Genotyping
Yersinia enterocolitica
Strains from Human and Porcine
Origins
S. Virtanen,aR. Laukkanen-Ninios,aP. Ortiz Martínez,bA. Siitonen,cM. Fredriksson-Ahomaa,aH. Korkealaa
Department of Food Hygiene and Environmental Health, Faculty of Veterinary Medicine, University of Helsinki, Helsinki, Finlanda
; Mikkelin Seudun Ympäristöpalvelut, Mikkeli, Finlandb
; Bacteriology Unit, National Institute for Health and Welfare, Helsinki, Finlandc
Sporadic and epidemiologically linkedYersinia enterocoliticastrains (nⴝ379) isolated from fecal samples from human pa-tients, tonsil or fecal samples from pigs collected at slaughterhouses, and pork samples collected at meat stores were genotyped using multiple-locus variable-number tandem-repeat analysis (MLVA) with six loci, i.e., V2A, V4, V5, V6, V7, and V9. In total, 312 different MLVA types were found. Similar types were detected (i) in fecal samples collected from human patients over 2 to 3 consecutive years, (ii) in samples from humans and pigs, and (iii) in samples from pigs that originated from the same farms. Among porcine strains, we found farm-specific MLVA profiles. Variations in the numbers of tandem repeats from one to four for variable-number tandem-repeat (VNTR) loci V2A, V5, V6, and V7 were observed within a farm. MLVA was applicable for serotypes O:3, O:5,27, and O:9 and appeared to be a highly discriminating tool for distinguishing sporadic and outbreak-related strains. With long-term use, interpretation of the results became more challenging due to variations in more-discriminating loci, as was observed for strains originating from pig farms. Additionally, we encountered unexpectedly short V2A VNTR fragments and sequenced them. According to the sequencing results, updated guidelines for interpreting V2A VNTR results were prepared.
Y
ersinia enterocoliticais a food-borne pathogen that causes hu-man yersiniosis. Yersiniosis is the third most commonly re-ported food-borne zoonosis (1) and often occurs in young chil-dren (2). The most frequent symptom is gastroenteritis with diarrhea, but additional sequelae such as reactive arthritis and erythema nodosum may occur, especially in adults.Y. enteroco-liticais a heterogeneous species that is divided into several bio-types and serobio-types (2). Bioserotypes 1B/O:8, 2/O:5,27, 2/O:9, 3/O:3, and 4/O:3 are associated commonly with human disease. Of these, bioserotype 4/O:3 strains have been responsible for yersiniosis cases in Europe, the United States, Canada, and Japan (3–6). Recently, bioserotype 2/O:9 infections have been on the rise (7–9). Besides humans, bioserotype 4/O:3 frequently is isolated from samples of pig origin (2,10–13).Pulsed-field gel electrophoresis (PFGE) has been one of the most commonly used genotyping methods for epidemiological studies ofY. enterocolitica (14–17), but more-sophisticated and automated methods are needed and recently have been applied. Multiple-locus variable-number tandem-repeat analysis (MLVA) has been developed for several pathogenic bacterial species (18,
19). The method is based on finding variable-number tandem repeats (VNTRs) in the genome and multiplying these fragments in a PCR. Different genotypes are distinguished by the number of repeats in each locus (18). The application developed forY. en-terocolitica4/O:3 uses six VNTR loci, i.e., V2A, V4, V5, V6, V7, and V9 (20). MLVA also has proved successful withY. enterocolitica serotype O:9 (7,8,21,22). A separate MLVA application has been developed forY. enterocoliticabiotype 1A (23).
MLVA has proved to be a promising tool forY. enterocolitica outbreak investigations, due to its high discriminatory power (7). However, the number of strains studied has been limited. There-fore, MLVA should undergo further testing with several sporadic and epidemiologically linked strains, to define its suitability for practical use. In this work, we used a previously developed MLVA
typing method to investigate a collection ofY. enterocoliticastrains from both sporadic and epidemiologically linked origins that had been acquired over 12 years, between 1995 and 2007. The strains originated from Finland, Germany, England, and Russia and were isolated from human patients as well as from pig farms, meat stores, and slaughterhouses. The discriminatory power, advan-tages, limitations, interpretation, and use of MLVA in epidemio-logical studies ofY. enterocoliticawere evaluated. Among strains from the same epidemiological origin, we commonly observed instability in loci V2A, V5, V6, and V7. Lastly, our observation of some unexpectedly short V2A PCR products was further studied.
