Copyright © 1974 AmericanSocietyforMicrobiology Printed in U.S.A.
Intracellular
Virus-Specific Structures and RNAs in
Oncornavirus-Producing Human Cells
A. G. BUKRINSKAYA, G. G. MILLER, E. N. LEBEDEVA, AND V. M. ZHDANOV
TheD.I.IvanovskyInstituteof Virologyand the N. F.GamaleyaInstituteofMicrobiologyandEpidemiology, Academyof Medical Sciences, Moscow, U.S.S.R.
Received forpublication 18September1973
Two kinds of virus-specific structures were isolated from the cytoplasm of
Detroit-6 and human amnion cells producing oncornavirus-like particles.These
structures represented A particles with the diameter of 70 to 80 nm and
aggregated strands of nucleocapsids with the diameter of 3 and 6 nm. The
structures were separated from cellular contaminants by isopycnic banding in
linear sucrosegradients and subsequently further purified by sedimentation in
velocity sucrose gradients. Their sedimentation coefficient was 250 and 150S,
respectively. Both structures contain 60, 45, and 35S RNA species, and 150S
structuresalsocontained20SRNA. The 35and 20S RNA from the 150S structure
formed hybrids with DNA enzymatically synthesized on extracellular virions.
The structures displayed endogeneous polymerase activity, DNAproductofthe
reactionbeingpredominantly associated with60SRNA. No 70S RNAwasfound
in the cellstructures ofvarious densities. Also, the virions purifiedfrom tissue
culture fluid contained 70SRNA. These findings are consistent with those on
extracellular maturation ofoncornavirusRNA.
Little information is available concerning the properties of intracellular precursors of
oncor-naviruses and virus-specific RNA in
oncor-navirus-infected cells. Coffin and Temin (4) isolated, from Rous sarcoma-infected cells, vi-rus-specific particles which banded in an equi-librium sucrose density gradient at 1.24 g/ml.
These particles possessed RNA-dependent
DNApolymeraseactivitywhich was resistant to ribonucleasedigestion. However, they were not further studied. Smith and Wivel, and Wivel
and Yang (14, 17) reported the presence of A
particles in cells producing oncornaviruses of
B andCtype. Theseparticles contained,among
otherheterogeneous RNA species, high
molecu-larweight RNA. Intracellular A particles were
described also in monkey cells producing
Ma-son-Pfizeroncornavirus of B type (8).
The data on the properties of intracellular
virus-specific RNA and the site of itssynthesis
are also very scarce. Parental 60 to 70S RNA
wasshown topenetrate the nuclei early in
infec-tion (5), and de novosynthesized heterogeneous
virus-specific RNA including high molecular weight species were isolated from the nuclei and
thecytoplasm(9). On the other hand, Tsuchida
et al. (15, 16) extracted from
oncornavirus-infected cells 35 and 20S RNA species, and did
not find anyhigh molecular weight RNA.
The results mentioned above were obtained
mainly on the systems of animal cells producing
oncornaviruses type C. As forthe cells of human
originproducingoncornavirus typeB, there are
no available dataconcerningthisquestion.
In this investigation we present the data on
the properties of cytoplasmic virus-specific
structuresand RNAextracted fromthese
struc-tures ofDetroit-6 and human amnioncells, two
of humancontinuous lineswhich spontaneously
produce oncornavirus type B. The
characteris-tics of these cell systems have been published
earlier (1, 18).
MATERIALS AND METHODS
Cells. Cell monolayers were grown in 199 medium supplied with 2% calfserum, and within 72to 96 h they were fed with fresh medium containing 3H-uri-dine(2 gCi/ml, specificactivity 16 Ci/mmol) or car-rier-free 32P04 (1 iACi/ml) inthe phosphate-free me-dium. After 24 h of incubation, cell cultures were washed withTris-saline (Tris-hydrochloride, 0.01 M, pH7.4;NaCl,0.1M) andthecellswereremoved from theglass by0.5%trypsin.
Virus. The virus from tissue culture fluid was concentrated as described (3) and subjected to equi-librium densitygradient centrifugation in 20 to 60% sucrose (wt/wt) in TNE buffer (Tris-hydrochloride 0.05M, pH8.3;NaCl,0.15M; EDTA,0.001M). The viruswasrecovered fromdensity gradientfractionsby pelletingandresuspended inthe same buffer.
