Sry
Expression Level and Protein Isoform Differences Play a Role in Abnormal
Testis Development in C57BL/6J Mice Carrying Certain
Sry
Alleles
Kenneth H. Albrecht,
1Maureen Young, Linda L. Washburn and Eva M. Eicher
2The Jackson Laboratory, Bar Harbor, Maine 04609
Manuscript received October 7, 2002 Accepted for publication January 28, 2003
ABSTRACT
Transfer of certain Mus domesticus-derived Y chromosomes (SryDOMalleles, e.g.,SryPOSand SryAKR) onto
the C57BL/6J (B6) mouse strain causes abnormal gonad development due to an aberrant interaction between theSryDOMallele and the B6-derived autosomal (tda) genes. For example, B6 XYPOSfetuses develop
ovaries and ovotestes and B6 XYAKRfetuses have delayed testis cord development. To test whether abnormal
testis development is caused by insufficientSryDOMexpression, two approaches were used. First, gonad
development and relativeSryexpression levels were examined in fetal gonads from two strains of B6 mice that contained a singleM. domesticus-derivedanda singleM. musculus-derivedSryallele (B6-YPOS,RIIIand
B6-YAKR,RIII). In both cases, presence of theM. musculus SryRIIIallele corrected abnormal testis development.
On the B6 background,SryPOSwas expressed at about half the level ofSryRIIIwhereasSryAKRandSryRIIIwere
equally expressed. On an F1 hybrid background, bothSryPOS andSryRIII expression increased, but SryPOS
expression increased to a greater extent. Second, sexual development andSry expression levels were determined in XX mice carrying a transgene expressingSryPOScontrolled by POS-derivedorMUS-derived
regulatory regions. In both cases one B6 transgenic line was recovered in which XX transgenic mice developed only testicular tissue but cord development was delayed despite normal Sry transcriptional initiation and overexpression. For three transgenes where B6 XX transgenic mice developed as females, hermaphrodites, or males, the percentage of XX transgenic males increased on an F1background. For
the one transgene examined, Sryexpression increased on an F1background. These results support a
model in which delayed testis development is caused by the presence of particular DOM SRY protein isoforms and this, combined with insufficientSryexpression, causes sex reversal. These results also indicate that at least onetdagene regulatesSryexpression, possibly by directly binding toSryregulatory regions.
N
ORMALLY in mammals, XX individuals develop as domesticus-derivedSrygenes are transferred onto specific inbred strains, such as C57BL/6J (B6).females with ovaries, and XY individuals develop as
males with testes. Although rare, complete sex reversal Standard mouse inbred strains are a composite of two species,M. musculusandM. domesticus(reviewed in (SR) occurs in which XX individuals develop testes and
XY individuals develop ovaries. In humans the easiest SR Bonhomme1986). Most strains, such as B6, contain a M. musculus(MUS) Y chromosome, but a few, such as cases to explain are XY females who carry a
nonfunc-tional SRY (sex-determining region, Y chromosome, AKR/J, contain aM. domesticus(DOM) Y chromosome (Bishop et al. 1985; Nishioka and Lamothe 1987; symbolized as Sry in mice) gene and XX males who
Tuckeret al.1992). Of interest to gonadal sex determi-carry a normal SRY gene located on their paternally
nation is the finding that transfer of some DOM Y chro-derived X chromosome due to an abnormal meiotic
mosomes (i.e., Sry alleles) to the B6 strain interferes recombination event (reviewed inSchafer1995;
McEl-with testis development (Table 1). For example, B6 reaveyandFellous1999). Several intriguing but
unex-XYPOSmice carrying an SryDOM allele from wild-derived
plained SR conditions occur, however, including XY
M. d. poschiavinus mice develop ovaries and ovotestes females and XY hermaphrodites who carry an
appar-but not normal testes (referred to as B6-YPOSsex reversal;
ently normal SRY gene and XY females who carry a
Eicheret al. 1982;Eicher andWashburn 2001). B6 mutatedSRYgene inherited from their father who
car-XYAKR mice carrying an SryDOM allele from the AKR/J
ried the same mutatedSRYallele (reviewed inSchafer
inbred strain have delayed testis cord development but 1995;McElreaveyandFellous1999). These cases are
do not develop ovarian tissue (WashburnandEicher reminiscent of what occurs in mice when certainMus
1983, 1989;EicherandWashburn1986;Nagamineet al.1987). OtherSryDOMalleles do not interfere with
nor-mal testis development when present on B6, an example
1Present address:Genetics Program, Department of Medicine, Boston
being B6 XYBUBmice (Sryallele from the BUB/BnJ
in-University Medical School, 715 Albany St., E325, Boston, MA 02118.
bred strain;Washburnet al.2001). These phenomena
2Corresponding author:The Jackson Laboratory, 600 Main St., Bar
Harbor, ME 04609. E-mail: [email protected] are highly sensitive to genetic background. That is, an
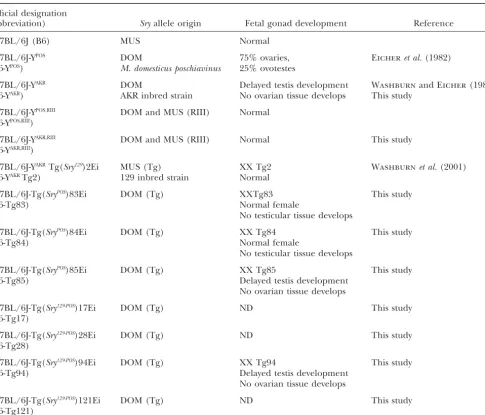
TABLE 1
Mouse strains, origin ofSryallele, and gonadal phenotype
Official designation
(abbreviation) Sryallele origin Fetal gonad development Reference
C57BL/6J (B6) MUS Normal
C57BL/6J-YPOS DOM 75% ovaries, Eicheret al.(1982)
(B6-YPOS) M. domesticus poschiavinus 25% ovotestes
C57BL/6J-YAKR DOM Delayed testis development WashburnandEicher(1983)
(B6-YAKR) AKR inbred strain No ovarian tissue develops This study
C57BL/6J-YPOS,RIII DOM and MUS (RIII) Normal
(B6-YPOS,RIII)
C57BL/6J-YAKR,RIII DOM and MUS (RIII) Normal This study
(B6-YAKR,RIII)
C57BL/6J-YAKRTg(Sry129)2Ei MUS (Tg) XX Tg2 Washburnet al.(2001)
(B6-YAKRTg2) 129 inbred strain Normal
C57BL/6J-Tg(SryPOS)83Ei DOM (Tg) XXTg83 This study
(B6-Tg83) Normal female
No testicular tissue develops
C57BL/6J-Tg(SryPOS)84Ei DOM (Tg) XX Tg84 This study
(B6-Tg84) Normal female
No testicular tissue develops
C57BL/6J-Tg(SryPOS)85Ei DOM (Tg) XX Tg85 This study
(B6-Tg85) Delayed testis development
No ovarian tissue develops
C57BL/6J-Tg(Sry129-POS)17Ei DOM (Tg) ND This study
(B6-Tg17)
C57BL/6J-Tg(Sry129-POS)28Ei DOM (Tg) ND This study
(B6-Tg28)
C57BL/6J-Tg(Sry129-POS)94Ei DOM (Tg) XX Tg94 This study
(B6-Tg94) Delayed testis development
No ovarian tissue develops
C57BL/6J-Tg(Sry129-POS)121Ei DOM (Tg) ND This study
(B6-Tg121)
ND, not determined.