MATERIALS AND METHODS
Y. enterocoliticastrains and isolation of DNA.For this study, we chose a total of 379 strains ofY. enterocoliticafrom Finland (n⫽288), Germany (n⫽46), England (n⫽34), and Russia (n⫽11). The strains were isolated between 1995 and 2007 and originated from clinical samples from human patients (n⫽150), tonsil and fecal samples from pigs collected at slaugh-terhouses (n⫽183), and pork samples collected at meat stores (n⫽46). Of these, 128 samples collected at slaughterhouses were traced to have originated from 35 pig farms. We included both sporadic (n⫽278) and epidemiologically linked (n⫽101) strains. The latter strains were consid-ered epidemiologically linked when they originated from pigs from the same farm. The strains belonged to serotypes O:3 (n⫽363), O:9 (n⫽9),
Received15 March 2013Returned for modification12 April 2013
Accepted22 April 2013
Published ahead of print1 May 2013
Address correspondence to S. Virtanen, [email protected].
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JCM.00710-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.00710-13
on May 16, 2020 by guest
http://jcm.asm.org/
and O:5,27 (n⫽7). We previously divided all serotype O:3 strains into 81 genotypes using PFGE with the enzyme NotI, and we divided 206 of those strains into 105 genotypes using the additional enzymes ApaI and XhoI (16,17,24–26). The strains were grown on tryptic soy agar (Difco, Law-rence, KS) at 28°C for 18 to 24 h, and the DNA was extracted using guanidium thiocyanate (27).
Multiple-locus variable-number tandem-repeat analysis.The MLVA was based on the method described by Gierczynski et al. (20), with mod-ification of the PCR run conditions (7). The forward primers for VNTR loci V2A, V4, V5, V6, V7, and V9 were fluorescently labeled with 6FAM, VIC, NED, and PET dyes (Applied Biosystems, Foster City, CA) and di-vided into two PCRs. One microliter of the template was added to the PCR mixture containing a 0.2 mM deoxynucleoside triphosphate mixture (Thermo Fisher Scientific, Vantaa, Finland) and 0.5 U of DyNAzyme II polymerase (Thermo Fisher Scientific), in a total reaction volume of 25l. A 1.5-l volume of 10-fold diluted PCR products was denatured with 10 l of HiDi formamide (Applied Biosystems) mixed with 0.3l of the GeneScan 500 LIZ internal standard (Applied Biosystems), boiled for 3 min, and placed on ice. The sizes of the PCR products were determined by capillary electrophoresis with an ABI Prism 310 genetic analyzer (Applied Biosystems). An injection time of 5 s and an injection voltage of 15 kV were used for electrophoresis. The run time was 28 min, the voltage was 15 kV, and the run temperature was set at 60°C. Certain V2A VNTR frag-ments of unexpected size (less than 263 bp) were sequenced using a Big-Dye Terminator v3.1 cycle sequencing kit (Applied Biosystems) with an ABI 3730xl DNA analyzer (Applied Biosystems).
Data analysis.The MLVA data were collected with GeneScan software (Applied Biosystems) and analyzed with BioNumerics v. 5.1 (Applied Maths NV, Sint-Martens-Latem, Belgium). The tandem-repeat number for each locus was saved as character data type in the BioNumerics soft-ware. The clustering of the MLVA results was calculated using Euclidian distances. To compare the discriminatory abilities of the VNTR loci, Simpson’s discriminatory indexes were calculated according to the method described by Hunter and Gaston (28). The chi-square test was used to compare MLVA and PFGE results.
RESULTS
Discrimination of strains.Among the 379 strains examined, we detected 312 different MLVA types. A dendrogram of the results appears in Appendix S1 in the supplemental material. After ex-cluding strains with the same epidemiological source, we detected 262 different MLVA types among 278 strains. The Simpson dis-criminatory index for the sporadic strains was 0.999. All of the MLVA types in the present study were discovered to be country specific, as none of the types was detected in several countries.
Groups of similar MLVA types were detected among strains originating from the same source, i.e., humans or pigs (see
Appen-dix S1 in the supplemental material). Within several MLVA pro-files, the PFGE results obtained by using three enzymes aggregated accordingly. However, a few strains with similar MLVA genotypes revealed diverse PFGE results. Interestingly, we observed in the MLVA results a significant (P⬍0.05) association between PFGE type NA/AA/HA and repeat numbers of two in locus V4 and three in locus V9 (see Appendix S1 in the supplemental material). Fi-nally, MLVA showed higher discriminatory power than the stan-dard method PFGE, even with three enzymes. Among the 206 serotype O:3 strains that were divided into 105 genotypes using PFGE with the enzymes NotI, ApaI, and XhoI, we detected a total of 193 different MLVA types.