Preparaton and analysis of cytoplasmic ex-478
on November 10, 2019 by guest
http://jvi.asm.org/
tracts. Thecells were washed at 4 C with Tris-saline and thenwith RSB (Tris-hydrochloride, 0.01 M pH 7.4; NaCl, 0.01 M;MgCl2,0.003 M) and disrupted in RSBby Dounce homogenization. Nuclei and cell rem-nantswere pelleted at 800 x g. Post-nuclear fraction was centrifuged at 15,000 xg to remove mitochondria and microsomes, supernatants (cytoplasmic extracts) inthe volume of5ml were layered over a 25 to 65% (wt/wt) 10-ml linear sucrose gradient prepared on TNEbuffer, and the structures were banded isopycni-callyby centrifugation in anSW 27.1 bucket rotor at 25,000 rpm for 16 to 18 h at 4 C. The upper layer containing the material was removed, the gradients weredivided into fractions, 0.1-volume sample from each was precipitated by 5% trichloroacetic acid along with 25 to 50 gg ofcasein. Radioactivity was countedin aPackard Tri-Carbscintillation spectrom-eterby usinga toluene-based scintillant. The appro-priate fractionscorresponding to the peaks of radio-activity werepooled, twice diluted with thesame buf-fer, and either recentrifuged in velocity sucrose gradients (17 to 40% wt/vol) in an SW 27.1 bucket rotor at 24,000 rpm for1.5hat 4C, orusedforRNA, analysis.
Polymerase assay.Cytoplasmicextractsfrom
un-labeled cells were prepared as described above and
centrifuged in 25 to 65% equilibrium sucrose
gradi-ents.DNApolymeraseactivity wasassayedeither in crudeextractsorinportions ofeach fraction in
reac-tion mixture containingthe following: Tris, pH 8.3,
0.05M;KCl, 0.15M;MgCl2,0.006M; dATP,dCTP,
anddGTP,0.1 mMeach;dithiothreitol (DTT),0.2%;
'H-TTP, 1
;iCi
(specific activity 26 Ci/mmol). Themixture in avolume of0.5mlwasincubatedat30C for15minandchilled, and acid-insolubleradioactivity
was counted. In some experiments, the material of each fractionwasdivided intotwoparts,onepartwas
used for radioactivity determination, and the other part foranalysis of nucleic acids by velocity sucrose gradient centrifugation.
Extraction of RNA from thecytoplasmic struc-tures and its velocity sucrose gradient centrifu-gation. RNA from sucrose gradient fractions after centrifugation ofcytoplasmicextracts was extracted eitherby1%sodiumdodecyl sulfate (SDS) or by the
SDS-phenol method followed by precipitation with
ethanol. Both methodsyielded thesameresults. RNA precipitates werecollected bycentrifugation and redissolvedinTNEbuffercontaining0.5%SDS. All RNA samples were centrifuged in 15 to 30%
(wt/vol) sucrose gradients prepared on STE
(Tris-hydrochloride,0.01 M, pH 7.4; NaCl, 0.15M; EDTA,
0.001M) containing0.5%SDS in anSW27.1bucket rotor at14,000to 16,000 rpmfor 14 to16 hat25C.
RNA-DNA hybridization. 3H-labeled DNA was obtainedas aproductofreversetranscriptasereaction by usingdetergent-disrupted virions from tissue cul-ture fluid as a template. Polymerase reaction was
performedin a2-ml volume byusingthepolymerase
mixturementioned above andcontaining0.02%NP40 which was incubated for 1 h at 35C. Then the polymerasemixturewaschilled,and thenucleic acids wereextracted twice with SDS-phenol. Purified calf thymus DNA was added as a carrier. The nucleic acidswereprecipitated bytwovolumes ofethanoland
left overnight. The pellet was redissolved in STE, RNA was hydrolyzed by 0.5 MNaOH at 37 C during a 16-h period, and after neutralization, DNA was pre-cipitated by ethanol and left overnight. DNA was then dissolvedin70%formamide, melted at 90 C for 2 min, and chilled andmixed with an equal volume of RNA dissolved in 30% formamidecontaining 0.8 M NaCl. The mixture was incubated at 37 C for 16 to 20 h and the extent ofRNA-DNA hybrid formation was mea-suredby the method of Hehlman, Kufe, and Spiegel-man (7) by using isopycnic centrifugation in pre-formed caesium sulfate gradients in a Ti5O rotor at 35,000 rpm for 60h.
Electron microscopy. Cytoplasmic extracts from unlabeled cells mixed with a little portion of the extract from labeled cells was centrifuged in equilib-rium sucrose density gradients. After counting radio-activity in portions, the appropriate fractions were pooled and diluted with TNEbuffer, and the material waspelletedat80,000 x g for30min, dissolved in the same buffer, negatively stained with uranyl acetate, and examined in an electron microscope (JEM 100B).
Forultra-thin sections, the monolayer culture cells were fixed in Dalton's chrome-osmium, dehydrated, embedded in Epon-araldite, and sectioned and stained withuranyl acetate and lead citrate.
