0022-538X/83/050371-07$02.00/0
CopyrightC1983,American Society for Microbiology
Characterization of the Herpes
Simplex
Virion-Associated
Factor
Responsible for the Induction
of
otGenes
WILLIAMBATTERSONANDBERNARDROIZMAN*
MarjorieB.KovlerViralOncology Laboratories, TheUniversityof Chicago, Chicago, Illinois 60637 Received27October1982/Accepted 25 January 1983
Herpes simplex virus (HSV) genes form three groups, a.,
(3,
and -y, whosesynthesis
iscoordinately
regulated andsequentially
ordered inacascadefashion.Earlier studies by Post et al. (Cell 24:555-565, 1981) have shown that chimeric
genesconstructedby fusion of 5' noncoding leader andupstreamsequences ofa
genes tothe 5'noncoding leader and structural sequencesof the viralthymidine kinase (TK),a
1B
gene, areregulatedasagenes uponrecombination into the viralgenome. In cells converted from TK- to TK+ phenotype, these chimeric genes are induced by infection with homologous TK- virus. The induction of the resident chimericgenedoesnotrequire viral protein synthesis and isindependent of the presence of functional at gene 4product
required
fortheexpression
of,B genes. In this paper, we report on theproperties
of the a-TK gene chimera resident in converted TK+ murine (L316) and human(1316)
cells. Our resultswere asfollows.(i)
Thepatternof induction of L316 cellsexposed
to0.1, 1.0, and 10PFUpercellsuggested thatexposure to competentvirus is
required
forinduction and that in untreated preparations this virus correspondsto infectious virus. (ii)UV
light-irradiated
viruswasjust
aseffectiveasuntreatedvirus ininducing
a-TKchimeras.
(iii) HSV-1(HFEM)tsB7
induced the a-TK gene chimeras at thenonpermissive (39°C) temperature; at
39°C
theparental HSV-1(HFEM)tsB7
capsids
accumulateatnuclearporesand donotreleaseviralDNA.(iv)
Thea-TKgenechimeras were notinduced by infection withspontaneous TK- mutants of pseudorabies virus and bovine mammillitis virusorwith human
cytomegalovirus
oradenovirustype 2 or
by
exposure tolysates
ofHSV-1-infected cells from whichthevirus was removed by centrifugation. These results indicate that theagene
inducer is a virion component located outside the capsid and that its function might beto stimulate thetranscription ofagenesby recognizing regulatory sites
on viral DNAsorhost cellproducts orboth.
A characteristic of the reproductive cycle of herpes simplex virus 1 (human herpesvirus 1,
HSV-1) is that its gene expression is tightly regulated.
Specifically,
the HSV-1 genes formthree groups, a,
1,
and y, whose synthesis is coordinately regulated and sequentiallyorderedina cascadefashion(10, 11). The five known a
genes are expressed first, and at least one, a
gene 4 (6, 16, 31), is required for the expression
of 1 genes. The1 genesturn offthe synthesis of
a genes and enable the expression ofy genes. On thebasis of studieson viral gene expression
in sensitive and resistant cells exposed to
a-amanitin, Costanzo et al. (5) concluded that cellular RNA polymerase II is involved in the
transcription of viralDNAthroughout the repro-ductive cycle. Central to our understanding of
the regulation of HSV gene expression is the
mechanism by which the cell transcriptional
machinery differentiates between theot,
1,
and-y genes.In an attempt to deal with this
question,
we mapped the site of initiation oftranscription
of fourax genes,i.e., aO,
a4,oa22,
andat27 (19), and constructed chimeric genesconsisting
ofapor-tion ofthe 5'
noncoding
leader sequences andthe sequences upstream from the initiation of transcriptionofagenes fused to the 5'
noncod-ingsequence and the structural sequences of the HSV-1 thymidinekinase (TK), a13 gene(19-22, 26).Theo4-TKgenechimera,whenrecombined into the viral genome, was regulated like an a
gene (26). Ofparticular interest, however, was the observation that in cells converted from
TK- toTK+phenotype, the a4-TK gene
chime-ra was inducedby infectionwith HSV-1 TK- in the presence of cycloheximide and at 39°C in cells infected withtemperature-sensitive (ts)
vi-371
on November 10, 2019 by guest
http://jvi.asm.org/
ruses with mutations in a gene 4 (26). These
observations suggested that unlike the induction of the natural
P-TK
gene(13, 17), the inductionof the chimeric a-TK gene can occur in the
absence of viral protein synthesis and specifical-ly does notrequire thepresence ofafunctional
a4 gene product. Similar results were obtained
with cellsconvertedtotheTK+ phenotype with aO- and a27-TK chimeras (21, 22).
