Vol.53,No. 2 JOURNALOFVIROLOGY, Feb. 1985, p.456-461
0022-538X/85/020456-06$02.00/0
CopyrightC 1985,American Society for Microbiology
Novel
Rearrangements
of
Herpes
Simplex
Virus DNA Sequences
Resulting from Duplication of
a
Sequence Within
the
Unique Region
of the
L
Component
KAY L. POGUE-GEILE, GLORIAT.-Y. LEE, ANDPATRICIA G. SPEAR* Department ofMicrobiology, University of Chicago, Chicago, llinois 60637
Received 29May 1984/Accepted 15 October 1984
We constructed insertion mutants of herpes simplex virus type 1 that contained a duplication of DNA sequences from the BamHI-L fragment (map units 0.706 to 0.744), which is located in the unique region of the Lcomponent (UL)ofthe herpes simplex virus
type
1genome. The second copy of the BamHI-L sequence was inserted ininverted orientation into the viral thymidine kinase gene (map units 0.30 to 0.32), also located Within UL. Asignificant fraction of the progeny produced by these insertion mutants had genomes with rearranged DNA sequences, presumably resulting from intramolecular or intermolecular recombination between the BamHI-Lsequences at the twodifferentgenomic locations. The rearranged genomes either had an inversion of theDNA sequenceflankedby theduplication or were recombinant molecules in whichdifferentregions ofthe genome had beendfplicated
and deleted. Genomicrearrangements similar to those described here have been reported previously but only for herpes simplex virus insertion mutants containing an extra copy of the repetitive a sequence. Such rearrangements have not been reported for insertion mutants that contain duplications of herpes simplex virus DNA sequences from largely unique regions of the genome. The implications of these results are discussed.The genome of herpes simplex virus (HSV) is 96 x 106
daltons (Da) and is composed oftwo segments, long (UL) and short (Us), of unique DNA. These two segments are each flanked by inverted repeats, the ends of which are common toboth segments andarerepeated in direct
orien-tationatthe endsof thegenome (4, 18, 20, 26, 28)(seeFig.
1).Recombination apparentlyoccursbetweentheseinverted
repeats, resulting in the inversion ofthe unique segments
flanked by the repeats. Because UL and
Us
can invertindependently ofeach other, populations ofHSV genomes usually contain equimolar quantities of the four possible isomers (5, 10, 29, 37).
Previous studies showed that, when specific HSV type 1
(HSV-1) DNA sequences are duplicated by insertion of a
second copyintothethymidine kinase(tk) gene of the virus,
inversions ofthe DNA sequence flanked by the duplicated
sequence can occur. These inversions could be detected in the genomes ofprogeny virus, provided that the inserted
sequence and the sequence at the normal locus were in
inverted orientation and that the inserted sequence
con-tained the repetitive sequencedesignated a(17, 18, 20, 30).
Inversions could not be detected when inserted sequences were from unique regions of the genome (7, 14, 17). We report here, however, that duplication ofa sequencefrom
the UL component can also result, at apparently high fre-quency, either in inversionofthe DNAsequences flankedby
invertedcopiesoftheduplicationor in other DNA rearrange-mentsresultingfromintermolecular recombinationbetween an inserted BamHI-L sequence and the same sequence at thenormallocus.
Thestudies described here wereinitiated for the purpose
ofcharacterizing a putative
fISV-1
gene product that gov-erns theprocess ofHSV-inducedcellfusion. MutantHSV-* Correspondingauthor.
tPresent address: School of Medicine, University of Miami, Miami, FL 33101.
1(MP) causes the fusion of infected cells under conditions in which awild-type sibling strain,HSV-1(mP), does not (12). The mutation was mapped first to mapunits 0.70 to 0.83(27)
and more recently to map upits 0.735 to 0.740 (2, 22), a region contained within the BamHI-Lfragment of the HSV-1 genome. Insertion mutants were constructed which con-tained the
BamHIH-L
fragment ofHSV-l(mP)DNA inserted into the tk gene ofHSV-1(MP) DNA and vice versa; this was done tocharacterize aproductexpressed from the insert that could either cause HSV-1(mP) to induce cell fusion or complement the mutation of HSV-1(MP). The insertion mutantsisolated did not serve the original purpose,because marker rescue and marker transferfromtheinsertedlocus to the normal locus(and viceversa) appeared to occur at high frequency, possibly by mechanisms proposed and discussedpreviously (4, 13, 32). This precluded the
correlation
of phenotype with expression of products from the inserted DNA. This study therefore focuses on the novel DNA arrangementsoccurringin these mutants.MATERIALS ANDMETHODS
Cells and viruses. Vero and HEp-2 cells were obtained from theAmerican Type CultureCollection, Rockville,Md.
