Joint Effects of Pleiotropic Selection and Stabilizing Selection on the
Maintenance of Quantitative Genetic Variation at Mutation-Selection Balance
Xu-Sheng Zhang
1and William G. Hill
Institute of Cell, Animal and Population Biology, University of Edinburgh, Edinburgh EH9 3JT, United Kingdom
Manuscript received April 16, 2002 Accepted for publication June 10, 2002
ABSTRACT
In quantitative genetics, there are two basic “conflicting” observations: abundant polygenic variation and strong stabilizing selection that should rapidly deplete that variation. This conflict, although having attracted much theoretical attention, still stands open. Two classes of model have been proposed: real stabilizing selection directly on the metric trait under study and apparent stabilizing selection caused solely by the deleterious pleiotropic side effects of mutations on fitness. Here these models are combined and the total stabilizing selection observed is assumed to derive simultaneously through these two different mechanisms. Mutations have effects on a metric trait and on fitness, and both effects vary continuously. The genetic variance (VG) and the observed strength of total stabilizing selection (Vs,t) are analyzed with
a rare-alleles model. Both kinds of selection reduceVGbut their roles in depleting it are not independent:
The magnitude of pleiotropic selection depends on real stabilizing selection and such dependence is subject to the shape of the distributions of mutational effects. The genetic variation maintained thus depends on the kurtosis as well as the variance of mutational effects: All else being equal,VGincreases
with increasing leptokurtosis of mutational effects on fitness, while for a given distribution of mutational effects on fitness,VGdecreases with increasing leptokurtosis of mutational effects on the trait. TheVGand
Vs,tare determined primarily by real stabilizing selection while pleiotropic effects, which can be large,
have only a limited impact. This finding provides some promise that a high heritability can be explained under strong total stabilizing selection for what are regarded as typical values of mutation and selection parameters.
T
HE presence of genetic variation in quantitative genes that affect the trait per generation per haploid genome, andVs,ris the strength of real stabilizingselec-traits is important for the selective breeding of
do-tion, the “variance” of the fitness profile, with a large mestic animals and crops, evolution, and adaptation
value of Vs,r implying weak selection. It is difficult to
(Charlesworthet al.1982;BartonandTurelli1989;
account for the observed high variance with this model FalconerandMackay1996;BartonandKeightley
for what are regarded as typical values ofVs,r(e.g., 20Ve),
2002). The existence of genetic variation is, however,
mutation rate per locus, and number of relevant loci paradoxical because stabilizing selection acting on the
(Turelli1984;FalconerandMackay1996). Further-population usually depletes genetic variation (Wright
more, simple genetic load arguments suggest that real 1935;CrowandKimura1970;Bu¨ rgerandGimelfarb
stabilizing selection cannot operate independently on 1999;Bu¨ rger2000). As the ultimate source of genetic
many characters (Robertson1967;Turelli1985; Bar-variation is mutation, an intuitively appealing
explana-ton 1990). In a recent review, however, Kingsolver tion for the maintenance of polygenic variation is that
et al. (2001) found that estimates of the strength of there is an equilibrium between the input of new
varia-stabilizing selection vary greatly, and the typical selec-tion by mutaselec-tion and its erosion by natural selecselec-tion.
tion may be much weaker than previously assumed. For real stabilizing selection it is assumed that natural
In an alternative model, the purepleiotropic model, selection acts directly and solely on the metric trait,
natural selection is assumed not to act directly on the relative fitness having a quadratic relationship with the
metric trait in question, but through pleiotropic side trait. Under the rare-allele model and the assumption
effects of mutant alleles on fitness (Robertson 1967; of Gaussian fitness function, predictions for the
equilib-Hill andKeightley1988). This model can generate rium genetic variance are given by the house-of-cards
apparent stabilizing selection, shown as a negative corre-approximation VG ⫽ 4tVs,r (Turelli 1984; Bu¨ rger
lation between relative fitness and phenotypic deviation 2000), wheretis the average number of mutations of
from the mean (Robertson1967;HillandKeightley 1988; Barton 1990; Gavrilets and de Jong 1993): Extreme individuals for the trait tend to carry more
1Corresponding author:Institute of Cell, Animal and Population
Biol-harmful mutations. There is observational evidence for
ogy, University of Edinburgh, W. Mains Rd., Edinburgh EH9 3JT,
United Kingdom. E-mail: [email protected] apparent selection in nature (Kruuket al.2002). This
model can provide an explanation for observed levels though the assumption of an equal fitness effect for all mutations is a convenient way to obtain analytical ap-ofVGbut for only part of the strength of apparent
stabi-lizing selection observed (Vs,t) and has the further defect proximations forVGandVs,t(Barton1990;
Kondras-hovandTurelli1992;Tanaka1996), it lacks rigorous thatVGincreases without bound as the effective
popu-lation size increases when the mutational effects are not support, and experimental data illustrate the highly lep-tokurtic distribution of mutational effects on fitness completely correlated and the distribution of fitness
ef-fects is leptokurtic (KeightleyandHill1990;Cabal- (Mackayet al.1992). As shown byZhanget al.(2002), the shape of the distribution of mutational effects does lero andKeightley1994). Such stabilizing selection
induced solely by pleiotropic effects on fitness of mu- affect the predictions of the pleiotropic model, so that it is necessary to take into account variation in effects tations is referred to as “pleiotropic selection” in this
article. There is a general relationshipVsⱖVG2/Vm(Bar- of mutations both on the trait and on fitness.
In this study, a compound model of continuously vary-ton1990;KondrashovandTurelli1992;Gavrilets
and de Jong 1993; Zhang et al. 2002); therefore the ing effects of mutations on the trait and on fitness is constructed to investigate the maintenance of genetic pure pleiotropic model cannot in principle explain both
the observed levels of genetic variances and typical es- variance and the observed strength of total stabilizing selection. The interaction between both kinds of selec-timates of strengths of stabilizing selection, provided
the mutational variance Vm is of the order 10⫺3Ve as tion and their overall impact on genetic variation and
strength of total stabilizing selection are explored. We observed (Houle et al.1996;LynchandWalsh1998;
Lynchet al.1999). hope thereby to provide a possible explanation for the
observations of both high genetic variance and the In addition to the above two hypotheses, many others
such as overdominance (Wright 1935; Robertson strong observed stabilizing selection. 1956;Gillespie1984;Barton1990),
frequency-depen-dent selection (Slatkin1979;Barton1990),
genotype-MODEL
by-environment interaction (Gillespie and Turelli
1989; Gimelfarb 1990; Zhivotovsky andGavrilets We assume additivity of gene action, linkage equilib-rium, a random-mating diploid population, and rare 1992), and epistatic interaction (Zhivotovskyand
Gav-rilets1992;Gavriletsandde Jong1993) have been mutant alleles. In accordance with the model of real stabilizing selection (Turelli1984, 1985), the relative proposed to explain the maintenance of polygenic
varia-tion. All these models have their respective appeal and fitness of individuals that have a phenotypic value P, the sum of the contributions from each locus plus a weaknesses in explaining the maintenance of polygenic
variation. random independent environmental effect of mean
zero, is assumed to be given byW(P)⫽exp(⫺P2/22).
Nevertheless, the real stabilizing selection and
pleio-tropic models are not mutually exclusive. Individual The mean fitness of individuals with genotypic value G⫽兺iaiisW(G)⫽exp(⫺G2/2Vs,r)⬇1⫺G2/2Vs,rwith
mutant alleles can have both deleterious pleiotropic
effects on fitness and effects on the metric trait in ques- Vs,r⫽ 2 ⫹ Vemeasuring the intrinsic strength of real
stabilizing selection. Ve is the environmental variance
tion (FalconerandMackay1996). If the metric trait
is not completely neutral, that is, the extreme pheno- and is scaled as a unit of variance.