In this paper, we reportthat theinduction of the chimeric a-TKgeneinTK+ converted cells requires exposure of the cellsto virus and that the chimericgeneis inducedby UV
light-irradi-ated virus and by ats mutant defective in the release of viral DNA from capsidsatthe
nonper-missive temperature, but not by several other herpesviruses orby adenovirus type2 (Ad-2).
MATERIALSANDMETHODS
Virusand cells.Theproceduresforthepropagation of virus and cellsand thecomposition ofgrowth and
maintenancemediahave beenpublishedelsewhere(7, 29). The construction and properties of
HSV-1(F)ts502A305, temperature sensitive inthe a4 gene andcontaininga700-basepairdeletion in the TKgene,
have been previously described (26).
HSV-1(HFEM)tsB7 syn+ wasisolated by
bromodeoxyuri-dine mutagenesis of HSV-1(HFEM)-infected cells
(14).Previous studieshave shown that incells infected withHSV-1(HFEM)tsB7 and maintainedat39°C, viral capsids accumulateatthe nuclearmembrane andno
viralgene expression can bedetected (1). Ad-2 was
obtained from M.Green, Institute forMolecular
Virol-ogy, St.Louis, Mo. Humancytomegalovirus (human herpesvirus5,CMV), strain AD 169,wasprovided by
F. Rapp, The Pennsylvania State University,
Her-shey. HSV-2(333-B3) TK- stock wasobtained from
R. B.Tenser, The PennsylvaniaStateUniversity.
TK-negative variants of bovine mammilitis virus (bovine herpesvirus-2, BMV) and ofpseudorabiesvirus(Suid herpesvirus-1, PRV)were selected bypreviously
de-scribed methods(26).
Ltk-cells(12)wereobtained from S. Kit,Medical
College of Baylor University, Houston, Tex. The
143(TK-) humancell line (4) was obtained from C.
Croce, The Wistar Institute, Philadelphia, Pa. L316 cells and 1316 cellswereprepared byconverting
Ltk-and143(TK-) cells, respectively,toaTK+ phenotype
withpRB316,which contains DNAsequences
encod-ing the a4 gene promoter-regulator fused to DNA
sequencescoding for the structuralportionof the TK
gene(19-22, 26). L103 cellsprepared bytransfection of Ltk- cells with theHSV-1(F) BamHI Q fragment
were aspreviously described(26).
TK+cellswereselected and maintained inmedium
containing10-5Mhypoxanthine,1.6 x10-5M
thymi-dine,and 4.4x 10-7M methotrexate.
TK assays. Procedures forsuperinfection of
trans-formedTK+cells and for extraction andassayofTK activity in these cells were as previously described
(19-22, 26).
Preparationofsupernatant fluid from HSV-1-infected
cell lysates. The preparation of infected cell lysate
stocks has been described elsewhere in detail(7, 29).
Specifically, Vero cells infected with
HSV-1(F)ts502A305andmaintained at 33°C were disrupted in a Dounce homogenizer and then sonicated at the maximum intensity with a Branson Cell Disruptor (model 200). The infected cell lysate was cleared of cell debris bycentrifugation, the titer was determined tobe 1.9 x 109PFU/ml in Vero cells, and thelysate
wasstored at-70°C. The virus stocks were diluted to concentrations of4 x 107 PFU/1.5 ml and 4 x 108 PFU/1.5 ml withphosphate-buffered saline containing 1% (wt/wt) glucose and centrifuged in a Beckman SW41 rotor for 90 minat30,000 rpm and4°C to pellet thevirions. Thesupernatantfluids were then exposed to L316cells as described above.
UV irradiationofvirus. Stock HSV-1(F)ts502A305
virus was suspended in phosphate-buffered saline con-taining 1% glucose to yieldaconcentration of 2 x 107
PFU/1.5 ml. A2-ml amount of virus suspension in a
10-cm-diameter petri dish was shaken on a rotary
platform and exposed to a germicidal lamp (wave-length, 254 nm). A total dose of 1,000ergs/mm2 was deliveredatthe rate of 15ergs/s permm2tothesurface of the fluid. Samples (1.5 ml) of irradiated virus were thenused to infect L316 cells. A portion of the UV light-irradiated virus was assayed for infectivity.