SryDOM allele that causes abnormal testicular develop- quence analysis of several DOMSryalleles revealed that
this hypothesis alone is inadequate because no correla-ment in B6 mice may not do so when present on another
genetic background (Eicher et al. 1982; Eicher and tion between the open reading frame (ORF) sequence and SR was found (Carlisleet al.1996;Albrechtand Washburn 1986; Nagamine et al. 1987; Biddle and
Nishioka 1988). For example, DBA/2J XYPOS (D2 Eicher1997). A second possible mechanism to explain
B6-YPOSSR is that expression of theSryPOSallele is aberrant.
XYPOS) and (D2 ⫻ B6)F
1 XYPOS mice develop normal
testes. These findings suggest that the ability of a specific Support for this hypothesis was obtained byNagamineet al. (1999), who assayedSryexpression in mouse fetal go-Sry allele to function correctly is dependent on other
genes, designated tda (testis-determining autosomal) nads from three B6 consomic strains carrying different DOMSryalleles,SryFVB,SryAKR, andSryTIR. [Sequence
anal-genes, and genetic mapping experiments have
sup-ported this hypothesis (Eicheret al.1996). If the proper ysis indicates that theSryTIRandSryPOSORFs are identical
(Albrecht and Eicher 1997)]. They found that Sry functioning of the humanSRYgene likewise is sensitive
to genetic background, this could clarify the unex- expression was highest in B6 XYFVBfetal gonads, which
develop as normal testes, and lowest in B6 XYTIR fetal
plained human XY SR conditions noted above.
The simplest mechanism to explain B6-YPOSSR is that gonads, which develop as ovotestes or ovaries. Moreover, Sryexpression was increased in (SWR⫻B6)F1XYTIRfetal
theSryPOSallele encodes a protein that does not interact
correctly with downstream genes if they are derived from gonads, which develop as normal testes.
se-inbred strain.) TheSxr Y chromosome carries a duplication
proaches designed to further our understanding ofSry
of most of the short arm (Yp), including theSrygene, at the
function in B6-YPOSSR. First, we determinedSry
expres-distal end of the long arm (Yq; Evans et al. 1982; Bishop
sion levels in fetal gonads from B6 mice carrying a Y et al. 1988). This duplicated copy of Sry is adjacent to the chromosome containingboth a MUS-derived Sry allele pseudoautosomal region and can be transferred to the X
chro-mosome by homologous recombination.
(SryRIII) and a DOM-derivedSry allele (SryPOSorSryAKR).
The basic strategy to create theSrybiallelic strains was to
This approach differed from that used byNagamineet
first transfer the duplicatedSxrsegment from theSxrY
chro-al. (1999) because it directly comparedSryDOMandSryMUS
mosome to an X chromosome and then to transfer it from
expression within the same gonad so that the results the X chromosome onto the YPOSor YAKRchromosome. Because
were independent of the number ofSry-expressing cells. XXSxrmice are sterile, we used the T(16;X)16H translocation
(T16H) to cause preferential X inactivation of the XSxr
chromo-We found thatSryPOStranscript levels were significantly
some: If X inactivation spreads to theSrygene, these T16H/
reduced compared toSryRIII, whereasSryAKRandSryRIII
tran-Sxrmice will develop as females (Cattanachet al.1982;
Simp-script levels were equivalent. We then assayedSrytranscript
sonet al.1984). T16H⫹/⫹EdaTafemales were mated to XYSxr
levels on a (D2⫻B6)F1genetic background previously males (EdaTais ectodysplasin-A, also known as tabby).
Non-shown to allow normal testis determination in XYPOS
tabby females (i.e., inherited T16H) that inheritedSxron their paternally derived inactive X chromosome were mated to B6
mice (EicherandWashburn1986). Unexpectedly, we
XYPOS hermaphrodites or to B6 XYAKR males to generate
found that although both SryDOM and SryMUS transcript
XSxrYPOSor XSxrYAKR males, respectively. XSxrYPOSand XSxrYAKR
levels increased,SryDOMtranscript levels increased more
males were mated to B6 females, and XYPOS,Sxrand XYAKR,Sxr
thanSryMUStranscript levels. Together, these data suggest
male offspring, respectively, were identified (i.e., theSxr
seg-that B6 XYPOSSR is caused, at least in part, by insufficient
ment was transferred from the XSxrchromosome to the YPOS
or YAKR chromosome by homologous recombination in the Sryexpression and that one or moretdagenes directly
pseudoautosomal region). Thereafter, XYPOS,Sxr and XYAKR,Sxr
regulateSry expression. These results confirm and
ex-males were backcrossed to B6 feex-males. All ex-males used to
pro-tend those ofNagamine et al. (1999).
duce fetal offspring in this study were at backcross generation
Our second approach was based on the premise that N10 or greater. Hereafter, the C57BL/6J-XYPOS,Sxrand C57BL/
if B6-YPOS SR is caused by insufficient Sry expression,
6J-XYAKR,Sxrstrains are referred to as B6-YPOS,RIIIand B6-YAKR,RIII,
respectively, to reflect theSryalleles present.
then overexpression ofSryPOSwould rescue testis
devel-Srytransgene construction:TheSry129-POSchimeric transgene
opment in B6 mice. B6 transgenic mouse lines carrying
is based on a 14.6-kb MUS-derived genomic DNA fragment
either a chimericSryconstruct in whichSryPOSexpression
from a 129 inbred strain (Gubbayet al.1990) and previously
was regulated by MUS regulatory regions or an SryPOS
shown to sex reverse XX mice carrying it as a transgene (
Koop-genomic DNA clone were produced. In two transgenic manet al.1991;Eicheret al.1995; Figure 1). TheSry129open
reading frame and a portion of the 3⬘ untranslated region
lines, B6 XX mice carrying either type of transgene
were replaced by a homologousSryPOSDNA segment as follows:
developed testicular tissue exclusively. However, testis
The replacement fragment was PCR amplified using
high-cord development was delayed despite normal transcrip- fidelityPfuDNA polymerase (Stratagene, La Jolla, CA), geno-tional initiation and overexpression ofSry. These data mic DNA template containing the YPOSchromosome, and
prim-suggest that delayed testis cord development is caused ers Sry-8312 (5⬘-CCATGTCAAGCGCCCCATGAATGC) and
Sry-9816 (5⬘-AGCTGTTTGCTGTCTTTGTGCTAGCC). (Sry
by SRY protein isoform differences that are exacerbated
primers are designated by the 5⬘ base using numbering in
by insufficientSry expression leading to ovarian tissue
GenBank entry X67204.) The PCR product was gel purified
development in B6 XYPOSgonads. The above hypothesis
and cloned into pCR-Script using the manufacturer’s
proto-is supported by the finding that the MUSSryB6allele is
cols (Stratagene). The DNA sequence of individual clones was confirmed prior to further use. The replacement fragment
expressed at relatively low levels (LeeandTaketo2001)
was inserted into the 14.6-kbSry129genomic DNA clone at the
without causing delayed testis cord development or SR.
uniqueHhaI (base pair 8312) andSpeI (base pair 9704) sites.
In five Sry B6 transgenic lines, sex reversal was not (TheSryORF is located between base pair 8304 and base pair complete. In two lines, XX transgenic mice developed 9491.) The nucleotide sequences 8304–8312 and 9491–9704 as females, and in three lines XX transgenic mice devel- are identical inSryPOSandSry129(AlbrechtandEicher1997).
