JOURNAL OF VIROLOGY, Mar. 1977,p.1042-1055 Copyright©D 1977 AmericanSociety for Microbiology
Vol.21,No. 3 Printed inU.S.A.
Distinguishable
Transformation-Defective Phenotypes
Among
Temperature-Sensitive Mutants of Rous Sarcoma
Virus
DOROTHEA BECKER,* REINHARD KURTH,' DAVID CRITCHLEY,2 ROBERTFRIIS, AND HEINZ
BAUER
InstitutfarVirologie,FachbereichHumanmedizin, 6300 Giessen,FederalRepublicof Germany,*and
Imperial
Cancer Research FundLaboratories,
Lincoln's InnFields,London WC 2A3PX,EnglandReceived for publication28October 1976
Eight transformation-defective, temperature-sensitive (ts) mutants of the
Prague strain of Rous sarcoma virus, subgroup A, have been isolated after
mutagenesis with 5-bromodeoxyuridine followed by selection on the basis of
focus tests. Five of these mutants, ts GI201, GI202, G1203, G1204, and G1205,
exhibit properties like most previously reported isolates in that they show a
temperature-sensitive response to each of a variety of transformation-specific
parameters tested. Interestingly, GI201, in addition to the
temperature-sensi-tive defect, carries a lesion that was observed as a nonconditional loss of
expression ofplasminogen activator protease. Three mutants, ts GI251, GI252,
and GI253, have been designated partial transformation-defective (PTD)
mu-tants sincethey behave as ts mutants accordingto sometestsfortransformation
andaswild typeaccordingtoothers. These threemutantsfail to form foci at the
nonpermissivetemperature (4100) andarenontumorigenicin3-week-old
chick-ens (body temperature, 420C). The agglutinability by concanavalin A ofcells
infected with thesemutants shows a definite temperature sensitivity, as do the
rate of 2-deoxyglucose uptake and the disappearance of the 250,000-dalton
normal cell glycoprotein (large, external, transformation sensitive [LETS]).
Although the PTD mutant-infected cells, unlike cells infected with other
trans-formationmutants, exhibit a cell-bound plasminogen activator protease atthe
nonpermissivetemperature, this activator is not detectable as afree proteasein
the medium, as it is with wild-type, virus-infected cells. The PTD mutants
behavelike the wild-typeparent intheir abilityto inducetransformed growth
properties in the infected cells, i.e., growth beyond normal cell saturation
density with or without serum-supplemented medium and growth leading to
colony formationinsoft-agar- or methyl cellulose-containing suspensionmedia.
A great variety ofchanges in cell behavior,
form, and metabolic activity are characteristic
of fibroblasts that have undergone
virus-in-duced transformation. Transformed cells fail to
respondto conditions thatinhibitthe growth of
normal cells, such as high population density (39),absence ofserumfrom themedium (13), or
absence ofa substrate for anchorage (23, 37).
Formation of characteristic foci makes
trans-formed cells apparentinamonolayer of normal
cells (38). Striking changes in the rate of trans-portofcertainsugars,which greatly exceed the range of values obtainedwith logarithmically growing or stationary normal fibroblasts (11,
IPresent address: Friedrich Miescher Laboratorium der MaxPlanck-Gesellschaft,74Thbingen,Federal Republic of Germany.
2Present address: Department of Biochemistry, Univer-sity of Leicester, Leicester LE1 7RH, England.
43), have been observed in transformed cells.
The increased mobility or altered distribution
of appropriatereceptorsresultsinsignificantly
enhanced agglutinability of transformed cells
by concanavalin A and other lectins (5, 32).
Increasedexpression ofaproteasemeasured as
aplasminogen activator (27, 30, 40, 41)occurs
ascells becometransformed. Itisprobable that
some proteolytic activity, in turn, influences
the structure of the cell surface by degrading
cellsurface proteins, among them the
250,000-dalton large, external,
transformation-sensi-tive (LETS) glycoprotein (12, 14, 34, 37, 45).
Various temperature-sensitive (ts) mutants of avian sarcoma viruses have been studied
with respect to certain ofthese parameters of
transformation (2, 4, 6, 9, 20, 22, 24, 47). In
general, mutants isolated by selection, based
on a focustest, have been temperature
sensi-1042
on November 10, 2019 by guest
http://jvi.asm.org/
DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS 1043
tive for all otherparameters tested. Two
nota-ble exceptions have beenreportedinthe
litera-ture: ts LA334 bears a transformation defect, which, although it preventsfocus formation at thenonpermissive temperature, does not inter-dict formation of colonies in soft-agar
suspen-sion (9); andts LA25, which, though defective
for focus and colony formation at the
nonper-missive temperature, does stimulate cells to
grow todensities much higher than those ob-tained by normal cells (22, 47).
This paper will document the isolation of
eight transformation-defective (TD)avian
sar-comavirus mutants, three of which exhibit an interesting dissociation of transformation pa-rameters, i.e., respond as temperature-sensi-tive mutants accordingto certain testsbut
be-have like wild-type virus according to other
tests. Hence,twophenotypically different cate-gories emerged as analyses of these mutants
werecarried out, and theprofiles of these two
categories will bedescribed.
MATERIALS AND METHODS
Cells and viruses. Chicken embryo cell cultures were prepared from fertile white leghorn embryos after 9 to10daysof incubation. Eggs for this purpose weregenerously supplied from a leukosis virus-free flock by E. Vielitz, Lohmann Tierzucht GmbH, Cux-haven, West Germany. Tissue cultures of normal and infected chicken embryo cells were grown in Dulbecco-modified Eagle medium prepared from a
powder(L and S Labor Service, Munich, West Ger-many)supplemented with antibiotics, 5% heat-inac-tivated calf serum, and 10% tryptose phosphate broth (Difco Corp., Ann Arbor, Mich.).
The parent virus used for the mutagenization and isolation of mutants was the Prague strain of Rous sarcomavirus,subgroupA.The original parent had beenrepeatedlyclonedbyisolation, with an aspirat-ing pipette,of single foci occurring at high dilution inafocus test (33), and mutants subsequently ob-tained were routinely cloned at eachstock prepara-tion step. The mutant designations (ts GI201
through205;ts GI251 through253)conformto
sug-gestionsputforthby Vogtetal. (42).
The procedures for preparation of soft-agar
sus-pensioncultures(8)andmethylcellulosesuspension
cultures (31, 46) usedin tests forcolonyformation have been describedpreviously. The compositions
andprotocolsfor the agar caseinplaqueoverlaytest
and the soft-agar casein suspension culture have been reported by Goldberg (10) and Balduzzi and Murphy (3), respectively. In allcases Dulbecco-mod-ified Eagle medium wasused withtheappropriate supplements.
Testforin vivotumorigenicity ofmutants.
Chick-ens wereinoculatedinthewing webs of bothwings
at 21days of age and examinedbiweeklythereafter for appearance oftumors atthe siteof inoculation. Based on the method of Bauer(unpublished obser-vation), it was found that both wings could beused, using differentdoses to givetwoindependent
titra-tions of the inoculated sample in a single animal. Concanavalin A agglutination test. The concana-valin A agglutination test used was a modification ofthat of Pollack and Burger (28), which has been described in detail elsewhere (21). It was found nec-essary to use plastic pipettesand test tubes for all manipulations to minimize damage to the cell plasma membranes. Briefly, subconfluent chicken embryo cells were washed twice for 5 min each with phosphate-buffered saline (PBS), pH 7.2, on a rock-ing platform. Cells were suspended by 15 min of incubation at 37°C in PBS containing 0.02% (wt/vol) EDTA and, subsequently, were washed once in warm serum-free medium followed by centrifuga-tionfor 5 min at 1,200 x g. The cell number was determined with an electronic cell counter and ad-justed to 2 x 106/ml in PBS. Lyophilized concana-valin A (Sigma, St. Louis, Mo.) wasreconstituted in distilled water, dilutedinPBS tothe desired concen-trations, and predispensed in 5-,ul quantities into wells of a Microtest plate (type I, Falcon Plastics). Each well then received 104 cells in a
5-,gl
volume; microplates were inverted to produce hanging drops and incubated for 15 min at room temperature on a slowly rocking platform. Scoring was performed us-ing alow-magnification microscope (Dynascope, Vi-sionEngineeringLtd., Send, Surrey).Determination of LETS glycoprotein. Chicken
embryo cells grown in90-mmdishes were washed three times with PBS and labeled by adding1mlof PBS containing 5 mM glucose, 0.1 U of glucose
oxidase, 20 ,Ag of lactoperoxidase, and 400 ,uCi of carrier-free sodium[251]iodide.After 10 min at room temperature, the reaction was stopped by adding8
mlof PBS containing 50 mM sodium iodide, and the cells were washed three times with thesamebuffer. Cells were scraped into PBS containing50 mM sodium iodide and2 mM phenyl sulfonyl fluoride, pelleted, and dissolvedat100°Cin 0.4mlofsample
buffer (0.1MTris, pH 6.8, 15% glycerol, 2%sodium dodecyl sulfate). Prior to electrophoresis, samples weremade 0.1 Mwith respect todithiothreitol and reheated to 100°C. The procedure has been described
indetail elsewhere (17).