RESULTS
Effect of
multiplicity
ofinfectionon induction of the a-TK chimeric gene in L316cells. The pur-poseof this series ofexperiments wasto deter-mine theeffect of the multiplicityof infectionontheinduction of thea-TKchimericgeneresident in L316 cells.
In these experiments, replicate cultures of
L316cellswere exposedtoHSV-1(F)ts502A305
at a multiplicity of infection of 0.1, 1, and 10
PFU per cell for 2 h. Viral inocula were then removed, and the infected monolayers were
washed, replenished with fresh medium, and
additionally
incubated at eitherpermissive
(33°C)ornonpermissive (39°C)temperatures. At
appropriate
times,
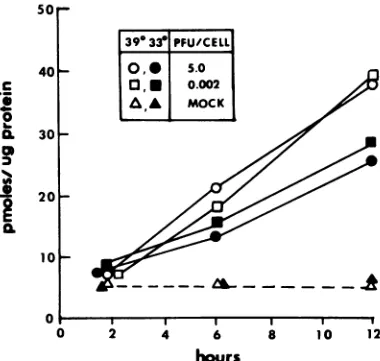
the cultures were harvested, and cell extracts prepared from the infected cultures were assayed for TK activity. Theresults of these experiments (Fig. 1)
demon-stratethat atbothpermissiveandnonpermissive
temperatures, inductionrequires infection of the
cellcarryingtheresidenta-TKgene chimera.As
has been previously observed, the extent of
a-TKinductionwasgreater at39°Cthanat33°C in L316 cells infected with HSV-1(F)ts502A305
(26).
Theseresults thusaffirmand extendprevious observations that induction ofthe a-TK
chime-ric gene residentin L316cells isaconsequence
ofviralinfection.
Ability of UV light-irradiated virus to induce the a-TK gene. Inasmuch as induction of
tran-scription of thea-TKgene can takeplaceincells infected in the presence of
cyclohexmide
(26),thepurpose of these experiments was to deter-mine whether UVlight-irradiated virus retained its abilitytoinduce TK activity inL316 cells.
on November 10, 2019 by guest
http://jvi.asm.org/
O,,o,39° 010 L316 cells were infected with 5 PFU of HSV-50 -
*-330
1(HFEM)tsB7,
or ofHSV-1(F)ts502A305,
perl. .UJ./cell or were mock
infected,
and the cultures40- I were then incubated at the permissive (33°C) or
nonpermissive (39°C) temperature. At various
timesafter infection, the cultures were harvest-ed,and cellextracts wereassayed for TK
activi--3/
/ ty. The results (Fig. 3A) indicate that at 390C,HSV-1(HFEM)tsB7was ascapable ofinducing
10 a-TK activity as was
HSV-1(F)ts502A305
and20 - thatat
330C,
HSV-1(HFEM)tsB7
inducedsome-1 what more a-TK
activity
than didHSV-01
1(F)ts502A305. Since HSV-1(HFEM)tsB7 isnot10- 0.1 a TK- mutant, its ability to express TK activity
--k
was assayed in 143(TK-) cells (Fig. 3C). Theresults of this experiment showed that
HSV-, , , . , 1(HFEM)tsB7 expressed a small amount of TK
0 2 4 6 8 10 12 activity at 330C, but none at 390C.
hours These results indicate that the a-TK gene
chimera can be induced at the nonpermissive FIG. 1. Effect ofmultiplicity ofviral infection on temperature by
HSV-1(HFEM)tsB7.
Previousluctionof TK activity from a4-TK gene chimeras in studies have shown that in cells infected with
16cells. Replicatecell cultureswereinfected with
,1.0,or10.0PFU of
HSV-1(F)ts502A305
virusper HSV-1(HFEM)tsB7 parentalvirus, capsids
ac-ll, and TK activity was measured as previously cumulate at390C
at the nuclear membrane and scribed (20-22) in cells harvested at the times do not release theviral
DNA into the nucleus )wn. TK activity is expressed as picomolesof TMP until thetemperature is shifted to the permissivemedpermicrogram of protein. After adsorptionof range. Consistent with this
observation,
noviral us to cells at 100C, the cells were transferred to gene expression was detected at 39°C (1, 14). her 33 or 390C. Time 0 representsthe time at which Thus,the data presented here demonstrate that lls were placed at 33 or 390C. theinduction of the chimeric a-TK gene in L316cellsmustbemediated byanextracapsid virion
component.
