertili£er was applied at the time of transplantation in the form of two 6 gm Agriform@ tablets per plant. These contained nitrogen, phosphorous and potassium in the weight ratios of 14:1:8.5 plus
trace amounts of calcium, sulphur and iron.
A detailed ciP..script::t:;:~ of the morphogenesis of cotton plants is provided by Tharp (1960) and will be discussed in Appendix 5. Under the prevailing glasshouse conditions employed in this study it grew with a single main stem to a height of approximately 120 cm with 15-20 side branches. These branches have a leaf at the junction with the main stem
and ~ ~.e ar:ranged in a spiral pattern about the main stem (frontispiece B). The
branches at the base of the main stem are usually vegetative (monopodia) and have a similar gro~th pattern to the mainstem. The number of
vegetative branches varies with plant density (Hearn 1969, Gutierrez ~ al. 1975) b\lt ranged between 2-6 in the glasshouse. Fruiting branches
(sympodia) have a leaf and a fruiting form at each node. The initial
\)
0 '
20.
opens into a flower and then develops into a boll. The developing fruit may be shed during the square and small boll (
<
2 cm diameter) stages. The fruiting forms are distributed in a decreasing series with respect to age, both along the fr:iiting branches and for equivalent positions on each branch up the mainstem .2.4.3 Pest control
The tTNO spotted spider mite (Tetranichus urticae Koch), whitefly
(Trialeurodes vaporariorum Westwood) and aphids were frequent pests in the glasshouses. The spider mite was controlled by the introduction of a
predatory mite, Typhlodromus occidentalis Nesbitt and whitefly was controlled by a hymenopterous parasite, Encarsia formosa Gahan. These pests occasionally escaped biological control and Propargite (Omite<!>), a specific miticide, or Dichlorvos (Shclltox pest strip) were then used. A specific aphicide drench, Pirimicarb (Pirimor@) , was used to control aphids. Although specific insecticides were used wherever possible, plants which had been tre21ted with chemical insecticides were not used in
experiments until after the suggested toxicity period had expired and then only when tests had shown that the plants were not toxic to Heliothis larvae.
Occasional outbreaks of a bacterial infection on the plants were suppressed by spraying them with a mixture of 1 gm each of penicillin G
.;;10 '"d
'
0
di
3.1
CHAPTER 3
A_LABORATORY STUDY OF INTERFERENCE BEHAVIOUR
IN ll.· ARMIGERA AND li· PUNCTIGERA.
Introduction
21.
Fox (197 5 c) has documented evidence for the widespread occurrence of cannibalism among natural populations in a wide variety of animal taxa. Of particular interest is the evidence of a high incidence of cannibalirrm among those terrestrial insects which are generally regarded as herbivores. The larvae of many lepidopteran species are cannibalistic (Dethier 1939, Brower 1961, Gefr 1953, Abdel-Salam and El-Lakwah 1973), including all Heliothis species which have been studied [Quaintance and Brues 1905,
(!!_.~Boddie), Brazzel et al. 1953 (!!_. virescens Fabricius), Cullen 1969 (!!_. punctigera), Twine 1971 (!!_. armigera)]. Although it has been observed in all larval stages of Heliothis, the frequency of cannibalistic attacks tends to increase with age (Brazzel ~t al. 1953) and density
(Twine 1971). Such behavio1ur has led these authors to speculate that cannibalism among Heliothis larvae may be important in population self-regulation. Simulation models developed by Stinner et al. (1977) support the regulatory role of cantt.ibalism and indicate that, as well as affecting survival, cannibalism also appears to reduce the mean generation time by up to 15%. This reduction was obtained because the authors assumed that the probability of larv~e eating smaller and hence slower developing individuals upon encounter was greater than for the other possible interactions. This assumption is justified (Quaintance and Brues 1905), but it is also possible that the development rate of cannibalistic larvae may b1a enhanced, as in Triboliurn larvae (Mertz and Robertson 1970).