Proteins were separated by using discontinuous sodiumdodecylsulfate-polyacrylamideslabgel
elec-trophoresis (17). Gelswerefixed and stained in 30%
methanol-10% acetic acid containing Coomassie
blue, dried ontopaper, andexposedtoKoderex X-ray film.
Determination of plasminogen activator. Insolu-ble ['25I]fibrinplates werepreparedasdescribedby
Unkeless et al. (41). Harvest fluidsweretaken from cellcultures that hadbeen washedoncewith
serum-freemedium,incubated for2hwith serum-free me-dium,andfinallywashed athirdtime with serum-free medium prior to incubation for12h inmedium
supplemented with 2.5% chicken serum (for cell-bound plasminogen activator determination) or in
unsupplementedmedium(for supernatant
plasmin-ogen activator determination). After collection of harvest fluids, the cell numberwasdetermined for eachplate, and the harvest fluidswerediluted with the appropriate unincubated medium so that the harvestunitcould be definedastheactivityin 1 ml
of medium incubatedover106 cellsfor12h. Harvest
VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
1044 BECKER ET AL.
fluidswerecentrifuged for15minat1,200 x gprior
to storage at -80'C until tested on insoluble
[1251]fibrin plates. The fibrinolytic reactionwas al-waysperformed at 370C, as previous experiments
(Friis, unpublished observations) had shown that
neither supernatant plasminogen activator nor
plasmin frommutantpreparations was more tem-perature sensitive than that fromwild-type
virus-infectedpreparations. The fibrinolyticreactionwas
performedwith 1 ml of harvest fluidperplate
con-taining,orsupplementedwith(accordingtothe type
of harvest fluid),2.5% chickenserum as a sourceof
plasminogen. The fibrinolytic reaction was
moni-tored over a period of 12 h, with 0.1-ml samples takenatintervals and countedinaliquid
scintilla-tioncounter. Forevaluationofplasminogen activa-toractivities, reaction timeswere chosen atwhich notmorethan 30 to50% ofthe available insoluble
[1251]fibrin (accordingto trypsindigestion controls) had beenreleased with the wild-type virus-infected preparationsormutantvirus-infectedpreparations from cellsgrownatthepermissivetemperature.
RESULTS
Isolation of mutants after
5-bromodeoxy-uridine mutagenesis. Cells infected with a
stock of cloned Prague strain Rous sarcoma
virus, subgroup A, were treated according to
the method of Bader andBrown (1) inthe dark
from the time of infectionuntil 12 h
postinfec-tion with100 jigof5-bromodeoxyuridineperml
in medium lacking the usual
tryptose-phos-phate broth supplement. At40hpostinfection,
thesupernatantswereharvested, andthe titer
of infectious virus in treated and untreated
sampleswasdeterminedinafocus test. Itwas
found that 5-bromodeoxyuridine treatment
re-ducedthe titerof treatedsamples byafactor of
about20. The 5-bromodeoxyuridine-treated
vi-rus wasplatedathigh dilutionsinafocustest,
and single foci were isolated as presumptive
mutagenized virus clones, whichweregrownto
small stocks in aMicrotestplate. Cloneswere
screenedfor theabilitytoreplicate andtoform
foci at permissive (350C) and nonpermissive
(41°C) temperatures in Microtest plate wells
usingaTiter-tecsemiautomatic dilutorsystem
(Flow Laboratories, Bonn) to facilitate
screen-ing. Out of440 clones examined, 8werefound
tobe temperaturesensitivefor focusformation.
None ofthe 440 clones exhibited temperature
sensitivity for replication ofprogeny; rather,
the titers produced at 41°C were consistently
higher byafactor of2or3than those produced
at35°C. The eighttemperature-sensitiveclones
observedwereserially clonedthreetimes,after
which their properties in various assays for
transformation were examined. Table 1shows
asummaryof results withsomeof themutants,
showing their relative abilities to form fociat
permissive and nonpermissive temperatures. Growth-related parameters of
transforma-tion. The ability of transformed cells to form
coloniesinasuspension medium of soft agar or
methyl cellulose is a measure of
anchorage-independent growth capacity, a property not
exhibited by normal fibroblast cells (23, 36).
Table 2 shows theresults of a test for the ability
ofseveral temperature-sensitivemutants to in-duceanchorage-independent growthininfected
cells at permissive and nonpermissive tempera-tures. Prague strain Rous sarcoma virus
(sub-group A) wild-type-infected cells formed
colo-nies with aboutthe same efficiency at either 35
or41'C,whereas normal cells failed todosoat
both temperatures. Significantly, the group of
ts mutants, GI251, G1252, and GI253,
desig-nated partial transformation-defective (PTD)
mutants, produced colonies like the wild-type
virus at both temperatures, in either a
soft-agar or methyl cellulose suspension culture.
The conventional TD ts mutants, GI201 and
GI202,aswell asGI203, GI204, and GI205 (data
not shown), failed to form colonies at the
non-permissive temperature.
The ability of a virus to induce
anchorage-independent growth is usually considered a
useful measure of the oncogenicity of a virus
strain (35). Therefore, several
temperature-sensitive mutantswere testeddirectly for their
ability to induce tumor formation in chickens,
which have a body temperature of 420C. The
birds were each inoculated with two dilutions,
oneineach wing web. The results of the
experi-ment aresummarized in Table 3, which shows
that although 60% of the animals formed
tu-mors with tsGI251, none formed tumors with ts
GI252, and only 10% formed tumors with ts
GI253 and with tsGI202, even though the doses
TABLE 1. Focus-formingtitersof differentmutantsa
Focus titer(FFU/ml) Focus-forming
Virus efficiency
350C 410C
(410C/350C)
WTPrA 8 x 105 8 x 105 1
ts GI201 8 x 105 <101 <10-4 tsGI202 1 X 106 1 X 102 1 X 10-4
ts GI251 2 x 106 <101 <10-5 ts GI252 7 x 105 <101 <1O-4
tsGI253 8 x 105 <101 <10-4
a Cloned virus prepared insmall stocks at
350C
wastitrated in a focus test performed in duplicate at
35 and 41'C. The number of foci appearing was
countedonday8after plating. FFU, Focus-forming units;WTPrA, wild-type Rous sarcoma virus, Pra-gue strain, subgroup A.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.267.458.500.587.2]DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS 1045
TABLE 2. Efficiency of colony formation in soft-agar and methyl cellulose suspension cultures
Fraction of inoculated cells formingcoloniesin:
Virus Soft agar Soft methyl cellulose
35°C 410C Efficiencyb 35°C 410C Efficiency
None <0.001 <0.001 <0.0001 <0.0001
WT PrAc 0.09 0.08 0.9 NDd ND
ts GI201 0.1 <0.001 <0.01 0.02 <0.0001 <0.005
ts GI202 0.3 <0.001 <0.003 0.02 <0.0001 <0.005
ts GI251 0.04 0.1 2.5 0.08 0.2 2.5
ts GI252 0.3 0.2 0.7 0.02 0.002 0.1
ts GI253 0.2 0.2 1.0 0.07 0.2 2.9
a Suspension culture mediawerepreparedaspreviously described (8, 31, 46). For the soft-agar colony test, 103cells infected at amultiplicity of infection of approximately 2 focus-forming units (FFU)/cell were plated on a35-mmplate. For the soft-methyl cellulose colony test,104cells, alsoinfected at2FFU/cell,were
usedon50-mmplates.
bThe efficiency of colony formation expresses the ratio of the number of colonies obtained at the nonpermissive temperature to that obtained at the permissive temperature. The values are underlined to
stresstheir derivative nature.
e WTPrA, Wild-type Rous sarcoma virus, Prague strain, subgroupA.
dND, Not done.