L316cellswereinfected with5PFUof HSV- Effect of supernatant fluids of infected cell
7)ts502A305 percellor withanequalamount lysateson the a-TK gene. To test the hypothesis
of virions that had been exposed to UV-light irradiation as described above. After 2 h of
adsorption,
the cells were treated as describedabove. The UV-light dosage was sufficient to
reduce the effective input multiplicities of the inocula from 5 PFU per cell in the untreated viral suspensions to 0.002 PFU per cell in the
UV
light-irradiated
samples. The L316 cellswereextractedand assayedforTKactivityat 2, 6, and 12 h afterexposure to untreated or UV light-irradiated virus.
The results (Fig. 2) demonstrate that virus exposedto
UV-light
irradiation retained itsabili-ty to induce TK activity in L316 cells atlevels comparabletothose inducedby untreated virus, notwithstanding the loss in infectivity. These results are consistent with the conclusion that thefactor responsible for induction ofTK activi-ty inL316 cells is a structuralcomponentofthe
virion.
Effect ofHSV-1(HFEM)tsB7 on TKactivityin L316cells. The purpose ofthis seriesof
experi-ments was to determine whether
HSV-l(HFEM)tsB7 could induce the resident a-TK
chimeric genes at the nonpermissive tempera-ture.
In these experiments, replicate cultures of
50r-c
0
h.
06
.2
40?-30h
20H
lo0
___ _ --.A
0 2 4 6 8 10 12
hours
FIG. 2. Induction ofa4-TKgenechimeras inL316 cellsby UV-irradiated virus. Cellswereinfected with
5PFUofHSV-1(F)ts502A305 viruspercellorwithan
equivalent amount of HSV-1(F)ts502A305 virus that
had been UV irradiated as described in the text. Infectedcultureswereincubated atthetemperatures
andforthetimesshown.
c 0 0. 0
A6
06 E 0.
nd L3 0.1 cel des shc for vir
eiti
cel
1(F
VOL.46,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.490.47.238.58.270.2] [image:3.490.251.441.436.617.2]BATTERSON
observation that induction of TK activity is
demonstrable after exposure of L316cells to 1 tsB7 PFU per cell (Fig. 1), thedataindicate that the
ts502A305
supernatantfluidsaredevoid ofinducing
activi-tyand that the inducing activity partitions with the virus.
Effect of infection withHSV-2(333-B3)TK-on
the a-TK gene. In this series of experiments, ftB7 L316 cells were infected with HSV-2(333-B3) TK- virus at a multiplicity of infection of10
PFUper cell. The results (not shown) indicate that HSV-2 induces the HSV-1 a-TKgene chi-meras in the same fashion as the homologous
hS02A305
virus. Inasmuch as the studies on HSV-1 xHSV-2 recombinants failed to reveal forbidden
juxtapositions of HSV-1 and HSV-2 DNA se-quences(23-25), the resultswere not unexpect-ed.
Effect of other viruses on the a-TKgene. The purposeof these experiments was to determine
whether otherviruses, particularly PRV,BMV, CMV, and Ad-2, were able to induce resident a-PlRv TK gene chimeras in transformed cells. Since
iBMV
BMVandPRVeachspecifyaTK, we selected~MOCKCS
mutants deficient in thisfunction,
asdescribed above.The experimental proceduresfor infection and maintenance of cells withtheseviruseswerethe
same as those described above. The
multiplic-ities ofinfectionwere10 PFUof PRV andBMVftB7
,percell, 100PFUof Ad-2 per
cell,
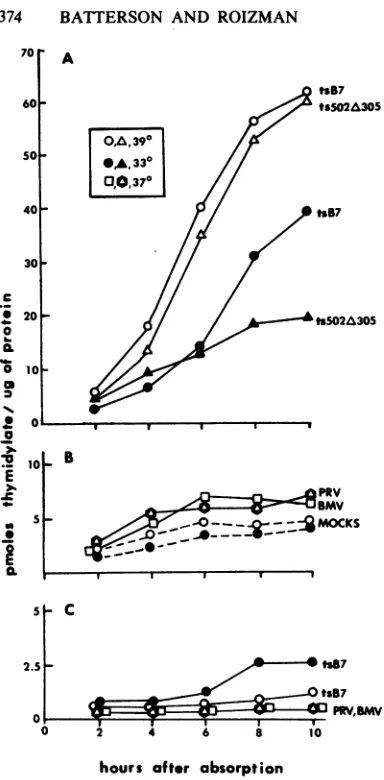
and 50 PFU of CMV percell. The results (Fig. 3) show theDtsB7 following.