Cannibalism may be regarded as a form of interference or contest competition which acts to regulate the population size in either of
0
lb
/}
Q
22.
below the carrying capacity of the environment, thus ensuring that food does not become a limiting factor. Examples of this form of cannibalism include the queen and monarch butterflies (Danaus) (Brower 1961), Dineutes assimilis (whirligig beetle) (Is tock 1966), Triboli~ (Park ~~ al. 1965)
and Poecilid fish (Thibault 1974). Alternatively, cannabili$m may be common only when the population density has exceeded the carrying capacity of its environment and food is limiting, when it will serve to ensure that at least some individ•~als obtain sufficient food to reach a minimum size necessary for reproduction. Examples include Spodoptera littoralis
(cotton leaf worm) (Abdel Salam and El-Lakwar 1974), Chrysomyia albk~~ (sheep blowfly) (Ullyet 1950), Dineutes horni and .Q_. nigrior (Is tock 1966).
The experiments described in t:,is chapter concern an experimental study of :i.nterference behaviour, ~specially cannibalism, it1
.!:!.·
armigeraand
.!:!.·
punctigera larvae. The experiments were performed in the laboratoryunder conditions of both intraspecific and interspecific competition. Because food limitation would necessarily be confuundctl with space limitation due to physical crowding, space w~s considered the limiting resource in all experjments, with food provided in excess. Support for this approach is provided by evidence that the frequency of cannibalism in Tribal~ species is more dependent on space and hence encounter frequency, than on the quantity of food available (MacDonald 1968).
The investigation had two major objectives:
A) To understand and compare the nature of intraspecific interference behaviour in
.!:!.·
!!.E.!!1igera and.!:!.·
punctigera. This included studies of the effects of larval density and food quality on the intensity of interference behaviour.~ as indicated by the survival, dispersion pattern and frequency of movement of the larv.'.le.~~---~
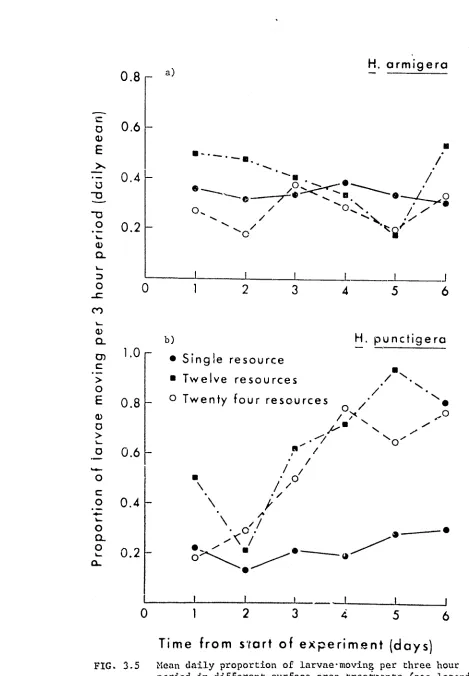
[image:38.640.42.612.76.803.2]a) Continuous resource (standard treatment)
b) Twelve discrete resources
c) Twenty-four discrete resources
4 ... ~.7
.. .,
"
0
.,
.
..
\•. "" i
0
'\
\
23.
3.2 Methods
3.2.1 Experimental Design
All experiments were conducted in the rearing room under the same environmental conditions described in Ch,2.2.1,
The st·andard experimental system employed, with which all other treatments were compared, consisted of a 30 cm square x 5 cm deep
aluminium tray, filled to a depth of 1 cm with soya bean flour-based artificial diet (900 mls) 1 (Appendix 2), A uniform, continum.J.s, two
dimensional feeding surface of 900 cm2 was thus provided, onto which twenty larvae, each weighing between 30-45 mg (late third instar), were introduced. A clear perspex lid, marked with a 5 cm square grid, was then taped in place over each tray (Fig. 3.la). The trays were placed in the centre of the rearing room at the same level as the wall light (Ch.2.2), such that each tray was evenly illuminated. Each tray was turned through an angle of 90° at every observation to further reduce the effects of any photctactic response to the wall lights.
The number of larvae surviving and the position of each larva with respect to the grid were recorded at each observation, usually made at three hourly intervals between 0900-2100 hours each day. The trials lasted for six days, after which time the larvae usually beg~n pupating. Deaths due to disease were easily distinguishable from those due to cannibali5m and trials were discarded if evidence of disease induced deaths were found.
