Identification
and
Characterization
of
Intragenic Sequences
Which
Repress Human
Immunodeficiency
Virus
Structural
Gene
Expression
ALAN W. COCHRANE,' KATHRYN S. JONES,2 SARY BEIDAS,'t PATRICK J. DILLON,'
ANNA MARIE SKALKA,2AND CRAIG A.
ROSEN'*
Department ofMolecular Oncology & Virology, RocheInstituteofMolecular Biology, Roche Research Center,
340Kingsland Street, Nutley, NewJersey07110-1199,1 andInstituteforCancerResearch, Fox Chase Cancer Center, Philadelphia, Pennsylvania 191112
Received 15 April1991/Accepted 11July 1991
Examination of the lifecycle of the human immunodeficiency virus (HIV)hasshown that multiple levels of
regulation exist, includingsomewhich require the virus-encoded Rev protein.In the absenceof Rev,mRNAs
encoding thestructuralproteins remainuntranslated,aphenomenon whichappears, inpart, tobecausedby
nuclear entrapment of these RNA species. To examine the basis for repression of structural gene mRNA
expression,aheterologousassaysystemwasutilizedtodetermine whether regionspresentwithingagandpol
containelements capableofsuppressinggeneexpression whenpresentin cis. Bothgenes werefoundtocontain cis-acting repressor sequences (CRS) that block gene expression when present within the 3' untranslated portionofaheterologous genetranscript. Theelementwithinpolwasfoundtohave the strongestrepressive effect.While Rev alonewasunabletoreversetherepression observed with thepolsequence,addition oftheenv
Rev-responsive element(RRE) incisandRev in transdidcause reversal ofinhibition. Deletion mutagenesis
defineda260-bp element withinthe3' portionofpolthatcontainsapotentCRSwhich functions whenpresent
in thesenseorientation. Thecorresponding regioninHIV-2polwasfoundtocontainafunctionally similar CRS
element. Toexaminethemechanism ofrepression,theeffects oftheCRSelementsonboth the abundance and subcellular distribution ofthemRNAswereexamined. Neitherwasdramatically alteredwhenexaminedin the contextofaheterologousreporter (chloramphenicol acetyltransferase) mRNA. Theseresultssuggestthatthe CRS element defined within pol operates at a splicing-independent, posttranscriptional level to prevent
expressionofstructuralgenes.We suggest thatmRNA bearingtheseCRS elementscannot reactproductively with thetranslational machinery ofthecell.Theabilityof Rev,anuclearprotein,toreversethiseffectindicates that Rev must alterthe nuclearmetabolism of these mRNAssuch thatthe mRNAreachingthecytoplasmis capableofbeing translated.
Investigationof the humanimmunodeficiencyvirus(HIV) lifecycle has revealedacomplexpatternofgeneexpression.
In addition to elements common to other retroviruses, the
HIV genome encodes numerous regulatory proteins that
affect different facets of HIV replication and pathogenesis. At present, at least six nonstructural regulatory proteins which participate in the regulation ofgene expression or
replication havebeenidentified (29, 44). Ofthese
nonstruc-turalproteins,two,TatandRev, have beendemonstratedto
beessentialforreplication (7, 11, 20, 39, 41).Themechanism of action of Tat remains controversial, with both posttran-scriptionalandtranscriptionalmechanismsbeingconsidered (la, 17, 23, 27, 31, 33, 38). Studiesof the mechanism of Rev indicate that it operates at the posttranscriptional level to
permit synthesis oftheviralstructuralproteins (10, 39).How this isachieved isnottotallyclear. As with allretroviruses, expression of the full coding capacity of the viral genome requiresdifferential splicingofthefull-lengthgenomicRNA initially generated upontranscription. However, splicingof the genomic RNA, by necessity, must be an inefficient
process, sinceexpressionof thegagandpolgenesrequires
the transport and translation of the full-length genomic RNA. While with the simpleretroviruses, genomic RNA is
*Correspondingauthor.
t Present address: Mercer University School ofMedicine,
Ma-con,GA 31201.
transported from the nucleus tothe cytosol constitutively, with HIV, the genomic RNA and single spliced forms encoding env appear to be sequestered within the nucleus, whilethemultiplyspliced formsareefficientlytransportedto
thecytoplasmintheabsence ofRev (9, 15, 16, 25). Expres-sion of Rev results in the partial transport and subsequent translation of the sequestered mRNAs. Although it is well established that this release from sequestration requires binding of the Rev protein to its cis-acting target sequence
within env (26, 28), termed the Rev-responsive element
(RRE), the basis for the selective retention of the RNA encoding structural proteins is currently unclear. Work by Changand Sharp (2), usinga heterologous beta-globin sys-tem, has demonstrated that a mRNA can be rendered Rev responsive byboththe addition of RRE and the mutation of either the splicedonororacceptor site ofthe intron. These
results have ledtothehypothesisthatsequestration ofHIV
structural mRNA in thenucleusisduetotheinefficiency of the HIVsplicingreaction. However, the failuretoobservea
similar pattern ofregulation with thesimpleretroviruses,in
which a similar inefficiency in env mRNA splicing occurs
(12, 19, 40), suggests that either the basis for the splicing inefficiency of HIV must differ or that additional elements arepresent within the HIVgenomewhich contribute to this
novel form ofregulation.
In support ofthe latterhypothesis, studiesusing a heter-ologousassay systemtoexamineRev functionhave shown 5305
0022-538X/91/105305-09$02.00/0
Copyright © 1991, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
5306 COCHRANE ET AL.
that cis-acting repressive sequences (CRS) encoded in the envgene of HIV contribute to the inefficientexpression of this mRNA(34). In this system,theenvgenewasplaced3' of thetermination codonofareportergene(chloramphenicol acetyltransferase [CAT]), resultinginadramatic decreasein CAT expression, which could be reverseduponadditionof Revin trans. Deletionanalysisof theenvsequencesuggests that several regions within env contain negative regulatory elements, because their deletion resulted in an increase in gene expression in the absence of Rev. Furthermore, dele-tionof thesplicedonororacceptorsites in this construct had
noeffectonexpressionand the constructremained respon-siveto Rev. These observations, coupled with the demon-stration that expression of other HIV structural proteins displayedasimilardependencyonthepresence of theRRE and Rev (8, 15), suggest that functionally identical CRS elements might be present within thegag andpolgenes of HIV.
Toidentify potentialCRSelements within the gag andpol genes ofHIV, segmentsof thesegeneswerefused3' of the CATterminationcodon and theireffectsonCATexpression following transfection were examined. Our results indicate thatboth gag andpolcontainCRSelementsthatareableto repressgeneexpressionwhenpresentin cis.Repressionwas reversedbyaddition of the RRE in cis andexpressionof Rev in trans. Deletionanalysisof thepolgenedemonstrated the presence ofaninhibitory sequencewithina260-bp regionof theintegrase domain. RNAanalysis suggeststhatinhibition ofgene expression mediatedby this CRS element is alsoa
posttranscriptional effect.