RESULTS
Distribution of radioactivity after isopyc-nic centrifugation of cytoplasmic extracts. Cytoplasmic extracts were prepared from the cells labeledwith 3H-uridineor37P0o,and sep-arated in
equilibrium
sucrosedensity
gradi-ents. For comparison, cytoplasmic extracts of
an HeLa cell strain, which does not produce virus-like particles (as revealed by electron
microscopy), were analyzed. Figure la shows
that radioactivity in this case is found in the
region ofdensity 1.18 g/ml. According to
pub-lished data (14), thisregioncontains mitochon-dria, microsomes, and cytoplasmic fragments.
An essential part of radioactivity was also
found in this region after isopycnic
centrifuga-tion ofAO and Detroit-6 cytoplasmic extracts.
Besides, some part of radioactivity was
regu-larly revealed in the region of density 1.22 to
1.23 and 1.27 to 1.29
g/ml
(Fig. lb). Thesecomponents werereproduciblyobserved in both
cell lines irrespective ofwhether the cells were
labeled with
32po0
or 3H-uridine. The 3-h cen-trifugation yielded the same results as the centrifugationduring
16 h. Addition of 0.4%DTT to the bufferdid notchangethe
distribu-tion ofradioactivity (data notshown).
Polymerase activity assay of the gradient fractions. To identify the possible precursors
of oncornavirusesin thegradient, each fraction
ofthegradient similar to that shownin Fig. lb
was assayed for endogenous
polymerase
activ-ity in theabsenceofdetergent.Thereactionwas
479
on November 10, 2019 by guest
http://jvi.asm.org/
performed for a short period of time (15 min) to
detect DNA still associated withthetemplate.
It is seen from Fig. 2a that the highest
polymer-aseactivity is revealed at the density of 1.22 and
1.19
g/ml,
and thelower activity is found also at1.27to 1.29 and 1.23g/ml.
To identify thereaction as virus specific, the nucleic acids were extracted from the
incuba-a!
.2', 1.21 1.18 1.15
g,
1.29 I.? 1.23 1.20 1.17
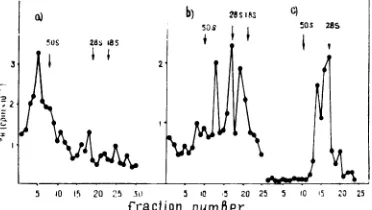
FIG. 1. Distribution of radioactivity after equilib-riumdensity centrifugationofcytoplasmicextractsof HeLa (a) and humanamnion (b) cells. Cytoplasmic
extractsfrom3H-labeledHeLa cells (a)and AOcells (b) were centrifuged in a 25 to 65% equilibrium
sucrosegradient prepared on TNE buffer inanSW
27.1 bucketrotor at25,000rpmfor 16 hat4C.After fractionation, radioactivitywascounted in0.1-volume
portions.
tion mixture after polymerase reaction with
crude cytoplasmic extracts as well as from the
gradient fractions corresponding to peaks of
radioactivity and were analyzed in velocity
sucrose gradients. Figure 2b shows that an
essential part of 3H-labeled DNA extracted
fromcytoplasmic extract sediments at 60S. The
similar sedimentation profile was observed for
DNAextractedfromthe 1.22 (Fig.2c) and 1.19
zone of the gradient (data not shown), and the
lesser part of the label was found in the region
lighterthan 60S (Fig. 2c). The60S peak
disap-peared after treatment with RNase (Fig. 2b)
indicating that DNA in this position is associ-ated with RNA.
Sedimentation characteristics of the com-ponents from different
density
regions.
The material from the regions of the gradient dis-playingthepolymerase activitywasfurtherana-lyzed in velocity sucrose gradients. Figure 3a
shows that an essentialpart oflabeledmaterial
from the 1.22 to 1.23 region of both cell lines
sediments at 250 to 300S and the lesser part of radioactivity is found near the bottom of the gradient. The last component has not been
regularly observed and may be a product of
aggregation of the abovementioned component.
The similar sedimentation profilewasobtained
when the material from the 1.27 to 1.29 zone wasanalyzed (datanotshown).Thematerialof density 1.19g/ml consistedof two components,
1.260 1.235 1.210 1.185 1.160 5 10 15 20 25
[image:3.493.58.248.161.303.2]g rraction number 5 Fraction10 15 nuuiner20 25
FIG. 2. Distributionof polymeraseactivityinequilibriumsucrosegradient fractions (a) after centrifugation
ofcytoplasmic extractsfromDetroit-6cells and analysis of DNA product (b-c). Theendogenouspolymerase activitywasassayedineachfraction ofthegradientsimilartothatshowninFig.lb in thevolume of0.5mlby using 9H-TTPas aradioactiveprecoursor, andradioactivity wascounted in '/8-volume portions (a). b, The
polymerasereaction withcytoplasmicextractswasperformedat30C for15min, andthen thenucleic acidswere
extractedfrom theincubation mixturewithSDS-phenolanddividedintwoparts,onepart(solid circles)was
left untreated, and theotherpart(empty circles)wastreated with 20ugofpancreaticribonucleasepermlat room temperature for20 min. The material was analyzed by centrifugation in 15 to 30% (wt/vol) velocity sucrosegradientpreparedonSTE buffercontaining0.5%SDS inanSW27.1bucketrotorat15,000rpmfor15h
at25C.c,Thefractionscorrespondingtothe peak ofradioactivityat1.22g/mlwerepooled,diluted with STE,
andnucleic acidswereextracted with 1%SDS and analyzedasinb. Nucleicacidwasextracted from purified
32P-labeled Sendai virions by 1% SDS and usedas a marker (50S) run in thesame gradient. 28and 18S ribosomal RNA extracted from Ehrlichtumorcellswerealsorunin thesamegradient andrevealed byoptical
density.