It is assumed that there are infinitely many loci on types of the metric trait are less fit, natural selection
takes place simultaneously through two different mech- each individual and at each locus there is a continuum of possible mutational effects, but each locus has the anisms: the deleterious pleiotropic effects on all other
aspects of fitness and real stabilizing selection on the same mutation distribution and loci are exchangeable. There are at most two alleles segregating at each locus: metric trait under study. Individuals that carry mutants
are therefore selected against because of both deleteri- the wild type, which is assumed to be at optimum, and the mutant. Mutations have effects on a metric trait (a) ous pleiotropic effects of mutants (i.e., pleiotropic
selec-tion) and phenotypic deviations of the trait value from and pleiotropic deleterious effects on fitness (s ⱖ 0), with a bivariate distributionh(a,s). If the metric trait the optimum (i.e., real stabilizing selection). The strength
of total stabilizing selection is therefore attributed to undergoes real stabilizing selection due to mutations, the observed stabilizing selection would come from both kinds of natural selection. As Kondrashov and
Turelli(1992, p. 615) noted, “A complete treatment these two parts and the equivalent total selection coeffi-cient within each individual is given by s˜⫽ s ⫹ (1 ⫺ should consider both direct and indirect selection on
the quantitative trait.” Tanaka (1996) used a cohort- 2x)a2/(4V
s,r) (seeappendix a), wherexis the frequency
of the mutant allele. The equivalent total selection coef-of-mutations model to combine both pleiotropic and
real stabilizing selections and assumed that all mutations ficient is in general not independent of the frequency of mutant alleles in this compound model. It is therefore had an equal deleterious effect on fitness and a Gaussian
distribution of effects on the target trait. However, this less tractable (seeappendix a) than the pure pleiotropic model (Barton 1990;Keightleyand Hill1990), in cannot readily account for both high heritabilities and
pleiotro-pic effect on fitness of each mutant allele and the coeffi- and Walsh1998; Lynch et al. 1999; and this value is used in this study). ThereforeVs,rcannot be very large
cient is always independent of the frequency of the
if a high VG is to be maintained under strong total
mutant allele. With the assumption of real stabilizing
stabilizing selection (i.e., smallVs,t).
selection (Turelli1984; Keightleyand Hill1988),
Although the properties of mutant effects on the met-however, selection acts on the total effect of all mutants
ric trait and on fitness are crucial to evaluatingVGand
within individuals and hence depends on the frequency
Vs,t, the distribution of mutational effects is hard to
esti-of mutant alleles (Robertson 1956). Such frequency
mate accurately (MackayandLangley1990;Hilland dependence of selection leads to multiple equilibria
Caballero1992;Mackayet al.1992;Davieset al.1999; (Bulmer 1985; Barton1986) but, unless population
ElenaandMoya1999;Keightleyet al.2000;Shawet size is very small, mutant alleles cannot increase to a
al.2000; ImhofandSchlo¨ tterer2001;Wloch et al. high frequency without passing through an
intermedi-2001). Even for Drosophila, for which there are many ate frequency, against which there is selection. The
fre-studies, the data seem to suggest a highly skewed and quency of mutant alleles therefore remains very low
leptokurtic distribution of mutational effects (Mackay (Bulmer1989). With rare mutant alleles, the equivalent
andLangley1990;HillandCaballero1992;Mackay total selection coefficient within each individual
organ-et al. 1992), but fine-scale information is still lacking. ism can therefore be approximated by
As inKeightleyandHill(1990), the distribution of s˜⫽s⫹a2/(4V
s,r) . (1) mutant effects on the metric trait is assumed to be
sym-metrical about a ⫽ 0, and only deleterious effects of In an infinite population the equilibrium genetic vari- mutations on fitness are assumed to occur, in accord
ance is with the classical view (FalconerandMackay1996).
The variability of the distribution ofais defined in terms ofεa⫽
√
E(a2) and fors isεs⫽√
E(s2). For theoreticalVG⫽ 4Vs,r
冮
∞
⫺∞
冮
∞
0
h(a,s)a
2/(4V s,r)
s˜ dads ⫽4Vs,rI2, (2) comparison, it is assumed in this study that mutational effects on the trait are, in increasing order of lepto-in whichI2 is determined by the distribution of muta- kurtosis, Gaussian, reflected gamma (1⁄
2), reflected
tional effects (see appendix b), is the genome-wide gamma (1⁄
4), reflected gamma (1⁄8), reflected squared
mutation rate over all loci, and the strength of total gamma (1⁄
2), and reflected quartic gamma (1⁄2);
muta-stabilizing selection (i.e., that which would be observed tional effects on fitness are equal, one-sided Gaussian,
regardless of its source) is gamma (1⁄
2), gamma (1⁄4), gamma (1⁄8), squared gamma
(1⁄
2), and quartic gamma (1⁄2), where gamma () denotes
the gamma distribution with shape parameter. Those Vs,t⫽
V2
G/Vs,r ⫹Vm⫺Covp
V2
G/Vs,r⫹ Vm
Vs,r. (3)
distributions, whose shapes are illustrated in Figure 1, cover a very wide range of all possible mutational effects. Here Covp is the covariance of relative fitness and
squared deviation due to pleiotropic effects on fitness
of mutations (see appendix a). When the pleiotropic RESULTS
selection is much stronger than real stabilizing
selec-Analytical approximations are obtained for some spe-tion, Covp→ Vm (cf.Bu¨ rger2000;Zhang et al.2002)
cial cases for an infinite population and a rare-allele and the strongest total selection applies with strength
approximation, and numerical calculations were per-Vs,t ⫽ VG2/(V2G/Vs,r ⫹ Vm)⬅ Vbs,t; when the pleiotropic
formed to provide support and to extend the results to effect is very weak in relation to real stabilizing selection,
more general situations. Simple results for some special Covp→0 and the strength of total stabilizing selection
situations are also presented within Keightley and approachesVs,t. In general, the following inequality
ap-Hill’s (1990) framework usingKimura’s (1969) diffu-plies for the strength of total stabilizing selection:
sion approximation.
Pure real stabilizing selection within a finite popula-Vb
s,t⬍Vs,t⬍ Vs,r. (4)
tion,i.e.,s⫽0, thus˜s⫽(1⫺2x)a2/(4Vs,r):The observed
Because the total covariance of relative fitness and strength of real stabilizing selection isVs,t⫽VG2/(2 Covr)⫽
squared deviation, Cov⫽ Vm ⫹ V2G/Vs,r, is larger than Vs,r and the genetic variance is given by (A2).