MATERIALSANDMETHODS
Preparationofexpressionvectors. ToidentifyHIVgagand
pol sequences capable of inhibiting gene expression, an
approach similar to that employed to study the CRS ele-ments of the env gene was used (34). In brief, the SacI-EcoRI(gag andpol, nucleotides 227 to4193), SacI-EcoRV (gag, nucleotides 227 to 2524), EcoRV-EcoRI (pol, nucleo-tides 2524 to 4193), or KpnI-EcoRI (pol 490-bp fragment,
nucleotides 3703 to 4193) fragment of strain Hxbc-2 was
placed 3' of the termination codon of the bacterial CAT gene. Nucleotidepositions are numberedby thenumbering systemof Ratneret al. (32). Vectors utilized contained the small t antigen intron and early polyadenylation signal of simianvirus40 (SV40) andexpression wasdrivenbyeither
the HIVtype 1 (HIV-1) longterminalrepeat(LTR)orSV40
early promoter. The plasmid pSVAR is a derivative of
pIlIAR(34) and the only difference betweenthe two is the
replacementof the HIVLTRwith the SV40 originandearly promoter.
The3'mutants ofthepol490-bpsequenceweregenerated by using the site-directed mutagenesis protocol ofKunkel (22). The 5'deletionmutantsweregeneratedby polymerase
chainreactionamplification (37)of theregionof interest and
cloned 3' of the terminationcodonof the CATgene.
The vectors pBC12BI/HIN and pBC12BI/HIN+RRE
were preparedby isolation oftheBamHI-NdeIfragment of pCG6 (a bacterial HIV integrase [IN] expression clone) which contains, in addition to the IN codingsequence, the lambda PL promoter. This fragment was ligated into the pBC12eukaryotic expression vector3' of the rat
preproin-sulin intron. ThelambdaPLpromoter was theneliminated,
and the 5' end of HIV IN was repositioned immediately
downstreamfrom the insulin ATG initiation codon by oli-gonucleotide-directed mutagenesis (22), usinga 36-base
oli-gonucleotidethat contained the 15 nucleotides
immediately
upstream of the ATGcodon,theATGitself,and the first18 nucleotides of HIV IN. The RRE utilized for both
pBC12BI/
HIN+RRE andpSVCP490RRE corresponds tonucleotides 7204 to 7684 of Hxbc-2 (32) and was placed 3' of the
inhibitory sequence and 5' of thepolyadenylation
signal.
Transfections andanalysis of expression. For initialstudies,
vectorsdrivenbytheHIV-1 LTRwereintroduced into CHO zip tatIII cells (34), which stably express tatprotein, using
theDEAE-dextran calciumphosphatetransfection
protocol
(34). Subsequent experiments utilized COS cells whichwere transfected by the DEAE-dextran protocol (5). Cells were
harvested 48 h posttransfection, and CAT activity was determined as previously outlined (13). For quantitative
assessment of the extent ofchloramphenicol modification, thin-layer chromatogram plates were scanned by using a
Betagen imaging device and theimages generated were used todetermine percent conversion(chloramphenicolto
acety-lated forms). For the analysis of CAT RNA subcellular distribution, cells were cotransfected with pgTat, another Rev-responsive vector(25).
To examineexpression of HIVIN, COS cells transfected with theINexpression vectors were labelled with 300 ,uCi of [35S]methionine for 3 to 6 h 48 h posttransfection. Cellswere subsequently lysed in RIPA buffer (0.1% sodium dodecyl
sulfate [SDS], 1% Triton X-100, 1% sodium deoxycholate,
0.15NaCl, 0.01 M TrisHCI [pH 7.4], 1 mM EDTA, 0.25 mM phenylmethylsulfonyl fluoride) and immunoprecipitation
performed by using rabbit polyclonal antisera directed against bacterially produced HIV-1 IN. Antibody-antigen complexes were precipitated with Staphylococcus aureus protein A, the precipitated proteins were analyzed on SDS-polyacrylamide gels, and bands were visualized by autora-diography.
Subcellular fraction and RNA extraction. Harvested cells were washed twice with ice-cold phosphate-buffered saline and then fractionated into nuclear and cytosolic fractions (14). Cells were resuspended inlysis buffer (10 mM TrisHCl [pH 7.5], 5 mMMgCl2, 10 mM NaCl, 0.50% NonidetP-40),
vortexedfor S s, and then placed on ice for Smin. Thelysate was then centrifuged at 2,000 rpm in a Sorvall RT6000B centrifuge for 5min at 4°C. The supernatant, designated the cytoplasmic fraction, was collected and immediately added to3 volumes ofasolutioncontaining6 M guanidine thiocy-anate,37.5 mM sodium citrate(pH 7.0), 0.75%Sarkosyl, and 0.15M
P-mercaptoethanol.
Thepellet was suspended inlysisbuffer, and the nuclei were pelleted by a second
centrifuga-tion at 2,000 rpmfor5 minto remove residual cytoplasmic material. The supernatant was removed, and the pellet (designated the nuclear fraction) was resuspended in a solution containing 4 M guanidine thiocyanate, 25 mM sodium citrate (pH 7.0), 0.5% Sarkosyl, and 0.1 M 1-mer-captoethanol. Both fractionswere processed as previously described(3). Following the final precipitation, RNApellets were washed with 75% ethanol, allowed to dry, and then redissolved in diethylpyrocarbonate-treated water. RNA was quantitated by determining the A260 of the solution.
Subcellular distribution of RNA. ForSianalysis of Tat and CATmRNA,theBl-pgTat S/K and pGM4CAT AEco clones,
respectively, wereused. TheBl-pgTat S/K plasmid contains theSalI-KpnI fragmentof HIV-1 (nucleotides 5331 to 5893 of
Hxbc-2)and encompassesthefirst Tat exon and a portion of the env gene. The pGM4CAT AEco plasmid contains the
HindIII-EcoRI fragment of the CAT gene derived from
pSVCAT (13). For labelling, Bl-pgTat S/Kand pGM4CAT AEco were linearized with Sall and HindIII, respectively.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HIV-1 LTR CAT PoiyA
i-i
-*0
pillCATgag/pol HV- 1LTR. CAT 4 PoIyA +
+
HIV-1 LTR; CAT Pol2//// y
227 2524
':'AA _ ,
pillCAT pol iIV-1LTRCAT poI PoyA
2524 4193.
'AA
-pillAR !HIV-1LCAT PoIyA]
5331 9177
-AA 7204 -S46.
pillCATpolRRE'HIV-1 LTR CAT RRE'PoyA
2524
FIG. 1. Detection of inhibitory sequences in the gag and pol genes of HIV-1. To the left are the schematic diagrams of the expression vectors used to identify the presence ofCRS-likeelements within the gag andpolregions ofHIV-1. Numbers correspond to the nucleotide
positions ofthe 5' and 3' termini of the HIV-1 fragments present in the vectors according to the numbering system of Ratner et al. (32). To theright,the thin-layer chromatograms used to analyze CAT activities present in celllysatesfollowing transfection of the vectors into CHO
zip tatIll cells in the absence (-) or presence (+) of Rev. Quantitation of the thin-layer chromatograms shown revealed the percent conversions: pIlICAT without Rev, 32.3; pIlIlAR without Rev, 1.12; pIlIAR with Rev, 31.0; pIIICATgag/pol without Rev, 4.2;
pIIICATgag/pol with Rev, 5.8;pIIICATgagwithout Rev, 4.29;pIlICATgagwith Rev,7.17;pIIICATpolwithoutRev, 0.8;pIlICATpol with Rev, 1.44;pIIICATpolRRE without Rev, 0.8;pIIICATpoIRREwith Rev, 35.3.