TheSryPOStransgene was derived from a 13.5-kb genomic
oped as females, hermaphrodites, or males. However,
DNA clone (L961) isolated from a mouse carrying a YPOS
chro-for the three transgenes tested, the percentage of XX
mosome (Gubbay et al.1992; Figure 1). Sequence analysis
transgenic males increased on a (D2⫻ B6)F1 genetic indicated that this clone extends from approximately base
background. In the one case assayed, transgene expres- pair 2355 to approximately base pair 15,825 relative to the 14.6-kbSry129clone described above and that a 36-bp deletion
sion was increased in F1gonads compared to B6 gonads,
was present in the glutamine repeat region downstream of
presumably due to the presence of an enhancer element
the DOM stop codon. The deleted region was replaced with
responsive to genetic background. a wild-typeSryPOSDNA fragment as follows: The replacement
fragment was PCR amplified usingPfuDNA polymerase, geno-mic DNA template containing the YPOSchromosome, and
prim-ers Sry-8386 (5⬘-CCAGCATGCAAAATACAGAGATCAGC)
MATERIALS AND METHODS
and Sry-9839 (5-ATGGCATGCTGTATTGACCACAAAGC).
Creating B6-YPOS,Sxrand B6-YAKR,Sxrconsomic strains:TheSxr The PCR product was digested with
SphI, gel purified, and (sex-reversed) Y chromosome rearrangement [formally, Tp(Y)- ligated into p961 (aNotI plasmid subclone of L961) digested 1Ct);Cattanachet al.1971] was used to generate mice bial- withSphI (base pair 8389 and base pair 9831 in GenBank entry lelic forSrycarrying a single copy of a MUS and DOMSry X67204). Correct orientation was confirmed by restriction
Figure 1.—Diagrammatic representation of theSrytransgenes. (A)Sry129. A 14.6-kb genomic
DNA clone isolated from the MUS 129 inbred strain. (B)SryPOS. A 13.5-kb genomic DNA clone
isolated from a strain carrying a DOM YPOS
chro-mosome. (C) Sry129-POS. A chimeric 14.6-kb
con-struct in which expression of the DOMSryPOSORF
is controlled by MUS regulatory regions derived from the clone in A. The HMG box DNA-binding domain, GRC, and the transcriptional start site at ⵑ8035 are indicated. MUS-derived regions are solid and DOM-derived regions are shaded. The inverted repeats are indicated by the double arrows. Numbering is based on GenBank entry X67204 and is approximate for B and C.
Srytransgenic mice:B6-Sry129-POSand B6-SryPOStransgenic mice CTA were used to amplify a 470-bp DNA fragment (Gubbay et al.1990) that was digested withMboI. Three fragments are were produced by micro-injecting the constructs described
above, without the plasmid backbone, into fertilized B6 eggs diagnostic forSryMUSalleles (199, 187, and 84 bp) and two are
diagnostic forSryDOMalleles (271 and 199;Eicheret al.1995)
using standard methods (Wagneret al.1981).
FourSry129-POStransgenic lines were recovered and formally and (2) primers Sry-9431 5⬘-TGGTGAGCATACACCATACC
and Sry-9808 5⬘-TTGCTGTCTTTGTGCTAGCC were used to
designated C57BL/6J-Tg(Sry-129-POS)17Ei, . . . 28Ei, . . . 94Ei,
and . . . 121Ei, hereafter referred to as Tg17, Tg28, Tg94, and amplify a 377-bp DNA fragment that, when digested with
NlaIV, produces a 377-bp undigested fragment diagnostic for Tg121. ThreeSryPOStransgenic founders were recovered and
are formally designated C57BL/6J-Tg(Sry-POS)83Ei, . . . 84Ei, SryMUS alleles and two comigrating fragments diagnostic for SryDOMalleles (189 and 188 bp). The Y chromosome was
de-and . . . 85Ei, hereafter referred to as Tg83, Tg84, de-and Tg85.
Transgenic line C57BL/6JEi-YAKRTg(Sry-129)2Ei (hereafter tected by multiplex PCR using primers for the YMT2/B locus
(5⬘-CTGGAGCTCTACAGTGATGA and 5⬘-CAGTTACCAAT
Tg2), carrying the original 14.6-kbSry129Tg (from which the
Sry129-POSTg was derived), was used as a control for some analy- CAACACATCAC) in conjunction with primers for the
autoso-mal myogenin gene (5⬘-TTACGTCCATCGTGGACAGCAT and
ses (Figure 1). All XX Tg2 animals present as males at weaning
(Washburnet al.2001). 5⬘-TGGGCTGGGTGTTAGTCTTAT) as a control (Capelet al.
1999).
Assessment of sexual phenotype in weaning-age mice:
Ani-mals were classified at weaning as female, male, or hermaphro- The Sry129-POS transgenes were detected by multiplex PCR
using the YMT/2B and myogenin primers described above in dite by the appearance of the external genitalia and by the
presence of yellow pigmented hairs associated with the mam- conjunction with transgene specific primers (5⬘-GAGGGCAT
GGTCAGTTGAAC and 5⬘-CTCAGTGTGGAATTCATCTGC;
mary glands. These pigmented hairs are present in B6 XX
females, absent in B6 XY males, and present in most B6 XYPOS Capelet al.1999). TheSryPOStransgene was detected by
multi-plex PCR as just described but using different transgene spe-hermaphrodites and in all B6 XYPOS females (Eicher and
Washburn2001). cific primers (5⬘-CTAATACGACTCACTATAGGGC and 5⬘-GCT GACTCCATGCACAGGC).
For histological analysis, gonads were dissected and fixed in
Bouin’s fixative, embedded in paraffin, sectioned, and stained Transgene copy number:Transgene copy number was deter-mined by semiquantitative PCR using B6 XYB6transgenic
geno-with hemotoxylin and eosin using standard procedures.
Fetal gonad analysis:Fetuses were collected from overnight mic DNA and the Sry-9431 and Sry-9808 primers. The assay was similar to that described below for semiquantitative RT-matings where noon on the day a vaginal plug was observed
is designated day 0.5 or from timed early morning matings. PCR except that 20 PCR cycles were employed. The results from at least three independent DNA samples were averaged. For more precise staging of fetuses younger than E13.0 (E,
embryonic day) the number of tail somites (ts) posterior to RT-PCR:Paired urogenital ridges or gonad/mesonephros complexes were dissected and nongonadal and nonmeso-the hind-limb bud was determined: E10.5 isⵑ8 ts, E11.5 is
ⵑ18 ts, and E12.5 isⵑ30 ts (Hackeret al.1995). E13.0–15.5 nephric tissues were removed. The mesonephros was trimmed to the length of the gonad. The gonad and mesonephros were fetuses were staged by fore-limb and hind-limb morphology
(Theiler1989). dissected apart in some later developmental stage samples. RNA was extracted from the dissected tissues using the RNeasy To assess fetal gonad development morphologically, gonads
with attached mesonephroi were dissected from E13.5–15.5 mini kit (QIAGEN, Chatsworth, CA). Lysed tissue was stored at⫺80⬚in RLT buffer (QIAGEN) until processed. The RNA fetuses and examined in whole mount using an inverted
micro-scope and transmitted light. This developmental stage was was DNased during isolation using an on-column protocol (QIAGEN) or after elution from the column using the DNA-chosen for analysis because a small amount of ovarian tissue
is easily visualized in an ovotestis and after this stage the rapid free protocol (Ambion, Austin, TX). After elution in 30l water, 2l of each RNA sample was tested for DNA contamination growth of testicular tissue can obscure detection of ovarian
tissue (Eicher et al.1980). A tissue sample was saved from by PCR amplification (35 cycles) using the 9431 and Sry-9808 primers. Any sample contaminated with DNA was re-each fetus for genotype determination, as described below.
Genotyping: PCR was used to detect the presence of an DNased, purified, and retested.