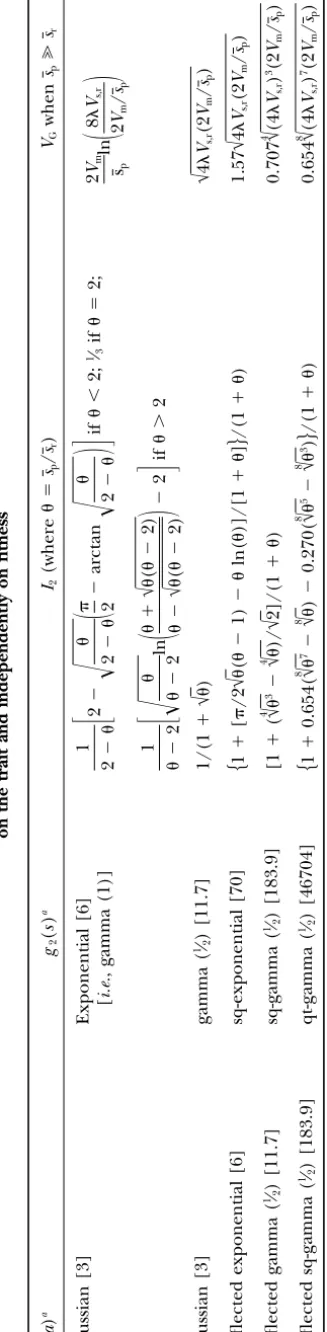
Numeri-that both for the pure pleiotropic model,Vm(Bu¨ rger cal calculation shows that, as the effective population
2000;Zhanget al. 2002), and for real stabilizing selec- sizeNeincreases,VGincreases and approaches the
rare-tion,VG2/(2Vs,r), the total stabilizing selection is certainly allele approximation 4Vs,r (Turelli 1984; see Figure
stronger than either individual component. The muta- 2). Theoretically this is because the equilibrium fre-tional variance on the traitVm⫽1⁄2ε2a, whereε2a is the quencies of mutant genes,x, become very small and the
variance of mutational effects on the trait, is observed heterozygosity can thus be approximated byH(s˜)⫽4/ s˜ as the effective population size Ne → ∞ and thus
to be of the order 10⫺3V
Figure 1.—The distribution of mutation ef-fects,s, on fitness used in the study withE(s2)⫽
1.0 in each case. Here sq-gamma (1⁄
2) and
qt-gamma (1⁄
2) represent squared gamma (1⁄2) and
quartic gamma (1⁄
2). The distribution of
muta-tional effects,a, on the trait was symmetrical about
a⫽0, but with that of |a| having the same range of distribution ass.
Nes˜Ⰷ 1. Figure 2 also shows that the genetic variance andHill1990;CaballeroandKeightley1994;
Cha-varras et al. 2001; Wloch et al. 2001). If, however, maintained in a finite population depends on the
distri-bution of mutational effects (cf.Keightley andHill the pleiotropic effects vary among mutants, substantial variation can occur; indeedVGbecomes unbounded for
1988). Further, if mutational effects on the trait follow a reflected gamma (1⁄
2),VGdepends little on the mutation an infinite population if neutral mutants predominate
(seeKeightleyandHill1990;Caballeroand Keight-rates; whereas for equal mutational effects on the trait,
VGin small populations depends heavily on the mutation ley1994;Zhanget al.2002). Moreover, the pure
pleio-tropic model can only partially account for the “typical” rates for givenVs,r.
Pure pleiotropic effects, where the target trait is com- strength of stabilizing selection (Barton 1990;
Kon-drashovandTurelli1992;Zhanget al.2002).
pletely neutral in itself (i.e., Vs,r →∞) ands˜⫽ s:With
all mutants having equal pleiotropic effects, the genetic Joint effects of both pleiotropic and real stabilizing selections within an infinite population, but assuming variance is VG ⫽ 2Vm/s (Barton 1990), which is too
small, given that the estimates of selection coefficients equal mutational effects on both the trait (εa) and fitness (s):From Equation 1,s˜⫽s⫹ε2
a/(4Vs,r) and the
approxi-with detectable effects in the laboratory are in the range
s ⫽ 0.02–0.08 (Crow andSimmons 1983; Keightley mation H(s˜) ⫽ 4/s˜ for an infinite population, the
Figure2.—Genetic variance maintained in the metric trait as a function of the effective popula-tion size under pure real stabilizing selecpopula-tion (i.e.,
s⫽0). Two cases are investigated: ⫽0.01,Vs,r⫽
10 and ⫽0.001,Vs,r⫽100. Results are shown
for two distributions of mutational effects on the trait: gamma (1⁄
2)-distributed effects (solid lines,
genetic variance is given by highly leptokurtically distributed effects of mutations on the trait lead to a low genetic variance (see also VG⫽H(s˜)ε2a/4⫽ 2Vm/(s⫹ ε2a/(4Vs,r)) . (5) Figure 2 for finite populations),Tanaka’s (1996)
for-mula gives a good approximation for the situation in As the fourth moment and covariance arem4⫽VGε2a/4
which sⰇ sr. The numerical results in Figure 3 show
and Covp⫽VGs/2, the strength of total stabilizing
selec-that expression (5) provides a close approximation to tion (real and apparent) is
VGwhenis either⬎10⫺2or⬍10⫺6for Gaussian effects
of mutations on the trait or when ⬎ 0.1 for gamma Vs,t⫽(ε2a/4⫹ 2VG)/[s⫹(ε2a/4⫹2VG)/Vs,r] (6)
(1⁄
2) effects of mutations. For other values of mutation
(seeappendix a). Equation 6 is a special case of Equa- rate, however,Tanaka’s (1996) results are much larger tion 3. With no pleiotropic effect of mutants (i.e.,s⫽ than numerical results for both Gaussian and reflected 0), selection comes solely from real stabilizing selection gamma (1⁄
2) mutational effects. When ⫽10⫺4, for
ex-on the metric trait, Vs,t ⫽ Vs,r; with some pleiotropic ample, (5) gives VG ⫽ 0.028, which is ⵑ1.5 and 2.3
effects on fitness, selection becomes stronger (i.e., Vs,t times as large as the numerical results for Gaussian and
decreases). The inclusion of a pleiotropic deleterious reflected gamma (1⁄
2) mutational effects, respectively.
effect therefore decreases both the genetic variance and Figure 3 clearly shows how both effects interfere and the strength of total stabilizing selection. Equation 5 is contribute to the overall outcome inV
GandVs,t. When
the same as that ofTanaka(1996) who assumed equal the mutation rate is very low (e.g., ⬍10⫺5) and each
deleterious effects of mutations on fitness but Gaussian mutant has large effects on the trait relative to its effect effects on the metric trait. In this case, the population on fitness, the results approach the house-of-cards ap-average of the total selection coefficient is simply equal proximation (Turelli 1984). If the mutation rate is to the sum of the selection coefficients due to both high (e.g., ⬎ 0.1) and each mutant has a relatively kinds of selection (cf.KondrashovandTurelli1992; small effect on the trait, the pleiotropic effect must be
Tanaka1996, 1998). widespread and becomes the main force of selection,
Joint effects, but assuming mutations have an equal the genetic variance tends to that of Barton (1990)
pleiotropic effect on fitness (s) and a continuous distri- but the strength of total stabilizing selection approaches
butionf(a) of mutational effects on the metric trait:In V
s,t⫽ (V2G/Vm)/[1⫹2VG/(sVs,r)], which is smaller than
this situation Kimura’s (1969) diffusion theory leads that ofBarton(1990). If the mean pleiotropic effect to H(s˜) ⫽ C(s˜)⫽ 4/s˜approximately, and K(s˜) ⫽ 0 is stronger than that of real stabilizing selection, i.e., approximately for an infinite population (Zhanget al. s ⬎ s
r, expression (7) can give better approximations
2002). The genetic variance isVG⫽ 兰∞⫺∞[a2/(s⫹ a2/ forV
GthanTanaka’s (1996). One interesting
phenome-(4Vs,r))]f(a)da, and the strength of total stabilizing selec- non can be noted by comparingV
GandVs,tin Figure 3:
tion is given by (A7), where the fourth moment ism4⫽ VG rises as the mutation rate increases while the total
兰∞
⫺∞((a4/4)/(s⫹a2/(4Vs,r)))f(a)da, and the covariance stabilizing selection becomes stronger (i.e., Vs,t
de-between relative fitness and squared deviation due to creases). This is in sharp contrast to both real stabilizing pleiotropic effects is Covp⫽VGs/2. In the following we selection, where asincreasesV
Gincreases butVs,t(⫽
denote the population mean of the selection coeffi- V
s,r) remains unchanged (Turelli1984), and the pure
cients arising from real stabilizing selection by sr⬅ pleiotropic model, where asincreasesVGremains
un-E(a2/4V
s,r)⫽ε2a/4Vs,r⫽2Vm/(4Vs,r),i.e., twice the ratio changed butVs,tdecreases (Barton1990;cf.Figure 5,
of mutational variance to the genetic variance main- c and d below).