Thelinear DNA was then labelled by incubation in a solution
containing 50 mM Tris HCl (pH 8.3), 40 mM KCl, 6 mM MgCl2, 1 mM dithiothreitol, 200 ,uCi ofat-32P-labelled
deoxy-nucleoside triphosphates (50 pLCi [each] of dATP, dTTP, dGTP, and dCTP), and 15 U of avian myeloblastosis virus reversetranscriptase at37°Cfor 30 min. The reactions were terminated byaddition of EDTA to 20 mM, and unincorpo-rated label was removed by passage of samples through
Sephadex G-50 spincolumns. The eluant was ethanol
pre-cipitated andresuspended in restriction enzyme buffer, and
plasmids Bl-pgTat S/K andpGM4CAT AEco were digested with PvuII and NarI, respectively. The desired fragments were isolated from 5% polyacrylamide gels, denatured by
heatingat 100°C for5 min, and used for S1 analysis. DNA
probes were added to 10
[Lg
of RNA, and the mixturewasethanol precipitated and dissolved in 20
[lI
of a solution containing 80% formamide, 40 mM piperazine-N,N'-bis(2-ethanesulfonicacid) (PIPES) (pH 6.4),400mMNaCl,and 1 mMNa2EDTA. Sampleswereheatedat70°C for 10 min and thenplacedinaH20bath setatthepredetermined annealingtemperature(49°Cfor Tat mRNA and42°Cfor CATmRNA).
Followingincubation for 4 to 6h, 200,ulof S1 buffer (30mM
sodium acetate [pH 4.6], 250 mM NaCl, 1 mM ZnCl2) containing 300 to 600 U of S1 nuclease was added. For
analysis of Tat mRNA, the S1 reaction mixtures were
incubated at 37°C for 1 h. For
analysis
of CATmRNA,
samples were incubated at room temperature for 1 h. S1 reactions were terminated by addition of 50[lI
of S1 stop buffer (4 M ammonium acetate, 20 mM EDTA, 40 ,ug of tRNA per ml), and the samples were ethanolprecipitated.
Pelleted DNA was resuspended in 70% formamide-5 mM Tris HCI (pH8.0)-0.5mMEDTA,heatedat100°Cfor 5min,
and then run on a 5% denaturing
polyacrylamide
gel. Fol-lowingelectrophoresis, gelsweredried and used forautora-diography.
Determination of HIV IN RNA distribution was carried
out by Northern (RNA) blot analysis (43). Aliquots (10 ,ug each) of nuclear andcytoplasmic RNAfrom control cells or cells transfected with pBC12BI/HIN+RRE in the presence orabsenceof Revwerefractionatedon1.2%
formaldehyde-agarosegels, andfollowing electrophoresis, the RNAs were transferred to nitrocellulose. The blots were baked at 80°C for 90 min and prehybridized in a solution containing 50% formamide, 6x SSC (lx SSC is 0.15 NaCl plus 0.015 M
sodium citrate) 1% SDS, 0.1% Tween 20, and 100 ,ug of
tRNA perml. Prehybridizationwascarriedoutat55°C for2 h, and then hybridization wasperformed in the same buffer at55°C overnight. The probe used for detection ofIN RNA was an antisense RNAprobe corresponding to nucleotides
3703to4193of Hxbc-2 (32). The blotswerewashed twice in 1x SSC-0.1% SDS at room temperature and twice in 1x
SSC-0.1%SDSat65°C.Toremoveresidualhybridizationto 28S rRNA, blots were washed three times in 2x SSC at roomtemperature, oncewith 2x SSCplus 1 ,ugofRNase A permlfor30min atroomtemperature, and thenonce with 0.1x SSC-0.1% SDS at 50°C. Signals were detected by autoradiography.
RESULTS
Identification of CRS-like elements in thegagpolregionof HIV-1. Having
successfully
used a heterologous assay toexamine the CRS present within the env region of HIV-1 (34), a similar strategy was utilized to
identify
functionally
identical sequences within othercodingregionsof the HIV-1 genome. gag,pol, or gagpol
regions
ofHIV-1 were fused downstream of the termination codon of the CAT gene and the effect of the fusion on CATexpression
was assessedfollowingtransfection
(Fig.
1).Analysis
of the CAT expres-sion levels (Fig. 1) revealedthataddition of the gag and gag polregions resulted inanapproximately
sevenfold decreasein the levelof CAT
expression.
However, additionof thepolpill CAT
227
pIlCAT gag
S.
*
.
1- 0
on November 10, 2019 by guest
http://jvi.asm.org/
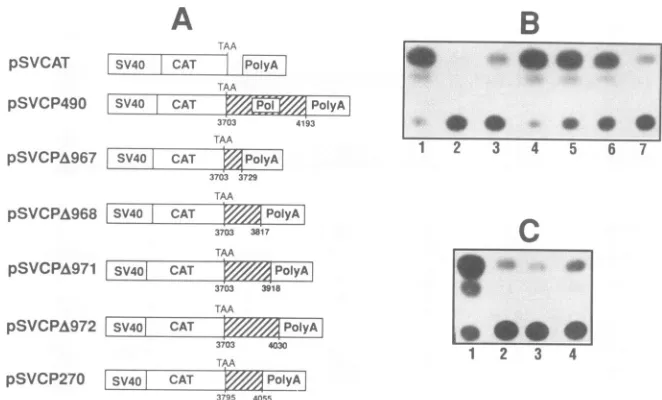
[image:3.612.151.469.81.279.2]5308 COCHRANE ET AL.