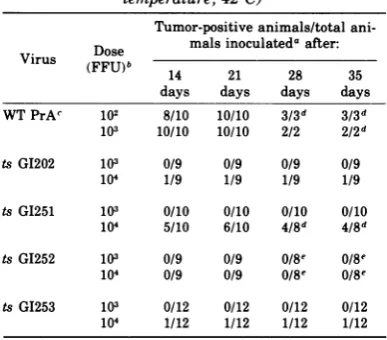
used with the mutant viruses were 100-fold
greaterthan thoseneededtoproducetumorsin
100%of the animals with the wild-type parent. Hence, asignificant degree of defectiveness for
tumorinductionwasobserved with allmutants
tested; thus the focus test, according towhich all mutants studied were temperature
sensi-tive, was abetter measure ofoncogenicity in
this case than the suspension culture test,
which had indicated that PTDmutantswere as
capable of inducing anchorage-independent
growth at the nonpermissive temperature as
the wild-type virus.
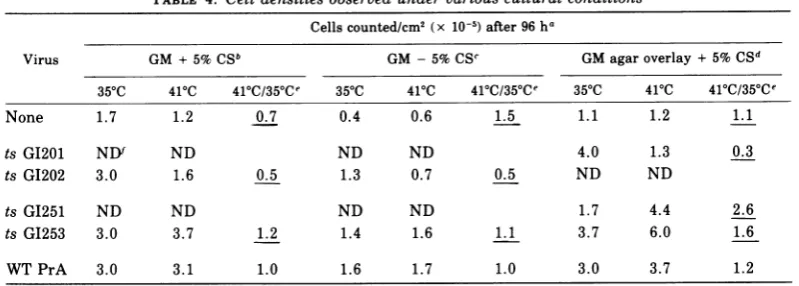
Anothermeasureof growth characteristic for
transformed cells, butnotnormal cells, is the
absence of, or increased, saturation density
(39).Table4shows that after 96 h, normal cells cultured inmedium containing5% calfserum
attain a maximum density much lower than
that ofmutant-transformed cellsatthe permis-sive temperature. Thisdensity, about 1.5 x 105 cells/cm2, remains relativelyconstant with
nor-mal cellsfor severaldaysmoreinculture,after
which the cells detachspontaneouslyas a
mon-olayer. Transformed cells, on the otherhand,
continuetoincreaseindensityuntilmedium is
exhausted or cellsspontaneously detach. It is
apparent from these results (Table 4) that
whereasts GI202-infected cellsgrowlike
trans-formed cells at 35°C and like normal cells at
41°C, ts GI253-infected cells behave like the
wild-typevirus-infected cells atboth
tempera-tures. Likewise, an experiment (Table 4)
con-TABLE 3. In vivo tumor formation in chickens (body temperature,420C)
Tumor-positive animals/total
ani-Dose malsinoculated after:
Vlrus(FFU)b 14 21 28 35
days days days days WT PrAc 102 8/10 10/10 3/3d 3/3d
103 10/10 10/10 2/2 2/2d
tsG1202 103 0/9 0/9 0/9 0/9
104 1/9 1/9 1/9 1/9
tsGI251 103 0/10 0/10 0/10 0/10 104 5/10 6/10 4/8d 4/8d
ts G1252 103 0/9 0/9 0/8e 0/8e
104 0/9 0/9 /8Me 0/8e tsG1253 103 0/12 0/12 0/12 0/12 104 1/12 1/12 1/12 1/12
a Groups of9 to 12 3-week-old white leghorn chickens fromaleukosis-free flockwereinoculatedinthewingwebs, eachwith 0.05 mlofvirusdilutedto contain theindicated dose. The right wing webs received a 10-fold-lower dose than the left, sothat the titrationcould be done at two dilutionswith aminimal number ofanimals. Independent titration of inoculainseparatewings ofasingle animal has been demonstrated (Bauer, unpublished observation). Ex-aminationsfortumorformationweremadebiweekly.
bFFU, Focus-formingunits.
cWTPrA,Wild-typeRous sarcoma virus,Prague strain,
subgroupA.
dSelected animals weresacrificed after21days because largenecrotictumorshad developed, renderingtheanimals moribund.
eOne animal died of causesunrelatedtovirus inocula-tion.
VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.253.447.336.506.2]TABLE 4. Cell densitiesobserved under variouscultural conditions
Cellscounted/cm2 (x 10-5)after 96 ha
Virus GM + 5%CSb GM -5%CSC GM agaroverlay+5%CSd
35°C 410C 41C/35aCe 355C 410C 41C/35aCe 355C 410C 415C/35Ce
None 1.7 1.2 0.7 0.4 0.6 1.5 1.1 1.2 1.1
ts G1201 ND ND ND ND 4.0 1.3 0.3
ts G1202 3.0 1.6 0.5 1.3 0.7 0.5 ND ND
ts GI251 ND ND ND ND 1.7 4.4 2.6
ts GI253 3.0 3.7 1.2 1.4 1.6 1.1 3.7 6.0 1.6
WTPrA 3.0 3.1 1.0 1.6 1.7 1.0 3.0 3.7 1.2
a Chickenembryo cellswereinfected with the various viruses at amultiplicity of infection of approxi-mately 2focus-forming units/cell, and after4days of incubation at 350C thecellswereseededintoduplicate
30-mmplates at a density of0.5 x 105cells/cm2 inmedia and attemperatures as indicated. After96 h, without medium change during the dayssinceseeding,liquid mediumculturesupernatantswerecollected
andpooled with cells suspended by trypsin treatment (0.25% trypsinfor 10 min at roomtemperature)from
the corresponding plate.Cells that had been overlayedwithagar-containingmedia werecollected after agar
had beengently aspirated using suction throughaPasteur pipette,followedby trypsintreatment tosuspend the cells. Thecell numbers weredetermined by an electronic cell counter,and the results areexpressedas
the cell number per squarecentimeterofplate surfacearea.
b Growthmedium (GM) wasDulbecco-modifiedEaglemediumcontaining10%tryptosephosphatebroth
and5%calfserum (CS).
c GMwasmade for footnoteb exceptthat CSwasabsent.
d GMwasmadeasfor footnote b except that24hafterseeding, plates received GMcontainingafinal
concentration of0.5%agar (Difco Corp.).
eThe ratio of the cell number per square centimeterobserved at 410Ctothatobserved at 35°Cispresented
tosummarize thedifferences betweenTDandPTDmutant-infected cells and betweennormal cells and
wild-type Rous sarcoma virus strain Prague,subgroupA (WTPrA)-infected cells. These derivative valuesare
underlined.
fND,Notdone.
ducted without serum in the medium showed
thatthe characteristic inhibition ofgrowth
ex-pected with normal cells (13)wasseen withts
GI202 only atthe nonpermissivetemperature,
and not at all with ts G1253. Finally, Table 4
showsanexperimentinwhichanagaroverlay,
inhibitory to normal cell growth, perhaps
ow-ing tothehigh concentration ofanionicgroups
containedinthe crudeagar, was usedto
dem-onstrate that whereas ts GI201-infected cell
growth is temperature sensitive under these
inhibitory conditions, ts GI251- and
G1253-in-fected cells exhibited no growth inhibition at the nonpermissive temperature. ts G1201, ts
GI202,ts GI203,tsG1204,andts GI205 behaved
alike in such experiments (data not shown),
whereas the PTD mutants behaved like the
wild type.