HEp-2cells weregrown in DulbeccomodifiedEaglemedium with 10% fetal calf serum, and Vero cells were grown in medium 199with5%fetal calf serum. The viralstrainsused wereHSV-1(MP) andHSV-1(mP) (6, 11, 12).
Plasmids. All vector sequences present in the plasmids
used in
thi5
study were from pBR322 (1). pRB142 andpRB103,containingthe BamHI-D' andBamHI-Q fragments, respectively, from HSV-1(F) DNA (23), were kindly
sup-plied by Bernard Roizman, University of Chicago. pKPG103R was subcloned from pRB103 by digestion with
SalI and BgIlI followed
by
religation
at37°C
for 1 h. To obtain pKPG103L, pRB103 was digested with HindIII andBglII, thelargerfragmentwas isolatedfromanagarosegel,
and the ends were filled in with DNApolymerase I Klenow fragment
Iand
religated at 16°C for 8 h(15). pKPG103R and 456on November 10, 2019 by guest
http://jvi.asm.org/
REARRANGEMENTS OF HSV DNA SEQUENCES 457
mapunits I.. 0.0
. a b
HSV-I(MP) C2Z
, , , -I
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
tk L - b aac' S ca
pTK 106
Ba
a l HSV- (mP) Iii
Bg/Ba Ba/Bg Ba
b tk Lt
_~~~--_ ~
_ -
:
ll*
aa'c S ca
pTK 133
Ba Bg/Ba Ba/Bg Ba
molecular
weights - 1.7 -3.5 1 0.5H
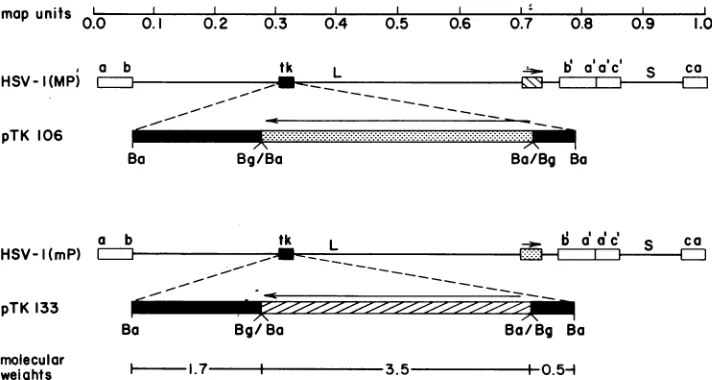
FIG. 1. Mapsof the HSV-1(MP) andHSV-1(mP)genomesand of theplasmids used in these studies. Theorganizationof thegenomeswith respect toUL andUsand therepetitive sequences (LI )is shownfor bothHSV-1(MP) and HSV-1(mP). Thetkgene (i ),the BamHI-L sequenceofHSV-1(MP) (
M
),andtheBamHI-LsequenceofHSV-1(mP) (@ )areshown in theappropriate viralgenomesandplasmids. The arrows indicate the directions of the two BamHI-L sequences withrespect to eachother in the insertion mutants. The restriction endonucleases used for cloning these HSV-1 sequences were BglII (Bg) andBamHI (Ba). To construct pTK133, the cloned BamHI-L sequencefromHSV-1(MP)presentinplasmid pTYL133wasrecloned into thesingle BgII site of the tkgeneinpRB103. Thesameprocedure was used to construct pTK106, except that the BamHI-L fragment was from pTYL106, which contains the BamHI-L fragment from HSV-1(mP). TheBglII-BamHI (Bg/Ba) ligation joints are resistant to cleavage by bothenzymes. The sizes(106 Da) ofthe ligatedDNA fragments areindicated.pKPG103L contain theBamHI-Q sequences thatare to the right and left, respectively, ofthe single BgIII site in this fragment (Fig. 1). pTK133andpTK106contain the BamHI-Lfragment (mapunits 0.706to0.744)fromHSV-1(MP)and HSV-l(mP) DNAs, respectively, insertedinto theBgIII site ofpRB103. Construction of these plasmids isdescribed in thelegendtoFig.1.pKPG101containsaSall fragment (map units 0.702to0.752) fromHSV-1(MP)DNA. pKPG101 was digestedwith BamHI andreligatedtoformpKPG102,which contains HSV-1 DNAsequencesfrom the BamHIsitetothe SalIsiteat0.744 and 0.752mapunits, respectively.