A
TAA
pSVCAT SV40 CAT rPoIYA 1
-___
_"AA
pSVCP490 SV40 I CAT iPo1yA
3703 93
rAA
pSVCPA967 SV40 CAT Pol
3703 3729
IAA
PSVCP,&968 USV401 CAT YA
3793 3817 TAA
pSVCPA971 [j0o CAT PoPolyA
3703 3918
TAA
pSVCPA972 isv40o =AT Poly
3703 4030
B
1 .3 4 5
1 2 3 4 5 6 7
C
_
.*
* *
t 2 3 4
TAA
pSVCP270
LSV4o
CAT3795 4055
FIG. 2. Deletionmapping of thepolCRSelement. TomaptheCRSelementwithin thepolfragmentutilized inFig. 1,subregionsofthis fragmentweretestedforinhibitoryactivity. (A) Schematic diagramofconstructsusedtodefinetheCRSelement withinpol.Numbersshown indicatethe 5' and 3' boundaries of the fragmentstested andarenumberedbythenumbering systemof Ratner et al.(32). (B)CATactivities generatedfollowing transfection of 3' deletionconstructsinto COS cells. Lanes:1,pSVCAT, 91.9%; 2,pSVAR,7.1%; 3,pSVCP490,21.7%; 4, pSVCPA967, 91.5%; 5, pSVCPA968, 75.8%; 6, pSVCPA971, 56.3%; 7, pSVCPA972, 15.4%. (C) CAT activities generated following transfection of 3' and 5' deletionconstructs into COS cells. Lanes: 1, pSVCAT, 83.4%; 2,pSVCP490, 11.8%; 3, pSVCPA972, 8.0%; 4, pSVCP270, 13.9%. Percentages indicated refertopercentconversionofchloramphenicoltooneof theacetylatedforms.
region alone effected an approximately 30-fold decrease in the level of expression. The failure to observe a similar
30-folddecrease with thepIIICATgag/polconstructsuggests that positional effects may influence the expression of the
inhibitory elements and that the CRSelements within both
gag and pol probably act independently of one another. Alternatively, the presence ofa major splice donor within
the gag fragment used in these experiments may have permitted excision of the repressive sequences by using a
downstream cryptic splice acceptor, thus permitting CAT expression. While addition of thepol region was found to
elicitthesamedecreaseasthatpreviouslyobserved withenv
(pIIIAR), in the case ofpIIICATpol, expression of Rev in
trans did not rescue gene expression. This observation suggested that either the mechanism ofrepression effected
bypolwas different from that ofenv or that the construct
lackedtheelement requiredtomediatetheeffect ofRev.To
test the latter hypothesis, the RRE within env was added ontothepIIICATpolconstruct(generatingpIIICATpolRRE) and theeffectof Revexpressionwasexamined. Additionof
Revin trans, andthe RREin cisresultedinrescue of CAT expression to levels observed with pIIICAT (Fig. 1). No
rescueofexpression wasevident uponaddition of theRRE
alone.Together these data suggestthat the CRS element(s) identified withinpol are at least functionally equivalent to those previously characterized in env.
Deletion mapping of CRS elements within pol. Deletion analysis was subsequently used to identify a minimal CRS
element in pol. To facilitate subsequent RNA analysis, experimentswereperformed inCOS cells, necessitating the
use of SV40 promoter-based vectors. A 490-bp region, corresponding to nucleotides 3703 to 4193, was found to significantly repress CAT expression when placed in cis (Fig. 2). Whilethe490-bpsequenceisinhibitory, itsextentof inhibition isnotasgreatasthatobserved usingthefull1.7-kb
pol region. This observation suggests that additional
se-quences within the 1.7-kb pol region may operate in
con-junction with the CRS element in the 490-bp fragment to effect the full inhibitory capacity of the region (indeed, a second inhibitory sequence has beententatively mapped to the deleted region in subsequent experiments;Cochraneand Rosen [4a]). Subsequent work focused on the 490-bp
frag-ment. By introducing further deletions,we found that (Fig.
2)greatestinhibitionwasobserved withaconstruct contain-ing nucleotides 3703 to 4030 (pSVCPA972), with further 3' deletions resulting in a significant increase in CAT expres-sion. Having established a 3' boundary for the repressive element,asimilarstrategywasemployedtocharacterizethe 5' boundary. The conclusion of these studieswasthe dem-onstration that a 260-bp sequence, comprising nucleotides 3795to4055, wascapable ofinhibiting CAT expressiontoa
similar extent as that observed with the 490-bp fragment
(pSVCP270; Fig. 2).
While the repression observed with pSVCP490 was not
relievedby expression of Rev intransalone, addition of both the RRE in cis and Rev intranselicitedamarked increasein
CAT expression (Fig. 3). These observations indicate that the490-bp regiondoes containabona fideCRS-like element.
The repressive effect of this sequence was found to be orientation dependent; inversion of the 490-bp sequence
resulted in an increase in CAT expression. The orientation
dependence of the sequence and its location within the construct strongly suggest that its effect ismediated at the posttranscriptional level.
The presence ofa CRS element within one of the
struc-turalgenesofHIV-1suggestedthatasimilarelementmaybe present within homologous regions of related viruses. To
test this hypothesis, regions of HIV-2 (Rod) and simian immunodeficiency virus (SIV) with extensivesequence
sim-ilaritytothe490-bp sequencedefined abovewere cloned 3'
ofthe termination codon of CAT and their effects on CAT
expression following transfection were examined. The
se-quence from HIV-2 inhibited CAT expression (pSVCAT
Rod490)toasimilarextentasthatobservedwiththe490-bp
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
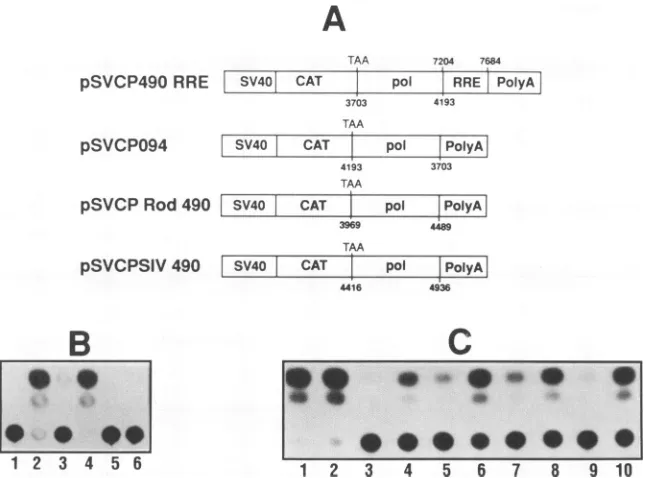
[image:4.612.139.470.79.279.2]A
pSVCP490 RRE
pSVCP094
pSVCPRod 490
pSVCPSIV490
TAA 7204 7684 SV40 CAT | pol tRRE PoyA
3703 4193
TAA
SV40 CAT pol PolyA
4193 3703 TAA
SV40 CAT 1 pol JPolyA|
3969 4489
TAA
SV40 CAT pol |PolyA
4416 4936
B
1 9 A r R
C
IL 123 4 5 7* 8910
1-
-o
o o o_ ..0... 0I ' a Lf D 0 1 2 3 4 5 6 7 8 9 10
FIG. 3. Effect ofRevexpressionandsequence polarity on the inhibition mediated by thepol490-bp sequence of HIV-1 and examination
of its homologs inHIV-2 andSIV.(A)Schematic diagram of constructs used to assess the capacity of Rev to reverse the inhibitory effect of
thepol 490-bp sequence and the dependency of the inhibitory effect on thepolarityof the sequence. The RRE used in theindicated constructs
corresponds to theregion ofenv encompassed by nucleotides 7204 to 7684 (32). For constructs pSVCPRod490 and pSVCPSIV490, the
indicated nucleotidesdelineate the regions of HIV-2 (Rod) or SIV found to have extensive homology to the HIV-1pol490-bp sequence
(numbering ofRod andSIVregions according to GenBank sequence). (B) Effect of Rev expression in trans on the inhibition mediated by the
pol490-bpsequence. Lanes: 1, untransfected cells, 0%; 2, pSVCAT, 85.1%; 3, pSVAR without Rev, 11.5%; 4, pSVAR with Rev, 92.3%; 5,
pSVCP490 withoutRev,1.61%;6,pSVCP490 with Rev, 4.57%. (C) Effect of pol 490-bp sequence inversion and addition of RRE. Lanes: 1,
pSVCAT withoutRev,96.5%;2,pSVCATwithRev, 95.0%; 3, pSVAR without Rev, 2.8%; 4, pSVAR with Rev, 23.0%; 5,pSVCP490RRE without Rev, 10.5%; 6, pSVCP49ORRE with Rev, 54.2%; 7, pSVCP490 without Rev, 15.7%; 8, pSVCP094 without Rev, 35.8%; 9, pSVCPRod490 without Rev, 4.6%; 10, pSVCPSIV490 without Rev, 45.1%. Percentages shown indicate the percent conversion of
chloramphenicolto one of the acetylated forms.