tained in real stabilizing selection. For a neutral trait, General case:As shown above and by previous work
sr⫽0. (Barton1990;KondrashovandTurelli1992;Tanaka
If the mean pleiotropic effect on fitness is much 1996), the equal fitness effect assumption cannot pro-weaker than that from real stabilizing selection (i.e.,sⰆ vide a simultaneous explanation for the observed high sr), the genetic variance approaches the rare-allele ap- heritability and strong stabilizing selection. If
muta-proximation VG ⫽ 4Vs,r. In general VG ⫽ 4Vs,r兺∞i⫽1 tional effects on fitness vary across loci in the absence
(⫺1)i⫺1E(a2i)/(4sV
s,r)i (Moran 1968, p. 296). Ifs Ⰷ sr, of real stabilizing selection a huge genetic variance can
the genetic variance can be approximated by be generated (KeightleyandHill1990;Zhanget al.
2002), so it is important to investigate the influence of VG⬇ 2Vm/(s⫹ 4sr) . (7)
variation in fitness effects onVGandVs,t.
The first check is whether the unbounded VG with
Noting that the kurtosis4⬅E(a4)/E2(a2)⫽1, 3, and
increasing population size is avoided with the inclusion 35/3 for effects that are equally distributed, normally
of a real stabilizing selection on the trait. The example distributed, and distributed as a gamma (1⁄
2),
respec-in Figure 4 shows that with even a weak real stabilizrespec-ing tively, Tanaka’s (1996) formula (i.e., Equation 5) is
selection (e.g., Vs,r ⫽ 1000), the genetic variance
in-therefore accurate only for equal mutational effects on
Figure3.—(a) Genetic variance maintained in the metric trait and (b) the strength of stabilizing selection on it as functions of the mutation rate. Mutational effects on the trait follow either a Gaussian or a reflected gamma (1⁄
2) distribution
or are equal (|a|⫽ εa⫽√2Vm/) and those on
fitness are equal (s⫽0.02). The effective popula-tion size is infinite andVs,r⫽100. The curves for
equal are given by Equations 5 and 6.
but asymptotes whenNeexceeds some large value. This 4Vs,r; while forspⰇsr, the genetic variance reduces to
asymptotic value ofVGdepends on the value ofVs,r, with
VG⫽
√
4Vs,r(2Vm/sp) . (9)a high VG for a weak real stabilizing selection (i.e., a
largeVs,r). At the same time, the value ofVs,talso increases This is the geometric mean of the genetic variance
main-and approaches a limit that is less thanVs,r. This implies tained by real stabilizing selection (Turelli1984) and
that selection becomes weaker as the effective
popula-for the pleiotropic model with equal fitness effects ( Bar-tion size increases, but the total stabilizing selecBar-tion is
ton1990; Kondrashov and Turelli1992). This ge-stronger than the real stabilizing selection. netic variance approaches infinity if the metric trait is
Suppose that mutational effects on the trait are Gaus- neutral (i.e.,
Vs,r →∞) (cf.KeightleyandHill1990),
sian and mutational effects on fitness follow a gamma consistent with the results shown in Figure 4a. Equation (1⁄
2) with mean sp⫽ E(s)⫽ εs/
√
3. If these mutational 8 clearly shows that both kinds of selection reduce V Geffects are independent, the genetic variance for an but the impact of pleiotropic selection depends on the infinite population can be expressed exactly as magnitude of real stabilizing selection. This unequal
influence of both kinds of selection on the genetic varia-VG⫽ 4Vs,r/(1⫹
√
sp/sr)⫽ 2Vm/(sr ⫹√
srsp) (8)tion is due to the fact that large pleiotropic effects on fitness can induce only a high fitness deficit whereas (seeappendix b), in which spis the population mean
of selection coefficients due solely to pleiotropic effect large effects on the trait can lead to a high genetic variance as well as a high fitness deficit. If the total on fitness and sr, as in (7), is due to real stabilizing
selection. For an extreme situation where the pleiotro- selection coefficient were defined as the ratio of muta-tional variance to the equilibrium genetic variance fol-pic effect is very weak (i.e., sp Ⰶ sr), (8) tends to the
Figure 4.—(a) Variation maintained in the metric trait and (b) the strength of real stabilizing selection as functions of the effective population sizeNe. BothVGandVs,tare evaluated by Monte
Carlo integration. Absolute values of mutational effects on the trait (|a|) and on fitness (s) are independent and both marginal distributions are gamma (1⁄
2). Parameters of mutations ⫽0.1 and εs ⫽ 0.1. Results are shown for three intrinsic
strengths of real stabilizing selection,Vs,r.
(1992), andTanaka(1998), (8) implies that the expec- are Gaussian, or gamma (1⁄
2), or gamma (1⁄4),
respec-tively, leads to the conclusion that a more leptokurtic tation of the total selection coefficient is not simply the
sum of both components, but a function, sT ⫽ sr ⫹ distribution of mutational effects on the trait induces a
smaller genetic variance given the same distribution of
√
srsp. Further results are listed in Table 1.mutational effects on fitness and the same other proper-Numerical results are shown in Figure 5 for a range
ties (cf. Keightley and Hill 1988). Figure 5b shows of distributions of effects of mutations on the trait and
that an increase in pleiotropic selection (εs) leads to an
on fitness such as equal, Gaussian, gamma (1⁄
2), gamma
increase in total stabilizing selection (i.e., decreasing (1⁄
4), and gamma (1⁄8) (except symmetrical foraand
one-Vs,t). For equal mutation effects, Figure 5d shows that
sided fors). With all other properties being the same,
an increase in mutation rate can induce stronger total Tanaka’s (1996) formula (i.e., Equation 5 for equal
stabilizing selection, while for other distributed muta-effects |a| and s) predicts the smallestVG andVs,t (i.e.,
tion effects there is a value of mutation rate at which the strongest selection). Further, the genetic variance
the total stabilizing selection is strongest. This behavior maintained increases and total stabilizing selection
be-of Vs,t may differ from the pure pleiotropic model
comes weaker as mutational effects on fitness become
(KeightleyandHill1990;Zhang et al.2002). more leptokurtic (see also Table 1). This, albeit in
agree-In a realistic model, mutational effects on the trait ment with the conclusion drawn byZhanget al.(2002),
differs from situations when the pleiotropic effect is and on fitness must be correlated (KeightleyandHill 1990). Although analytical treatment is never easy (if assumed to be equal (see Figure 3). Comparison of the
three curves in Figure 5a in which mutational effects possible) when a correlation between mutational effects is included (e.g.,Turelli1985), it is important to con-on fitness follow the gamma (1⁄
sider the impact of such a correlation on the results for VGandVs,t. Using the method ofKeightleyandHill
(1990), the mutational effects |a| and s were sampled from a bivariate gamma (1⁄
2) distribution. The numerical
calculations show that when this correlation is only inter-mediate ( ⬍0.5), its impact onVGandVs,ris not large
(see Figure 6). Unless the correlation between |a| and s is very high, the results based on the assumption of independent mutational effects apply approximately.