Typing of human and porcine strains.We observed similar MLVA types between strains isolated from (i) human fecal sam-ples, (ii) human patients and pigs, and (iii) pigs originating from the same farm but also from two farms. The same MLVA types were found in human clinical isolates from Finland. Evidence of epidemics limited to certain provinces was observed between 1996 and 1999 (Table 1). Six groups of closely related MLVA types that included 19 humanY. enterocolitica cases were limited to one province. Additionally, four groups of similar MLVA profiles that were detected over a period of 3 years were found in two or three provinces within the country. Among strains from humans and pigs, similar MLVA results were found for human fecal samples and samples originating from pigs (Table 2).
[image:2.585.44.544.87.216.2]A farm-specific MLVA type was found for 17 farms (49% of all farms studied). Two separate farm-specific MLVA profiles were discovered for 13 (37%) farms, whereas three, four, and five dif-ferent MLVA profiles were found for two farms, three farms, and one farm, respectively. Interestingly, all English pig farms that
TABLE 1Similar MLVA results forYersinia enterocoliticastrains isolated from clinical fecal samples from human patients in certain provinces of Finland
No. of VNTRs in loci V2A, V4, V5,
V6, V7, and V9 No. of strains Province Yr(s) isolated
8, 2, 11, 16, 9, 3 3 Satakunta 1996
9, 7, 7, 10, 9, 4 4 Southwest Finland 1996–1997
12, 2, 10, 16 or 17, 5, 3 4 Southwest Finland 1996–1998
11 or 12, 2, 9, 8, 6, 3 3 Ostrobothnia 1997–1998
11, 2, 10, 8, 8, 3 3 Ostrobothnia 1998
7, 2, 15, 4, 6, 2 2 Åland 1999
15, 2, 9, 13, 9, 3 2 Tampere region and southwest Finland 1996–1998
6, 2, 8, 7, 7 or 8, 3 2 Uusimaa and Åland 1996–1998
11 or 12, 2, 13 or 14, 13, 6–8, 3 5 Tampere region and Ostrobothnia 1996–1998
8 or 9, 7, 8, 9, 8–10, 4 3 Several 1997–1999
TABLE 2Similar MLVA results forYersinia enterocoliticastrains isolated from pig and human sources
No. of VNTRs in loci V2A, V4,
V5, V6, V7, and V9 Sample material Yr isolated
3, 6, 10, 8, 9, 5 Pig heart 1998
3, 6, 10, 8, 9, 5 Human feces 1997
12, 4, 12, 10, 11, 3 Pig tongue 1996
12, 4, 12, 10, 11, 3 Human feces 1997
8, 2, 11, 9, 6, 3 Pig liver 1997
8, 2, 11, 9, 6, 3 Human feces 1997
13, 10, 9, 12, 7, 5 Pig tongue 1998
13, 10, 9, 13, 7, 5 Human feces 1996
on May 16, 2020 by guest
http://jcm.asm.org/
[image:2.585.299.544.616.723.2]were included in this study had more than one MLVA type. Sim-ilar MLVA types were found for the pig farms GB2 and GB4, GB4 and GB5, FI5 and FI6, FI12 and FI13, FI17 and FI18, and RU1 and RU2, which were located in England, Finland, and Russia (Table 3). Vast variations in distances were observed for the geographical positions of the farms with similar MLVA types, ranging from 10 km between farms FI5 and FI6 to 300 km between farms RU1 and RU2. Samples from pigs that originated from farms FI5 and FI6 and also from farms FI12 and FI13 were collected at the same time and at the same slaughterhouse, whereas samples originating from the other farms were collected separately.