HIV-1 element. In contrast, the homologous region of SIV
wassubstantially less efficient in repressinggeneexpression
(Fig. 3), with only a maximum twofold repression in CAT
expression observed.
Demonstration of the inhibitory effect ofthepolsequences
using an INexpression system. Theuseof the CAT
expres-sion systemto testforthepresenceof CRS elements, while sensitive, suffers from the caveat that results obtained in sucha heterologous system may notreflectwhat occurs in virus infection. To address this possibility, a second assay
system was employed to verify the repressive effect ofthe pol region. The 490-bpsequencedefined aboveencompasses alargeportionof the HIV-1 IN domain.Therefore, onemay
anticipate that IN expression would display a Rev
depen-dencysimilartothatobserved forpSVCP49ORRE. By using thepBC12vectorsystem(5), the regionofpol corresponding
totheINgenewascloned 3'oftheratpreproinsulinIIintron andINexpressionwasexamined after transfection into COS
cells. No IN expression was detectable after transfection with either pBC12BI/HIN or pBC12BI/HIN+RRE in the
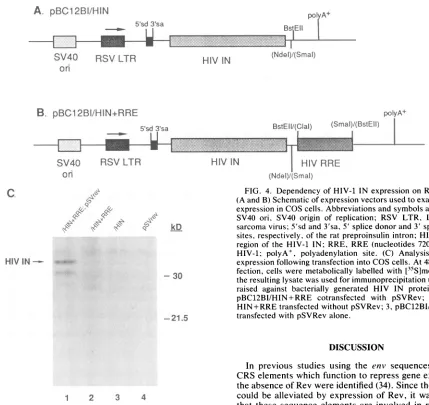
absence of Rev (Fig. 4). However, a significant level of IN
expressionwasdetected from thepBC12BI/HIN+RRE con-structwhen Rev was expressed in trans. The second band observed in cells transfected with plasmids pBC12BI/ HIN+RREand pSVRev would appeartobe a degradation
product, given its absence in any of the other lanes. This
observation supports the previous findings using the CAT
assay system andconfirms that this domain ofpolcontains
repressive elements whoseaction can be reversedby addi-tion ofthe RRE and Revin cis and trans, respectively.
Examination of the mechanism of pol 490-bp sequence inhibition. In light ofprevious studies which indicate that inhibition of viral structural gene expression is due to the
sequestrationof thecorrespondingmRNAsinthe nucleus of thecell(9,15,16,25),itwasofinteresttodeterminewhether a similar mechanism applied to the inhibitory effect of the
CRSelements definedabove. Accordingly, cellstransfected
with plasmidscontainingthepol490-bp CRS elementwere
fractionated into nuclear andcytosolic componentsand the distribution of expressed mRNA was determined. Analysis
of CAT mRNA generated from plasmids pSVCP490 and
pSVCP49ORRE
(Fig. 5)revealed that incontrast toprevious
studies (9, 15, 16, 25), the inhibition of CAT
expression
effectedby thepol490-bp sequence could notbeexplained
by sequestering of mRNA containing the sequence in thenucleus ofthe cell, despite the marked reduction in CAT
expression. AsignificantamountofCAT-containingmRNA was detected in the cytoplasm. In addition, the inhibition observed couldnotbeattributedsolelytoalterations in CAT RNA steady-state levels, because the abundance of RNA
generatedfrom thepSVCP490vector wascloseto that from
pSVCAT. Furthermore, rescueof CAT
expression,
effectedby addition of the RREwith Rev in trans, wasnotfound to
be associated with a dramatic alteration in CAT RNA
subcellular
distribution,
although
some increase in CATmRNAabundancewasnoted. To address the
possibility
thaton November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.148.471.83.322.2]5310 COCHRANE ET AL.
A. pBC12BI/HIN
5SV40 RSV LTR
on
sd 3'sa II
HIV IN
B.
pBC12BI/HIN+RRE5'sd 3'sa
SV40
RSV
LTRon
polyAs BstEll/(CIal) (Srmal)/(BstEll)
HIV IN
...:.
rS:'alwEma:
HIV RRE (Ndel)/(Smal)
FIG. 4. Dependency of HIV-1 INexpression onRREand Rev.
(A and B)SchematicofexpressionvectorsusedtoexamineHIV IN
expressioninCOScells.Abbreviationsandsymbolsareasfollows:
SV40 ori, SV40 origin of replication; RSV LTR, LTR of Rous sarcomavirus;5'sdand3'sa,5' splice donor and 3' spliceacceptor
sites, respectively,of the ratpreproinsulin intron; HIVIN, coding
region of the HIV-1 IN; RRE, RRE (nucleotides 7204to7684) of HIV-1; polyA+, polyadenylation site. (C) Analysis of HIV IN expressionfollowing transfection into COScells. At 48h
posttrans-fection,cells weremetabolically labelled with [35S]methionine and
theresultinglysatewasusedforimmunoprecipitation using antisera raised against bacterially generated HIV IN protein. Lanes: 1, pBC12BI/HIN+RRE cotransfected with pSVRev; 2, pBC12BI/
HIN+RREtransfectedwithoutpSVRev; 3,pBC12BI/HIN;4, cells
transfected withpSVRev alone.
DISCUSSION
1 2 3 4
thepresenceof RNA in the cytosol wasduetoleakage from
the nucleus during fractionation, cells were cotransfected
withthepgTatvector(25). The pgTatvectorconsists ofthe
twoTatexons separated by an intron corresponding to the
env gene of HIV-1. Previous work with this vector has established that in the absence ofRev, the spliced form of the RNAis transportedtothe cytoplasm while theunspliced form remains entrapped in the nucleus. Analysis of the distribution ofthe unspliced form ofpgTat RNA withinthe fractions used to examine the distribution of pSVCP490-generated RNA (Fig. 5) revealed that unspliced RNA was
detectable only in the nuclear fractions. Thus, results with pSVCP490cannotbe attributed toleakage of material from thenucleus into the cytoplasm.