DISCUSSION
The assumptions for the origin of both kinds of selec-tion are distinct. In models of real stabilizing selecselec-tion, selection is assumed to arise solely from the deviations of the metric traits from their optimum due to mutational effects (i.e., phenotypic selection, selection directly act-ing on the trait), whereas in pure pleiotropic models the apparent stabilizing selection is assumed to arise as a consequence of direct effects of deleterious mutations on overall fitness, ignoring any effect on the trait itself (i.e., selection acting directly on genes). By assuming that the total stabilizing selection observed on individu-als comes simultaneously from both kinds of selection, the joint effect model presented in this article includes the properties of both the real stabilizing selection (Turelli 1984) and the pure pleiotropic models (Keightleyand Hill 1990). It is important to know whether new findings about the genetic variation and the stabilizing selection emerge from the analyses of the joint effect model.
The pure pleiotropic model (Keightley and Hill 1990; Kondrashov and Turelli 1992; Zhang et al. 2002) can account for substantial quantitative genetic variation, but the apparent stabilizing selection of strengthVs,t⬎V2G/Vmseems too weak. Moreover, a defect
of such a model with continuously varying and leptokur-tic mutational effects is that the geneleptokur-tic variance keeps increasing with the population effective size ( Keight-leyandHill1990;CaballeroandKeightley1994), although this can be avoided by assuming there is a minimum mutational effect (Zhanget al.2002). Such divergence ofVGas population size becomes infinite is
actually an artifact of allowing a density of mutations that are precisely neutral. Equations 3 and 4 show clearly that when V2
G/Vs,r Ⰷ Vm, which might usually be true
(Endler 1986; Falconer and Mackay 1996), the strength of total stabilizing selection is significantly less than the constraintV2
G/Vmimposed by the pure
pleiotro-pic model. Therefore the complete treatment of both pleiotropic effects and real stabilizing selection breaks down the constraint between Vs,t and VG in the pure
pleiotropic model (Barton 1990; Kondrashov and Turelli1992;Zhanget al.2002). With weak real stabi-lizing selection on the trait, it is easy to produce a high VGbut the total stabilizing selection also appears weak,
whatever the pleiotropic effect. For there to be strong
TABLE 1 Exact results of I2 ⫽ E [ 2/( 2⫹ s )], where ⫽ a / √ 4 Vs,r and the approximations for VG when sp Ⰷ sr for some d istributions of mutational e ffects on the trait and independently on fitness g1 ( a ) a g2 ( s ) a I2 (where ⫽ sp / sr ) VG when sp Ⰷ sr Gaussian [3] Exponential [6] 1 2 ⫺
冤
2 ⫺冪
2 ⫺冢
⫺ 2
arctan
冪
2 ⫺冣冥
if ⬍ 2;1⁄3
if ⫽ 2; [ i.e. , g amma (1)] 2
Vm sp
ln
冢
8 Vs,r 2 Vm / sp冣
1 ⫺ 2冤冪
⫺ 2 ln冢
⫹ √ ( ⫺ 2) ⫺ √ ( ⫺ 2)冣
⫺ 2冥
if ⬎ 2 Gaussian [3] gamma (1⁄)2
[11.7] 1/(1 ⫹ √ ) √ 4 Vs,r (2 Vm / sp ) Reflected e xponential [6] sq-exponential [70]
兵
1 ⫹ [ /2 √ ( ⫺ 1) ⫺ ln( )]/[1 ⫹ ]其
/(1 ⫹ ) 1 .57 √ 4 Vs,r (2 Vm / sp ) Reflected g amma (1⁄)2
[11.7]
sq-gamma
(
1⁄)2
[183.9] [1 ⫹ ( √ 4 3⫺ √ 4 )/ √ 2]/(1 ⫹ ) 0 .707 √ 4 (4 Vs,r ) 3(2 Vm / sp ) Reflected sq-gamma (
1⁄)2
[183.9]
q
t-gamma
(
1⁄)2
[46704]
兵
1 ⫹ 0.654( √ 8 7⫺ √ 8 ) ⫺ 0.270( √ 8 5⫺ √ 8 3)其
/(1 ⫹ ) 0 .654 √ 8 (4 Vs,r ) 7(2 Vm / sp ) asq-exponential, sq-gamma (
1⁄),2
and
q
t-gamma
(
1⁄)2
represent squared exponential, squared gamma (
1⁄)-,2
and
q
uartic
gamma
(
1⁄)-distributed2
mutational effects (one sided for fi tness and symmetrical for the trait), respectively. The numbers in brackets [ ] are the ratios o f the fourth moment to the squared variance, 4 ⫽ E ( a 4)/ ε
4,a
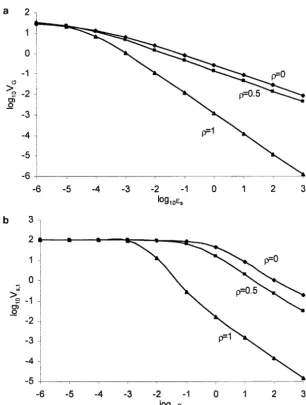
Figure5.—Variation maintained in the metric trait in an infinite population and the strength of real stabilizing selection as functions of (a and b) the pleiotropic effect and (c and d) the mutation rate. The interaction of both effects of selection (pleiotropic effect and real stabilizing selection) has been investigated by either (a and b) fixing the effect of real stabilizing selection (i.e., ⫽0.1) or (c and d) fixing the pleiotropic effect (i.e.,εs⫽0.1). Results are shown forVs,r⫽100. For differently
distributed mutational effects on the trait and on fitness, the same variablitiesεaandεsare assumed, and mutational effects are independent. The symbols eq, gs, gm1/2, gm1/4, and gm1/8 represent equally, Gaussian-, gamma (1⁄
2)-, gamma (1⁄4)-, and gamma
(1⁄
8)-distributed mutational effects (one sided for fitness and symmetrical for the trait), respectively.
total stabilizing selection, the real stabilizing selection and Kondrashov and Turelli (1992, p. 615) con-cluded that real stabilizing selection was “essentially ir-should be strong enough while the impact of pleiotropic
effects is relatively small [see inequality (4)]. relevant to the dynamics of the alleles responsible for variation in the trait.” Within the joint effect model, In contrast to Tanaka’s (1996, 1998) pleiotropic
model, which includes both kinds of selection but as- the total selection coefficient, which is a complicated function of both components [see (8) and appendix sumes an equal deleterious effect on fitness for all
mu-tants, the joint effect model presented here, which b], is⬎sr, butⰆspas well ifspⰇsr. This of course leads
to a largerVG. Therefore pleiotropic effects on fitness
allows both mutational effects to vary, leads to quite
different pictures of how both kinds of selection are can be large but their impact onVGis limited.