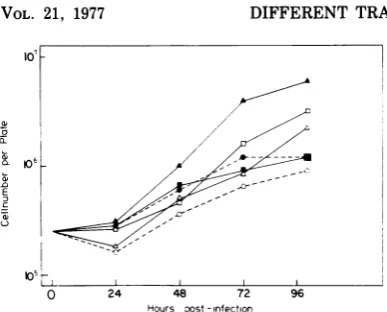
Figure 1 shows agrowth kineticexperiment
similar to that described in Table 4, which
clearlyshows thetemperatureindependence of
ts G1251-infected cellsfor growth inliquid
me-diumon asolid substrate and thetemperature
sensitivity ofts GI201-infectedcells.
ConcanavalinAagglutinability and hexose
transport as measuresof transformed cell
al-tered membrane functions. Lectin
agglutina-bility of cells infected by the variousmutants,
atpermissiveandnonpermissive temperatures,
was studied using concanavalin A. Table 5
shows thataccordingtothiscriterionof altered
membrane function in transformed cells,
pre-sumably resulting from increased mobility of
the lectin binding sites (32), the PTD and TD
mutants do not differ. A definite temperature
sensitivitywas apparentforall mutants.
Greatly increased transport of certain hex-osesandaminoacidsisratheruniquely charac-teristicoftransformedfibroblasts, sincenormal
cells, underany state of growth, never attain
transport levels equivalent to those of
trans-formed cells (11, 43). Therefore, the mutants
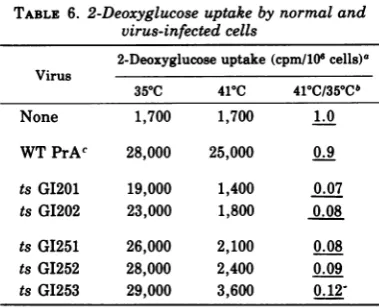
were examinedfor changes in the
2-deoxyglu-cose transport rate,which might occurafter a
shift ofinfectedcells from the nonpermissive to
the permissive temperature. Figure 2 shows
that the increase in rate of 2-deoxyglucose transport seen with ts GI201 is precisely
re-J. VIROL.
1046 BECKER ET AL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.58.458.79.222.2]DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS
05L
0 24 48 72
Hours post-infection
96
FIG. 1. Growth curves with ts GI201 (U)-and ts GI251 (A)-infectedcells, as well as with normal cells (0), is shown at 359C (open symbols) and 419C (closed symbols). Cells were infected with a multi-plicity of infection of2 focus-forming units/cell at time 0 and plated after absorption for 30 min at roomtemperature in suspension on 35-mm plates at
an initial density of 3.5 x 105 cells/plate in liquid medium. Cell numbers per plate were determined at intervals as shown.
TABLE 5. Agglutinationofmutant-infectedcells with concanavalinAa
Growth Degree of agglutination at concan-Virus temp avalin Aconcn(gg/ml)b of:
mutant tOC_
('C) 0 62.5 125 250 500
None 35 - - - - +
41 - - - - +
tsGI201 35 - ++ ++ +++ ++++
41 - - - - +
tsGI202 35 - +++ +++ ++++ ++++
41 - - - - +
tsGI251 35 - +++ ++++ ++++ ++++
41 - - -
-tsGI252 35 - +++ +++ ++++ ++++
41 - - - + +
tsG1253 35 - +++ ++++ ++++ ++++
41 - - - -
-aTheprocedureused is describedinMaterials and Meth-ods.
bAgglutinationwasscoredmicroscopicallyasfollows:-, 90 to100%; +,70 to90%; ++,50 to70%; + ++, 30to50%; + + ++,0 to 30%singlecells.
flected inasimilarincreasedrate seenwithts
GI251after shifttothe permissivetemperature.
Table 6 shows the normalized values obtained
withseveral mutant-infected celltypes,normal
cells, and wild-type virus-infected cells. A
sig-nificanttemperaturesensitivitywas seenwith
allmutantsfor thistransportfunction(datafor
ts G1203,tsG1204, andtsGI205notshown). The
values obtained for the three PTDmutants at
the nonpermissive temperature were,
regu-larly, slightly higher thanthoseseen with the TD mutants or normal cells. This is perhaps
explainable inthat PTD-infected cells are not
subject, atthe nonpermissive temperature, to
the growth inhibition that, even though the
cultures were still subconfluent, may already
have begun to retard mutant-infected or
nor-mal cell growth. That the state of cell growth
influences hexose transport rates to some
de-gree (two- to threefold) is recognized (24, 43).
The verylargedifferential between uptake by
normal phenotype and transformedphenotype
cells is probably attributable to the compound
influences of transformation and density
inhi-bition ofnormal cellgrowth.
Examination of expressionof LETSprotein
inmutant-infected cells.Transformed cells
ex-hibit reduced amounts or a total absence ofa
250,000-daltonglycoprotein,the so-called LETS
glycoprotein (12, 14, 34, 37, 45). Inthepresent
study the level of LETSglycoproteinwasfound
to be temperature sensitive in cells infected
with ts GI202, ts GI251, ts GI252, andts GI253
as detectedeitherbylactoperoxidase-catalyzed
cell surface iodination (Fig. 3) or Coomassie
blue stainingoftotalcell protein(Fig.4).
Inter-estingly, cells infectedby ts GI201 retained a
considerable amount of LETS glycoprotein
35-A
? 30
-u
25-in 0
:o
20-0o 15
-'a 5 Wa
0 2 6
Hours after shift ('2'C-351C) 18
FIG. 2. Uptake of 2-f3H]deoxyglucose bytsGI201 (i)-andtsGI251 (A)-infectedcellsatvarious inter-valsafterashift from the nonpermissive tothe
per-missivetemperatureis shown.Chicken embryocells
were infected as described in footnote a, Table 6,
transferred into 30-mmdishes, and maintained for 18 hat41'C.Atintervals,plateswereshiftedto35'C;
at36 hafter transferallplates werewashed,and
2-PH]deoxyglucose uptake was determined as
de-scribed in Table 6. The results are expressed as
countsperminuteper106rcells.
obL
CL;
aB
co
c,I~~~~~~~~~~~~~~~~~~~
VOL. 21, 1977 1047
lo'
2
C3
Cl--C,
-1D-I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.51.245.55.211.2] [image:6.501.51.242.339.529.2] [image:6.501.257.452.408.545.2]1048 BECKER ET AL.
TABLE 6. 2-Deoxyglucoseuptake bynormal and virus-infected cells
2-Deoxyglucoseuptake(cpm/101cells)a Virus
350C 410C 410C/350Cb
None 1,700 1,700 1.0
WTPrAc 28,000 25,000 0.9
tsGI201 19,000 1,400 0.07
ts G1202 23,000 1,800 0.08
tsGI251 26,000 2,100 0.08
ts GI252 28,000 2,400 0.09
ts G1253 29,000 3,600
0.12-a Chicken embryo cells were infected at a
multi-plicity of approximately 2 focus-forming units/cell
and, after3days of incubation at350C,were trans-ferred to 30-mm plates at the temperatures indi-cated, with an initial density of 0.5 x 105cells/cm2.
Two days later, when the cells were nearly confluent with approximately 1.2 x 105 cells/cm2, the plates weregently washed twice with glucose-free Hanks balanced salt solution (H minus) at410C,incubated with 0.5,uCi of2-[3Hldeoxyglucose-supplemented H minussolution for 10 min at410C,andwashed three times withice-cold H minus solution. The cells were suspended using trypsin (0.05%trypsinfor 10 min at roomtemperature), asample was counted with an electronic cell counter, and 80% of the total was lysed with 1% Triton X-100 followed by liquid scin-tillation counting in a water-compatible mixture. The results obtained are expressed as counts per minuteobserved per 106 cells present in the sample. bThe ratio of the counts per minute uptake mea-sured withcells grown at410Ctothatobserved with cells grown at 350C is presented to illustrate the differences observed with the different mutant-in-fected andwild-type virus-infected cells. The values
areunderlined to stress their derivative nature. e WTPrA, Wild-type Rous sarcoma virus, Prague strain, subgroupA.
even when grown at 35°C (50% of the level
observedat 41°C), although accordingto other
parameters, asshown above,ts GI201-infected
cellsat35°C arefully transformed. This result
wouldsuggest that ts GI201 isnonconditionally
defective foranactivity thatisresponsible for
the loss of LETS glycoprotein in transformed
cells.