GuPWV,BMV
[image:4.490.54.246.54.444.2]hours ofter absorption
FIG. 3. Induction of TKactivity byTK- HSV-1, BMV,and PRVherpesviruses. Theexperimental
pro-tocols are described in the text. The reference box
symbolsin A denote thetemperatureofincubation and
typeof virus usedin theexperiments illustratedinA, B, and C. (A) Inductionof TKactivityin L316 cells
infected with HSV-1(HFEM)tsB7 and by
HSV-1(F)ts502A305. (B) Induction of TKactivity in L316 cellsbyTK-mutantsof BMV and PRV. Selection of these TK- viruses is described in the text. (C) TK
activity in 143(TK-) cells infected with
HSV-1(HFEM)tsB7, BMV(TK-), or PRV(TK-) viruses. Note thatHSV-1(F)ts502A305 carriesa700-basepair
deletion inthe TKgene(26).
thatthe a-TKgeneinL316 cellsis inducedbya
nonvirion-associated factor accumulating in in-fected cell lysates, L316 cells were exposed to
supernatantfluids obtainedafterhigh-speed
cen-trifugation oflysates of Vero cells infected with HSV-1(F)ts502A305 as described above. The
L316 cells were notinduced after exposure to
supernatant fluids containing the equivalent of
10or 100 PFUpercell(Table1). Inlight of the
TABLE 1. TKactivity of L316 cells exposed to supernatantfluids oflysates of Vero cells infected
with HSV-1(F)ts502A305 at 33°C
Hourspost- pmol ofthymidylateper
p.g
ofproteinin:exposure or Cellsex-
Mock--infection' Infected posed to Mock-cells supernatant
ifctedl
fluidcelExpt1
2 9.7 7.1 6.3
6 26.2 5.2 7.4
12 49.4 6.9 6.5
Expt2
2 NDb 5.3 5.8
6 ND 6.1 4.9
a The
supernatant
fluids were adjusted to yield avirus-equivalent concentration of the cell lysateof 10 PFU (experiment 1) or100 PFU (experiment 2) per cell. The procedure forpreparation ofthesupernatant fluid is described inthe text.
bND, Not done (preliminary experiments have
es-tablishedthat at highmultiplicities of infection [e.g.,
100 PFUpercell], theinduction of the oa-TK gene in
L316 cells wasgenerally not as highas atlower (10
PFU percell) multiplicities ofinfection).
7or
A60
50
40
OA, 390
OA, 330
0,0,370
30-c
* 20
a
0.6 10 0 10 co 5
10
E
£ 5
5
- c
2.5_
2 4 6 a 10
J. VIROL.
- -
-4
la C
- IO
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.490.258.447.459.586.2]INDUCTION OF a GENES BY HERPES SIMPLEX VIRUS
(i) None of the viruses
selected for
this study induced TK activity in human 143(TK-) cells. Theresultsobtained for BMV and PRVmutants areshowninFig. 3C.(ii) Neither BMV nor PRV significantly in-duced the resident a-TKgene chimerain L316 cells(Fig. 3B).
(iii) NeitherCMV norAd-2induced the resi-dent a-TK gene chimera in I316 cells
(Fig.
4). Attempts to induce the enzyme in L316 cells with these viruses alsofailed.DISCUSSION
Inthispaper, weaffirm and expand the
previ-ousreport (26) from this laboratory that
(i)
a4-TK gene chimeras are induced in
converted,
TK+ cells by infection withTK- virus, (ii) the induction isindependent of the synthesis ofa4 gene product, and
(iii)
the induction does notrequire the synthesis of viral proteins directed by the DNA
of
infecting
virus. The results presented in thisreport indicate that the induc-tion isdependent
onexposure tovirus,
that UVlight-inactivated
virusisaseffectiveasuntreatedvirus, and that the chimeric gene is induced at
the
nonpermissive
temperatureby
anHSV-1 tsmutant defective in release of viral DNA from
capsids,
but notby
otherherpesviruses
norby
Ad-2.