For a simple explanation of why a distribution of responsible for VG. As found by Tanaka (1996) and
intuitively argued byKondrashovandTurelli(1992), pleiotropic effects allows the model to generate high VG, suppose that new mutations are divided into two
the total selection coefficient should be equal to a linear
sum of that arising from real stabilizing selection and equally possible classes: one with equal pleiotropic effect s1, the other withs2, but with both having the same effect
that solely attributable to pure pleiotropic effect:sT⫽
sr⫹sp. As in generalspⰇsr(Gillespie1991;Kondra- on the trait (i.e.,sr). The two classes contribute toVGas
2Vm/(s1 ⫹ sr) and 2Vm/(s2 ⫹ sr), respectively, from
shovandTurelli1992), the total selection coefficient
main-Figure 6.—The influence of the correlation () between absolute values of mutational effects on the metric trait (|a|) and on fitness (s) on (a) the genetic variance and (b) observed strength of apparent stabilizing selection. Mutational ef-fects on the trait and fitness follow a bivariate gamma (1⁄
2). The mutation rate is ⫽0.1 and the
intrinsic strength of real stabilizing selection is
Vs,r ⫽ 100. Results are shown for three
correla-tions.
tained is then larger than if all mutations have the same by the house-of-cards approximation (see Equation 8, Table 1, and Figure 5); but if the pleiotropic effect is mean pleiotropic effect (s1 ⫹ s2)/2 because [1/(s1 ⫹
sr)⫹1/(s2⫹sr)]/2⬎1/[(s1⫹s2)/2⫹sr]. The numeri- large, the genetic variance maintained is given by (9)
for Gaussian effects on the trait and gamma (1⁄
2) effects
cal results show that if a very small minimum total
selec-tion coefficient, say 10⫺10, is assumed, the genetic vari- on fitness of mutations. As the genome-wide mutation
rateexceedst, our prediction ofVGmay not be smaller
ance maintained is nearly the same as that without such
than the house-of-cards approximation (cf. Tanaka minimum fitness effect. As the mutant alleles of large
1996, 1998). For the typical estimate of strength of real effects on fitness would be quickly eliminated from the
stabilizing selection,Vs,r⫽20 (Turelli1984),VG⫽0.4
population, the genetic variance is attributable
primar-ily to mildly deleterious mutations. The huge genetic
√
/spfrom (9) under the conditionspⰇ sr(i.e.,spⰇvariation generated in the joint effect model of continu- 2.5⫻10⫺5). This implies that if both the mutation rate
ously varying pleiotropic effects on fitness, therefore, and the mean pleiotropic effect are of similar order, comes mainly from “a class of alleles with significant abundant genetic variation can be maintained, and less effects on the character, but very little effect on fitness” restrictive conditions are required if the mutational
ef-(Barton1990, p. 779). fects are more leptokurtic (see Table 1 and Figure 5).
The mutation rate assumed in this study is the It is also interesting to compare the prediction of the
joint effect model with the house-of-cards approxima- genome-wide mutation rate. Although all of the muta-tions may affect fitness to a varying degree, only a small tionVG⫽4tVs,r, wheretrefers solely to the total rate
of mutations that affect the metric trait under study fraction of them may be considered to appreciably affect the trait under study. It is, however, unrealistic to assume (Turelli1984, 1985). When the pleiotropic effect is
kurtic than on fitness (seeRobertson1967;Keightley trait as low asVm⫽10⫺3Ve(cf.Barton1990). Combining
both kinds of selection and allowing mutational effects andHill1988;HillandCaballero1992). The
analy-ses of the joint effect model show that the genetic vari- on the metric trait and on fitness both to vary change the picture of the mutation-selection model and therefore ance maintained at mutation-selection balance depends
not only on the variance of mutational effects but also enable the mutation-selection balance to be a plausible on their leptokurtosis. For a given distribution of muta- cause of quantitative variation.
tional effects on fitness, a more leptokurtic mutational We are grateful to Nick Barton, Brian Charlesworth, Peter Keightley, effect on the trait induces a smaller genetic variance, Jinliang Wang, and a referee for helpful comments and Ian White for help in proving Equation 8. This work was supported by a grant
consistent with the results of Keightley and Hill
from the Biotechnology and Biological Sciences Research Council
(1988) who studied pure real stabilizing selection in
(R35396).
finite populations. Even for this more realistic model, the joint effect model can still generate abundant ge-netic variation if mutational effects on fitness are suffi-ciently leptokurtic, say gamma (1⁄
2), and the genome- LITERATURE CITED
wide mutation rate is not⬍0.01 (see Figure 5). Barton, N. H., 1986 The maintenance of polygenic variation The scanty data for multicellular eukaryotes are con- through a balance between mutation and stabilizing selection.
Genet. Res.47:209–216.
sistent with any value of between 0.1 and 100
Barton, N. H.,1990 Pleiotropic models of quantitative variation.
(Charlesworthet al.1990;KondrashovandTurelli Genetics124:773–782.
1992; Lynch et al. 1999; Kumar and Subramanian Barton, N. H.,andP. D. Keightley,2002 The nature of quantita-tive genetic variation. Nat. Rev. Genet.3:11–21.
2002). Recent studies onCaenorhabditis elegans, however,
Barton, N. H.,and M. Turelli, 1989 Evolutionary quantitative
show that the mutation rate for life history traits isⰆ1.0 genetics: how little do we know? Annu. Rev. Genet.23:337–370. and is of the order 10⫺3 (Keightleyand Caballero
Bulmer, M. G.,1985 The Mathematical Theorey of Quantitative Genetics.
Clarendon Press, Oxford.
1997; Garcia-Dorado et al. 1999; Vassilieva and
Bulmer, M. G.,1989 Maintenance of genetic variability by
mutation-Lynch1999). The best estimate of the average selection selection balance: a child’s guide through the jungle. Genome coefficient against heterozygous mutations isE[s/2]⫽ 31:761–767.
Bu¨ rger, R.,2000 The Mathematical Theory of Selection, Recombination,
0.02 (Crowand Simmons1983). Data for Drosophila
and Mutation.Wiley, Chichester, UK.
bristle traits show that is in the range 0.09–1.0 and Bu
¨ rger, R.,andA. Gimelfarb,1999 Genetic variation maintained
εs in the range 0.01–0.2 (Keightley and Hill 1990; in multilocus models of additive quantitative traits under stabiliz-ing selection. Genetics152:807–820.
CaballeroandKeightley1994). Data for competitive
Caballero, A.,andP. D. Keightley,1994 A pleiotropic
nonaddi-viability in Drosophila suggest that ⱖ0.01 andE[s]ⱕ
tive model of variation in quantitative traits. Genetics138:883–
0.08 (Chavarriaset al.2001). Data for yeastSaccharo- 900.
Charlesworth, B., R. LandeandM. Slatkin,1982 A
neo-Darwin-myces cerevisiaeshow thatis of the order 10⫺3andE[s/
ian commentary on macroevolution. Evolution36:474–498.
2] is in the range 0.01–0.05 (Wlochet al.2001). Even
Charlesworth, B., D. CharlesworthandM. T. Morgan, 1990
with such large pleiotropic effects, our joint effect Genetic loads and estimates of mutation rates in highly inbred model, which assumes leptokurtic effects both on the plant populations. Nature347:380–382.
Chavarrias, D., C. Lo´ pez-Fanjul and A. Garcı´a-Dorado, 2001
trait and on fitness of mutations, predicts high
heritabil-The rate of mutation and the homozygous and heterozygous
ities under strong total stabilizing selection unlessis mutational effects for competitive viability: a long-term experi-very small, say ⬍0.01 (see Equation 8, Figure 5, and ment withDrosophila melanogaster.Genetics158:681–693.
Crow, J. F.,andM. Kimura,1970 An Introduction to Population
Genet-Table 1). But the estimates of mutation and selection
ics Theory.Harper & Row, New York.
parameters are not very reliable (Kondrashov 1998; Crow, J. F.,andM. J. Simmons,1983 The mutation load in Drosoph-Lynchet al. 1999; Kingsolver et al.2001). Mutation ila, pp. 1–35 inThe Genetics and Biology of Drosophila, edited by
H. L. Carson, M. AshburnerandJ. N. Thompson.Academic
rates are usually underestimated and mean fitness
ef-Press, New York.
fects are usually overestimated as the effects of most Davies, E. K., A. D. PetersandP. D. Keightley,1999 High fre-mutants may be too small to be detected (Kondrashov quency of cryptic deleterious mutations inCaenorhabditis elegans.