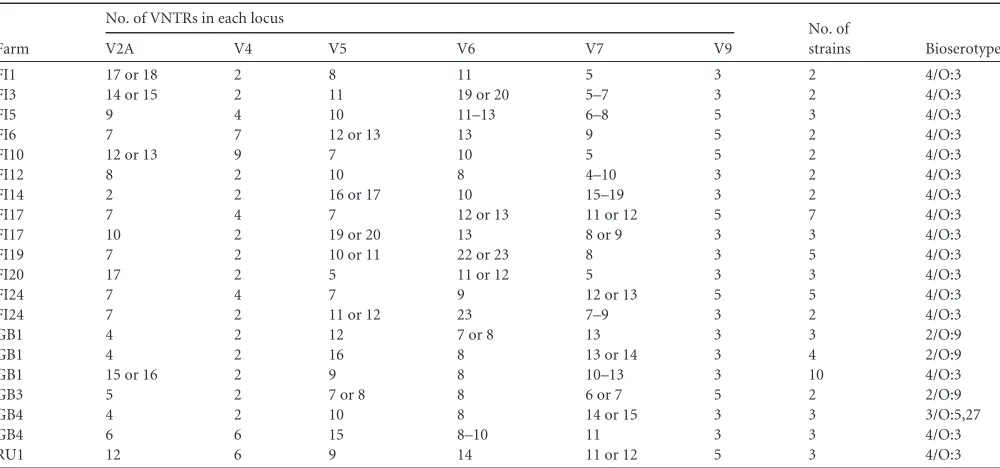
Variations in V2A, V5, V6, and V7 VNTR loci.Within the
MLVA results for strains that originated from the same pig farms, variations in tandem-repeat numbers were noted commonly for the loci V2A, V5, V6, and V7. Among the strains from 15 farms, we observed variations in one to three of these loci (Table 4). How-ever, such variations were not observed for loci V4 and V9. The discriminatory index values for tandem-repeat loci V2A, V4, V5, V6, V7, and V9 were 91.1%, 53.7%, 83.3%, 84.9%, 82.1%, and 46.0%, respectively. For one strain, the PCR product for VNTR locus V7 primers remained absent, despite several repeated reac-tions.
[image:3.585.38.550.77.300.2]Unexpectedly short V2A VNTR fragments.For 100 strains, the V2A VNTR fragments obtained were shorter than expected,
TABLE 3Similar MLVA results detected on two farms
Country and farm pair
No. of VNTRs in loci V2A, V4, V5, V6, V7, and V9
Distance between
farms (km) Sampling date
England
GB2 4, 2, 10, 8, 15, 3 145 3 June 2003
GB4 4, 2, 10, 8, 14 or 15, 3 30 December 2003
GB4 6, 6, 15, 8–10, 11, 3 233 30 December 2003
GB5 6, 6, 15, 10, 11, 3 9 June 2003
Finland
FI5 7, 7, 12, 13, 9, 5 10 18 September 2007
FI6 7, 7, 13, 13, 9, 5 18 September 2007
FI12 8, 2, 10, 8, 4, 3 90 16 October 2007
FI13 8, 2, 10, 8, 4, 3 16 October 2007
FI17 7, 4, 7, 12 or 13, 11 or 12, 5 55 28 January 2004
FI18 7, 4, 7, 12, 12, 5 23 April 2004
Russia
RU1 12, 6, 9, 14, 11 or 12, 5 300 23 June 2006
RU2 12, 6, 9, 14, 11, 5 28 June 2006
TABLE 4Variations detected for different VNTR loci ofYersinia enterocoliticastrains originating from the same pig farms
Farm
No. of VNTRs in each locus
No. of
strains Bioserotype
V2A V4 V5 V6 V7 V9
FI1 17 or 18 2 8 11 5 3 2 4/O:3
FI3 14 or 15 2 11 19 or 20 5–7 3 2 4/O:3
FI5 9 4 10 11–13 6–8 5 3 4/O:3
FI6 7 7 12 or 13 13 9 5 2 4/O:3
FI10 12 or 13 9 7 10 5 5 2 4/O:3
FI12 8 2 10 8 4–10 3 2 4/O:3
FI14 2 2 16 or 17 10 15–19 3 2 4/O:3
FI17 7 4 7 12 or 13 11 or 12 5 7 4/O:3
FI17 10 2 19 or 20 13 8 or 9 3 3 4/O:3
FI19 7 2 10 or 11 22 or 23 8 3 5 4/O:3
FI20 17 2 5 11 or 12 5 3 3 4/O:3
FI24 7 4 7 9 12 or 13 5 5 4/O:3
FI24 7 2 11 or 12 23 7–9 3 2 4/O:3
GB1 4 2 12 7 or 8 13 3 3 2/O:9
GB1 4 2 16 8 13 or 14 3 4 2/O:9
GB1 15 or 16 2 9 8 10–13 3 10 4/O:3
GB3 5 2 7 or 8 8 6 or 7 5 2 2/O:9
GB4 4 2 10 8 14 or 15 3 3 3/O:5,27
GB4 6 6 15 8–10 11 3 3 4/O:3
RU1 12 6 9 14 11 or 12 5 3 4/O:3
on May 16, 2020 by guest
http://jcm.asm.org/
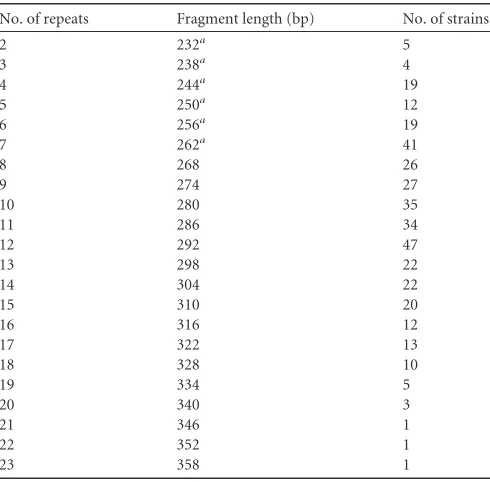
[image:3.585.43.543.488.722.2]being less than 263 bp. However, the lengths of these exceptionally short fragments varied in 6-bp intervals, with results of 232, 238, 244, 250, 256, and 262 bp. The short V2A fragments of all lengths were sequenced in order to clarify our results. The real repeat numbers of these fragments, as confirmed by sequencing, were 2, 3, 4, 5, 6, and 7 when the VNTR fragment lengths were 232, 238, 244, 250, 256, and 262 bp, respectively (Table 5). These results indicate an updated interpretation of the results for the V2A VNTR locus.