The discovery that a4-TK genechimeras are inducibleby infection (26)wasunexpected inas-much as the infectivity of the
deproteinized
DNAsuggested that the introduction of compe-tent viral DNA is all that is
required
for aprogrammatic
execution of the reproductivecy-clein
permissive
cells. The observations thata-TK gene chimeras are
inducible
(26), that the inducibility is conferred by amovable "regula-tory" sequence locatedatleast 100nucleotidesupstreamfrom the site of initiation of
transcrip-tion ofagenes(20),
and that alla genescontain within the regulatory regions homologs of a consensus sequence (22) suggest that a genes areindeedinduced andraise questionsregarding thenatureof theageneinducer and itsfunction in thereproductive
cycle of the virus.Signal for induction ofa genes. The observa-tions reported in this andprevious publications indicate that the
signal
for the induction of a genes is HSV-1 and HSV-2 specific and mostlikelyastructuralcomponentlocatedoutside the
capsid.
Specifically,
theeffect ofthemultiplicity of infectionsuggests thatinduction ofa-TKgenechimeras
requires
exposure to competent virusand that the concentration of this virus in the inoculum approximates that of the infectious
virus. However, theexperimentwith UV light-irradiated virus as well as the results of induc-tion ofthe a4-TK genechimera inthepresence
of inhibitory concentrations of cycloheximide
*ts502A305,390
OtS5024305,330
ACMV, 370 OAD-2,370 XMOCK,370
c 0
a.
0
0 2 4 6 8 10
hours
FIG. 4. Induction of TK activity in 1316 cells
1143(TK-)cells transformedtoTK+ phenotypewith anoa-TK gene chimera contained inplasmid pRB316]
(26)byHSV-1(F)ts502A305, Ad-2,andCMV.
(26)
indicate thatthesynthesis
of viralproteins
by
theinfecting
virus isnotrequired.
The stud-ies with theHSV-1(HFEM)tsB7
mutantarecon-sistent withthisconclusionandsuggestthat the
structuralcomponent
responsible
forthe induc-tionofthe virusis located outsidethecapsid.
Itis of interest to note that the HSV-1 virion containsatleast 24 structural
proteins
(9, 27),
of whichnomorethanhalfareeitheratthesurfaceofthe
envelope
orin thecapsid.
Thefunctionofthe
noncapsid,
nonenvelope
proteins
is un-known.Conceivably,
among theseproteins
arethose
responsible
for the induction ofaL genesand forotherfunctions suchasthe virion-associ-ated shutoff of host macromolecular
synthesis
(8)
and theputative
virionprotein
kinase(18).
Function ofageneinducer. Inasmuchasviral
mutants
lacking
aLregulatory
regions
have notbeen
constructed,
the functionoftheregulatory
regions
and therequirement
forinduction of theagenes afterinfection havenotbeen
explored.
Itisconceivable that the function of the oLgene inducer is to enhance
transcription
of the agenes
by
binding
eithertotheviral genomeortohost gene
products.
We cannotdifferentiatebe-tween these alternatives. However, BMV and
375 VOL.46,1983
16
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.490.253.437.71.360.2]PRV share many biological properties with
HSV-1 and HSV-2 and belong to the same subfamilyof herpesviruses (28); the observation thatneitherPRVnorBMV, aherpesvirus struc-turally and genetically related to HSV-1 and HSV-2 (2, 3, 30),induced the a-TKgene chime-rasuggeststhat theageneinducer hasa recogni-tion site for specific sites in the viral DNA in addition to any putative recognition sites for
hostgeneproducts. The observation that the
a-TKgene chimerasare induced atahigher level
at39 thanat33°C isof considerable interest and
potential helpindetermining thefunctionofthe inducer but does not differentiate between the
alternatives listed above.
A central question is whether, in the absence of the a gene inducer or in the absence of
regulatory regions, the agenes contained in the viralDNA would betranscribed atalevel suffi-ciently high to sustain viral multiplication. The observation that a-TK gene chimeras are
tran-scribed in the absence of the a gene inducer is
notcompelling evidence that they wouldbe
tran-scribed as partofthe DNA in the infected
cell;
thus, in the absence of functional a4 gene
prod-uct, thenatural ,-TKgeneistranscribedwhenit
is a resident gene in cells converted to TK+
phenotypebut notwhen it ispartofthegenome
ofan infecting virus.