In contrast to our findings with the CAT-based vectors,
analysis of the subcellular distribution of pBC12BI/HIN+ RRE-generated RNA (Fig. 5) gave results similar to those previously reported (9, 15,16, 25). Inthe absence of Rev, IN mRNA was detectable only in the nuclear fraction of the
cell.Addition ofRevresultsin theappearanceof IN mRNA
in thecytosol.
In previous studies using the env sequences of HIV-1, CRS elements which functionto repressgene expressionin
the absence of Revwereidentified(34). Since therepression could be alleviated by expression ofRev, it was suggested
that these sequence elements are involved in regulation of HIV structuralgeneexpression. Other studies (8, 15), dem-onstratingasimilardependency uponRev forexpression of the gag gene products, suggested that CRS elements are
likely present within other structural genes of HIV. In this study, wehaveverified this hypothesis usingaheterologous
assaysystem. Weobserved that addition of either thegag or
pol sequences in cis resulted in reduction of CAT gene
expression. Sequences present within pol were found to
elicit the greatest inhibitory affect. To delineate the CRS elements, subregionsof thepolgene wereexaminedforthe
presenceofinhibitorysequences. Initialexperiments
identi-fieda 490-bpsegment thatcontained a CRSelement which wasfurthermapped to a260-bp sequence correspondingto nucleotides 3795 to 4055 of strain Hxbc-2. The deletion mapping demonstrates that the splice acceptor within this region is not required for the repressive capacity of the
sequence, since equivalent inhibition of CAT expression is
observed with mutant pSVCPt\972, which lacksthe known splice acceptor. Thiswould suggest that splicing signals do not contribute to the inhibitory effects of this region. Our datadonotruleoutthepossibilitythat theCRS elementsare
capable ofinhibiting splicing events which occur in other
regions of the vector. However, experiments designed to
examine thispossibilityhavefailedtodetect suchanactivity
(data not shown). Furthermore, deletion of the intron from
poIA+
BsiEll
(Ndel)/(Smal)
C.
kD
HIV
IN-- 30
-21.5
[image:6.612.79.508.78.483.2]:: I I
...
I
X..:
... ?x
M.
W-M
::::
....
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
pSVCAT
I
-'--CP490
+neo +rev +neo
N
C
N
C
N
C
e No
S_
cmO
C o
0.O
CO- - + -
re
C
pSVCAT pSVCP49ORRE
r--
+I _ 1 [
F--REV: N C N C N C N C
_ *
p
a
pSVCAT pSVCP49ORRE +
thepBC12BI/HIN+RREconstructhadnoeffecton
expres-sion orRev responsiveness (datanot shown). Thefunction of theCRS elementis orientationdependent,because inver-sion ofthe sequence alleviated a portion of the repressive
effect. Since the HIV-2 virus isvery similartoHIV-1 in its
d
b
pSVCAT
pSVcp49D
+neo
+Rev
+neoI
I
111
N
C
N
C
N
C
M
HIN
ARRE
Rev +
.. I
Nuclei
-
probe
- unspliced
-
spliced
M HIN
RRE
FI771
Cytosol
FIG. 5. Analysis of subcellulardistribution of RNAs containing
pol CRS element. (a) (Topgel) Aliquots of RNA from nuclear (N)
andcytoplasmic (C) fractions of cells transfected with pgTat
to-gether with pSVCAT or pSVCP490 and pSVNeo (+neo) or Bl-SVRev(+rev). TheseRNAs were hybridized with a CAT-specific
probeand treated with S1,and samples were run on adenaturing polyacrylamide gel. (Bottom gel) CATassays ofcells utilized for RNAanalysis. (b) Aliquots ofRNAtreated as described above were
hybridized with aprobefor spliced andunspliced Tat RNA. The
three bands visible following S1 treatment represent reannealed
probe, the unspliced form ofTatmRNA,and thesplicedform of Tat mRNA. AfterS1treatment, sampleswereanalyzed ondenaturing polyacrylamide gels.(c)(Topgel) Aliquotsof RNA from nuclear (N) andcytoplasmic(C) fractions of untransfectedcells or cells
trans-fected with pSVCAT or pSVCP49ORRE in the presence (+) or
absence(-) ofRev were used forhybridization withCAT probe. (Bottomgel) Corresponding CATassaysfrom cells used forRNA
analysis. (d) Analysis of subcellular distribution ofHIV IN mRNA.
Following fractionation of cells into nuclear (nuclei)orcytoplasmic (cytosol) fractions, aliquots ofRNA were analyzed by Northern blotting. Lanes: M, mock-transfected cells; HIN+RRE, pBC12BI/
HIN+RRE transfected cells inthepresence(+)orabsence(-)of
Rev.
regulationof geneexpression,weanticipatedthepresenceof
functionally equivalent CRS elements within the
corre-sponding regionof the HIV-2polgene. Ouridentification of ahomologous sequenceinHIV-2,withinhibitory properties
similartothoseof the element inHIV-1,suggeststhat these
a
a
I-A 5* a
REV:
ireAM
q
*
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.328.551.89.461.2]5312 COCHRANE ET AL.
elementsare likely important in regulation of virus structural gene expression. As observed with the entire
pol
sequence,
reversal of the inhibition with the 490-bp element could be achieved by addition of the RRE in cis together with
expressionof Rev in trans. This ability of Rev to reverse the
inhibition imparted by these sequences indicates that the
repressive element within pol is at least functionally related tothose previously characterized in env (34).
Upon analysis of the CRS-containing CAT mRNA, no gross alteration in abundance or subcellular distribution was
observedthat could account for the lack of gene expression. However, analysis of RNA generated from the IN expres-sion vectors was consistent with previous observations and
suggested that RNA containing a CRS element can be
entrappedin the nucleus in the absence of Rev. The basis for the difference between the two expression systems is un-clear. There is some support for the notion that sequence
context can affect the function of particular sequences. The iron-responsive element can affect mRNA translation or
stability, depending on the mRNA species analyzed (42). Theability to replicate previous findings in our system using the pgTat vector (25) as well as the IN expression vector
indicates that methodological problems such as gross nu-clear leakage of RNA did not occur during fractionation.
Therefore, our inability to detect similar changes in subcel-lular distribution with the heterologous CP490 transcripts is noteasily explained. One possibility is that because of some
peculiarity of sequence context in the
pSVCP490
transfec-tants, a significant amount of CAT mRNA is transported to the cytoplasm through a nonproductive route. In this case, Rev might still effect transport but we would not be able todiscriminate between RNA entering the cytosol through a
nonproductive route and the Rev-mediated exported RNA which is eventually translated. Regardless of the specific
mechanism, observations made with the heterologous sys-tem clearly showed that the requirement for Rev in gene
expressionis not solely in RNA transport from the nucleus but that Rev is capable of affecting the utilization of target mRNA by the translational machinery within the cytoplasm. A similar observation has been made by Ariggo et al. (1).