One-third of the RNA sample (10l) was reverse transcribed
SryMUSand/orSryDOMallele in genomic DNA using one of the
following methods: (1) Primers Sry-8207 5⬘-AGATCTTGATT at 42⬚ for 1 hr in a 20-l reaction using the RNA PCR kit (Applied Biosystems, Foster City, CA). Parallel reactions were
performed, one with reverse transcriptase (⫹RT) and one without (⫺RT). A no-template (H2O) negative control was
included in each experiment. The reverse transcription (RT) reaction (2l) was PCR amplified with primers specific for
theHprtgene (5⬘-CCTGCTGGATTACATTAAAGCACTG and
5⬘-GTCAAGGGCATATCCAACAACAAAC) as a positive
con-trol for the presence of intact RNA (Koopmanet al.1989). Semiquantitative RT-PCR was used to determine the relative expression of theSryMUS(SryRIII)vs.theSryDOM(SryPOSorSryAKR)
alleles (Bergstrom et al. 2000). The reverse transcription reaction (4l) was amplified by PCR in the presence of [␣
-32P]dCTP using the Sry-9431 and Sry-9808 primers and
restric-tion digested withNlaIV. The resulting fragments were sepa-rated on 2% agarose gels and Southern blotted using standard methods. The amount of radioactivity in each band was deter-mined using Phosphor imaging plates and Image Gauge soft-ware (Fuji Medical Systems USA, Stamford, CT).
Sryexpression levels were compared to the expression levels ofLhx1(LIMhomeoboxprotein1) using a semiquantitative RT-PCR assay.Lhx1was chosen as the control because it is expressed only in the mesonephric component of the genital ridge and expression is relatively constant during the develop-mental stages analyzed (Barneset al.1994;Fujiiet al.1994;
Nagamineet al.1999).Lhx1-specific primers were designed to amplify a 139-bp fragment that spanned a region with no
NlaIV restriction sites and contained a 97-bp intron (Lhx1-1660 5⬘-GGCGAGGAGCTCTACATCATAG and Lhx1-1798 5⬘
-CTTGGGAATCCGGAGATAAAC). The Lhx1 primers were
Figure2.—Transmitted light microscope images of E13.5– combined with Sry-9431 and Sry-9808 in a multiplex PCR
15.0 fetal B6 gonads and mesonephroi. Testis development reaction containing [␣-32P]dCTP and 2l of the RT reaction.
in B6 XYPOSfetuses is rescued by a single copy ofSryMUS. (A)
The PCR reaction was digested withNlaIV, separated on 3%
E14.5 XX ovary. Note the lack of cords and reticular appear-agarose gels, Southern blotted, and analyzed as outlined
ance. (B) E14.5 XYB6testis with testis cords present throughout.
above.
(C) E14.5 XYPOSovary, which looks similar to the XX ovary in
The number of PCR cycles corresponding to the
exponen-A. (D) E14.5 XYPOSovotestis. Two or three testis cords are present
tial amplification phase was determined empirically for each
in the center of the gonad and are flanked by ovarian tissue at RT-PCR assay (data not shown). Twenty-seven cycles were used
the cranial and caudal ends. (E) E13.5 XYPOS,RIIItestis. Even at
for theSry-only assay and 29 cycles were used for the multiplex
this earlier stage, testis cords are present throughout. (F)
Sry/Lhx1assays. PCR used 1.5 mmMgCl2and a 57⬚annealing
E14.5 XYPOS,RIII testis appears similar to the control XY testis
temperature.
in B. Testis development is normal in E14.5–15.0 XX Tg85
Statistical analysis:A two-way analysis of variance (ANOVA)
and XX Tg94 fetuses. (G) E14.5 XX Tg85 testis. (H) E15.0 was used to determine if there was a significant effect of fetal
XX Tg94 testis. Both gonads appear similar to the control XY age, genetic background, or interaction of these two variables
testis in B with testis cords present throughout. In A–H the onSry expression. Analyses were performed using
ln-trans-gonad is at the top and the mesonephros is below; cranial formed data to better meet the assumptions of ANOVA.
Schef-(anterior) is to the right and caudal (posterior) is to the left. fe´’s F was used for post-hoc multiple comparisons when the
ANOVA identified a significant effect. All effects were evalu-ated using␣ ⫽0.05.
ment in B6 XYPOSand B6 XYAKRmice. Previous
experi-ments demonstrated that the presence of a multi-copy MUS-derivedSry129transgene restored normal testis
de-RESULTS
velopment in B6-YPOSmice (Eicheret al.1995).
To assess if ovarian tissue development in B6 XYPOSmice
A single copy ofSryMUScorrects testis development in and delayed cord development in B6 XYAKR mice are
B6 XYPOS and B6 XYAKR mice: All B6 XYPOS,RIII and B6
caused by insufficientSryexpression, we developed two XYAKR,RIII mice presented as normal males at weaning. Srybiallelic B6 lines: One line carried a Y chromosome Moreover, gonad differentiation in both types of Sry containing the MUS-derivedSryRIIIallele and the
DOM-biallelic fetuses was normal at E13.5–15.5. The 16 B6 derivedSryPOSallele (B6-YPOS,RIII), and the other line
car-XYPOS,RIIIfetuses analyzed had two normal testes whereas
ried a Y chromosome containing the SryRIIIallele and
the 14 B6 XYPOScontrol sibs had ovaries (N⫽22 gonads)
the DOM-derivedSryAKRallele (B6-YRIII,AKR). We reasoned
or ovotestes (N⫽6 gonads; Figure 2). In addition, the that these B6 lines would allow a direct comparison of 19 B6 XYAKR,RIIIfetuses analyzed had two normal testes
the relative expression of twoSryalleles within the same whereas 10 of 11 B6 XYAKRcontrol sibs had testes with
gonad and therefore the results would be independent delayed cord differentiation. (One B6 XYAKR fetus had
develop-Figure4.—AverageSry/Lhx1expression levelvs. develop-mental age in B6 XYPOS,RIIIand (D2 ⫻B6)F
1 XYPOS,RIIIE11.5
urogenital ridges. Expression of bothSryPOSandSryRIIIincreases
in the F1 genetic background. As expected, Sry expression
peaks at 18–19 ts and then decreases. However, peak expres-sion occurs later in B6 urogenital ridges. For B6 XYPOS,RIII, two
samples were analyzed at 16 ts, three at 17 ts, five at 18 ts, three at 19 ts, and six at 20 ts. For (D2⫻B6)F1XYPOS,RIII, four
samples were analyzed at 16 ts, two at 17 ts, two at 18 ts, and
Figure3.—Analysis ofSryexpression inSrybiallelic urogeni- one at 19 ts. tal ridges. (A) Representative radioactive semiquantitative
RT-PCR results for E11.5SryPOS,RIIIandSryAKR,RIIIurogenital ridges.
Lanes labeled⫹and⫺represent samples with and without
(D2 ⫻ B6)F1 XYPOS fetuses develop testes (Eicher et
reverse transcriptase, respectively. Lhx1 expression also was assayed in these samples. Note thatSryPOStranscripts are
pres-al. 1982). If B6 XYPOS SR is caused by insufficient Sry
ent at lower levels than those of SryMUS, whereas SryAKR and
expression, expression of Sry should be increased in SryMUSare present at about equal levels. (B) Average expression
(D2⫻B6)F1XYPOSfetal gonads. We analyzed the relative
ofSryDOM/SryRIIIin E10.5–E13.0 urogenital ridges. The average
expression of SryPOSand SryRIIIin urogenital ridges dis-SryDOM:SryRIIIratio, number of samples analyzed (N), and
stan-dard error are indicated for each genotype. sected from E10.5–13.0 (D2⫻B6)F1XYPOS,RIIIfetuses. If
the expression ofSryPOSandSryRIIIwere increased
equiva-lently, the ratio of SryPOS:SryRIII would remain at 0.59.