Construction of recombinant viruses. Rabbit skin cells werecotransfected eitherwithmixtures ofHSV-1(MP) DNA and BamHI-digested pTK106 DNA or with mixtures of HSV-1(mP)DNAandBamHI-digested pTK133DNA (8,14, 36). When thetotalcytopathiceffectwasobserved,progeny
viruswereplated underagarose containingthymidine arabi-noside (araT) at 100 ,Lg/ml (17). A second plating under
agarose with araTwasdone to purifythe virus.
DNApreparationandanalysis. Plasmid DNAwaspurified in cesium chloride gradients by equilibrium centrifugation (25) of cleared lysates (9). HSV-1(MP) and HSV-1(mP) DNAsused for transfections orcloningwere purified in Nal
density gradients as described by Walboomers and ter Scheggt (35) and modified by Zezulak and Spear (38). Recombinant viral DNAs usedfor Southern blotting exper-iments were obtained from Vero cells infected with
recom-binantvirus that had beenplaque purifiedtwice. These viral DNAs wereprepared by phenol extraction ofthe cytoplas-mic fraction of infected cells lysed with Nonidet P-40 (38). The DNAs weredigested with restriction enzymes as
spec-ified by the manufacturer (New England BioLabs, Inc.), electrophoresed on agarose gels, and blotted to nitrocellu-lose(31). Hybridizations were doneat65°Cin 6x SSC(0.9 M NaCl and0.09 M sodium citrate)-30% formamide-salmon
sperm DNA (50 p.g/ml)-lx Denhardt solution (15)-sodium dodecylsulfate 0.5% for 8to15 h. Nitrocellulosefilterswere
washed three times in 2x SSC, one time in lx SSC, and
finally in 0.4x SSC. Nick-translation kits (New England
NuclearCorp.) wereusedto make 32P-labeled DNAprobes fromundigested plasmids.
RESULTS
Selectionandcharacterizationof viruses witha duplication
of the BamHI-L sequence. pTK133 and pTK106 were
con-structedbyinsertingthe BamHI-Lfragment of either HSV-1(MP) or HSV-1(mP) DNAinto the single BgiII site of the cloned BamHI-Q fragment present in
pRB103 (Fig.
1). Insertions ofDNAatthisBgIII
site resultinanonfunctional tk gene (14, 16, 24, 34). After cotransfection of cells with HSV-1(mP) or HSV-1(MP) DNA and one ofthe plasmids shown in Fig. 1, recombinant virusescontaining
theappro-priate insertion in their tk genes were selected by their
resistanceto araT(17). AraT-resistant isolateswere plaque purified, and the DNA was extracted from cytoplasmic fractionsof the infected cells. Mostof theviral DNA in the
cytoplasmic fractionswas packagedin progeny virions. DNAsfrom mostofthe araT-resistantplaqueisolates we
analyzed were shown by Southern blot analysis tocontain
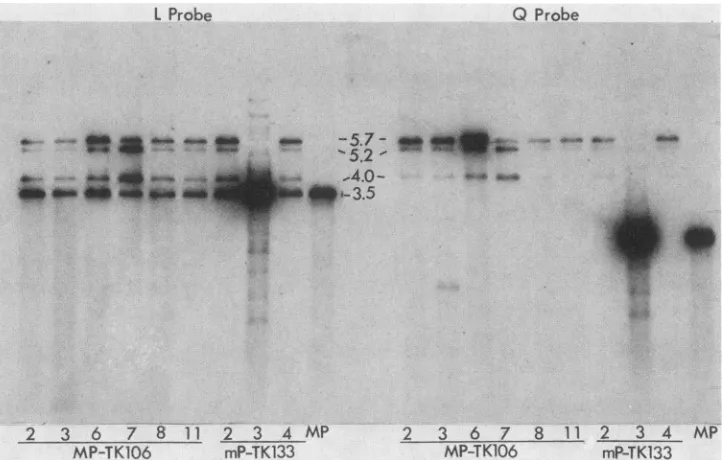
the BamHI-L sequence inserted into the tk gene. The genomes ofeight of the viruses analyzed (Fig. 2) were all
missing the BamHI-Q fragment (2.2 x
106
Da); theycon-tained insteada new
fragment
(5.7x 106 Da)thathybridizedwith both the BamHI-Lprobe and theBamHI-Q or tk probe and was the size predicted for the new BamHI fragment generatedby theinsertion. All viruses(includingthe parent MPstrain)hadabandat3.5 x 106 Da,whichisthe BamHI-L
fragment atits normal locationon the genome.