Analyzing the subcellular distribution of env mRNA, these
investigatorsobserved that RNA is present in the cytoplasm inthe absence and presence of Rev but is found associated with polysomes only after Rev expression. The ability of Rev toalter the translational properties of target mRNAs has alsobeenobserved in a similar heterologous system utilizing
CRS elements from the HIV-1 env gene (23a). The observa-tionthat Rev can affect the translational capacity of its target mRNA bears striking similarity to the effects of Tat on
TAR-containing mRNA, as demonstrated in the Xenopus
oocyte system (la). In this system, it was observed that
injected mRNA containing the TAR element is translated only when RNA is injected into the nucleus in the presence ofTat. The requirement for Tat and nuclear
injection
sug-gests that Tat interaction with the RNA results in the mRNA being translationally competent upon transport into thecytoplasm. The close similarity of the effects of Tat in the Xenopus oocyte system and those of Rev on the reporter system used here are of particular interest, because both
proteins demonstrate strong accumulation in the nucleoli of
transfected cells (21, 24, 30, 36) and both are RNA-binding
proteins (4, 6, 18, 26, 35, 45), which have the potential of
modifyingthe interaction of other cellular factors with their
respective target mRNA.
Results from other Rev-dependent expression systems have suggested that nuclear entrapment of HIV structural
mRNA is the
by-product
of an inefficient splicing process.Results obtained
by Chang
andSharp
(2),
demonstratingthatmutation of either
thebeta-globin
splice
donor or acceptorsequence
in aheterologous
expression
vectorelicits nuclearentrapment
ofmRNA,
which can then be efficientlyex-ported
to thecytoplasm
by placement
ofthe HIVRRE incisand
expression
ofRev
intrans,
lendsfurther support to thishypothesis.
Theability
of theCRS
elements, whichappar-ently
lack functional
splicing
signals,
to elicit the sameoutcome
as that obtained with inefficient splicing does notnecessarily
conflict
withthis
interpretation.
We speculatethat cellular factors which
interact with the HIV splicingmachinery may
also interact withcryptic
elements presentwithin the CRS
sequences.
Insupport
of this hypothesis, wehave found
thatspecific
cellular factors interact with theHIV-1
and HIV-2 CRS elements and that monoclonalanti-bodies directed
against
specific
splicing
components caninterfere with
complex
formation(unpublished
data).Fur-ther studies on
the nature of these interactionsand their rolein nuclear
entrapment
of HIV
RNA will likely add to ourunderstanding
of thoseprocesses
that controlmRNA metab-olism.ACKNOWLEDGMENTS
We thank Tina Rose for
preparation
of this manuscript andJ.
Kulkosky
forproviding
antibodies to HIV IN. Alan Cochraneis therecipient
of anAmFAR/F.L.
andE.L.
Cummings ScholarAward.This work was
supported
inpart
by a National Drug DiscoveryAward(C.A.R.),
by
National Institutes of Health grant CA49042,CA-06927, andRR-05539, a
grant
from the Pew Charitable Trust,and also
by
anappropriation
from the Commonwealth of Pennsyl-vania (A.M.S.).REFERENCES
1.
Arrigo,
S.J.,
andI.
S.Y.
Chen. 1991. Rev is necessary fortranslation but not
cytoplasmic
accumulationof HIV-1vif, vpr,and
envlvpu
2 RNAs. Genes Dev. 5:808-819.la.Braddock,
M.,
A. M.Thorburn,
A. Chambers, G. D. Elliot,G.J.
Anderson,
A. J.Kingsman,
and S. M. Kingsman. 1990. Anuclear translational block
imposed
by
the HIV-1 U3 regionisrelieved
by
the tat-TAR interaction. Cell 62:1123-1133.2.
Chang,
D.D.,
and P. A.Sharp.
1989. Regulation by HIV Revdepends upon
recognition
ofsplice
sites. Cell59:789-795.3.
Chomczynski,
P.,
and N. Sacchi. 1987. Single-step method ofRNA isolation
by
acidguanidinium
thiocyanate-phenol-chloro-formextraction. Anal. Biochem. 162:156-159.
4.
Cochrane,
A.W.,
C.-H.Chen,
and
C. Rosen. 1990. Specificinteraction of the HIV Rev transactivator proteinwith a struc-tured
region
in the env mRNA. Proc. Natl. Acad. Sci. USA87:1198-1201.
4a.Cochrane,
A.W.,
andC. Rosen.Unpublished
data.5.
Cullen,
B. R. 1988. Use of
eukaryotic
expression
technology inthe
functional
analysis
of cloned
genes.
Methods
Enzymol.152:684-704.
6.
Daly,
T.,
K.Cook,
G.Gray,
T.Maione,
and J. Rusche. 1989.Specific binding
of HIV-1 recombinant Rev protein to theRev-responsive
element invitro.
Nature(London)342:816-819.7.
Dayton,
A.I.,
J.
G.Sodroski,
C.A.
Rosen, W. C. Goh, andW. A.
Haseltine.
1986. The transactivator gene ofthe humanT-cell
lymphotrophic
virustype
III is required for replication.Cell 44:941-947.
8.
Dayton,
A.I.,
E. F.Terwilliger, J.
Potz, M. Kowalski, J. G.Sodroski,
and W. A. Haseltine. 1988. Cis-acting sequencesresponsive
to the revgene
product
of the humanimmunodefi-ciency
virus. J.Acquired
Immune Defic. Syndr. 1:441-452.9.
Emerman, M.,
R.Vazeux,
and K. Peden. 1989. The rev geneproduct
ofthe humanimmunodeficiency
virusaffectsenvelope-specific
RNAlocalization. Cell 57:1155-1165.10.
Feinberg,
M.B.,
R. F.Jarrett,
A.Aldovini,
R. C. Gallo,andF.Wong-Staal.
1986.HTLV-III
expression
and productioninvolveJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
complex regulation at the levels of splicing and translation of viral RNA. Cell 46:807-817.
11. Fisher, A. G., M. B. Feinberg, S. F. Josephs, M. E. Harper, L.M.Marselle, G. Reyes, M. A. Gonda, A.Aldovini, C. Debouk, R. C. Gallo,and F. Wong-Staal. 1986. The transactivator gene of HTLV-III is essential for virus replication. Nature (London) 320:367-371.
12. Fu, X.-D., R. A. Katz, A. M. Skalka, and T. Maniatis. 1991. The role of branch point and 3' exon sequences in the control of
balanced splicing of avianretrovirus RNA. Genes Dev. 5:211-220.
13. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express chloramphenicol
acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 14. Greenberg, M., and E. Ziff. 1984. Stimulation of 3T3 cells
inducestranscription of the c-fos proto-oncogene. Nature
(Lon-don) 311:433-438.
15. Hadzopoulou-Cladaras, M., B. K. Felber, C. Cladaras, A. Atha-nassopoulos, A. Tse, and G. N.Pavlakis. 1989. The rev(trslart) protein ofhuman immunodeficiency virus type 1 affects viral mRNAand protein expression via a cis-acting sequence in the envregion. J.Virol. 63:1265-1274.