However, if the expression of one allele was increased tion in B6 XYPOSmice and delayed testis cord
differentia-relative to the other, the ratio would change. We found tion in B6 XYAKR mice. This result also provided the
that the mean ratio (⫾95% confidence interval) ofSryPOS:
opportunity to perform Sry expression-level
experi-SryRIII
wasⵑ0.74⫾0.05 (Figure 3), indicating thatSryPOS
ments using B6 XYPOS,RIIIand B6 XYAKR,RIIIgonads to
exam-is expressed at a significantly lower level thanSryRIII. The
ine relativeSryexpression in gonads destined to develop
ANOVA indicates that this difference is constant from as normal testes.
E10.5 to E13.0 (P⫽0.958). However, the ANOVA also SryPOS
(DOM) transcript levels are reduced compared
indicates that the increased expression ofSryPOSon the to SryRIII (MUS) transcript levels between E10.5 and
F1genetic backgroundvs. the B6 background is signifi-E13.0:Semiquantitative RT-PCR was used to determine
cant (P⬍0.0004). We conclude that the expression of the relative expression of SryPOS vs. SryRIIIin urogenital
SryPOS is increased relative toSryRIIIon a hybrid genetic
ridges dissected from E10.5–13.0 B6 XYPOS,RIIIfetuses.Sry
background. expression normally is first detectable atⵑE10.5 (ⵑ8-ts
SryPOS expression is more sensitive thanSryRIII to ge-stage), peaks at ⵑE11.5 (18-ts stage), and is absent by
netic background:To determine if the expression level
ⵑE13.0 (Koopmanet al.1990;Hackeret al.1995;Jeske
of one or both Sry alleles is increased on the F1 back-et al. 1995). As indicated in Figure 3, the mean ratio
(⫾95% confidence interval) of SryPOS:SryRIII was 0.59 ⫾ ground, the expression level of each allele and Lhx1
were compared. The analysis was conducted using E11.5 0.04 during this time, indicating thatSryPOS
is expressed
at a significantly lower level thanSryRIII. Statistical analysis (16–20 ts) urogenital ridges because this is the timeSry
normally is maximally expressed. As indicated in Figure using ANOVA indicates that this difference is constant
between E10.5 and E13.0 (P ⫽ 0.958), implying that 4, expression of both SryPOS and SryRIII was increased
relative toLhx1in 16- to 18-ts gonads from F1XYPOS,RIII
during this time SryPOS and
SryRIII are temporally
regu-lated in a similar manner. We conclude that SryPOS is compared to gonads from B6 XYPOS,RIIIfetuses. The
AN-OVA indicates that the difference between the B6 and expressed at significantly reduced levels relative toSryRIII
in B6 mice. F1genetic backgrounds is significant (P⬍0.003). This
result, coupled with the finding that the ratio ofSryPOS:
Srytranscript level is affected by genetic background:
B6 XYPOSfetuses develop ovaries or ovotestes whereas SryRIIIwas increased to 0.74 in F
TABLE 2
SryPOSandSry129-POStransgenes and XX sex reversal
% XX Tg sex reversal (weaning)
F1
Tg copy Expression level
Transgene designation B6 (D2⫻B6) (C3H⫻B6) number (Tg:B6)
SryPOS
Tg85 100 (N⫽47) NA NA 17 3:1
Tg83 0 (N⫽84) 80 (2乆:8么) 83 (1乆:5么) 3 1:1
Tg84 0 (N⫽71) 0 (23乆:0么) 61 (7乆:11么) 9 2:1
Sry129-POS
Tg94 100 (N⫽33) NA ND 11 5:1
Tg17 56 (20乆:15 :45么) ND ND 12 ND
Tg28 9 (49乆:1 :4么) 53 (8乆:6 :3么) ND 14 ND
Tg121 18 (37乆:1 :7么) ND ND 8 ND
NA, not applicable because 100% of B6 XX Tg offspring are sex reversed. ND, not determined.
thatSryPOSis more sensitive than
SryRIIIto genetic
back-SryPOS transcripts are present at later developmental stages thanSryRIII:Previous results indicated that expres-ground. (These data also confirm thatSryPOSis expressed
at lower levels thanSryRIII.) Additionally, the data suggest sion ofSry persisted longer in B6 XYTIRgonads, which
develop abnormally, than in B6 XYB6 (Leeand Taketo
that peakSry expression occurs at an earlier
develop-mental stage (18 tsvs.19 ts, orⵑ2 hr) in the F1back- 1994) and B6 XYFVB(Nagamineet al.1999) gonads, which
develop normally. (YFVBis DOM derived.) However, in
ground. Whether this small difference in timing is
sig-nificant is unknown. an F1 hybrid background where XYTIR gonads develop
into normal testes,SryTIRexpression was not prolonged.
SryAKRandSryRIIIare expressed at equivalent levels in
E10.5–13.0 gonads:In contrast to B6 XYPOSgonads, B6 Between E10.5 and E13.0SryPOS is present atⵑ60% of
the SryRIII level in B6 XYPOS,RIII gonads. However, after
XYAKRgonads develop as normal testes, but have delayed
testis cord differentiation. The relative expression of E13.0, the situation is reversed and SryPOS is present at
higher levels thanSryRIII. For example,SryPOS,SryRIII, and SryAKRvs. SryRIIIwas determined by semiquantitative
RT-PCR using RNA from E10.5–13.0 B6 XYAKR,RIIIurogenital Lhx1 transcript levels were determined by RT-PCR in
seven E13.5 B6 XYPOS,RIII gonads with attached
meso-ridges. The mean ratio (⫾95% confidence interval) of SryAKR:
SryRIIIis 1.02⫾0.1 (Figure 3), indicating that
SryAKR nephroi (three pairs and four single complexes). The
averageSryPOS:
Lhx1ratio was 0.03 whereas the average andSryRIIIare expressed at equivalent levels. As indicated
by the ANOVA, the relative expression ratio was con- SryRIII:
Lhx1ratio was 0.007, indicating that at this stage SryPOSis present at about four times the level ofSryRIII.
stant throughout this time (P ⫽ 0.958). We conclude
thatSryAKRandSryRIIIare expressed at essentially equiva- These data suggest that expression of SryPOS persists
longer than expression ofSryRIIIin B6 XYPOS,RIIIgonads.
lent levels and are similarly regulated temporally. These
results suggest that delayed testis cord development in Transgenic overexpression ofSryPOSrescues testis de-termination:The comparativeSryexpression results sug-B6 XYAKRfetuses is not caused by insufficient or delayed
Sryexpression. gested that B6-YPOS SR is caused, at least in part, by
insufficient SryPOS expression. If this hypothesis is
cor-SryB6expression is lower than SryAKR expression:
Re-cent data indicated that SryB6 (MUS) is expressed at rect, transgenic overexpression ofSryPOSin B6 XX mice
should initiate normal testis determination. Two differ-lower levels thanSryAKR(DOM;LeeandTaketo2001).
We assayed the relative expression ofSryin E11.5 (18- ent SryPOS transgenic constructs were employed to test
this hypothesis (Figure 1). The first was a genomic DNA ts stage) B6 XYB6and B6 XYAKRgonads usingLhx1as a
control. The averageSry:Lhx1ratio was 0.95 in B6 XYAKR clone isolated from the M. d. poschiavinus Y
chromo-some. Analyses of three B6 transgenic lines (Tgs 83–85, gonads (N⫽5 gonad pairs) and 0.38 in B6 XYB6gonads
(N ⫽ 10 single gonads). In (D2 ⫻ B6)F1 XYB6gonads Table 2) carrying this construct (SryPOS) are presented.