15
10
a
C:L
In5
480
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.112.396.401.526.2]ONCORNAVIRUS-PRODUCINGHUMAN CELLS
one of whichsedimentedatabout150Sand the other whichwas in alighterregion of thesucrose gradient (Fig. 3b).
Analysis ofRNA. RNA was extracted with SDS or SDS-phenol from cytoplasmic extracts andsucrose gradientfractionsand wasanalyzed
in 15 to 30% linear sucrosegradients.
Figure4ashows thatbesides28and18SRNA species, total cytoplasmic RNA from AO cells contains a relativelyhigh percentage of 35 and
a) 115s
-3 n
.4J E
I' 2
[image:4.493.44.234.187.309.2]Fraction iLimBer Fract-on nwr'8tr
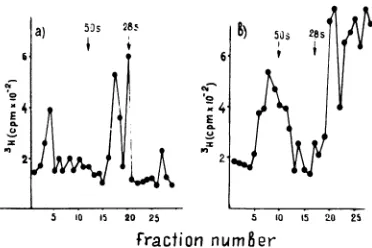
FIG. 3. Velocity sedimentation of cytoplasmic structures from Detroit-6 and AO cells. Cytoplasmic extracts from 32P-labeled Detroit-6 cells and 3H-labeled AO cells were separated in an equilibrium
sucrose gradient prepared on TNE; the fractions
corresponding tothepeak ofradioactivity at1.22to
1.23g/ml (a), and1.19g/ml (b)werepooled, diluted with TNE, andrecentrifugedina17 to 40%(wt/vol)
velocitysucrosedensity gradientat24,000 rpm for1.5
hat4C. Tobaccomosaic virus(185S) was runin the
same gradients and revealed by optical density.
Symbolsfora: 0, 32P;0, 3H.
28s
a) i
5 10 15 20 25
45S RNA (about 10to 15% and 1to 2%of total RNA,
respectively).
The similar results wereobtained
with Detroit-6 cells.When RNA wasextractedfrom the fractions
of equilibrium sucrose gradient, 28 and 18S RNA specieswere invariably found throughout the gradient. Therefore, to
obtain
more pure material containing virus-specific RNA species,RNA wasextracted from theappropriate zones
of velocity sucrose gradients after recentrifu-gationofequilibrium sucrose gradient fractions. The 250S structure of density 1.27 to 1.29 and
1.22 to 1.23
g/ml,
the 150S structureofdensity1.19
g/ml,
and the top component of the same density shown inFig. 3b wereanalyzed.RNA extracted from the top component of density 1.19 g/ml contained only 28 and 18S
species. Thus, this component apparently
rep-resents a cellularcomponent.
The mainsize-classesofRNA in the 250 and 150S structures were 60, 45, and 35S RNA. Besides this, the150S structures contained 20S RNA. The relationship between these RNA
species varied from experiment to experiment,
but usually60 and 35S dominated in the250S structureand 35 and20S dominatedin the150S structures (Fig. 4b, c). Rapidly sedimenting RNA species were also seen in a number of experiments (Fig. 4b) in accordance with the data
published
(9).DNA-RNA hybridization. The 35 and 20S RNAs extracted from the 150S component of density 1.19
g/ml
were annealed with3H-DNA
productenzymatically synthesizedon the
extra-5 10 15 20 25
Ft-action number
c)
28318S2
aoS
5 10 15 20 25
FIG. 4. Sedimentation analysis oftotal cytoplasmic RNA (a) and RNA extracted from the cytoplasmic
structures (b, c) of Detroit-6 and AG cells. a, RNA from cytoplasmic extracts ofAO cells extracted by SDS-phenol; b,RNAextractedfromthe250Sstructuresof density1.22 to1.23g/ml.Cytoplasmic extractsfrom 32P-labeledAOcells and 3H-labeled Detroit-6 cellswereseparatedinequilibriumsucrosegradients,fractions correspondingtothepeak ofradioactivityat 1.22 to 1.23g/mlwerepooledandrecentrifugedinavelocity
su-crosegradientasinFig. 3a,fractions corresponding tothe250Speak werepooledanddiluted withSTE,and RNAwasextractedwith 1% SDS.Symbols:0,32p;0,3H.c,RNAextractedfrom the150Sstructures ofdensity
1.19g/ml. Cytoplasmicextractsfrom'H-labeled AOcellswerecentrifuged, the150Sstructurewasobtainedas described inFig.3b, theappropriate fractions werepooledanddiluted, and theRNA wasextracted with 1% SDS. RNAwasanalyzedin 15to30%sucrosedensitygradients preparedonSTEcontaining0.5%SDSin anSW 27.1 bucketrotor at 16,000rpmfor14 to16 hat 25 C. ThesymbolsareasinFig.2.