Because of the limited availability of space and of larvae, replicate trials of each treatment usually had to be cotlducted at
different times. Since all treatments were compared with the 20 larvae per container> continuous resource, .soya flour diet treatment, a.t least
l
\)
0
D
24.
one trial of
tbis
type was attempted in conjunction with trials of other treatments. All trials of a given treatment were then considered together for analysis, irrespective of their field source and laboratory history(Appendix Table 3.1). Ineq<'alities in the degree of replication between treatments resulted from differences in the availability of larvae and from the rejection of those trials which showed evidence of disease.
3.2.2 The Analysis and Treatment of Data
a) Survival
To obtain a measure of the variation in the intensity of cannibalism, both between treatments and ~ith larval age, the mean
percentage of larvae survivir.g at twelve hourly intervals (as a fraction of the initial population) was plotted on a logarithmic scale against elapsed time. Ninety-five percent confidence intervals were calculated for each data poiut fr.om an analysis of the angularly transformed data. A single classification analysis of variance was used for statistical comparisons between treatments; ~ost of these are detailed in Appendix 3. The raw data for the number of Heliothis larvae surviving in each trial over time is provided in Appendix Tables 3.2 - 3.7.
b) Dispersion
The number of cells in the grid which contained 0,1,2, .... n larvae were recorded for each observation. The extent to which larvae were distributed randomly (i.e. independently) about their container was
then calculated by comparing the observed frequency distribution with that predicted by a Poisson model using a chi-square test. The inde~ of
dispersion, which measures the direction of the deviation from randomness, was then obtained by comparing the mean and variance of the observed distribution (Southwood 1966, Poole 1974).
I
0
0
25.
The analysis was first performed using the 5 cm grid as the sampling unit. Those trials which showed a regular dispersion pattern were re-analysed using a 10 cm square sampling grid, in order to estimate the distance over which interference behaviour betw1~en larvae influenced the dispersion pattern.
A contingency chi-square test demonstrated that the disper8ion pattern of larvae was independent of the time elapsed in all trials. The observed frequen~y of occurrence of each class was therefore added over all observations in each trial, in order to provide an adequate sample size for the test of departure from a random dispersion pattern.
c) Frequency of Movement
The net movement during a three hour period was calculated hy comparing the distribution of larvae at a given time with the distribution in the previous observation. A larva was regarded as having moved if its position on the grid was not represented in the previou3 observation. On.ly movements acrOES grid boundaries were considered. The frequency of larval movement was expressed as the proportion of surviving larvae which moved in a three hour period. Although this method could not detect reciprocal movements by two larvae to each others position, this was considered unimportant since relative changes in movement frequency, over time and between treatments, were of central interest.
The raw data for each trial are provided in Appendix Tables 3.8 -3.10. The movement frequencies were normalised using an angular
transformation before statistical comparison by an analysis of variance. Most of these ~re detailed in Appendix 3.
Section A: Intraspecific Interference Behaviour
3. 3 Comparis·on between 11. armigera and Ji. punctigera. in the standard system
---26.
pt:'r containet· on a soya bean flour artificial diet, arranged as a
~ontinuous uniform resource.