The second construct was derived from the original (N ⫽16 single gonads) the average Sry:Lhx1ratio was
0.43. Thus, the expression ofSryB6(MUS), like that of 14.6-kb
Sryclone but contained the DOMSryPOSORF in
place of the MUSSry129ORF so that expression ofSryPOS SryRIII (MUS), is less sensitive to genetic background
than the expression ofSryPOS(DOM). Our results con- was controlled by MUS-derived regulatory regions.
Anal-yses of four B6 transgenic lines (Tgs 17, 28, 94, and 121, firm those ofLee andTaketo (2001) and emphasize
Figure5.—Both Tg85 and Tg94 are overexpressed relative to the endogenousSryB6allele in E11.5 urogenital ridges.
Rep-resentative radioactive semiquantitative RT-PCR results. Lanes labeled⫹and⫺represent samples with and without reverse transcriptase, respectively. Tg85 is expressed three times and Tg94 five times greater than the endogenousSryB6allele.
At weaning, 75% of B6 XYPOSmice present as normal
females and 25% present as hermaphrodites (Eicher andWashburn 2001). In contrast, for both the SryPOS
andSry129-POS constructs, one transgenic line from each
Figure6.—Transmitted light microscope images of E13.5
was identified in which all XX transgenic (XX Tg) mice
fetal B6 gonads. Testis cord differentiation is delayed in XX
presented as normal males at weaning (SryPOS
-Tg85 and Tg85 (SryPOS) and XX Tg94 (Sry129-POS) gonads. (A) XX ovary. Sry129-POS-Tg94). Histological examination of testes from
(B) XYB6testis. (C) XYAKRtestis. Note that the caudal pole is
three XX Tg85 and seven XX Tg94 adult males demon- undifferentiated and lacks testis cords (arrow). (D) XX Tg94 testis. (E) XX Tg85 testis. Both types of XXSryPOStransgenic
strated the presence of Sertoli and Leydig cells, lack
testes appear similar to the XYAKR testis with delayed cord
of ovarian tissue, and absence of germ cells (data not
differentiation at the caudal pole (arrows). (F) XX Tg2 testis,
shown). (The absence of germ cells is expected in XX which appears similar to the control XY testis in B with testis males due to the lack of a Y chromosome and presence cords present throughout. This result indicates that the
14.6-of two X chromosomes.) kb transgenic construct, which shares regulatory regions with
Tg94, contains the regulatory elements necessary for normal
We then examined gonads from B6 XX Tg85 and
testis differentiation. In A–F the gonad is at the top and the
XX Tg94 E14.5–15.5 fetuses to determine if ovarian
mesonephros is below; cranial (anterior) is to the right and
tissue was present during fetal development. All XX caudal (posterior) is to the left. Tg85 (N⫽24) and XX Tg94 (N⫽26) gonads developed
testicular tissue exclusively (Figure 2). In contrast,
ovar-ian tissue is readily visible in all gonads from E14.5–15.5 expression is first initiated (Koopman et al. 1990; B6 XYPOSfetuses (EicherandWashburn2001).
Hackeret al.1995;Jeskeet al.1995). Tg94 transcripts Semiquantitative RT-PCR analysis revealed that Tg85 were detected in 7-ts-stage XY Tg fetuses whereas endog-and Tg94 were overexpressed relative toSryB6in B6 XY
enousSrytranscripts were not yet detectable (three pairs Tg fetal gonads at the 18-ts stage (E11.5), the timepoint of urogenital ridges, 35 PCR cycles). Tg94 expression when Sry is normally maximally expressed: Tg85 was also was detected in 9-ts XX Tg and 11-ts XY Tg fetuses expressed threefold greater and Tg94 fivefold greater (one pair of urogenital ridges each). Expression of Tg94 than the endogenous SryB6 allele (Figure 5). We
con-was clearly higher than in the endogenous SryB6 allele
clude that overexpression ofSryPOSallows normal testes
in the 11-ts XY Tg sample. Tg85 expression was detected
to develop in E14.5 B6 fetuses. in 9-ts-stage fetuses (two pairs of XX Tg and one pair
Testis cord development is delayed in XX Tg85 and of XY Tg urogenital ridges) and transgenicSry
expres-XX Tg94 fetal gonads: At E14.5–15.5 B6 XYAKR fetal
sion was clearly higher than in the endogenousSryB6in
gonads are normal appearing testes but at develop- the XY Tg sample. We conclude that testis cord develop-mental stages prior to E14.5, testis cord development is ment is delayed in XX Tg85 and XX Tg94 fetal gonads delayed relative to B6 XYB6gonads. We examined testis despite normal transcriptional initiation and
overex-cord differentiation in E13.5 XX Tg85 and XX Tg94 pression of the transgenes.
fetuses to determine if testis development was normal. Because theSryTg constructs might be missing regu-Similar to B6 XYAKR, all XX Tg85 (N⫽10) and XX Tg94 latory elements necessary for the initiation of normal,
(N⫽14) gonads had delayed testis cord development nondelayed testis cord development, we examined testis
(Figure 6). cord development in E13.5 B6 XX Tg fetuses from an
Because delayed testis cord development could be Srytransgenic line, Tg2, carrying an intact 14.6-kbSry129
caused by delayed initiation of transgene expression, (MUS) Tg. As shown in Figure 6, in contrast to the we assayed transgene expression in urogenital ridges delayed testis cord development observed in E13.5 B6 XX Tg85 (DOM), B6 XX Tg94 (DOM), and B6 XYAKR
fetuses, testis cord development in E13.5 B6 XX Tg2 (MUS) and B6 XYAKR Tg2 fetuses was complete (N ⫽
12 gonads). Because the Tg2 and Tg94 constructs tain the same MUS-derived regulatory regions, we con-clude that delayed testis cord development in B6 XX Tg94 fetuses is not caused by the absence of a critical regulatory region(s).
External sexual phenotype of XX Tg mice and trans-gene expression level are sensitive to trans-genetic back-ground:Of the sevenSrytransgenes analyzed, Tg85 and Tg94 were the only ones in which 100% of the B6 XX Tg offspring were completely sex reversed (Table 2).
In contrast, at weaning 56% of B6 XX Tg17 mice, 9% Figure 7.—Expression of Tg83 is increased on a (D2 ⫻ B6)F1 genetic background. AverageSry/Lhx1expressionvs.
of B6 XX Tg28 mice, and 18% of B6 XX Tg121 mice
developmental age in B6 Tg83 and (D2⫻B6)F1Tg83 E11.5
presented as male. No B6 XX Tg83 (N⫽84) or B6 XX
urogenital ridges. Initial (16- to 17-ts) expression was similar in
Tg84 (N ⫽71) mice presented as males. both backgrounds. However, at the 18- to 21-ts developmental Because the XX Tg females are fertile, we intercrossed stage, Tg83 expression was increased on the F
1background.
hemizygotes from the Tg28, Tg83, and Tg84 lines to The data presented represent average expression, and not all XX Tg 83 gonads are destined to develop as testes. Therefore,
determine if these transgenes caused XX SR when
ho-the difference in expression between ho-the B6 and F1genetic
mozygous. Insertion of the transgene created recessive
backgrounds probably is greater than represented. The
num-lethal mutations in the Tg28 and Tg83 lines (as sug- ber of gonads analyzed for each stage is indicated. gested by underrepresentation of transgenic offspring
in the intercross) so that the phenotype of Tg
homozy-gotes could not be examined. From Tg84 intercrosses, data presented in Figure 7 represent average expression, and not all of the XX Tg 83 gonads are destined to two XX Tg SR males were present among the 37 XX Tg
offspring. Because known XX Tg84/⫹mice are not sex develop as testes. Therefore, the difference in expres-sion between the B6 and F1genetic backgrounds
proba-reversed, we conclude that two copies of Tg84 can cause
XX sex reversal. The homozygous phenotypes for Tg17 bly is greater than represented. This idea is supported by the relatively large range of Tg83 expression obtained and Tg121 were not examined because B6 XX Tg
het-erozygotes are sometimes sex reversed. for these gonads (data not shown). These data suggest
that expression of Tg83 is sensitive to genetic back-Because B6-YPOSSR is highly sensitive to genetic
back-ground, we produced F1hybrid Tg mice by mating B6 ground, a finding that correlates with the external
sex-ual phenotype observed in F1XX Tg83 mice.