1.5-E-I
481
VOL.13,1974on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.97.391.432.554.2]cellular virions, and the hybridizationreaction
wasevaluated by centrifugation in a preformed
Cs2SO4 gradient. RNA extractedfrom the cyto-plasm of normal human fibroblasts was used as
a control.
The product synthesized duringl h of
polym-erase reaction ontheextracellularvirions
repre-sented DNA-RNA hybrids that banded at the
density of 1.54 g/ml in Cs2SO4 gradients, and
after alkaline hydrolysis a predominant
compo-nent was one-stranded DNA banded at 1.46
g/ml (Fig. 5a). Also, an essential part ofDNA
radioactivity shifted to the regions of higher
density after the hybridization reaction (Fig.
5b, c). No detectable hybridization was
ob-servedbetweenthe DNA product and the
cellu-larRNAfromfibroblast(Fig. 5d).
Properties of 60S RNA and RNA extracted
from the virions. To characterize further 60S
RNA, it was dissociated into its subunits. With
this purpose, RNA from the 1.23 cytoplasmic
structures was centrifuged, and fractions
con-taining 60S RNA were pooled, precipitated in
ethanol, and dissolved inSTE.
When recentrifuged immediately, 60S RNA
exhibited a rather broad peak ofradioactivity
at55 to65S(Fig. 6a). After heating at 90 C for 2
min, it dissociated into components with
sedi-mentation coefficients of 35, 28, and 20S (Fig.
6b). When recentrifugation was not performed immediately and the samples were left over-night in 1% SDS, 60S RNA spontaneously dissociated into 35 and 28S components (Fig. 6c).
Virions from tissue culture fluid werepurified
bycentrifugation on an equilibrium sucrose
gra-dient; RNA was extracted from the 1.17 peak
and analyzed similarly. High molecular weight
RNA was found in the heavier region of the
sucrose gradientthan 60SRNA extracted from
thecells, the components 35, 28, and 4Sbeing
possible products of its dissociation (Fig. 7a). The sedimentation profileofrecentrifuged cyto-plasmic60S RNA from the 1.19 zone of
equilib-rium sucrosegradient run under the similar
con-ditions is shown forcomparison(Fig. 7b). It can
be seen that radioactivity of RNA extracted
from the virions doesnot coincide with that of
RNA extracted from the cytoplasmic
struc-tures.
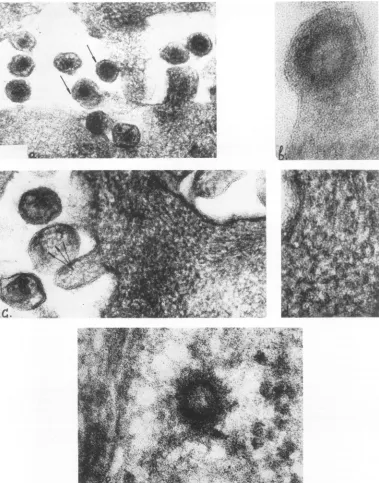
Electronmicroscopy. Examination of
ultra-thin sections of Detroit-6 cells revealed alarge
number of A and B particles in intercellular
spaces, and some of them were in the process of budding from the cell surface (Fig. 8a).Ahigh magnification ofa budding Aparticle is shown
in Fig. 8b. Ofspecialinterest are the
filamen-toushelicalstructuresseenunder the cell
mem-brane and showninFig.8c and d. The diameter
of these filaments is about 6 nm. Similar
a)
IOS OSs 80
uS5 28sWS
i
1'
b) 28s'1S C)
50S I 5OS 285
I I
5 ...101.5 20 25 301 5 10 I5 20 25 5 iO i5 210 25
[image:5.493.267.453.288.393.2]fraction numBer
FIG. 6. Dissociation of60S cytoplasmic RNA from Detroit-6 cells. Fractions correspondingtothepeakat 60Sfrom the gradients similartothat shown inFig. 4b werepooled, diluted with STE, divided in twoparts, and onepart (b) washeated to 90 C for2 min and quickly chilled. Both samples were recentrifuged in 15 to 30% sucrosegradientsasinFig. 4. c, 60S RNA from thesame zoneof the gradientasarecentrifuged
onethefollowing dayas described in Fig.4.