DISCUSSION
We typed a large number ofY. enterocoliticastrains from human and porcine sources and from four different countries by using MLVA with six VNTR loci. The method showed a high discrimi-natory capacity, since 312 different MLVA types were found among 379Y. enterocoliticastrains. MLVA found 45 genotypes among 62Y. enterocolitica4/O:3 strains in a previous study (20) and 77 types among 88Y. enterocoliticastrains in another study (7). When strains with the same epidemiological sources were excluded, we detected 262 MLVA types among 278 strains, whereas 21Y. enterocoliticastrains with no epidemiological link were previously discriminated into 20 genotypes using MLVA (20).
Interestingly, with randomly chosen clinical strains isolated from human fecal samples in Finland, we observed similar MLVA types over a period of several years. Some of these strains were isolated from the same provinces, indicating unidentified geo-graphically limited epidemics, while others were distributed in different provinces within the country but still seemed to be cases with apparently common sources. MostY. enterocolitica infec-tions are sporadic, and outbreaks are considered uncommon (2). Epidemics caused byY. enterocoliticaseem to exist, however, al-though many of them have remained unidentified. The
occur-rence of the same MLVA types in clinical samples collected over several years indicates long-term persistence and the availability of the same infectious sources.
Similar genotypes of strains that were isolated from humans and pigs in corresponding time periods were discovered with MLVA, suggesting a link between these two sources. Strains iso-lated from humans and porcine sources showed similar PFGE types previously (16,17). As MLVA has markedly greater discrim-inatory capacity than PFGE, our results confirm that edible pig offal is an important source in the transmission ofY. enterocolitica between pigs and humans. However, some strains that originated from human clinical samples were isolated before the correspond-ing MLVA types were obtained from porcine samples. As certain Y. enterocoliticastrains seem to persist on pig farms (29), the com-mon MLVA types found acom-mong pigs and humans likely existed on the farms before the isolation from human clinical samples in-cluded in this study.
Among strains from pig origins, similar MLVA types were de-tected mainly from pigs originating from the same farms. How-ever, some MLVA types were found on several farms.Y. enteroco-liticastrains can be transmitted from one farm to another with infected pigs (29). After pigs from different origins are mixed at the farms, different MLVA types are incorporated and spread rap-idly in the pig population within each unit. Previous contacts and animal sales between the farms might have enabled the spread of similar MLVA types at these farms. Interestingly, similar MLVA types found at several farms were isolated only within one coun-try. Of these, two MLVA profiles were obtained from samples collected on the same day from the same slaughterhouse, and the sampled pigs might have been exposed to contamination at the slaughterhouse instead of being infected at the farms.
Among the six VNTR loci used in this study, the discrimina-tory power of VNTR locus V2A was the highest, whereas VNTR loci V4 and V9 showed the lowest discrimination. This finding is in agreement with those of a previous study (7). Regional differ-ences in the discriminatory abilities of different loci seem to exist, as among Chinese strains; locus V5 exhibited the highest discrim-inatory power, and the least variation occurred in loci V4 and V2A (22). Besides serotypes O:3 and O:9, MLVA proved successful for serotype O:5,27 strains in this study. The 16 strains of serotypes O:5,27 and O:9 were divided into 10 MLVA types. MLVA grouped these strains according to their serotypes and farms of origin. All serotype O:5,27 and O:9 strains in this study originated from sam-ples from pigs from England. These serotypes are particularly common among English pigs, whereas serotype O:3 predominates among pigs from other European countries (13).