Tg carriers to D2 and C3H/HeSnJ (C3H) mice and examined the external sexual phenotype of XX Tg mice at weaning (Table 2). In the three transgenic lines tested
DISCUSSION
(bothSryPOSandSry129-POS), the phenotype of XX Tg mice
was modulated by genetic background. For example, at Transfer of certainM. domesticus-derived (DOM) Y chro-mosomes (SryDOMalleles) onto specific inbred strains, such
one extreme, 81% (13/16) of the (C3H or D2⫻B6)F1
XX Tg83 mice presented as males whereas all (N ⫽ as B6, causes abnormal testis determination (Eicheret al. 1982; Eicher and Washburn 1986; Nagamine et 84) B6 XX Tg83 mice presented as females. For Tg84,
different F1hybrid backgrounds gave different results: al.1987;Biddle andNishioka 1988). The degree of
abnormality depends on the particular Y chromosome All 23 (D2⫻B6)F1XX Tg84 mice were female whereas
7 (C3H⫻B6)F1XX Tg84 mice were female and 11 were transferred. The experiments presented were designed
to elucidate the mechanism ofSryDOMmisfunction when
male. Surprisingly, none of the F1XX Tg84 mice were
obvious hermaphrodites. We conclude that the external present on the B6 genetic background.
To determine if DOMSryalleles are expressed at differ-sexual phenotype and, by inference, testis
determina-tion of XX Tg mice is sensitive to genetic background. ent levels or in different temporal patterns from those of MUSSry alleles, we developed two B6 mouse lines Semiquantitative RT-PCR was used to determine if an
increase in transgenic RNA transcript levels correlated that each carry a single DOMSryallele (POS or AKR) and a single MUSSryallele (RIII). Gonads in B6 XYPOS,RIII
with sex reversal in F1XX Tg mice. We analyzed (D2⫻
B6) Tg83 E11.5 gonads because Tg83 seemed to be the and B6 XYAKR,RIIImice are phenotypically normal testes.
This finding confirms and extends results demonstra-most sensitive to genetic background. Tg83 expression
was compared toLhx1 expression in gonads from 16- ting that transgenic overexpression of a MUSSry129allele
rescues SR in B6-YPOS mice (Eicher et al. 1995) and
to 21-ts fetuses. As illustrated in Figure 7, initial
(16-to 17-ts) expression was similar in both backgrounds. delayed testis cord development in B6-YAKR mice (data
presented here). Also, the data indicate that the DOM However, at the 18- to 21-ts developmental stage, Tg83
alleles. It is interesting to note that, to our knowledge, Sry allele is more sensitive to genetic background than theSryRIIIallele. It is likely, therefore, that at least one
allSrytransgenic lines capable of sex reversing XX mice
contain multiple insertions of the Srygene (Koopman tdagene affectsSryexpression and that this interaction is direct. The simplest model is that one or more tda et al. 1991;Eicher et al. 1995; Washburnet al. 2001).
The reasons for this are not clear, but it may be that genes is a transcription factor that controlsSry transcrip-tion by directly interacting with theSrypromoter. How-expression of a transgene is dependent on the
chromo-somal site of insertion and on the presence of critical ever, other models are possible. For example, atdagene could interact with theSrytranscript and affect its stabil-cis-acting regulatory elements. In the case of the B6
XYPOS,RIIIand B6 XYAKR,RIIImice, a single copy of a MUS- ity or localization. Further functional studies are needed
to test these models. derivedSrygene was present and able to correct
testicu-lar abnormalities. We suggest that the reason this single We found that in B6 XYAKR,RIIIfetal gonads the DOM SryAKR and MUS
SryRIII alleles were expressed at equal
copy ofSryfunctioned normally was that it was present
on a segment of the mouse Y chromosome that normally levels. The question of whether theSryRIIIallele initiates
normal testis determination is complicated by the fact contains it.
If the misexpression hypothesis is correct, expression that we analyzed testis development in XXSxr fetuses
where random X inactivation can affect the expression of theSryPOSallele should be more “abnormal” than that
of the SryAKR allele. This was, in fact, the case: On the of theSryRIIIallele. However, 32 of the 40 B6 XXSxrgonads
examined between E13.25 and E14.5 were normal testes B6 background, the DOMSryPOSallele was expressed at
ⵑ59% of the MUSSryRIIIallele whereas the DOMSryAKR without delayed testis cord development. (The
re-maining 8 gonads were ovotestes.) This result suggests allele and the MUSSryRIIIallele were expressed at equal
levels. Moreover, if the misexpression hypothesis is cor- that in the absence of significant inactivation of the XSxr
chromosome, theSryRIIIexpression level is sufficient to
rect, expression ofSryPOSwould be more “normal” on a
hybrid genetic background known to rescue B6-YPOSsex initiate normal testis development on the B6
back-ground. BecauseSryAKRandSryRIIIare expressed at
equiv-reversal. This, too, was the case: The SryPOS allele was
expressed atⵑ74% of the MUSSryRIIIallele on a (D2⫻ alent levels yet B6 XYAKRgonads have delayed testis cord
development, delayed testis cord development cannot B6)F1genetic background. Because relativeSry
expres-sion was measured in genital ridges destined to develop be attributed solely to insufficient SryAKR
expression. Rather, delayed testis cord development probably is as normal gonads and independent of the number of
Sry-expressing cells, we conclude thatSryexpression per caused by reduced translation of the SryAKR transcript,
by reduced stability of the SRYAKR protein isoform, or
cell is reduced. The results, however, do not exclude
the possibility that the number of Sry-expressing cells by reduced ability of the SRYAKRprotein isoform [which
is approximately half the size of the MUS SRY protein also is reduced.
The fact that the relative expression of SryPOS/SryRIII isoform (Cowardet al.1994)] to initiate testis
develop-ment. These results are consistent with the finding that andSryAKR/SryRIIIwas constant between E10.5 and E13.0
suggests that the temporal expression of DOM and MUS SryB6 (MUS) is expressed at lower levels than SryAKR is
yet testis development in B6 XYB6mice is normal (Lee
alleles is similar during this time. Therefore, it is unlikely
that delayedSryexpression is responsible for either SR and Taketo 1994 and results herein). Furthermore, these results suggest that at least one tdagene partici-in B6 XYPOSgonads or delayed testis development in B6
XYAKRgonads. These results are consistent with those of pates in the sex-determination cascade downstream of
or in parallel withSry. LeeandTaketo(1994) andNagamineet al.(1999).
After E13.0, expression ofSryPOSpersisted longer than Overall, theSryexpression analysis indicates that
B6-YPOS SR is caused by insufficient SryPOS expression and
expression ofSryRIII. This result implies thatSryPOS
expres-sion is downregulated more slowly than SryRIII
expres- that delayed testis cord development in B6 XYAKRmice
is caused by reduced efficiency of the SRYAKRisoform.
sion. However, we cannot exclude the possibility that theSryPOS transcript is more stable than the
SryRIIItran- If this model is correct, then overexpression of
SryPOSin
B6 mice would rescue SR but might not rescue delayed script. We suggest that if persistent expression is due to
inefficient downregulation ofSryPOSexpression, then the testis cord development. Two different transgenic
con-structs were used to test this hypothesis: anSryPOS
geno-same regulatory elements that prevent efficient
upregu-lation ofSryPOSexpression may be identical to those that mic DNA clone and a chimeric construct in which
ex-pression of the SryPOS ORF was controlled by SryMUS
prevent efficient downregulation.