Science285:1748–1751.
andTurelli1992;Davieset al.1999;Lynchet al.1999).
Elena, S. F.,andA. Moya,1999 Rate of deleterious mutation and
The observation of high heritabilities and strong total the distribution of its effects on fitness in vesicular stomatitis stabilizing selection may then be interpreted in terms virus. J. Evol. Biol.12:1078–1088.
Endler, J. A.,1986 Natural Selection in the Wild.Princeton University
of the joint effect model of continuously varying
muta-Press, Princeton, NJ.
tional effects.
Falconer, D. S.,andT. F. C. Mackay,1996 Introduction to
Quantita-In summary, the joint effect model presented here tive Genetics, Ed. 4. Longman, Harlow, UK.
Garcia-Dorado, G., C. Lopez-Fanjul and A. Caballero, 1999
shows thatVGandVs,tare determined primarily by real
Properties of spontaneous mutations affecting quantitative traits.
stabilizing selection while pleiotropic effects, which can
Genet. Res.74:341–350.
be large, have only a limited impact. With an abundant Gavrilets, S.,andG. de Jong,1993 Pleiotropic models of polygenic
variation, stabilizing selection and epistasis. Genetics134:609–
supply of mutations and leptokurtic mutational effects
625.
on fitness, the joint effect model can induce a significant
Gillespie, J. H.,1984 Pleiotropic overdominance and the
mainte-amount of stabilizing selection as well as a substantial nance of genetic variation in polygenic characters. Genetics107:
321–330.
Gillespie, J. H.,1991 The Causes of Molecular Evolution.Oxford Uni- Turelli, M.,1984 Heritable genetic variation via mutation-selection balance: Lerch’s Zeta meets the abdominal bristle. Theor. Popul. versity Press, Oxford.
Biol.25:138–193.
Gillespie, J. H.,andM. Turelli,1989 Genotype-environment
inter-Turelli, M.,1985 Effects of pleiotropy on predictions concerning
action and the maintenance of polygenic variability. Genetics
mutation-selection balance for polygenic traits. Genetics 111: 121:129–138.
165–195.
Gimelfarb, A.,1990 How much genetic variation can be maintained
Vassilieva, L. L.,andM. Lynch,1999 The rate of spontaneous
by genotype-environment interactions? Genetics124:443–445.
mutation for life-history traits inCaenorhabditis elegans.Genetics
Hill, W. G., andA. Caballero, 1992 Artificial selection
experi-151:119–129. ments. Annu. Rev. Ecol. Syst.23:287–310.
Wloch, D. M., K. Szafraniec, R. H. BortsandR. Korona,2001
Hill, W. G.,andP. D. Keightley,1988 Interrelations of mutation,
Direct estimate of the mutation rate and the distribution of fitness population size, artificial and natural selection, pp. 57–70 in
effects in the yeastSaccharomyces cerevisiae. Genetics159:441–452.
Proceedings of the 2nd International Conference on Quantitative
Genet-Wright, S.,1935 The analysis of variance and the correlations
be-ics, edited byB. S. Weir, E. J. Eisen, M. M. GoodmanandG.
tween relatives with respect to deviations from an optimum. J.
Namkoong.Sinauer, Sunderland, MA.
Genet.30:243–256.
Houle, D., B. MorikawaandM. Lynch,1996 Comparing
muta-Zhang, X.-S., J. Wang andW. G. Hill,2002 Pleiotropic model
tional variabilities. Genetics143:1467–1483.
of maintenance of quantitative genetic variation at
mutation-Imhof, M.,andC. Schlo¨ tterer,2001 Fitness effects of
advanta-selection balance. Genetics161:419–433. geous mutations in evolving Escherichia coli populations. Proc.
Zhivotovsky, L. A.,andS. Gavrilets,1992 Quantitative variability
Natl. Acad. Sci. USA98:1113–1117.
and multilocus polymorphism under epistatic selection. Theor.
Keightley, P. D.,andA. Caballero,1997 Genomic mutation rate
Popul. Biol.42:254–283. for lifetime reproductive output and lifespan inCaenorhabditis
elegans.Proc. Natl. Acad. Sci. USA94:3823–3827.
Communicating editor:J. B. Walsh
Keightley, P. D.,andW. G. Hill,1988 Quantitative genetic
variabil-ity maintained by mutation-stabilizing selection balance in finite populations. Genet. Res.52:33–43.
Keightley, P. D.,andW. G. Hill,1990 Variation maintained in
APPENDIX A quantitative traits with mutation-selection balance: pleiotropic
side-effects on fitness traits. Proc. R. Soc. Lond. Ser. B242:95–100.
Let us assume that the gene action within and across
Keightley, P. D., E. K. Davies, A. D. PetersandR. G. Shaw,2000
loci is additive and loci are unlinked and in linkage
Properties of ethylmethane sulfonate-induced mutations
affect-ing life-history traits inCaenorhabditis elegansand inferences about equilibrium. A random-mating diploid population is as-bivariate distributions of mutation effects. Genetics156:143–154.
sumed. Mutations in a diploid individual have an effect
Kimura, M., 1969 The number of heterozygous nucleotide sites
on a metric traitzwithathe difference in value between
maintained in a finite population due to steady flux of mutations.
Genetics61:893–903. homozygotes and a net effect on fitness that includes
Kingsolver, J. G., H. E. Hoekstra, J. M. Hoekstra, D. Berrigan,
pleiotropic effects on all other traits, withsthe
differ-S. N. Vignieriet al., 2001 The strength of phenotypic selection
ence in fitness between homozygotes. There is therefore
in natural populations. Am. Nat.157:245–261.
Kondrashov, A. S.,1998 Measuring spontaneous deleterious muta- a bivariate distribution, h(a, s), of a and s for alleles
tion process. Genetica102/103:183–197.
affecting the trait. If there is real stabilizing selection,
Kondrashov, A. S.,andM. Turelli,1992 Deleterious mutations,
the total observed stabilizing selection would come from
apparent stabilizing selection and the maintenance of
quantita-tive variation. Genetics132:603–618. these two parts. Following the method of Falconer
Kruuk, L. E. B., J. Slate, J. M. Pemberton, S. Brotherstone, F. E.
andMackay(1996, p. 27), the mutant allele frequency
Guinnesset al., 2002 Antler size in red deer: heritability and
within a single-locus model is given byx1⫽{x⫺x(1⫺ selection, but no evolution. Evolution (in press).
Kumar, S.,andS. Subramanian,2002 Mutation rates in mammalian x)[s/2⫹a2/(8V
s,r)]}/wwith the mean fitness given by genomes. Proc. Natl. Acad. Sci. USA99:803–808.
w⫽1⫺x(1⫺x)a2/(4V
s,r) if the previous frequency is
Lynch, M.,andB. Walsh,1998 Genetics and Analysis of Quantitative
x.With weak selection (i.e.,w⬇1), the change in the
Traits.Sinauer, Sunderland, MA.
Lynch, M., J. Blanchard, D. Houle, T. Kibota, S. Schultzet al., mutant allele frequency is ⌬x ⫽ x1 ⫺ x ⬇ ⫺x(1 ⫺ x)
1999 Perspective: spontaneous deleterious mutation. Evolution
[s/2 ⫹ (1 ⫺ 2x)a2/(8V
s,r)]. Thus the equivalent total 53:645–663.
selection coefficients are
Mackay, T. F. C.,andC. H. Langley,1990 Molecular and
pheno-typic variation in theachaete-scuteregion ofDrosophila melanogaster.