b a) )c d)
8a4
112
1.61 156 151 146 1.,1 162 1.55 1.4s 141 163 155 i.47 141 163 158 i.52 146
FIG. 5. Hybridization of the 3H-DNA product of the endogenous virion DNA polymerase system with 20 and 35S RNA extracted from the 150Sstructures ofAO cells. RNA was extracted by SDSfrom the 150S structuresof density 1.19g/ml, and 20 and35Sspecieswere obtained as described in Fig. 4. Cytoplasmic RNA extracted byphenol-SDS from theprimary tissue culture ofhumanfibroblasts wasused as a control. The hybridization experiment was set as described (2), and all samples were analyzed by Cs2SO4 equilibrium density gradient centrifugationinTi5Orotor at35,000 rpmfor72h at 5 C.a,3H-DNA;b,c,d,reaction mixture after annealing with20SRNA, 35S RNA, and RNA ofhumanfibroblasts, respectively.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.113.398.492.591.2]5 ,0 ,5 20 25 5 10 I5 "0 25
[image:6.493.47.233.70.194.2]fraction
numBer
FIG. 7. Sedimentation analysis ofRNA extracted from virions (a) and from cytoplasmic structures of density 1.19g/ml (b). a, RNA was extracted by 1%
SDS from the virions purified by equilibriumsucrose
density centrifugation, and high molecular weight RNAwasrecentrifuged in 15to30%osucrosegradient
containing 0.5% SDS as in Fig. 4. b, Cytoplasmic RNA was extracted by 1% SDS from the region of
density1.19g/ml,andhigh molecular weightRNAwas
recentrifuged asin Fig. 4.
filamentous helical structures ofthe same
di-ameter are seen inside some extracellular
vi-rions. Figure 8e shows one of the A particles
regularly seeninthe hyaloplasm.
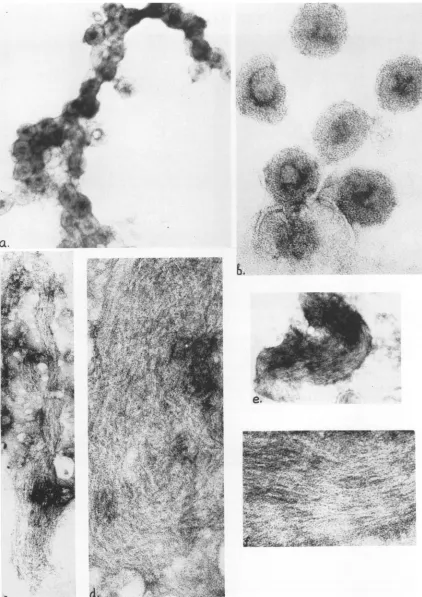
Figure 9 shows the material of equilibrium
sucrose gradient fractions correspondingto the peaks of radioactivity.
The material from the 1.22 to 1.23 region
contained a high concentration of A particles
with the diameter of 78 to 80nm (Fig. 9a, b).
Partly disaggregated A particles with released
RNP strands of diameter 6 and 3 nm were
also observed. Morphologically similar
struc-tureswere seeninthe material of density 1.22to
1.23 and 1.27 to 1.29 g/ml from AO cells (data
notshown).
The fractions from 1.19 zone of the gradient
contained rather homogeneous material which
represented clusters of aggregated filamentous
heliceswith the diameter of6nm(Fig. 9c-f). In
some partsthese helices were seen as unwound
threads of diameter 3 nm (Fig. 9d). The
scattered membraneous structures were also
seen inthis material.
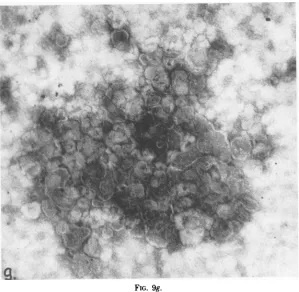
The fractions ofdensity 1.18 g/ml from
De-troit-6, AO and HeLa cells contained
mem-braneousstructures ofvarious sizesbutdidnot
contain filaments (Fig. 9g).
DISCUSSION
Detroit-6 and AO cellsbelongtothegroupof
cell strains of human origin which
spontane-ously produce oncornavirus particles of B type
(1, 18). We made an attempt to isolate the
intracellular precursors ofoncornavirus and to
investigate their RNA. The high concentration
of virus-specific structures was achieved by equilibrium sucrose density centrifugation of cytoplasmicextractsfollowedby centrifugation
in velocity sucrose gradients. Intracytoplasmic
A particles similar to that found in ultra-thin
sections were revealed in the region of density
1.22 to 1.23
g/ml
and 1.27 to 1.29 aftercentrifu-gation of cytoplasmic extracts, in accordance with the data of Smith and Wivel (14). Clusters ofaggregated filamentous heliceswere found in
the region of the gradient with the density of
1.19
g/ml.