Two novel fragments were detected in BamHI digests of DNA from the insertion mutants. The amounts of these fragments relativetothosepredicteddiffered frommutantto mutant. The novelfragments, 5.2 x
106
and 4.0 x106
Da, hybridized to both the BamHI-Q and BamHI-L probes.1-t-VOL. 53, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.129.485.76.266.2]458 POGUE-GEILE ET AL.
L Probe Q Probe
2 3 6 7 8 11 2 3 4 MP 2 3 6 7 8 11 2 3 4 MP
[image:3.612.132.493.75.305.2]MP-TK106 mP-TK133 MP-TK106 mP-TK133
FIG. 2. Analysis ofgenome structuresof araT-resistantprogenyobtainedfrom cells cotransfected withHSV-1(MP)DNAandpTK106or
withHSV-1(mP)DNAand pTK133.BamHIrestriction fragments of virion DNA fromHSV-1(MP) and from nine araT-resistant isolateswere electrophoresedon a0.7%agarosegel and transferredtonitrocellulose. Thefragments containing sequenceshomologoustothe BamHI-L sequence(Lprobe) and theBamHI-Qsequencecontaining the tkgene(Q probe)wereidentifiedbyhybridization. The sizes(106Da)of the fragments detectedareindicated.
Their sizes were as predicted for fragments from genomes
resulting fromtherecombinationaleventsdepictedinFig.3.
Genomes with an inversion ofthe DNA sequence between the duplicated BamHI-L sequences (product 5) could be
generated by intramolecular recombination or bya double-crossover intermolecular recombination event as shown.
Many of the defective products of intermolecular
recombi-nation (products 1 through 4) could theoretically be pack-aged into virions. These products were approximately ge-nomelength (since thetwocopies of theBamHI-L sequence
in the insertion mutants were approximately equidistant from the ends ofthegenome), andthey containedcleavage andpackagingsignals that have been showntobepresent in therepetitivea sequencelocated atboth ends of thegenome and at the L-Sjunction (33;N. Frenkel, L. P.Deiss,and R. R. Spaete, Abstr. Herpesvirus 1984, no.
1357,
p. 198).To test the hypothesis that the anomalous fragments
containing
sequences thathybridized
to boththeBamHI-Land BamHI-Q fragments were from recombinant genomes (Fig. 3), we digested DNA from the progeny of two insertion mutants withSail. This enzymedoes notcleavewithin the
BamHI-LorBamHI-Qsequence and generatesfragmentsin
whichsequences contiguousto theanomalousBamHI
frag-ments can be identified. Figure 4 shows the two separate
regions of the genome in which BamHI-L sequences were present in opposite orientations in the insertion mutants.
Alsoshown is thechangeincontiguousDNA sequences that
would result from recombination between thetwoBamHI-L
sequences. The sizes of theSall fragmentscharacteristic of each BamHI-L locus in parental orrecombinant form and theprobestowhichtheywouldhybridize are also indicated.
All four of the SalI fragments depicted in Fig. 4 were detected (Fig. 5), providing evidence for the predicted
recombinational events.
DNAsfrom theinsertionmutants werealsodigestedwith EcoRI or HpaI. Both of these enzymes cut within the
PARENTAL MOLECULES PRODUCTS
Inverted.Repeats a b b'a'
Intermolecular Recombinations 2)a}aa
ab b'a'ac' ca single b
crossove
b'a,
| bIntramolecular Recombination
ab
z:j
a'c' caa b X
[= ET=
~~~~~~~~~~~~~~-a b ba'a'c' ca
5)1 - - 2 EZIJZJ m
FIG. 3. Results of intermolecular andintramolecularrecombination between the two BamHI-L sequences present at differentlociin the insertionmutants. Left, Genome structures ofthe mutantsresulting from insertion of theBamHI-L sequenceinto thetkgeneinopposite orientationtotheresidentBamHI-L sequences. Symbols: ,repetitive sequences(superscript letters refertothe samestrand ofDNA withinamolecule: a, b, and c are sequences homologous to a', b', and c', respectively); EZZ,BamHI-L sequence (orientation is shown by thedirection of thecross-hatching);X,recombination sitesgenerating the products shown on the right. The molecules are drawn to scale with respect to theparticular featuresidentified, except that a portion of each genome has been omitted from most of these representations where indicatedby breaks between parallel diagonallines.