16. Hammarskjold, M.-L., J. Heimer, B. Hammarskjold, I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human
immunodeficiencyvirus env expression by the rev gene product. J.Virol. 63:1959-1966.
17. Hauber, J., A. Perkins, E. P. Heimer, and B. R. Cullen. 1987.
Trans-activationof humanimmunodeficiencyvirus gene
expres-sion ismediatedby nuclear events. Proc. Natl. Acad. Sci. USA
84:6364-6368.
18. Heaphy, S., C. Dingwall,I.Ernberg, M. J. Gait, S. M.Green, J. Karn, A. D. Lowe, M. Singh, and M. A.Skinner. 1990. HIV-1
regulator ofvirion expression (Rev) protein binds to a RNA
stem-loop structure located withinthe Rev response element.
Cell 60:685-693.
19. Katz, R. A., and A. M. Skalka. 1990.Controlof retroviral RNA
splicing through maintenance of suboptimal processingsignals.
Mol. Cell. Biol. 10:696-704.
20. Knight, D. M., F. A. Flomerfelt, and J. Ghrayeb. 1987.
Expres-sion of the art/trsprotein of HIV and study of its role in viral envelope synthesis. Science 236:837-840.
21. Kubota, S., H. Siomi, T. Satoh, S. Endo, M. Maki, and M. Hatanaka. 1989. Functional similarity of HIV-1 Rev and
HTLV-1 Rexproteins: identification ofa new nucleolar-target-ing signal in Rev protein. Biochem. Biophys. Res. Commun.
162:963-970.
22. Kunkel, T. A. 1985.Rapidandefficientsite-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
23. Laspia, M., A. Rice, and M. Mathews. 1989. HIV-1tatprotein increases transcriptional initiation and stabilizes elongation. Cell59:283-292.
23a.Lawrence,J., etal. Submitted forpublication.
24. Malim, M. H., S. Bohnlein, J. Hauber, and B. R. Cullen. 1989.
Functionaldissection ofthe HIV-1rev trans-activator-deriva-tion ofatrans-dominantrepressorofrevfunction.Cell 58:205-214.
25. Malim, M. H., J. Hauber, S.-Y. Le, J. V. Maizel, and B. R. Cullen. 1989. The HIV-1 rev transactivator acts through a
structured target sequence to activate nuclear export of
un-spliced viral mRNA. Nature(London)338:254-257.
26. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J.
Hauber,and B. R.Cullen. 1990. HIV-1 structural gene
expres-sion requires binding ofthe Rev trans-activator to its RNA
targetsequence. Cell 60:675-683.
27. Muesing, M. A., D. H. Smith, and D. J. Capon. 1987.Regulation of mRNA accumulation by a human immunodeficiency virus trans-activator protein. Cell 48:691-701.
28. Olsen, H., P. Nelbock, A. Cochrane, and C. Rosen. 1990.
Secondarystructureisthemajor determinantforinteractionof HIV Revprotein withRNA. Science247:845-848.
29. Pavlakis,G. N., and B. K. Felber. 1990. Regulation of expres-sionofhumanimmunodeficiency virus.The New Biol. 2:20-31. 30. Perkins, A., A. W. Cochrane,S. M. Ruben, and C. A. Rosen.
1989. Structural and functional characterization of the human
immunodeficiencyvirusrevprotein.J.Acquired Immune Defic.
Syndr.2:256-263.
31. Peterlin, B. M., P. A. Luciw, P. J. Barr, and M. D. Walker. 1986. Elevated levels of mRNA canaccountfor the transacti-vation ofhumanimmunodeficiencyvirus. Proc. Natl. Acad.Sci. USA83:9734-9738.
32. Ratner, L., W. Haseltine, R. Patarca, K. J. Livak, B. Starcich, S. F.Josephs, E. R. Doran, J. A. Rafalski, E. A. Whitehorn, K.
Baumeister,L. Ivanoff, S. R. Petteway, Jr., M. L.Pearson, J.A. Lauteenberger, T. S. Papas, J. Ghrayeb, N. T. Chang, R. C. Gallo, and F.Wong-Staal.1985. Complete nucleotide sequence
oftheAIDSvirus, HTLV-III.Nature(London) 313:277-283.
33. Rosen, C. A., J. G. Sodroski, W. C. Goh, A. L. Dayton, J. Lippke, and W. A. Haseltine. 1986. Post-transcriptional
regula-tion accounts for thetrans-activation ofthe human T-lympho-tropicvirus type III. Nature(London)319:555-559.
34. Rosen, C. A., E. Terwilliger, A. Dayton, J. G. Sodroski, and W. A.Haseltine.1988.Intragenic cis-actingartgene-responsive
sequences ofthe human immunodeficiency virus. Proc. Natl.
Acad. Sci. USA 85:2071-2075.
35. Roy,S., U.Delling, C.-H. Chen, C. A. Rosen, and N. Sonnen-berg. 1990. AbulgestructureinHIV-I TAR RNA isrequiredfor Tat binding and Tat-mediated trans-activation. Genes Dev.
4:1365-1374.
36. Ruben,S., A. Perkins, R. Purcell, K. Joung, R.Sia,R.Burghoff,
W. A. Haseltine, and C. A. Rosen. 1989. Structural and
func-tional characterization ofhuman immunodeficiency virus Tat
protein. J. Virol.63:1-8.
37. Sambrook, J., E. F.Fritsch, and T. Maniatis. 1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor
Laboratory, ColdSpring Harbor, N.Y.
38. Sharp, P., and R. Marciniak. 1989. HIV TAR: an RNA
en-hancer? Cell 59:229-230.
39. Sodroski, J., W. C. Goh, C. Rosen, A. Dayton, E.Terwilliger,
and W. A. Haseltine.1986. Asecondpost-transcriptional trans-activator generequired for the HTLV-III replication. Nature
(London) 321:412-417.
40. Stoltzfus, C. M., and S.J. Fogarty. 1989.Multiple regionsin the Rous sarcoma virus src gene intron act in cis to affect the
accumulation ofunsplicedRNA. J. Virol.63:1669-1676.
41. Terwilliger, E., R.Burghoff,R. Sia, J. Sodroski,W.Haseltine,
and C. Rosen. 1988. The art gene product of the human
immunodeficiency virus is required for replication. J. Virol. 62:655-658.
42. Theil, E. 1990. Regulation offerritin and transferrin receptor mRNA. J. Biol. Chem. 265:4771-4774.
43. Thomas, P. S. 1980.Hybridizationof denaturedRNAandsmall
DNAfragments transferredtonitrocellulose.Proc. Natl. Acad.
Sci. USA77:5201.
44. Varmus,H. 1988. Regulation ofHIV andHTLV gene
expres-sion.Genes Dev. 2:1055-1062.
45. Zapp, M.,and M.Green.1989.Sequence-specificRNAbinding bythe HIV-1 Revprotein. Nature(London)342:714-716.