Our results would havepredicted that
depro-teinized HSV-1 DNA wouldnotbe
infectious;
in fact,thespecific infectivityofHSV-1 DNA isoftheorder of106 moleculesperPFU(15). Among
the possible explanations are that (i) the
repro-ductive cycle initiated by deproteinized DNA
depends on the participation offunctional host
analogs of a proteins and is therefore very
inefficient or (ii) the reproductive cycle begins
with the synthesis of the a gene inducer and
continueswith aproteinsratherthanwith thea
proteins asisthe casein thereproductivecycle
initiated by virions. Additional studies should
reveal thenatureandfunction of the inducerand
its siteof action.
ACKNOWLEDGMENTS
Thesestudies wereaided byPublicHealth Servicegrants
CA-08494 and CA-19264 from the National Cancer Institute andby American CancerSocietygrantMV-2R.W.B. was a
PublicHealthServicepostdoctoral trainee(grant 5 T32 PHS
AI07182).
LITERATURECITED
1. Batterson, W., D. Furlong, and B. Roizrnan. 1983.
Molecu-largeneticsofherpes simplex virus.VIII. Further charac-terizationofatemperature-sensitive mutantdefective in
release ofviral DNA and inother stages ofthe viral
reproductive cycle.J.Virol. 45:397-407.
2. Buchman, T. G., and B. Roizman. 1978. Anatomy of
bovine mammillitis DNA. I. Restriction endonuclease
maps of fourpopulations ofmoleculesthatdiffer inthe
relativeorientation of their long andshortcomponents.J. Virol. 25:395-407.
3. Buchman, T. G., and B. Roizman. 1978. Anatomy of bovinemammillitis DNA. II.Size andarrangementsofthe deoxynucleotidesequences. J. Virol. 27:239-254. 4. Campione-Piccardo, J., W. E. Rawls, and S. Bacchetti.
1979. Selective assay for herpes simplexviruses express-ingthymidinekinase. J. Virol. 31:281-287.
5. Costanzo, F., G. Campadefli-Fiume, L.FoA-Tomasi, and E.Cassai. 1977.Evidence that herpes simplex virus DNA is transcribed by cellular RNA polymerase B. J. Virol. 21:996-1001.
6. Dixon, R.A. F., and P. A. Schaffer. 1980. Fine-structure mappingand functionalanalysis of temperature-sensitive
mutants in the gene encoding the herpes simplex virus
type1 immediate early protein VP175. J. Virol. 36:189-203.
7. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968. Characterization ofherpes simplex virus strains differing intheir effectsonsocial behavior of infectedcells. J.Gen. Virol. 2:357-364.
8. Fenwick, M. L., and M. J. Walker.1978. Suppression of thesynthesis of cellularmacromoleculesbyherpes sim-plex virus. J. Gen. Virol. 41:37-51.
9. Heine, J. W., R.W. Honess, E.Cassai,and B.Roizman.
1974.Proteins specifiedby herpes simplex virus. XII. The
virion polypeptides of type 1 strains. J. Virol.14:640-651. 10. Honess, R. W., and B. Roizman. 1974. Regulation of
herpesvirus macromolecular synthesis. I. Cascade regula-tionofthesynthesis of threegroups of viral proteins. J. Virol. 14:8-19.
11. Honess, R. W., and B. Roizman. 1975. Regulation of herpesvirus macromolecularsynthesis: sequential transi-tion of polypeptide synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A. 72:1276-1280.
12. Kit, S.,D.R. Dubbs, L. J.Piekarski,and T. C.Hsu. 1963. Deletion of thymidine kinase activity from L cells resis-tanttobromodeoxyuridine. Exp. Cell Res. 31:297-312. 13. Kit, S., D. R. Dubbs, and P. A. Schaffer. 1978.Thymidine
kinase activity of biochemically transformed mousecells after superinfection by thymidine kinase-negative,
tem-perature-sensitive,herpes simplex virus mutants. Virolo-gy85:456-463.
14. Knipe, D. M., W. Batterson, C. Nosal, B. Roizman,and A. Buchan. 1981.Molecular genetics of herpes simplexvirus. VI. Characterization of a temperature-sensitive mutant defective in the expression of all early viral gene products. J.Virol. 38:539-547.
15. Knipe, D. M., W. T. Ruyechan, and B.Rolzman. 1979. Molecular genetics of herpes simplex virus. III. Fine mapping of a genetic locus determining resistance to phosphonoacetate by two methods of marker transfer. J. Virol. 29:698-704.