3.3.1 Results
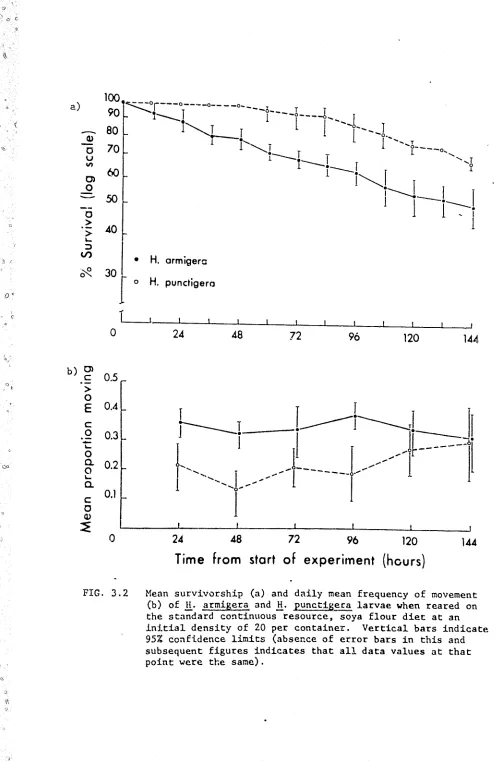
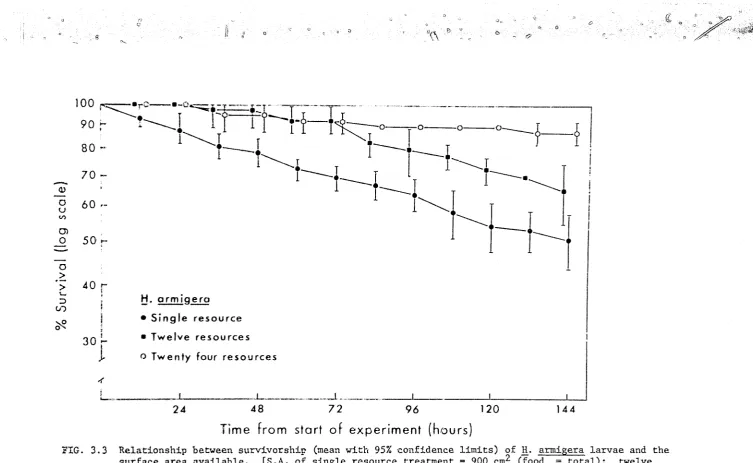
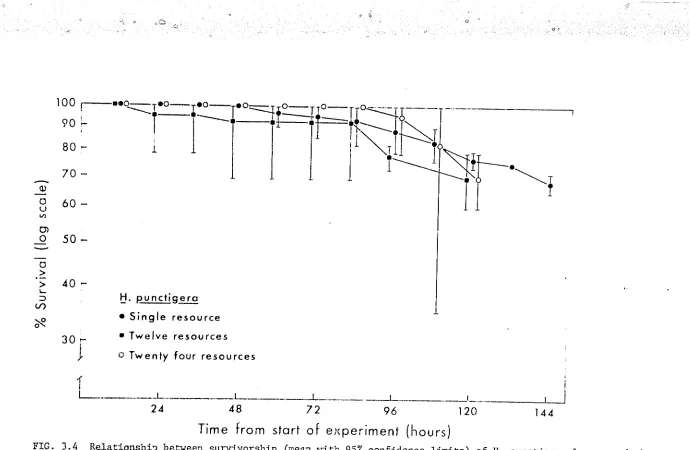
a) Survivm:-ship. (Appendix Tables 3.2, 3.4)
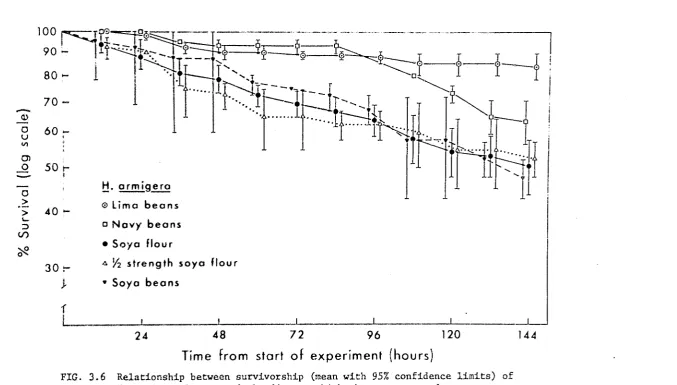
Fig. 3 • 2a indicates that _!i. armigera and H. E1.mctigera differed with respe~t to both overall survivorship and to the pattern of mortality
in these experiments. The mean survivorship of B_. armigera was l()wer than
that of!!· punctigera throughout the experimental period with 10.l
±
0.72 H. an11igera surviving to the end of the trial compared with 13. 7+
0. 33H. uuncti.gera (t(g) 2.89, .Ol<p<.02), The mortality rate for H. armigera rem2ined relatively constant through time, with the victims of all
ca.1rnibalistic encounters being eaten. In contrast, there were no daaths
d<1e to interference benaviour in the
.!!.·
punctigera trials during the first48 hours, after which the freql1ency of cannibalism increased until, between
% ·144 hours, the mortality rate was similar to that observed in
H. armigera. Furthermore most of the B_. punctigera victims of the last '
16 hours (4/5) were not eaten,
o) Dispersion
H. arnige.ra (Ta'!:>ie 3.la): A significant deviation from randomness
towards a regular dispersion pattE:·rn was observed in si.x out of the eight
r~plicates, while the:: other two trials showed a random pattern with
respect to the 5 cm grid. A re-analysis of those regularly dispersed trials using the 10 cm grid indicat~d that only one trial showed a
regular dispersion patte~n on this scale. Another trial showed an
agg<egated patt~rnt ~hile the larvae in the rem3inlng trials were
randomly distributed over the 10 cm grid. ThesP. r1~aults indicate that
interference behaviour tends to maintain a reJgulat' spacing between larvae
over sho::-t dtstanccs (up ::o S cm), with the c.ffect rHminishing over laq;e.r
t~""--..Y \ ., ~
">
o<
ao
'.i
:
a)
--
cu0
u
II)
C>
0
0
>
>
..._
:> C.I)
•
H.