These helices had a diameter of 6 nm; unwound threads of diameter 3 nm werealso seen in these preparations. The identical
filamentous helices were revealed in ultra-thin
sections ofthese cells. The similarity of these
structures totheinner component of the cores of
oncomaviruses (11) suggests a virus-specific
or-iginand allowsone to assume that these
struc-tures represent helical nucleocapsids of
oncor-naviruses. It is not clear why their density was
significantly lower than that expected for ribo-nucleoprotein structures. Apparentlythey were
either associated with membraneous material
whichwas seen inthese gradient fractions when observed in electron microscope, or contained lipids as aconstituent.
TheAparticleswereshownto cohtain 60and 35S RNA andthe 150Sstructurecontained 60, 45, 35, and 20S RNA. These RNA specieswere described as virus specific in the systems pro-ducing oncornaviruses ofC type (3, 10, 15, 16).
It must be mentioned that, unlike in that
system, a
significantly highet
percentage of 35and 45S RNA (about 10 to 15% and 1 to 2%, respectively, ofthe totalcytoplasmic RNA) was found inthecell lines used.
Both kinds of
virus-specific
structures, (Aparticles andfilaments) possessed endogeneous polymerase activity in the absence of
deter-gents. This result makes it evident that the
structures lack virus proteins which block the
polymerase activityinthe mature virions.Since the DNA product of polymerase reaction was predominantly assocated with 60S RNA, it is suggested that this RNA is a much better template for DNA polymerization than other RNA
species.
Thesignificance
of thesestruc-tures in virus
reproduction
and theirinterrela-tion in the cells requires further investigation.
Since35and 20SRNAextractedfromthe
150S
structures were shownto possess sequences
ho-mologous to those found in extracellular virion
RNAtheymay be
regarded
asitspossible
intra-cellular precursors.
ThesameRNAspecies, 35and
20S,
werealsofoundinthecell nucleibeingextractedat60C.
They also possessed a sequence homologous to
483
on November 10, 2019 by guest
http://jvi.asm.org/
BUKRINSKAYA ETAL.
.,,.v=,4stZ, B
%$w.v. X
*; Ss }
f$9.4...*.5
Fsws ,4g i, K gAlan;24t ss;X4
ttww'_
>f>sJffiffi
4;
vi
[image:7.493.70.449.74.557.2][F-A'
j''-
H
FIG. 8. Ultra-thin sections of Detroit-6 cells. a, Low magnification of the cell surface showing virions in intercellularspaces and budding A and B virions (arrows). x135,000. b, Highermagnification of theabove. Budding A particle is seen. x390,000. c, Sections of the cell surface showing the clusters offilamentous nucleocapsids underthecell membrane. x190,000.d,Higher magnification of the above. x390,000.e, Atypical nucleoid ofAparticle inthehyaloplasm. x390,000.
J. VIROL.
484
-7t
Ia
on November 10, 2019 by guest
http://jvi.asm.org/
'II
iV
L-I
. .4.
[image:8.493.29.451.40.637.2]_r I
|~~~~~~~~~~¶~ -
-r vs v v~~~~4 4'-
-FIG. 9. Negatively stained preparations of equilibrium sucrose gradient fractions of different density. The material from Detroit-6 cells. a, aggregated A particles in the 1.23 band. x 135,000. b, Highmagnification of the above. x390,000. c-f. Clusters of filamentous nucleocapsids in the 1.19 band. c, x 135,000; e, x65,000; d, f, x390,000. g, Membraneous structures in the 1.18 band. x135,000.
485
C.
.A
WI'
.41,
11.v
11-.. I,
4;
4.,
i< ---...
.. .. p
.k
V:,
I,..;.
,L:;..
.. -.I
iv t
eI.
ti I.. I
dOL
I V..-!
.i
on November 10, 2019 by guest
http://jvi.asm.org/
ET AL.
. .a
go
FIG. 9g.
thatinextracellularvirionRNA,indicatingthat
nuclei play someessential role in oncornavirus
multiplication (manuscript in preparation).
Our results show that eitherno70S RNAor a very small portion exists in the cells, and that
intracellular high molecular weight RNA
sedi-ments slower than RNA extracted from extra-cellular virions. These data are consistent with
the data obtained on oncornaviruses oftype C about extracellular maturation of virion RNA
(2, 3).
ACKNOWLEDGMENTS
Wewishtoexpress ourthankstoNatalyVorkunova for her excellent technical assistance.