Measures of plasminogen activator
pro-teasewith mutant-infected cells. The finding
that loss of LETS glycoproteinwastemperature
sensitive with all mutants tested suggested
thatexpression ofplasminogen activator pro-tease (40) or other protease activities (7, 44)
might be defective at the nonpermissive
tem-perature,sinceproteaseshavebeen proposedas
onepossible mechanism by whichLETSprotein
might be lost during transformation (15, 18). Experimentswerefirstperformed usingafocus test modified to include opaque casein in the overlay as a substrate for protease activities, especially plasmin (10). It was found that whereas TD mutant-infected cells failed to
pro-duce clear areasorplaques in the overlay at the nonpermissive temperature, the cells infected with PTDmutantsdid, though somewhat later than wild-type virus-infected cells (data not shown). At the permissive temperature, all
mu-tants tested, as well as wild-type virus, could induce plaques over cells, whichwerelater ap-parent as foci of transformed cells. Thedelayed positive plaque formation with PTD mutant-infected cells at the nonpermissive temperature suggested some protease activity, though prob-ably less than that induced by wild-type virus.
A second type of caseinolytic test was
per-formed basedonthe suspension cultureplaque
testof Balduzzi and Murphy (3). Figure5shows
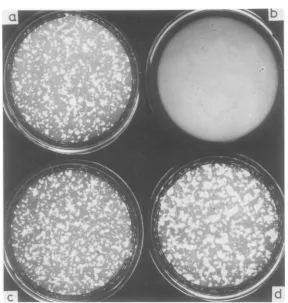
the results withtsGI202- andts GI253-infected
cells in suspension culture at 35 and 41°C. Cleared areas orplaquesare seen with tsG1202 only at the permissive temperature, whereas with ts GI253 plaques are numerous at both temperatures,being ofevenlargerdiameterat the nonpermissive temperature. These tests
had been incubated for only3days,sothe
abil-ityofts GI253toform coloniesinsuspensionat
the nonpermissive temperatureplayedno
ma-jor role. Single transformed cells are
appar-ently capableofactivatingsufficientplasminto
produceaplaque. Theplaques observedinthis
test with wild-type virus-infected cells (data
not shown) were similar to those seen with cells
infectedwith PTD mutants;hence,accordingto
thisassay, thePTD mutants, in contrast to TD mutants, were notdefective atthe nonpermis-sive temperature for a plasminogen activator protease.
Quantitative measurements of plasminogen
activator wereperformed using the fibrinolytic assay of Unkelessetal. (41). This fibrinolytic
activity has been identifiedasarisingfrom the
actionofurokinase-likeprotease (plasminogen
activator;40) onthezymogenplasminogen,
re-sulting in activation of plasmin, the protease
actually measured with the insoluble
[125I]fibrin test under the conditions used. To
meetthe requirements ofa testfor temperature
sensitivity, it was necessary toobtainharvests
of plasminogen activator from cells grown at
either 35 or41°Cand tobeabletomake compar-isons of their activities. Hence, separate
har-vestfluidsweretaken from cellsatthese
tem-peratures and reacted on cell-free plates
con-taining insoluble [125I]fibrinat asingle
temper-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.62.252.75.229.2]DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS
-OK
41
0c
Do-350
c
a
b
c
d
e
g
h
- -~~~-- wo
FIG. 3. Iodinated cell surface proteins of mutant-virus-infected cells grown at 35 and410C. The LETS band is prominent in cells infected with all mutants at 41'Cand with normal cells, but is significantly reduced with mutantsat350C and wild-type virus-infected cells. Cells were labeled using the lactoperoxidase system when nearing confluence. Gel tracks contain equal amounts of cell protein. (a-e) Cells grown at 41°C: a, ts GI201; b, ts GL202; c, tsG1251;d, tsGI252; e, ts G1253.(f-j) Cells, in the same order, grown at350C. (k) Nor-malcells. (1) Wild-type-infected cells grown at350C.The kand 1 LETS bands are displaced relative to others because these samples were subjected to electrophoresis on a separate slab gel.
-
41
0c
a
b
c
d
ON
-44
350
c
e
f
g
h
I
V"O
fflK * ~ 4_01
1. -I.
lt
:;3;
z~~lo4O
FIG. 4. Total cellular proteins of mutant-virus-infected cells grown at 35 and 41°C, as detected by
Coomassieblue stain. The tracksarelabeledasinFig.3.Tracks handjreceivedmarkerprotein of 130,000 (E. coli /3galactosidase) and 68,000 (bovine serum albumin) molecular weight, respectively. The band
correspondingtothemajoriodinatedprotein (LETS; molecular weight, 250,000) is also indicated.
*i
i
Jk
I
:.-j
4250
-v130
1049
VOL. 21, 1977
... . fi
t-I115-ft 'A"owV1.
."
la.'.W W
.wmmmw.p m.-Goo-VW-
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.101.386.73.315.2] [image:8.501.102.387.403.597.2]1050 BECKER ET AL.
[image:9.501.112.401.66.370.2]a
FIG. 5. Suspension culture system containing casein is presented to indicate the plasminogen activator expressionoftsGI202-infected cells (a and b) andtsGI253-infected cells (c and d)atpermissive (a and c) and nonpermissive (b andd) temperatures.
ature,
370C,
aspreliminarytestshad indicated that mutant-infected, cell-derived plasminogenactivator was no more temperature sensitive
than such plasminogen activator derived from
wild-typevirus-infected cells.
Based onthe experimentswith the
caseinoly-tic assay, a method was used to distinguish
betweencell-boundand supernatant activator.
This was possible because harvest fluids
ob-tained from well-washed cells, incubated in a
medium lackingserum asplasminogensource,
couldonlycontainplasminogenactivator. Such
plasminogen activatorwasmixed withchicken
serum directly on the insoluble [1251]fibrin
plate, so that the final measurement was
ac-tually plasmin-digested [1251]fibrin. On the
other hand, cell-bound plasminogen activator
could bemeasuredby obtaining harvest fluids
from cells incubatedinthe presence of chicken
serumas aplasminogen source. Of course,this
assay ofcell-bound activity must include also
the supernatant plasminogen activator, which
will have simultaneously activated
plasmino-genduring the incubation; however, the actual
problem with the mutantsstudiedhas beento
recognizecell-bound plasminogen activator
un-der conditions where supernatantplasminogen
activator is absent. Furthermore, supernatant
plasminogen activator, perhaps because its
half-life isshorter thanthat of plasminduring
the 12-h incubation on cells, always gives a
reaction of lower activity on the insoluble
[I25I]fibrin plates than is seen with the
cell-bound activatortest.