Among MLVA results obtained from the same pig farms, vari-ations were detected commonly for four (V2A, V5, V6, and V7) of the six VNTR loci. Allelic variation of a single VNTR locus was reported recently forMycoplasma pneumoniae(30). We detected such variations in several VNTR loci inY. enterocolitica. Both in-sertions and deletions of tandem repeats were observed. Varia-tions were evident in strains isolated from pigs originating from the same farms. Since the same strains are assumed to persist on farms (29), it is unsurprising that mutations occur in these strains over time and lead to variations in VNTRs. The generated varia-tions in one or more VNTRs might cause the MLVA results for identical clones to appear different (19). In addition, environmen-tal stress can affect the stability of tandem repeats (31). Most of the mutations in VNTR units were shown to be single-repeat
differ-TABLE 5Distribution of numbers of tandem repeats in the V2A VNTR locus
No. of repeats Fragment length (bp) No. of strains
2 232a 5
3 238a 4
4 244a 19
5 250a 12
6 256a 19
7 262a 41
8 268 26
9 274 27
10 280 35
11 286 34
12 292 47
13 298 22
14 304 22
15 310 20
16 316 12
17 322 13
18 328 10
19 334 5
20 340 3
21 346 1
22 352 1
23 358 1
a
Confirmed by sequencing.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.41.286.85.326.2]ences (32), but multiple-repeat mutations also occur. Of these, two-repeat mutations are more common than three-repeat muta-tions, which are more common than four-repeat mutations. However, we found no variations in the farm-specific results for VNTR loci V4 and V9, which have VNTR unit lengths of 7 and 12 bp, respectively. These less-discriminating loci thus are longer than the other loci, all of which have a VNTR unit length of six base pairs. It seems that the more-discriminating loci occasionally might be even hypervariable and might be too discriminating for long-term use. This should be taken into consideration when in-terpreting results, especially those obtained from pig farms. Dif-ferences in the VNTR numbers of one or several loci can, in the long run, occur within strains from a single epidemiological origin such as a pig farm.
Interestingly, PFGE type NA/AA/HA (26) was significantly as-sociated with MLVA results showing repeat numbers of two and three for VNTR loci V4 and V9, respectively (see Appendix S1 in the supplemental material). This was the most clearly observed congruity between the results of these two methods in the present study. The strains with this genotype originated from Finland, England, and Russia and were of both human and pig origins. All of these strains belonged to bioserotype 4/O:3.
Compared with the amplicon lengths and corresponding tan-dem-repeat numbers reported in previous studies (7,20), V2A primers produced PCR products that were unexpectedly short, as their lengths were only 232 to 262 bp. The minimal length of a V2A fragment has been reported to be 263 bp (20). However, the sequencing results of our study showed that these 232- to 262-bp fragments contained two to seven VNTR units. According to the previously publishedY. enterocoliticasubsp.palearctica4/O:3 Y11 genome (33), we also noticed that the actual number of repeats was nine when the size of the V2A amplicon was 274 bp. In pre-vious studies, the repeat number of nine equaled lengths of 282 and 310 bp for V2A fragments (7,20). Based on this finding, we prepared an updated table to interpret the results for the V2A VNTR locus (Table 5).
Performing MLVA was fast. With the protocol used, the MLVA results were ready within 24 h. MLVA results can be ana-lyzed with automated software, which reduces the possibilities for human error. MLVA results can be compared between laborato-ries more easily than obsolescent gel images, but this requires in-terlaboratory consistency in MLVA typing and universal guide-lines for interpretation of the results. However, precise guideguide-lines are difficult to develop due to the varying durations of outbreaks and the different environments surrounding the pathogens (34). The MLVA method has high discriminatory power, evidently higher than that of PFGE, as several previous studies have noted (7,22,35). MLVA appears to be a practical tool for genotypingY. enterocolitica, especially in short-term epidemiological studies. Among strains that originate from pigs, MLVA distinguishes the strains according to their farms of origin, but variations in loci V2A, V5, V6, and V7 are common. Reliable comparisons of results between laboratories require uniform interpretation of the results.
ACKNOWLEDGMENTS
This study was performed at the Centre of Excellence in Microbial Food Safety Research, Academy of Finland (grants 118602 and 141140), and was partly funded by the Ministry of Agriculture and Forestry of Finland (grant 2849/502/2008).
Maarit Penttilä, Erika Pitkänen, and Anu Seppänen are acknowledged for their technical assistance.
We declare that we have no competing interests.