J. VIROL.
.;_,.awl* or--5.7-
W
1:
.::. "'5.21
,4.o- #aft
l*,',.-3.5
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.95.529.555.664.2]PARENTAL FRAGMENTS
0( L 0(r) D L 8(1)
7.8 4.8 t
NOVEL FRAGMENTS
0() L Do 0(r) L B(M)
t
~~6.5 6JFIG. 4. Restriction fragments generated by Sall cleavages of rearranged ornonrearrangedgenomesfrom the insertion mutants. Top, Parental Sall fragments that will hybridize to a BamHI-L probe ifno recombination has taken place betweenthe BamHI-L sequences atthetwodifferent loci. Bottom, Novel Sallfragments that willhybridizeto aBamHI-L probe ifrecombination has taken place between thetwoBamHI-L sequences atthedifferentloci. The verticalarrowsshow the positions of theSall recognitionsequence; numbers between the vertical arrows indicate the sizes of Sall fragments(106Da). Horizontal arrowsindicate that theBamHI-L sequences ( E ) areininverted orientation. The letters above the lines are the names of BamHI fragments from HSV-1 DNA (or portions thereof) usedasprobes in the Southern blotanalysis (Fig. 5). Symbols: M,the HSV-1 DNA sequence contained in probe Q(l) or plasmid pKPG103L; MgE, the HSV-1 DNA sequence in probeQ(r)orplasmid pKPG103R; C,theHSV-1DNA sequence of the BamHI-D' fragmentpresent in pRB142(mapunits 0.700to 0.706); _, the HSV-1DNA sequence,designated B(l), contained inpKPG102.
BamHI-Lsequence so that no restriction fragments specific to the recombinant genomes would be generated. As pre-dicted, these restriction digests showed no novel bands (data not shown).
DISCUSSION
Evidence hasbeenpresented that the normalinversionof
the UL and Us components ofwild-type HSV genomes is due to site-specific recombination between inverted copies ofthe reiterateda sequences and is mediatedby viral gene products (3, 4, 17-21, 30). The evidencefor this site
speci-ficity
was obtained by inserting fragments from either theunique or the reiterated regions ofthe genome into the tk geneoftheHSVgenome (17). Insertions ofDNAfromthe
reiteratedregions ofthegenome resulted in novelinversions between inverted copies of the reiterated sequences. In contrast, insertions ofthree different Sacl fragments from
UL (map
units0.64to0.65)
orUs (map
units0.910to0.925 or0.925 to0.935) didnotresult in anynovelDNA rearrange-ments that could be detected in cytoplasmic packaged ge-nomes.Two other HSV DNAfragments from unique regions of
the genome, in addition to the BamHI-L fragment, have been insertedintothe tk geneofHSV. Noevidence forDNA rearrangements was obtained when a
SalI-to-BamHI
frag-ment containing the genefor gC-1 (map units 0.62 to 0.64) was inserted in either orientation into the HSV-1(MP) ge-nome (14). In contrast, when the homologous HSV-2 se-quence encoding gC-2 (map units 0.62 to 0.64) was inserted into the HSV-1(MP) genome in inverted orientation to thegC-1gene, one of the two novel restrictionfragments which
would be predicted by inversion was detected in a
BamHI
digest
ofpackaged viralDNA(K. M.Zezulak,Ph.D. thesis,University of Chicago, Chicago, Ill., 1983). Perhaps the second novelfragmentwasnotdetected because its size was very similar to that of aBamHIfragment fromthe parental viral DNA. Thus, it is possible that inverted duplication of
the sequence encoding gC-2 as well as the BamHI-L se-quence results in DNA rearrangements.