Results of a typical experiment using these
procedures to measure cell-bound (including
supernatant) plasminogen activator, compared
with those measuring supernatant
plasmino-gen activator alone, are shown in Table 7. ts
GI201 is ofparticular interest because it
ex-hibits almost no detectable supernatant
plas-minogen activator at either temperature, and
the cell-boundactivity is alsoextremelylow. ts
GI202 is simply temperaturesensitive for both
types of plasminogen activator. The PTD
mu-tants, on the other hand, although strikingly
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS 1051 TABLE 7. Temperaturedependence ofplasminogen
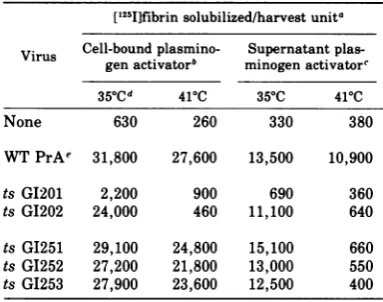
activator protease expression byvirus-infectedcells [1251]fibrin solubilized/harvest unita Virus Cell-bound
plasmino-
Supernatant
plas-gen activator" minogenactivatore
35aCd
410C 35 C 410CNone 630 260 330 380
WTPrAe 31,800 27,600 13,500 10,900
ts GI201 2,200 900 690 360
ts GI202 24,000 460 11,100 640
ts GI251 29,100 24,800 15,100 660
ts GI252 27,200 21,800 13,000 550
ts GI253 27,900 23,600 12,500 400
a Thirty-millimeter (7.06-cm2) plates prepared
with 8,000cpmof insoluble [1251]fibrin and 10 gg of
cold fibrinpercm2,accordingtothe method of
Unke-lessetal.(41),wereusedtomeasurethe proteolytic
activityoftwotypesofplasmin-containing
prepara-tions. One milliliter of harvest fluidwas incubated onthe fibrinplatesat370C. Atintervals, asample wastaken and countedin anaqueousmixture ina
liquid scintillation counter. The values shown are
for 8-h digestion of the fibrin plates corrected to
reflect the counts per minute 125I solubilized per
plateperharvestunit. One harvest unitwasdefined
as1ml ofharvest fluid obtained after12 hof incuba-tion of the appropriate medium at the indicated temperature with 106 cells. Cell counts were
per-formedonplatesfrom which harvest fluids had been
obtained, and,basedontheresults,the fluidsample
wasdiluted with unincubated mediumtonormalize for celldensity.
bFormeasurementofcell-boundplasminogen
ac-tivator, harvest fluids were prepared as described
aboveusing Dulbecco-modified Eaglemedium
sup-plemented with 2.5% chicken serum as asource of
plasminogen. Fluidswerefrozenat -80°C. e For measurement ofsupernatant plasminogen activator, harvest fluids were obtained using Dul-becco-modifiedEagle medium withoutsupplement. Fluidswerefrozenat-80°C.At the time ofassayfor
proteolytic activityon[125I]fibrin plates,chicken se-rum wasaddedtoafinal concentration of2.5% to the fluids as a source ofplasminogen. Controls done
without this source ofplasminogen showedonly a
background activity (approximately 600 cpm/plate
per harvest unit), which was subtracted individ-ually from the valuespresented.
d Chickenembryocellswereinfected ata
multi-plicity ofapproximately 2 focus-forming units/cell and,after 3daysofincubationat35°C,were trans-ferred to 30-mm plates at the temperatures indi-cated. Two days later, cells were washed and
pre-paredasdescribed abovefor harvest fluid harvests.
e WTPrA, Wild-typeRoussarcomavirus, Prague
strain, subgroupA.
temperature sensitive for expression of
plas-minogen activator in harvested supernatants,
display virtually undiminished activity ofthe
cell-bound type. Hence, the finding that foci
and colonies ofPTD mutant-infected cells
ex-hibited temperature-independent caseinolysis
maybe understood. The casein-containing
me-dia also contained serum as a matter of course. This serum made plasminogenavailableto the cell-bound plasminogen activator, and the re-sulting plasmin was responsible for caseino-lysis.
DISCUSSION
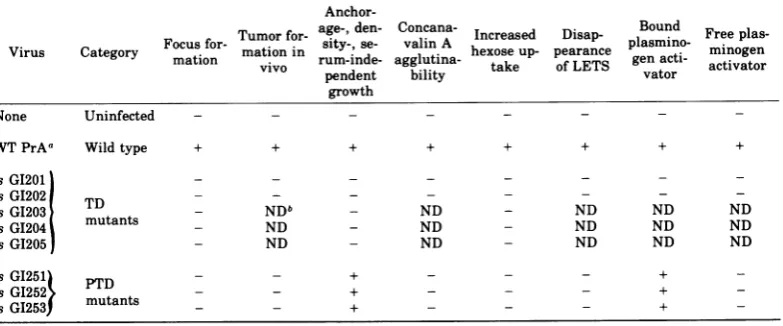
Table 8 presents a summary of the response
of cells infected with the various mutants in
separate parameters of transformed behavior.
For purposes of this discussion it is of central
importance todistinguish between the
parame-tersof cell behavior thatcanbemeasured and
the functions of the virus-coded transformation
proteinthat indirectly, after an unknown
num-ber of intervening processes, induces the
pheno-typic parameters we recognize. The TD
mu-tants, like most previously studied
tempera-ture-sensitive mutants of Rous sarcoma virus,
induce consistently temperature-sensitive
be-haviorinthe host cells. In contrast, the results
withthe PTDmutantsdelineateadissociation
betweencertain parametersof transformation.
PTD mutant-infected cells behave atthe
non-permissive temperature like wild-type
virus-infected cells according to three measures of
altered social behavior: loss ofanchorage,
se-rum, and density-dependent growth control.
Concomitant with, and perhaps causally
re-lated to, this altered growth control, the PTD
mutantsinduce intheir hostcells, atthe
non-permissive temperature, expression of
plas-minogen activator. Thisplasminogen activator
is indeed not really similar to that found with
wild-type virus-transformedcells, since it
can-notbe detected freeinthe supernatant butonly
withprocedures thatexposeplasminogentothe
cell surface. This mutant-inducedplasminogen
activator is, however, in terms of enzymatic
activity, comparable to that ofthe wild-type
virus-infected cells, and,mostsignificantly, the
cell-boundactivity found with thePTD mutant
systemislackingwith allotherTD mutants at
the nonpermissive temperature.
In reporting the existence of mutants like
those in the PTD category, the question of
whether these mutants may not be simply
"leaky" inthe conventionalgenetic sense (i.e.,
fractionally functional owing perhaps to a
merely statistical inactivation of the altered
VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.47.239.99.250.2]TABLE 8. Summary of parameters and responsesobserved with different mutants at41'C
Anchor-Turor for- age-, den- Concana- Increased Disap- Bound Free plas-Virus Category Focus for- mation in sity-,se- valinA hexose up- pearance
plasmino-
minogenmation vivo rum-inde- agglutina- take of LETS genacti- activator
pendent bility vator
growth None Uninfected
-WTPrAa Wild type + + + + + + + +
tsG1201 _
tsG1202 - -
-tsGI203 TD- NDb ND - ND ND ND
tsGI204 mutants _ ND - ND ND ND ND
tsGI205 - ND ND ND ND ND
tsGI251 PTD - - + - - +
-tsGI253 mutants - - +
aWT PrA,Wild-type Rous sarcomavirus,Prague strain,subgroupA.
bND, Notdone.
protein at the nonpermissive temperature) is immediately encountered. Admittedly, the PTD mutants might be loosely described as
leakyaccordingtosomeparametersof
transfor-mation, although according to other
parame-terstheyarenot moreleaky than manyother
conventional TD mutants. The questions of sensitivity of measurement with the various parameters and oflinearity of the assays
be-come problematic for arrivingataconclusion.
Theconceptof"leakiness"resolves into the first
oftwo hypothetical models that may be
con-structedtoexplain the existence ofPTDaswell as TDmutants. (i)Onemay assumethatas a
result of the nature of the lesion in certain
mutants, asmall fraction of the altered
trans-formationprotein retainsfunction.Then,ifone assumesthat thisfunctionalproteinacts inthe
sameway (withasingle active site)onseveral
discrete cellular sites, some of which have higherthresholds than others in ordertoexert
an effect on the total cellular phenotype, one
couldexpect toobtain results like those found with PTDmutants. TDmutantswouldsimply be mutants in which the altered protein
re-tainedno function. (ii) Avirus-coded
transfor-mation protein maynot be limited to asingle
typeoffunctioninitsinteraction with the cellu-lar machinery. Ifseveral transformation func-tions, presumably carriedoutby a single
pro-tein molecule withmultiple active sites, have independent effects on the host cell, one may assume thatundercertain conditions a
muta-tionin thisprotein would lameone functional
site without injuryto theother(s). This would also result in aphenotypic dissociation of
var-iousparameters measuring theindirect effects of the transformation functions. SoaPTD
mu-tant would be amutant inwhich aparticular
functionisselectively destroyed, andaTD
mu-tant would not necessarily be amultiple
mu-tant but one that might also have suffered a
lesion, which would alter the conformation of
the entire protein.