ADDENDUM IN PROOF
We haverecently found that RNA extracted from purifiedintracytoplasmic A particles doesnotcontain
the sequences homologous to extracellular virus
ge-nome. When RNA extractedfrom the 250 and 150S
intracellular structures was hybridized to excess of DNAenzymatically synthesized on the extracellular
viriontemplateinthepresenceofactinomycin D,the percentage ofhybridizationasrevealedbyRNA
frac-tion resistant to RNase hydrolysis was 0 and 80 to 90%, respectively. We assume, therefore, that the 150Sstructures butnotAparticlesaret!heprecursors
oftheextracellularvirions.Aparticlesmayrepresent
aseparate oncornavirus orits precursor(manuscript inpreparation).
LITERATURE CITED
1. Bykovsky, A. P., G. G. Miller, N. V. Klisunova, L. K. Gorokhova, V.A. Zakhaleva, and V. B.Martynenko. 1973.Morphologyof oncornaviruses types A, B and C of continuous humanand animal cell lines.Vopr. Virusol. 2:215-221.
2. Canaani, E., K. V. D. Helm, and P. Duesberg. 1973. Evidence for 30-40 SRNAas precursorofthe60-70S RNA of Rous sarcoma virus. Proc. Nat. Acad. Sci. U.S.A. 70:401-405.
3. Cheung, K. S., R. E. Smith, M. P. Stone, and W.K. Joklik. 1972. Comparison ofimmature (rapid harvest) and mature Rous sarcoma virus particles. Virology
50:851-864.
4. Coffin, J. M., and H. M. Temin. 1971. Comparison of Roussarcomavirus-specific deoxyribonucleicacid
po-lymerases in virions of Roussarcomavirus and inRous
sarcoma virus-infected chicken cells. J. Virol.
7:625-634.
5. Dales, S., and H. Hanafusa. 1972. Penetration and intracellular release of thegenomeof avianRNAtumor
viruses.Virology50:440-458.
6. Green, M., H. Rokutanda, and M. Rokutanda, 1971. Virus-specific RNA incellstransformedby RNA
tu-morviruses. Nature N. Biol. 230:229-232.
7. Hehlmann,R., D.Kufe,and S.Spiegelman.1972. RNA inhumanleukemic cells relatedtothe RNA ofa mouse
leukemia virus. Proc. Nat. Acad. Sci. U.S.A. 69:435-439.
8. Kramarsky, B., N. H. Sarkar, and D. H. Moore. 1971. Ultrastructuralcomparison ofavirusfrom a rhesus-monkey mammary carcinoma with four oncogenic RNA viruses. Proc. Nat. Acad. Sci. U.S.A. 68:1603-1607.
9. Leong,J.A., A. C.Garapin,N.Jackson,L.Fanshier,W.
486
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.493.107.406.70.362.2]Levinson, and J. M. Bihop. 1972. Virus-specific RNA incells producing Roussarcoma virus:detection and characterization.J.Virol,9:891-902.
10. McCain, B., N. Biswal, and M. Benyesh-Melnick. 1973. Thesubunitsof murinesarcoma-leukemia virus RNA. J.Gen.Virol. 18:69-74.
11. Nermut,M.V.,H.Frank, andW.Schiafer.1972. Proper-ties of mouse leukemia viruses. III. Electron micro-scopicappearance asrevealedafterconventional
prep-arationtechniquesaswellasdrying and
freeze-etching.Virology49:345-358.
12. Parsons, J. T., J.M. Coffin, R.K.Haroz,P.A.Bromley, andC. Weissmann. 1973. Quantitative determination and locationofnewly synthesized virus-specific ribonu-cleic acidin chicken cellsinfectedwithRoussarcoma
virus. J. Virol. 11:761-774.
13. Penman, S. 1966. RNA metabolism in the HeLa cell nucleus.J. Mol. Biol. 17:117.
14. Smith, G. H., and N.A. Wivel.1973.IntracytoplasmicA particles: mousemammarytumorvirus nucleoprotein cores? J. Virol. .11:575-591.
15. Tsuchida, N., S. Bhaduri, H. J. Raskas, and M. Green. 1973. Partial purification of intracellular murine
sar-coma-leukemiavirusRNA speciesby membrane filtra-tion.Intervirology1:27-33.
16. Tsuchida, N., M. S. Robin, and M. Green. 1972. Viral RNA subunits in cells transformed by RNAtumor
vi-ruses. Sciences 176:1418-1420.
17. Yang, S. S., and N. A. Wivel. 1973. Analysisof high-molecular-weight ribonucleic acid assocated with in-tracisternalAparticles. J.Virol.11:287-298. 18. Zhdanov, V.M., V. D. Soloviev, T. A.Bektemirov, K.
V. Ilyin. A.S. Bykovsky, N. P. Mazurenko. I. S. Irlin. and F. I. Yershov. 1973. Isolation of oncornaviruses. fromcontinuous human celllines.Intervirology 1:3-9.