ormigerc~ 30
H.
0 punctigera
l_J
0
24
48 7296
120 144b) C> c 0.5
>
0
E
c
0
·-
-
..._ 0CL
0 ..._
CL
c 0
cu
~
0.4
0.3
0.2
0.1
0
u
a
n
%
1w
Time from start of experiment (hours)
144
[image:44.613.50.548.23.784.2]TABLE 3.1 Dispersion characterstics of
.!:!.·
~igcra larvae on different diet treatments,Explanatory notes for Tables 3,1, 3.2
l, The probability that the observed dispersion pattern differs significantly from a random pattern w.:i.s obtained using chi-square tables for n-2 degrees of freedom, where n is the number of classes.
2. Index of dispersion ~ variance/mean
3, The dispersion pattern which tested for de.viation from random usirtg a 10 cm square grid was performed only on those replicates of the single continuous resource treatments which showed a regular pattern on the 5 cm square grid.
(\
0
,,,, u
1')
m
0
'ti 0
rb. of observations of x larvae
per coll l\"lplicate
0 l 2 ,3 >4
a)Soya flour diet : iunglc resource
July •7.;• obs 469 217 Jl 3
exp -184.6 191.8 38,0 5.6
Jutr '7.f obs 451 203 30
exp 465.7 179.0 39.l
~~uch '75 obs 643 298 26 3
exp 671.8 2-18.l 45.8 6.2
~1rch '75 obs 652 251 30 3
exp 667.1 225.9 38.3 ti. 7
:-'.lt'Ch •7c; cbs 525 235 27 5
exp 539,5 207.l 39.8 5.6
M1y '75 obs 468 236 48 4
exp 479 6 218,2 49.6 8.5
1-t1y '75 obs 500 204 47 5
exp 499.7 206.9 42.8 6.6
Au<] '76 obs 346 200 29 1
exp 366.l 165.9 37.6 6.4
b)Sciya flour diet. : t\>.'ClVI? resources
July '74 obs 29 109 SB l'J 13
exp 52.7 77.2 56.5 27.6 14.0
July '74 t;bs l(l 78 60 10 2
exp 41.2 57.9 40.7 19.0 9.1
c)SOya flour Giet : ~ty four resources
Jnly '74 cbs 181 232 52 2
exp 225.8 170.3 64.2 19.8
July '74 dlS 19!1 237 59 • 9
exp ..!36.2 li9.0 67.8 21.0
;;J j
""
I:1..-v1ation of dispersion pattern fron randoi
Index of
·.2 p
dis!X)rsion
6.25 * o.0a Regular
5.82
.
0.84 Regul.:ir21.5~
...
0.83 Regular5.53
.
0.91 Regular8.31
.
0.89 Rc..]ular4.15 n.s. 0.89 Randon
0.82 n.s. 0.98 Randon
14.62
...
0.80 Regular26.55
...
0.70 Regular39.08
...
0.46 Regular46.lJ
...
0.62 RegularJ:l.60
...
0.69 Regular;.;
ro
0.
0
0 O.:;
Deviation of dispersion pattern fron randon 110 an grid!
w.i. of Index of
classes x' p dispersion
6 2.30 n.s.
5 5.34 n.s.
• 6 9.65
.
1.19 J\ggreg.6 4.24 n.s.
6 1.51 n.s.
5 9.07 * 0.66 Regular
cont. over page