16. Knipe,D. M., W. T. Ruyechan, B.Rolzman, and I. W. Halliburton. 1978. Moleculargenetics of herpessimplex virus: demonstration of regions of obligatory and
non-obligatoryidentity within diploid regions of the genome by sequence replacement and insertion. Proc. Natl. Acad. Sci.U.S.A. 75:3896-3900.
17. Leiden, J. M.,R.Buttyan, and P. G. Spear. 1976. Herpes
simplex virus gene expression intransformed cells. I.
Regulation of the viral thymidine kinase gene in trans-formed Lcells by products of superinfecting virus. J. Virol. 20:413-424.
18. Lemaster, S., and B.Roizman. 1980. Herpes simplex virus phosphoproteins. II. Characterization of the virion pro-teinkinase and of the polypeptides phosphorylated in the virion. J. Virol. 35:798-811.
19. Mackem, S., and B.Roizman. 1980. Regulation of herpes-virus macromolecular synthesis: transcription-initiation sites and domains of a genes. Proc. NatI. Acad. Sci. U.S.A. 77:7122-7126.
20. Mackem, S., and B. Roizman. 1982. Differentiation
be-tween apromoterandregulator regions of herpessimplex
virus 1: the functional domains and sequence of a movable atregulator. Proc. Natl. Acad. Sci. U.S.A. 79:4917-4921.
on November 10, 2019 by guest
http://jvi.asm.org/
INDUCTION OF a GENES BY HERPES SIMPLEX VIRUS 377 21. Mackem, S., and B. Rolzman. 1982. Regulation ofagenes
of herpes simplex virus: thea27gene
promotor-thymi-dine kinase chimera is postively regulated in converted L cells. J.Virol. 43:1015-1023.
22. Mackem, S.,andB.Rolzman. 1982.Structural features of the herpes simplex virusagene4,0, and 27
promoter-regulatorysequenceswhich conferaregulationon chime-ric thymidine kinasegenes.J.Virol. 44:939-949.
23. Marsden, H. S., N. D. Stow, V. G. Preston, M. C.
Tinbury, and N.M.Wilkie. 1978. Physical mapping of herpes simplex virus-induced polypeptides. J. Virol.
28:624-642.
24. Morse, L. S., T. G. Buchna, B. Roizman,and P. A.
Schaffer. 1977. Anatomyof herpes simplex virus DNA. IX. Apparent exclusionofsomeparental DNA
arrange-ments inthegeneration of intertypic (HSV-1 x HSV-2) recombinants. J. Virol.24:231-248.
25. Morse, L. S., L. Pereira, B. Roizznan,andP.A.Schaffer. 1978.Anatomy of herpes simplex virus (HSV) DNA. X. Mapping of viralgenesby analysis of polypeptides and functions specified by HSV-1 xHSV-2 recombinants.J.
Virol 26:389-410.
26. Post, L. E., S. Mackem, and B. Roiznan. 1981.Regulation ofagenesofherpes simplex virus: expression of chimeric
genesproduced by fusion of thymidine kinasewithagene
promoters.Cell 24:555-565.
27. Spear, P. G.,andB. Rolzman. 1972. Proteinsspecified by herpes simplex virus. V. Purification and structural
pro-teins of the herpesvirion. J. Virol. 9:143-159.
28. Rolzna, B., L. E. Carmichael,F.Delnhardt, G. de-The, A.J. Nahmias, W. Plowright, F.Rapp,P.Shehdrick, M. Takahashi, and K. Wolf. 1981.Herpesviridae: definition, provisional nomenclature, and taxonomy. Intervirology 16:201-217.
29. Rolzman, B.,andP. G.Spear. 1968.Preparation of herpes simplex virus of high titer.J.Virol. 2:83-84.
30. Stetz,H.,H. Ludwig, and R. Rott.1973/1974.
Immuno-logic and genetic relationship between herpes simplex virusandbovinemammallitis virus. Intervirology2:1-13.
31. Watson, R. J., and J. B. Clements. 1980. A herpes simplex virustype 1functioncontinuously required for early and late virus RNA synthesis. Nature (London) 285:329-330. VOL. 46, 1983
![FIG. 4.an(26)1143(TK-) oa-TK Induction of TK activityin 1316cells cells transformed to TK+ phenotype with gene chimera contained in plasmid pRB316] by HSV-1(F)ts502A305, Ad-2, and CMV.](https://thumb-us.123doks.com/thumbv2/123dok_us/1446664.97127/5.490.253.437.71.360/induction-activityin-cells-transformed-phenotype-chimera-contained-plasmid.webp)