Adissociation of transformation parameters
has been reported with linesofmouse3T3cells
infected with athermosensitive mutant
poly-oma virus (25); however, in this case, though
transformed growth properties were
tempera-turesensitive, agglutinability byconcanavalin
A was enhanced at nonpermissive as well as
permissive temperatures. Hence, these cell
lines, assuming this dissociation is directly
brought about by a mutation in the viral ge-nome, exhibit not the same properties, but
rather properties complementary to those
shown by cells infected with the PTD mutants
discussed above.
Extensive analysis of the whole range of
transformationparametersexhibited bya
vari-etyof random clones ofmouse3T3cellsinfected
by simianvirus 40has beenreportedby Risser
and Pollack (31). In this work the viruswasnot
mutagenized; rather, the variability in
proper-ties detected among the different clonesis
as-sumed to derive fromthe nature ofthe
virus-cellinteraction. Thisapproach ledtothe recog-nitionof distinctcategories inthe alteration of
host growth controlafter infection; it was
possi-ble to identify clones that showed
anchorage-dependent growthbutretainedserum- and
den-sity-dependent growth properties.
Dissociation of transformation parameters like thatseenbetweenthedifferent simian
vi-rus40-infectedclones has beenrecognizedwith
cellsinfectedwithtwodifferent aviansarcoma
1052
BECKER ET AL. J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS
virusts mutants. tsLA334-infectedcellsexhibit
adegree of anchorage-independent growth,
al-though density-independent growth was not noted (9, 47). ts LA25-infected cells, although
almost completely anchorage dependent, show
a striking density- (22, 47) and
serum-inde-pendentgrowth pattern (Friis and Weber, un-published observations).
The involvement of a protease activity, now recognized as a urokinase-like plasminogen
ac-tivator(41) acting toproduce plasminin
serum-containing media (30), indetermining the
mor-phological (27) and many of the growth-related
properties of transformed cells (26) has been
further emphasized by the work of Pollack et
al. (29) and Shin etal. (35), which has shown
strong correlation among
anchorage-independ-ent growth, plasminogen activator protease,
and ability of cellsto inducetumors invivo in
"nude" mice.
Jones et al. (19) have further shown that
tumorsresulting from inoculation of low-level
plasminogen activator-producing cell lines are
invariably made up of high-level plasminogen activator-producing cells.
The results reported above with the PTD
mu-tantsdonotfundamentally contradict the
exis-tenceofacorrelation betweenplasminogen
ac-tivator production and oncogenicity. The PTD
mutants seem tobeasortof intermediate cate-gory in which the dissociation of
transforma-tion parameters isplaceddifferently. The
dem-onstrated anchorage-independent growth with
the PTD mutant-infected cells isparalleled by
protease activity, asthe caseinoverlay plaque
testshowed (Fig. 5).Furthermore, experiments
inour laboratory have shown that
anchorage-independent growth of transformed cells
re-quiresthe presenceof plasminogeninthe
soft-agar suspension culture medium (Becker, un-published observation). The fact that in vivo tumorformation is notefficient withthe PTD mutantsmayindeed reflectanimportant
func-tional difference between cell-bound and
super-natant plasminogen activator in contributing
totumorigenicity.
The presence of LETS glycoprotein on the
surfaces of PTD mutant-infected cells at the
nonpermissive temperature is an important
finding,because itarguesagainst the
hypothe-sis that LETS may play a regulatory role in
establishing density inhibition ofgrowth (15,
16).Ourfindings are, however, consistent with
the report that restoration of LETS
glycopro-tein to transformed cells results in increased
adhesion to substrate and more normal cell
morphology but failstoimpose
density-depend-entgrowth regulation (48), sincePTDmutants
at the nonpermissive temperature do exhibit
normal or near-normal morphology and
rela-tively high adhesion to substrate (Friis and
Weber, unpublished observations) under condi-tions approximating transformed cell growth.
The TD mutant tsGI201, which is unusual in
being non-conditionally defective for cell-bound and supernatant plasminogen activator pro-tease, brought useful insight into the question
ofthe mechanism by which LETS glycoprotein
is removed from transformed cells. ts GI201, which is transformed according to almost all
parameters at the permissive temperature,
shows only partial removal of LETS
glycopro-tein. Since the PTD mutant-infected cells at the
nonpermissive temperature, under conditions
where significant cell-bound plasminogen acti-vator is expressed and in medium containing
high plasmin activities, nonetheless exhibit
LETSglycoproteininnormal amounts, one can
speculate thatadifferent protease, onethat is
notplasmin and is perhaps not even involvedin
plasminogen activation, is responsible for
LETS removal.
The study of the transformation events
occur-ringafter avian sarcoma virus infection is very
complex because of the variety of parameters to
be monitored andthe unknown relationships of
the parameters to each other. Studies such as
that reported above may help to clarify the
relationships of the parameters, for example, the possible dependence on particular protease
activitiesfor loss of the LETS glycoprotein. Itis
to be hoped that examination of mutants such
as thesePTD mutantsandothers ofa
comple-mentary nature, i.e., mutantsdefective for
in-ducing growth parameters of transformation butlikethewild-typevirusfor inducingcertain
biochemical changes in infected cells (such as
increased hexose transport), could eventually
lead to recognition of distinct transformation
functions directly coded for by the sarcoma viral genome.
ACKNOWLEDGMENTS
Wewouldlike to express our thanks to Sigrun Bingel for veryable technicalassistance.
This work was supported by Sonderforschungsbereich47 of the DeutscheForschungsgemeinschaft.
LITERATURE CITED
1. Bader, J. P., and N. R. Brown. 1971. Inductions of mutations in an RNA tumor virusbyananalogueofa DNAprecursor. Nature(London) New Biol. 234:11-12.
2. Bader, J. P., D. A. David, and N. R. Brown. 1974. Accumulation ofwaterduringtransformation of cells byanaviansarcomavirus.Cell 3:307-313.
3. Balduzzi, P. C., and H.Murphy.1975.Plaqueassayof aviansarcomavirusesusingcasein. J.Virol. 16:707-711.
VOL. 21, 1977 1053
on November 10, 2019 by guest
http://jvi.asm.org/
1054 BECKER ET AL.
4. Biquard, J. M., and P.Vigier.1972.Characteristicsofa conditional mutant of Rous sarcoma virus defectivein abilitytotransform cells athigh temperature. Virol-ogy47:444-455.
5. Burger,M.M., and A. R.Goldberg.1967.Identification of a tumor-specific determinant on neoplastic cell surfaces. Proc. Natl. Acad. Sci. U.S.A.57:359-366. 6. Burger, M.M.,andG.S. Martin.1972. Agglutination
of cellstransformedbyRous sarcoma virusbywheat germagglutinin and concanavalin A. Nature (Lon-don)New Biol.237:9-12.
7. Chen,L.B.,and J. M.Buchanan.1975. Plasminogen-independent fibrinolysis by proteases produced by transformed chick embryo fibroblasts. Proc. Natl. Acad. Sci.U.S.A. 72:1132-1136.
8. Friis, R. R. 1972.Abortiveinfection of Japanesequail cells with avian sarcoma viruses. Virology 50:701-712.
9. Friis, R. R., K. Toyoshima, and P. K. Vogt. 1971. Conditionallethal mutants of avian sarcoma viruses. Physiologyof ts 75 and ts 149.Virology 43:375-389. 10. Goldberg, A. R. 1974. Increased protease levels in
transformedcells: a caseinoverlayassayfor the de-tection of plasminogen activator production. Cell 2:95-102.
11. Hatanaka, M.,and H. Hanafusa. 1970. Analysisof a functionalchangeinmembraneintheprocessofcell transformationbyRous sarcomavirus; alterationin the characteristics of sugar transport. Virology 41:647-652.
12. Hogg,N. M. 1974. Acomparison ofmembraneproteins of normal andtransformed cellsby lactoperoxidase labeling. Proc.Natl. Acad.Sci.U.S.A.71:489-492. 13. Holley,R. W., and J. A.Kiernan.1974.Controlof the
initiationof DNAsynthesisin3T3 cells: serum fac-tors.Proc.Natl. Acad.Sci. U.S.A. 71:2908-2911. 14. Hynes,R.0.1973.Alterationofcell-surfaceproteinsby
viraltransformation andbyproteolysis. Proc. Natl. Acad.Sci. U.S.A. 70:3170-3174.