Several factors other than the actualfrequency of recom-bination between the duplicated sequences could influence theability to detect rearranged genomes, and these factors couldhelp to explain the different consequences of inserting the BamHI-L sequence and other unique sequences. In all the experiments described here and elsewhere, the DNA that was analyzed was DNA packaged in virions. Thus, detection of recombination products can be limited by the
requirements for packaging.Theserequirementsinclude the presence of the a sequence on at least one end of the genome (33; N. Frenkeletal., Herpesvirus 1984)and anappropriate
genomesize. Although the range ofpermissible sizes is not known withcertainty,mutantscontaining deletions or
inser-tions that are greater than about 8% of the genome length have not yetbeenreported. The detectionof recombination
products may also depend in part on the ability ofvirions
containing rearrangedgenomes toreplicate. Defective rear-ranged genomes would not be amplified during successive roundsofreplication except under conditions ofhigh
multi-plicity of infection.
It seems unlikely, however, that packaging constraints
alone canexplain the different consequences of duplicating
various sequences from unique regions of the genome. Intramolecular recombination between duplicated se-quences in opposite orientations should always yield
packageable genomes. Recombinant products that might
have resulted from intermolecular recombination in this
study (i.e., products 1through4, Fig. 3) are an appropriate
L
B(l)
Q(r)
Q(O)
D-78 40 -6.5 6.1 MD*ft -4.8
s a
X 3 2 3 2 1 3 2 1 3 2 1 3 2 1
Hind)ll
FIG. 5. Sall restriction endonuclease digestion and Southern blotanalysis of DNA from the BamHI-L insertion mutants. This Southern blot includes Sall digests of DNA from HSV-1(MP) (sample 1)andfromtheinsertionmutantsMP-TK106-7 (sample 2) and mP-TK133-2(sample 3).Samplesof theseSall-digestedDNAs wereloaded into fourorfive wells,and theappropriatelanes were hybridizedwith the labeledprobesindicated at the topofeachpanel (as described in the legend to Fig. 4). The first panel shows
HindIll-digestedXDNAhybridizedwithlabeledX DNA. Thebands detected herewereusedassize markerstodetermine the sizes(106 Da) ofthe bands in theother.panelsasshownonthefarright.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.296.78.169.2] [image:4.612.322.553.395.627.2]460 POGUE-GEILE ET AL.
sizeforpackagingonly because the tk gene and the normal genomic location of the BamHI-L sequence happen to be equidistant from the ends of the genome. This is probably also true for at least two of the other insertion mutantsfor which no evidence ofgenomic rearrangement was obtained. These insertion mutants include those described by Lee et al. (14)and Mocarski et al. (17), which had sequencesfrom
map units 0.62 and 0.64 and 0.64 to 0.65, respectively,
duplicated in the tk gene.
The possible defectiveness of rearranged genomes also does not fully explain the different consequences of
dupli-cating BamHI-L sequences and other unique sequences.
Smileyet al. (30) detected packaged molecules fromwhich half ofthe genomehad been deleted due to recombination
between twocopiesof the a sequence in direct orientation in
infectionsatrelatively lowmultiplicity (1 PFU per cell). The deletedgenomes were an appropriate size to be packaged as dimers.
It seems likely, therefore, that an unusually high rate of
recombination between theduplicatedBamHI-L sequences
isatleast partlyresponsible for the resultsreported here. It
ispossible thata site-specific mechanism is responsible for
theserearrangements, because similarrearrangements have not been detected in HSV genomes with duplications of otheruniquesequences. This argument was used
previously
to propose the existence of a site-specific mechanism for recombination between a sequences (17). If both of these genome regions contain signals for site-specific recombina-tion, then presumably the signals arefordifferent systems; or, if thesame system isused, itdoes notpromote
recom-bination between thea andthe BamHI-L sequences.
Alter-natively, the recombinational events we observed may
re-quire
homology butnot aspecific
nucleotide sequence, withthe frequency of recombination between homologous se-quences depending onfactors not yetdefined.
ACKNOWLEDGMENTS
WethankB.Roizman forprovidingtwoof theplasmids,R.Aspell forgrowing and maintaining cell lines, and J. Hoshizaki fortyping themanuscript.
This workwassupportedby grantstoP.G.S. fromtheAmerican Cancer Society and the Public Health Service (CA21776 and CA19264).K.L.P.-G. isatrainee under National Research Service Award 5 T32PHS GM07281.
LITERATURE CITED
1. Bolivar, F.,R.L.Rodriguez,P.J. Greene,M.C.Betlach,H.L. Heynecker, H. W. Boyer, J. H. Crosa, and S. Falkow. 1977. Construction and characterization ofnewcloningvehicles. II.A multipurposecloningsystem. Gene 2:95-113.