REFERENCES
1.European Food Safety Authority.2011. The European Union summary report on trends and sources of zoonoses and zoonotic agents and food-borne outbreaks in 2009. EFSA J.9:2090. doi:10.2903/j.efsa.2011.2090. 2.Fredriksson-Ahomaa M, Lindström M, Korkeala H.2010.Yersinia
en-terocolitica andYersinia pseudotuberculosis, p 164 –180.InJuneja VK, Sofos JN (ed), Pathogens and toxins in foods. ASM Press, Washington, DC.
3.Van Ossel C, Wauters G. 1990. AsymptomaticYersinia enterocolitica infections during an outbreak in a day-nursery. Eur. J. Clin. Microbiol. Infect. Dis.9:148.
4.Thompson JS, Gravel MJ.1986. Family outbreak of gastroenteritis due to Yersinia enterocoliticaserotype 0:3 from well water. Can. J. Microbiol.
32:700 –701.
5.Zenyoji H, Maruyama T, Sakai S, Kimura S, Mizuno T. 1973. An outbreak of enteritis due toYersinia enterocoliticaoccurring at a junior high school. Jpn. J. Microbiol.17:220 –222.
6.Lee LA, Gerber AR, Lonsway DR, Smith JD, Carter GP, Puhr ND, Parrish CM, Sikes RK, Finton RJ, Tauxe RV.1990.Yersinia enterocolitica O:3 infections in infants and children, associated with the household preparation of chitterlings. N. Engl. J. Med.322:984 –987.
7.Sihvonen LM, Toivonen S, Haukka K, Kuusi M, Skurnik M, Siitonen A.
2011. Multilocus variable-number tandem-repeat analysis, pulsed-field gel electrophoresis, and antimicrobial susceptibility patterns in discrimi-nation of sporadic and outbreak-related strains ofYersinia enterocolitica. BMC Microbiol.11:42. doi:10.1186/1471-2180-11-42.
8.MacDonald E, Heier B, Stalheim T, Cudjoe K, Skjerdal T, Wester A, Lindstedt B, Vold L.2011.Yersinia enterocoliticaO:9 infections asso-ciated with bagged salad mix in Norway, February to April 2011. Euro Surveill. 16(19):pii⫽19866. http://www.eurosurveillance.org/View Article.aspx?ArticleId⫽19866.
9.Moriki S, Nobata A, Shibata H, Nagai A, Minami N, Taketani T, Fukushima H.2010. Familial outbreak ofYersinia enterocoliticaserotype O9 biotype 2. J. Infect. Chemother.16:56 –58.
10. Fredriksson-Ahomaa M, Hielm S, Korkeala H.1999. High prevalence of yadA-positiveYersinia enterocoliticain pig tongues and minced meat at the retail level in Finland. J. Food Prot.62:123–127.
11. Fredriksson-Ahomaa M, Björkroth J, Hielm S, Korkeala H.2000. Prev-alence and characterization of pathogenicYersinia enterocoliticain pig tonsils from different slaughterhouses. Food Microbiol.17:93–101. 12. Ortiz Martínez P, Fredriksson-Ahomaa M, Sokolova Y, Roasto M,
Berzins A, Korkeala H.2009. Prevalence of enteropathogenicYersiniain Estonian, Latvian, and Russian (Leningrad region) pigs. Foodborne Pathog. Dis.6:719 –724.
13. Ortiz Martinez P, Mylona S, Drake I, Fredriksson-Ahomaa M, Korkeala H, Corry JE.2010. Wide variety of bioserotypes of enteropathogenic Yersiniain tonsils of English pigs at slaughter. Int. J. Food Microbiol.
139:64 – 69.
14. Najdenski H, Iteman I, Carniel E.1994. Efficient subtyping of patho-genicYersinia enterocoliticastrains by pulsed-field gel electrophoresis. J. Clin. Microbiol.32:2913–2920.
15. Iteman I, Guiyoule A, Carniel E.1996. Comparison of three molecular methods for typing and subtyping pathogenicYersinia enterocolitica strains. J. Med. Microbiol.45:48 –56.
16. Fredriksson-Ahomaa M, Stolle A, Siitonen A, Korkeala H.2006. Spo-radic humanYersinia enterocoliticainfections caused by bioserotype 4/O:3 originate mainly from pigs. J. Med. Microbiol.55:747–749.
17. Fredriksson-Ahomaa M, Hallanvuo S, Korte T, Siitonen A, Korkeala H.
2001. Correspondence of genotypes of sporadicYersinia enterocolitica bio-serotype 4/O:3 strains from human and porcine sources. Epidemiol. In-fect.127:37– 47.