15. Hynes, R. 0. 1974. Role of surfacealterations incell transformation: the importanceof proteases and sur-faceproteins.Cell1:147-156.
16. Hynes, R. O., and J. M. Bye. 1974. Densityand cell cycle dependenceofcell surfaceproteins in hamster fibroblasts. Cell3:113-120.
17. Hynes, R. O., andJ. A. Wyke. 1975. Alterationsin surfaceproteins inchicken cells transformedby tem-perature sensitive mutants of Roussarcomavirus. Virology 64:492-504.
18. Hynes, R.O., J. A.Wyke,J. M.Bye,K. C.Humphryes, and E. S.Pearlstein.1975.Areproteasesinvolvedin altering cell surfaceproteinsduring viral tranforma-tion?, p. 931-944. In E. Reich, E. Shaw, and D. B. Rifkin (ed.), Proteases andbiological control. Cold Spring HarborLaboratory,ColdSpring Harbor,N.Y. 19. Jones, P. A.,W. E.Lang,and W. F.Benedict. 1975. Fibrinolytic activity in a humanfibrosarcoma cell line and evidence for theinduction ofplasminogen activator secretion during tumor formation. Cell 6:245-252.
20. Kawai,S.,and H.Hanafusa.1971.Theeffects of recip-rocal changes in temperature in the transformed state of cells infected with a Rous sarcoma virus mutant. Virology46:470-479.
21. Kurth, R., and H. Bauer. 1973. Avian oncornavirus-induced tumorantigensofembryonicandunknown origin.Virology56:496-504.
22. Kurth, R.,R. R.Friis,J. A.Wyke,and H.Bauer.1975. Expression oftumor-specific surface antigenson cells infectedwithtemperature-sensitive mutantsofavian sarcoma virus.Virology64:400-408.
23. Macpherson,I., and L.Montagnier.1964.Agar
suspen-sion culture for the selective assay ofcells trans-formedbypolyomavirus.Virology23:291-294. 24. Martin,G.S.,S.Venuta,J.M.Weber,andH.Rubin.
1971. Temperature-dependent alteration in sugar
transportincells infectedbyatemperature-sensitive
mutantof Roussarcomavirus.Proc. Natl. Acad.Sci. U.S.A.68:2739-2741.
25. Okada, Y.S., and A.Hakura. 1975. Temperature-de-pendent propertiesofcellstransformedbya thermo-sensitivemutant(ts-121)ofpolyomavirus. II. Char-acterization of 121-6 cells. Int. J.Cancer16:394-403.
26. Ossowski, L.,J.P.Quiqley,G.M.Kellerman,and E. Reich.Fibrinolysisassociatedwithoncogenic trans-formation. Requirement ofplasminogen for
corre-latedchangesincellularmorphology, colony forma-tion in agar, and cell migration. J. Exp. Med.
138:1056-1064.
27. Ossowski, L.,J. C.Unkeless,A.Tobia,J.P.Quiqley,
D. B.Rifkin,and E.Reich.1973.Anenzymatic func-tionassociatedwithtransformationoffibroblastsby oncogenicviruses.II.Mammalian fibroblast cultures transformedbyDNA and RNAtumorviruses. J.Exp. Med. 137:112-126.
28. Pollack, R. E.,and M. M.Burger. 1969. Surface-spe-cificcharacteristics ofa contact-inhibited cell line containingtheSV-40viral genome. Proc. Natl.Acad. Sci.U.S.A.62:1074-1076.
29. Pollack, R. E.,R.Risser, andD.Rifkin. 1974.
Plas-minogenactivatorproductionaccompaniesloss of
an-chorage regulationintransformationofprimaryrat embryocellsbySimian virus40. Proc. Natl. Acad. Sci.U.S.A. 71:4792-4796.
30. Quiqley,J.P.,L.Ossowski,and E.Reich.1974.
Plas-minogen:theserumproenzymeactivatedby factors frommalignantcells. J. Biol.Chem.249:4306-4311.
31. Risser, R., and R. E. Pollack. 1974. A nonselective
analysisof SV40transformationofmouse3T3 cells.
Virology59:477-489.
32. Rosenblith, J. Z., T. E. Ukema, H. H. Yin, R. D.
Berkin,and M. J.Karnowsky.1973. Acomparative evaluation ofthe distribution ofConcanavalin
A-bindingsitesonthe surface ofnormalvirally
trans-formed,andprotease-treatedfibroblasts.Proc.Natl. Acad.Sci.U.S.A. 70:1625-1629.
33. Rubin,H. 1960. Ananalysisof the assay of Rous
sar-comacells in vitrobytheinfectivecentertechnique.
Virology10:29-49.
34. Ruoslahti, E.,and A. Vaheri.1974. Novel human se-rumprotein fromfibroblastplasma membrane. Na-ture(London)248:789-791.
35. Shin, S.-I.,V. H.Freedman,R.Risser,andR.Pollack. 1975. Tumorgenicity ofvirus-transformed cells in nude mice iscorrelated specificallywithanchorage independent growthinvitro. Proc. Natl. Acad. Sci. U.S.A. 72:4435-4439.
36. Stoker, M. 1968. Abortivetransformationbypolyoma virus.Nature(London) 218:234-238.
37. Stone, K. R., R. E. Smith, and W. K. Joklik. 1974.
Changesinmembranepolypeptides thatoccurwhen chick embryofibroblasts and NRK cellsare trans-formed with avian sarcoma viruses.Virology 58:86-100.
38. Temin,H. M.,andH. Rubin. 1958.Characteristicsof anassayforRoussarcomavirus and Roussarcoma cells intissue culture.Virology6:669-688.
39. Todaro, G. J., H. Green, and B. J. Goldberg. 1964. Transformation of properties ofanestablished cell linebySV40 andpolyomaviruses. Proc. Natl. Acad. Sci.U.S.A.51:66-73.
40. Unkeless,J.C., K. D.Dan, G. M. Kellerman, and E. Reich. 1974. Fibrinolysis associated with oncogenic transformation. J.Biol.Chem. 249:4295-4305.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
DIFFERENT TRANSFORMATION-DEFECTIVE MUTANTS
41. Unkeless, J. C., A. Tobia, L. Ossowski, J.P. Quiqley, D.Rifkin, and E. Reich. 1973. An enzymatic function associated withtransformation of fibroblasts by
onco-genic viruses. I Chick embryo fibroblast cultures transformed by avian RNA tumorviruses. J. Exp. Med. 137:85-111.
42. Vogt, P. K., R. A. Weiss, and H. Hanafusa. 1974. Proposal of numberingmutantsof avian leukosisand
sarcomaviruses. J.Virol. 13:551-554.
43. Weber,J. M. 1973. Hexosetransportinnormal and in Roussarcomavirus-transformed cells. J. Biol. Chem.
248:2978-2983.
44. Weber, J. M. 1975. Inhibition ofprotease activity in culturesof Roussarcomavirus-transformed cells:
ef-fectonthetransformed phenotype. Cell 5:253-261. 45. Wickus, G. G., P. E. Branton,and P. W. Robbins.
1974.Roussarcomavirustranformation of the chick
cell surface,p. 541-546.In B. Clarkson and K. Bas-erga (ed.), Control of proliferation in animal cells.
Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
46. Wyke, J. A.1971.A method of isolating cellsincapable ofmultiplication in suspension culture. Exp. Cell. Res. 66:203-208.
47. Wyke, J. A., and M. Linidal. 1973.Temperature sensi-tive aviansarcomaviruses. Aphysiological compari-sonoftwenty mutants.Virology 53:152-161. 48. Yamada, K. M., S. S. Yamada, and I. Pastan. 1976.
Cell surface protein partially restores morphologi-cally, adhesiveness, andcontactinhibition of
move-mentoftransformed fibroblasts. Proc. Natl. Acad. Sci. U.S.A. 73:1217-1221.
VOL. 21, 1977 1055
on November 10, 2019 by guest
http://jvi.asm.org/