The relative expression results were confirmed by regulatory regions (Sry129-POS). Two B6 transgenic lines,
one from each type of construct, were established in measuring expression of the individual Sry alleles
against expression of a control gene (Lhx1). These data which all XX transgenic progeny developed testes. How-ever, testis cord development was delayed in both lines indicated that expression of both the DOM and MUS
Sry alleles was increased on the hybrid genetic back- despite overexpression of SryPOS and normal
transcrip-tional initiation from the transgenes. These results sug-ground, but the expression of the DOM allele was
relatively high levels; however, overexpression is not suf- genetic background. We do not know if the difference between the D2 and C3H inbred strains is due to differ-ficient to correct delayed testis cord development. The
transgenic results support a model where delayed testis ent alleles of the tda genes previously mapped or to differences in noveltdagenes. Molecular identification cord development is caused by the presence of
particu-lar DOM SRY protein isoforms that cause SR when un- of the tdagenes will clarify this.
As noted in the Introduction, several intriguing but derexpressed. The fact that (D2⫻ B6)F1XYPOSfetuses
develop normal testes without evidence of delay unexplained SR conditions are found in humans, in-cluding XY females and XY hermaphrodites who carry (Eicher et al. 1996) indicates that delayed testis cord
development requires that at least onetdagene be ho- an apparently normal SRY gene and XY females who carry a mutatedSRY gene inherited from their carrier mozygous for the B6 allele.
To our knowledge, all SryDOM ORFs analyzed have a father. We hypothesize that these human SR conditions
are like B6-YPOSSR and are caused by conditionally
insuf-stop codon in the glutamine repeat region downstream
of the HMG box, which means that SRYDOMproteins are ficientSRYexpression. Therefore, it is possible that the
human homologs oftdagenes implicated in B6-YPOSSR
about half the size of SRYMUS proteins (Cowardet al.
1994;Carlisleet al.1996;AlbrechtandEicher1997). play a role in these and other human SR conditions. However, the predicted “half-size” SRYDOMprotein
iso-We thank members of The Jackson Laboratory Microinjection and
form alone is not sufficient to account for either sex Microchemistry services for their technical assistance. Appreciation is expressed to Jason Stockwell of The Jackson Laboratory
Computa-reversal or delayed testis development when present on
tional Biology Resource for statistical analysis of the data and to Robin
the B6 genetic background because someSryDOMalleles,
Lovell-Badge (MRC, NIMR, London) for providing the L961SryPOS
such asSryFVBandSryBUB, initiate normal testis
develop-clone. We are grateful to Luanne L. Peters and Timothy P. O’Brien for
ment when on the B6 background (BiddleandNishi- critical reviews of the manuscript. This work was funded by National oka1988;Nagamineet al.1999;Washburnet al.2001). Institutes of Health research grant GM-20919 (to E.M.E.), fellowships GM-16726 (to K.H.A) and HD-08492 (to M.Y.), and by a National
DifferentSryDOMprotein isoforms that differ in the
num-Cancer Institute CORE grant CA34196 (to The Jackson Laboratory).
ber of glutamines encoded by the third glutamine re-peat cluster (GRC-3) have been identified. No correla-tion is found between the number of glutamines in
GRC-3 and sex reversal (Carlisleet al.1996;Albrecht LITERATURE CITED
andEicher1997). However, it is possible that the num- Albrecht, K. H., andE. M. Eicher, 1997 DNA Sequence analysis ber of glutamines in GRC-3 plays a role in whether a of Sryalleles (subgenus Mus) implicates misregulation as the cause of C57BL/6J-YPOSsex reversal and defines the SRY
func-given SryDOM allele causes delayed testis development
tional unit. Genetics147:1267–1277.
when on the B6 background.SryAKR, which causes
de-Barnes, J. D., J. L. Crosby, C. M. Jones, C. V. WrightandB. L.
layed testis development, has 13 glutamines in GRC-3 Hogan, 1994 Embryonic expression ofLim-1, the mouse homo-log ofXenopus Xlim-1, suggests a role in lateral mesoderm
differen-whileSryFVBandSryBUB, which initiate normal testis
devel-tiation and neurogenesis. Dev. Biol.161:168–178.
opment, have 12. The situation is complicated by the Bergstrom, D. E., M. Young, K. H. AlbrechtandE. M. Eicher, fact that differentSryalleles are expressed at different 2000 Related function of mouse SOX3, SOX9, and SRY HMG domains assayed by male sex determination. Genesis28:111–124.
levels. For example, the SryFVB allele is expressed at
Biddle, F. G., andY. Nishioka, 1988 Assays of testis development
higher levels than theSryAKRallele is (Nagamineet al.
of the mouse distinguish three classes of domesticus-type Y
chro-1999). mosome. Genome30:870–878.
Bishop, C. E., P. Boursot, B. Baron, F. BonhommeandD. Hatat,
Not all of the transgenes produced exclusively male
1985 Most classical Mus musculus domesticus laboratory mouse
XX Tg progeny, and for several the percentage of male strains carry a Mus musculus musculus Y chromosome. Nature XX Tg progeny was increased on a hybrid genetic back- 315:70–72.
Bishop, C. E., A. Weith, M. G. MatteiandC. Roberts, 1988
Molec-ground. For the one transgene examined, the increase
ular aspects of sex determination in mice: an alternative model
in male XX Tg progeny was correlated with increased for the origin of the Sxr region. Philos. Trans. R. Soc. Lond. B expression of the Sry transgene. The results indicate Biol. Sci.322:119–124.
Bonhomme, F., 1986 Evolutionary relationships in the genusMus. that the transgenes contain a DNA element that controls
Curr. Top. Microbiol. Immunol.127:19–34.
Sry expression level and is sensitive to genetic back- Capel, B., K. H. Albrecht, L. L. WashburnandE. M. Eicher, ground. We suggest that this element is likely to directly 1999 Migration of mesonephric cells into the mammalian
go-nad depends onSry.Mech. Dev.84:127–131.
interact with atdagene. Furthermore, this control
ele-Carlisle, C., H. Winking, D. WeichenhanandC. M. Nagamine,
ment is present in the region of minimal overlap
be-1996 Absence of correlation betweenSrypolymorphisms and
tween the two types of transgenes (i.e., betweenⵑ2355 XY sex reversal caused by theM. m. domesticus Ychromosome. Genomics33:32–45.
bp andⵑ14,625 bp). Future experiments are focused
Cattanach, B. M., C. E. PollardandS. G. Hawkes, 1971
Sex-on identifying theSryexpression control element.
reversed mice: XX and XO males. Cytogenetics10:318–337.
We are intrigued by the finding that all (D2⫻B6)F1 Cattanach, B. M., E. P. Evans, M. D. BurtenshawandJ. Barlow, 1982 Male, female and intersex development in mice of
identi-XX Tg84 mice are female whereas approximately half
cal chromosome constitution. Nature300:445–446.
of the (C3H⫻B6)F1XX Tg84 mice are female and the
Coward, P., K. Nagai, D. Chen, H. D. Thomas, C. M. Nagamineet remainder are male. This result nicely illustrates the fact al., 1994 Polymorphism of a CAG trinucleotide repeat within Srycorrelates with B6.YDOMsex reversal. Nat. Genet.6:245–250.