2. Bond,V. C.,and S. Person. 1984. Fine structurephysical map locations of alterationsthataffect cell fusion inherpessimplex virus type1. Virology 132:368-376.
3. Davidson, A.J., H. S.Marsden,and N. M. Wilkie. 1981. One functionalcopyof thelong terminal repeat gene specifyingthe immediate-early polypeptide IE110 suffices aproductive infec-tion ofhumanfoetallung cellsbyherpessimplex virus.J. Gen. Virol. 55:179-191.
4. Davidson, A.J.,and N. M.Wilkie. 1981. Nucleotide sequences ofthejoint between the Land S segments ofherpes simplex virus types 1 and2.J. Gen. Virol. 55:315-333.
5. Delius, H.,andJ.B.Clements. 1976. Apartial denaturation map ofherpessimplex virus type1DNA:evidence forinversionsof uniqueDNAregions.J. Gen. Virol. 33:125-133.
6. Ejercito,P.M.,E. D.Kieff,and B. Roizman. 1968. Characteri-zationofherpessimplexvirusstrainsdifferingin theireffecton social behavior of infected cells.J.Gen. Virol. 3:357-364. 7. Gibson, M. G., and P. G. Spear. 1983. Insertion mutants of
herpes simplexvirus have aduplication oftheglycoprotein D
gene and expresstwodifferent formsofglycoproteinD.J.Virol. 48:396404.
8. Graham,F.L.,and A.J.Vander Eb.1973. Anewtechniquefor the assayof infectivityof human adenovirus DNA. Virology 52:456-467.
9. Guerry, P.,D.J.LeBlanc,and S. Falkow.1973.General method for theisolation ofplasmiddeoxyribonucleicacid. J. Bacteriol. 116:1064-1066.
10. Hayward, G. S.,R.J.Jacob,S.C. Wadsworth,and B. Roizman. 1975.AnatomyofherpessimplexvirusDNA:evidence for four populationsof molecules thatdiffer in the relative orientations of theirlong and short segments. Proc. Natl. Acad. Sci. U.S.A. 72:4243-4247.
11. Heine, J. W.,R. W. Honess, E. Cassai,and B. Roizman.1974. Proteins specified by herpes simplex virus. XII. The virion polypeptides of typeI strains. J. Virol. 14:640-651.
12. Hoggan, M. D., and B. Roizman. 1959. The isolation and propertiesofavariant ofherpessimplexproducing multinucle-ated giant cells in monolayer cultures in the presence of antibody.Am.J.Hyg. 70:208-219.
13. Knipe,D.M.,W. T.Ruyechan,R. W.Honess,and B. Roizman. 1979.Moleculargeneticsofherpessimplexvirus: the terminala sequencesof theLand S componentsareobligatorilyidentical and constitute a part ofa structural gene mapping predomi-nantly in the S component. Proc. Natl. Acad. Sci. U.S.A. 76:4534-4538.
14. Lee, G. T.-Y.,K. L. Pogue-Geile,L. Pereira,and P.G. Spear. 1982. Expressionofherpessimplex virusglycoproteinC froma DNAfragment inserted into the thymidinekinase gene of this virus. Proc. Natl.Acad. Sci. U.S.A. 79:6612-6616.
15. Maniatis, T., E. F. Fritsch, andJ. Sambrook. 1982. Molecular cloning: alaboratorymanual.ColdSpringHarborLaboratory, ColdSpring Harbor,N.Y.
16. McKnight, S. L. 1980. Thenucleotide sequence andtranscript map of the herpes simplex thymidine kinase gene. Nucleic AcidsRes. 8:5949-5964.
17. Mocarski, E.S., L. E. Post, and B. Roizman. 1980. Molecular engineeringof theherpes simplexvirusgenome: insertion ofa
second L-Sjunctioninto thegenomecausesadditional genome inversions. Cell22:243-255.
18. Mocarski,E. S., and B. Roizman. 1981. Site-specificinversion sequence of the herpes simplex virus genome: domain and structural features.Proc.Natl.Acad. Sci.U.S.A. 78:7047-7051. 19. Mocarski,E. S.,and B. Roizman. 1982.Herpesvirus-dependent amplification and inversion of cell-associated viral thymidine kinase gene flankedbyviralasequences and linkedto anorigin of viral DNA replication. Proc. Natl. Acad. Sci. U.S.A. 79:5626-5630.