Nature348:64–66. s˜⫽s⫹ (1⫺2x)a2/(4V
s,r) (A1)
Mackay, T. F. C., R. F. LymanandM. S. Jackson, 1992 Effects
of P elements on quantitative traits in Drosophila melanogaster. (cf. Robertson 1956; Bulmer 1985; Keightley and
Genetics130:315–332. Hill1988). WithKimura’s (1969) diffusion
approxima-Moran, P. A. P.,1968 An Introduction to Probability Theory.Clarendon
tions under the infinite independent loci model, the
Press, Oxford.
Robertson, A.,1956 The effect of selection against extreme devi- genetic variance can be evaluated by the equation
ants based on deviation or on homozygosis. J. Genet.54:236–248.
Robertson, A.,1967 The nature of quantitative genetic variation,
VG⫽
冮
∞
0
冮
∞
⫺∞
冮
1⫺1/(2N)
1/(2N)
h(a,s)⌽(x;s˜)2x(1⫺x)a
2
4dxdads (A2) pp. 265–280 in Heritage from Mendel, edited byA. Brink.The
University of Wisconsin Press, Madison, WI.
Shaw, R. G., D. L. ByersandE. Darmo,2000 Spontaneous
muta-(Zhanget al.2002), the variance of squared deviations is
tional effects on reproductive traits ofArabidopsis thaliana. Genet-ics155:369–378.
VG2⫽
冮
∞
0
冮
∞
⫺∞
冮
1⫺1/(2N) 1/(2N)
h(a,s)⌽(x;s˜)
Slatkin, M.,1979 Frequency- and density-dependency selection on
a quantitative character. Genetics93:755–771.
Tanaka, Y.,1996 The genetic variance maintained by pleiotropic
mutation. Theor. Popul. Biol.49:211–231. ⫻兵2x(1⫺x)⫺3[2x(1⫺x)]2其a 4
16dxdads⫹2V
2
G⬅m4⫹2V2G,
Tanaka, Y.,1998 A pleiotropic model of phenotypic evolution.
and the covariance of relative fitness (taking positive in the same way as that inTurelli(1984) and thus has a dimension of genetic variance.
values because s˜ is defined to be positive) and the
squared deviation is For an infinite population, by using the
approxima-tionsH(s˜)⫽C(s˜)⫽4/s˜andK(s˜)⫽0, the expressions
Cov(w, (z⫺zm)2)⫽
冮
∞
0
冮
∞
⫺∞
冮
1⫺1/(2N) 1/(2N)
h(a,s)⌽(x;s˜) for the genetic variance and strength of the total
stabiliz-ing selection reduce to those given in (2) and (3). Equa-tion 3 is obtained by noting that
⫻2x(1⫺x)(1⫺2x)s 2
a2
4dxdads⫹
VG 2
2Vs,r
.
VG 2⫽4
冮
∞
0
冮
∞
⫺∞
h(a,s)[(a4/16)/
s˜]dads⫹2V2 G
(A4)
Thus the covariance is partitioned into two parts: one
⫽4Vs,r
冮
∞
0
冮
∞
⫺∞
h(a,s)[(a2/4)(s˜⫺s)/s˜]dads⫹2V2 G
due to pleiotropic selection and the other due to stabiliz-ing selection. In the above equations⌽(x;s˜) is the
equi-⫽2Vs,r兵Vm⫺4
冮
∞
0
冮
∞
⫺∞
h(a,s)[(a2/4)(s/2)/s˜]dads⫹V2 G/Vs,r其,
librium frequency distribution of mutations, given by
where Covp⫽4兰∞0 兰∞⫺∞h(a,s)[(s/2)(a2/4)/s˜]dads, and
⌽(x;s˜)⫽ 4Ne x(1⫺ x)G(x)
冮
1 xG()d冮
10G()d
,
the covariance of relative fitness and squared deviation Cov(w, (z⫺zm)2)⫽Covp⫹ VG 2/2Vs,r ⫽Vm⫹V2G/Vs,r.
with G(x)⫽exp
冦
⫺ 2Nex(s⫹(1⫺x)a2
4Vs,r
)
冧
.APPENDIX B
If a population has a large effective sizeNe such that We consider the evaluation of genetic variance
assum-2Nea2/(4Vs,r)Ⰷ1, numerical calculations show that the ing that the population is under stabilizing selection
distribution function⌽(x;s˜) is finite only for very small because of the joint effect of pleiotropic and real stabiliz-values ofx; that is, the equilibrium frequency of mutant ing selections and that both mutational effects are inde-alleles is very small,x ⬇ 0. With the assumption that pendent. If mutational effects on the trait and on fitness the mutant alleles are very rare, the equivalent total follow distributions g
1(a), where⫺∞⬍a⬍∞, and g2(s),
selection coefficient can be approximated by where 0⬍s⬍∞, respectively, then evaluation ofV
G⫽
4Vs,rI2according to (2) is equivalent to the expectation,
s˜⫽s⫹a2/(4V
s,r) . (A5)
Thus the equilibrium genetic variance is I2⫽
冮
∞
⫺∞
冮
∞0
g1(a)g2(s)[2/(2⫹s)]dy1dy2⫽E[2/(2⫹s)] ,
VG⫽
冮
∞
0
冮
∞
⫺∞h(a,s)H(s˜)(a
2/4)dads, (A6)
in which scaled effects on the trait ⫽a/
√
4Vs,raresym-metrical about 0 and distributed with mean 0 and vari-and the observed strength of total stabilizing selection,
ancesr⫽ε2a/4Vs,r. This integral can be obtained exactly
i.e., the variance of the total fitness profile as defined
for some types of mutational effects and the results are byBarton(1990) andKeightleyandHill(1990), is
listed in Table 1, showing thatI2depends on only the
Vs,t⬅⫺VG 2/[2 Cov(w, (z ⫺zm)2)] ratios
p/sr, confirmed by numerical calculations on other
types of mutational effects. The population mean of the
⫽ ⫺(m4 ⫹2V2G)/[2 Cov(w, (z⫺ zm)2)] . (A7)
total selection coefficient is thus given bysT⫽sr/I2. One
example is where mutations have Gaussian effects on The fourth moment is m4⫽ 兰∞0 兰∞⫺∞h(a,s)
兵
H(s˜) ⫺the trait and gamma (1⁄
2) effects on fitness (i.e., a squared
3K(s˜)} (a4/16)dadsand the covariance of relative fitness
Gaussian random variable). Making the transformation and squared deviation is Cov(w, (z⫺zm)2)⫽Covp⫹Covr,
(,s) to (,v), whereby the ratiov⫽(s/sp)/(2/sr) is
in which Covp⫽ 兰∞0 兰∞⫺∞h(a,s)C(s˜)(s/2)(a2/4)dads is
F-distributed with 1 d.f. in both the numerator and de-the contribution due to pleiotropic effects of mutations
nominator, integratingleads to the density function and Covr⫽VG 2/(2Vs,r) due to real stabilizing selection.
of v,φ(v)⫽1/[(1 ⫹v)
√
v] (Moran 1968, p. 332). The expressions for the heterozygosity, H(s˜), and forNoting that 2/(2 ⫹ s) ⫽ 1/[1⫹ (s
p/sr)v], we have
K(s˜) andC(s˜) are given byZhanget al.(2002) by
replac-E[2/(2 ⫹ s)] ⫽ 兰∞
0φ(v)[1 ⫹ (sp/sr)v]⫺1dv⫽
冮
∞
0{2/[
ingsbys˜. In contrast withKeightleyandHill(1990),
who assumed that the strength of stabilizing selection (1⫹t2)(1⫹(s
p/sr)t2)]}dt⫽1/(1⫹
√
sp/sr) .is measured in phenotypic standard deviation units and Thus the expectation is determined only by the ratio ofsptosr.