18. van Belkum A.2007. Tracing isolates of bacterial species by multilocus variable number of tandem repeat analysis (MLVA). FEMS Immunol. Med. Microbiol.49:22–27.
19. Lindstedt BA. 2005. Multiple-locus variable number tandem repeats analysis for genetic fingerprinting of pathogenic bacteria. Electrophoresis
26:2567–2582.
20. Gierczynski R, Golubov A, Neubauer H, Pham JN, Rakin A. 2007.
on May 16, 2020 by guest
http://jcm.asm.org/
Development of multiple-locus variable-number tandem-repeat analysis forYersinia enterocoliticasubsp.palearcticaand its application to biosero-group 4/O3 subtyping. J. Clin. Microbiol.45:2508 –2515.
21. Lindstedt BA.2011. Genotyping of selected bacterial enteropathogens in Norway. Int. J. Med. Microbiol.301:648 – 653.
22. Wang X, Gu W, Cui Z, Luo L, Cheng P, Xiao Y, Tang L, Kan B, Jing H.
2012. Multiple-locus variable-number tandem-repeat analysis of patho-genicYersinia enterocoliticain China. PLoS One7:e37309. doi:10.1371 /journal.pone.0037309.
23. Gulati P, Varshney RK, Virdi JS. 2009. Multilocus variable number tandem repeat analysis as a tool to discern genetic relationships among strains ofYersinia enterocoliticabiovar 1A. J. Appl. Microbiol.107:875– 884.
24. Laukkanen R, Ortiz Martínez P, Siekkinen K, Ranta J, Maijala R, Korkeala H.2009. Contamination of carcasses with human pathogenic Yersinia enterocolitica4/O:3 originates from pigs infected on farms. Food-borne Pathog. Dis.6:681– 688.
25. Fredriksson-Ahomaa M, Niskanen T, Bucher M, Korte T, Stolle A, Korkeala H.2003. DifferentYersinia enterocolitica4:O3 genotypes found in pig tonsils in Southern Germany and Finland. Syst. Appl. Microbiol.
26:132–137.
26. Fredriksson-Ahomaa M, Autio T, Korkeala H.1999. Efficient subtyping ofYersinia enterocoliticabioserotype 4/O:3 with pulsed-field gel electro-phoresis. Lett. Appl. Microbiol.29:308 –312.
27. Pitcher DG, Saunders NA, Owen RJ.1989. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Lett. Appl. Microbiol.8:151– 156.
28. Hunter PR, Gaston MA.1988. Numerical index of the discriminatory ability of typing systems: an application of Simpson’s index of diversity. J. Clin. Microbiol.26:2465–2466.
29. Virtanen S, Salonen L, Laukkanen-Ninios R, Fredriksson-Ahomaa M, Korkeala H.2012. Piglets are a source of pathogenicYersinia enterocolitica on fattening-pig farms. Appl. Environ. Microbiol.78:3000 –3003. 30. Benitez AJ, Diaz MH, Wolff BJ, Pimentel G, Njenga MK, Estevez A,
Winchell JM.2012. Multilocus variable-number tandem-repeat analysis ofMycoplasma pneumoniaeclinical isolates from 1962 to the present: a retrospective study. J. Clin. Microbiol.50:3620 –3626.
31. Cooley MB, Carychao D, Nguyen K, Whitehand L, Mandrell R.2010. Effects of environmental stress on stability of tandem repeats in Esche-richia coliO157:H7. Appl. Environ. Microbiol.76:3398 –3400.
32. Vogler AJ, Keys CE, Allender C, Bailey I, Girard J, Pearson T, Smith KL, Wagner DM, Keim P.2007. Mutations, mutation rates, and evolution at the hypervariable VNTR loci ofYersinia pestis. Mutat. Res.616:145–158. 33. Batzilla J, Hoper D, Antonenka U, Heesemann J, Rakin A. 2011.
Complete genome sequence ofYersinia enterocoliticasubsp.palearctica serogroup O:3. J. Bacteriol.193:2067.
34. Noller AC, McEllistrem MC, Shutt KA, Harrison LH. 2006. Locus-specific mutational events in a multilocus variable-number tandem repeat analysis ofEscherichia coliO157:H7. J. Clin. Microbiol.44:374 –377. 35. Noller AC, McEllistrem MC, Pacheco AG, Boxrud DJ, Harrison LH.
2003. Multilocus variable-number tandem repeat analysis distinguishes outbreak and sporadicEscherichia coliO157:H7 isolates. J. Clin. Micro-biol.41:5389 –5397.