20. Mocarski,E.S.,andB. Roizman.1982.Structure and role of the herpessimplexvirus DNAtermini ininversion, circularization andgenerationof virionDNA.Cell 31:89-97.
21. Poffenberger, K.L., E. Tabares,and B. Roizman. 1983. Char-acterization ofa viable, noninverting herpes simplex virus 1 genomederived by insertion and deletion of sequences at the junctionof componentsLandS. Proc.Natl. Acad. Sci. U.S.A. 80:2690-2694.
22. Pogue-Geile, K. L., G. T.-Y. Lee, S. K. Shapira, and P. G. Spear. 1984. Fine mappingofmutations in thefusion-inducing
MPstrainofherpessimplexvirustype1.Virology136:100-109. 23. Post,L.E.,A.J.Conley,E.S.Mocarski,and B. Roizman. 1980. Cloningofthereiterated andnonreiteratedherpessimplexvirus 1 sequences as BamHI fragments. Proc. Natl. Acad. Sci. U.S.A. 77:4201 4205.
24. Post, L. E.,S. Mackem, and B. Roizman. 1981. Regulationof alpha genes ofherpes simplex virus: expression of chimeric genesproduced byfusionofthymidinekinase withalphagene promoters. Cell24:555-565.
25. Radloff, R.,and W. Bauer.1967. Adye-buoyant-densitymethod forthedetection and isolation of closed circularduplexDNA: theclosedcircularDNA in HeLacells. Proc. Natl. Acad. Sci. U.S.A.57:1514-1521.
26. Roizman, B. 1979. The structure and isomerization ofherpes simplexvirusgenomes. Cell 16:481494.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
27. Ruyechan, W.T., L. S. Morse, D. M.Knipe, and B. Roizman. 1979.Molecular geneticsofherpes simplex virus.II.Mapping of
themajorviral glycoproteins andof geneticloci specifying the social behavior ofinfectedcells. J. Virol. 29:677-697. 28. Sheldrick,P., and N. Berthelot. 1974. Invertedrepetitions inthe
chromosome of herpes simplex virus. Cold Spring Harbor Symp. Quant. Biol. 39:667-678.
29. Skare, J., and W. C. Summers. 1977. Structureandfunction of herpesvirusgenomes. II.EcoRI,XbaI andHindIlI
endonucle-asecleavage sitesonherpessimplex virustype 1 DNA.
Virol-ogy76:581-595.
30. Smiley,J. R., B. S. Fong, and W.-C.Leung. 1981.Construction ofa double-jointed herpes simplex viral DNA molecule:
in-verted repeats are required for segment inversion and direct
repeatspromotedeletions. Virology 113:345-362.
31. Southern, E. M. 1975. Detectionofspecific sequences among
DNAfragmentsseparatedby gelelectrophoresis. J. Mol. Biol.
98:503-517.
32. Varmuza, S. L., and J. R.Smiley.1984. Unstableheterozygosity in a diploid region of herpes simplex virus DNA. J. Virol.
49:356-362.
33. Vlazny, D. A., A. Kwong, and N. Frenkel. 1982. Site-specific
cleavage/packaging of herpes simplex virus DNA and the selec-tive maturation of nucleocapsids containing full-length viral DNA. Proc. Nati.Acad. Sci. U.S.A. 79:1423-1427.
34. Wagner, M. J., J. A. Sharp, and W. C. Summers. 1981. Nucleotide sequence ofthe thymidine kinase gene ofherpes
simplex virus type 1. Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445.
35. Walboomers, J. M. M., and J.terScheggt. 1976. Anewmethod
for theisolation ofherpes simplex virustype2 DNA. Virology 74:256-258.
36. Wigler,M.,A.Pellicer, S. Silverstein,R. Axel,G.Urlaub, and L. Chasin.1979. DNA-mediated transfer of the adenine phospho-ribosyltransferaselocus into mammalian cells. Proc. Natl. Acad. Sci. U.S.A. 76:1373-1376.
37. Wilkie, N. M. 1976.Physicalmapsforherpes simplexvirustype
1 DNAfor restriction endonucleases Hind III, Hpa-1, and X. bad. J. Virol. 20:222-233.
38. Zezulak, K.M., and P. G. Spear. 1984. Mapping of the
struc-turalgene for the herpes simplex virus type 2 counterpart of herpessimplexvirustype 1glycoprotein C and identification of atype2 mutant which does not express this glycoprotein. J. Virol.49:741-747.