JOURNAL VIROLOGY, July 1981,
0022-538X/81/070150-12$02.00/0
Fine-Structure
Mapping
of
Herpes
Simplex
Virus
Type 1
Temperature-Sensitive
Mutations
Within the Short
Repeat
Region of the Genome
V. G. PRESTON
Medical ResearchCouncilVirology Unit, University ofGlasgow,GlasgowGll5JR, Scotland
Received 12December 1980/Accepted31 March 1981
Cloned herpes simplex virustype 1(HSV-1) DNA fragmentswereusedto fine-structure mapthetemperature-sensitive (ts) lesions from fourmutants, tsT,D, c75,andK, by markerrescue.Thesemutantsalloverproduced immediate-early viral polypeptides at the nonpermissive temperature.
Although
one of these viruses, tsK,gave a morerestrictedinfected-cellpolypeptideprofile
under these conditions than the other three, no complementation was detected between pairwisecrossesofthese mutants intheyieldtest.Recombination, however,was obtained between all mutant pairs except ts T and D. Inphysical mapping
experiments, ts+viruswasrecovered fromcells coinfected with DNA oftsT,D, or c75 andBamHI fragmentk fromwild-type strain 17 HSV-1 DNA cloned in pAT153, whereas ts K wasrescued by cloned HSV-1 BamHI-y. Both of these clonedDNAfragmentscontained sequencesfrom the shortrepeat
region
of the HSV-1genome. The tsmutationswere moreprecisely mapped by markerrescue, using restriction enzyme fragments within BamHI-k and-y from cloned DNA. Thesmallestfragment ableto rescue amutant was 320basepairslong.Theorder of thefour mutations derived fromthesestudieswasconsistent with theassign-ment by genetic recombination. All four lesions mapped within the coding sequences of the
immediate-early polypeptide
Vmw IE 175 (ICP4) which lie outside the "a"sequence.Theresults showedthatmutations in different regions of thegeneencodingVmw IE 175could produce similarphenotypic effectsatthe nonpermissivetemperature.Infection ofsusceptiblecells with herpes sim-plex virus type 1 (HSV-1) in the continuous presenceof theprotein synthesis inhibitor cyclo-heximide results in restricted transcription of the viral genome (5, 9). The
virus-specific
mRNA's transcribed underthese conditionsby thecellularRNApolymeraseII codefor imme-diate-early (IE) polypeptideswhich are synthe-sizedvery earlyin the normal growthcycle of the virus (8, 18). Cells infected with tempera-ture-sensitive(ts) mutant tsT,D, K, or c75 and maintained at the nonpermissive temperature (NPT) produce
abnormally
large amounts of these polypeptides and reduced amounts of othervirus-specific species (11, 14, 17). Of these four mutants, ts K has been themost extensively studied. This virus has a ts defect in an IE polypeptiderequired for the"switch on" of tran-scriptionof HSV earlygenes,including the virus-coded thymidine (pyrimidine deoxynucleoside) kinasegene, and the subsequent "switch off" of transcriptionof viral IE genes (16, 28, 29). The IE polypeptide may also have a role in the activation of late viral genes, since itsfunctionis required continuously throughout the viral growthcycle, both beforeandafter theonsetof viral DNA synthesis (29). At the NPT, ts K inducesa Vmw IE 175polypeptide with abnor-malproperties (16). On cellfractionation, large amounts of this
polypeptide
were recovered in thecytoplasmic fractionin contrast totheVmw IE 175induced incells infected withtsKatthepermissive
temperature (PT) orwithwild-typevirusateither the PT orNPT, which wasfound predominantly inthenuclear fraction. Further-more, Vmw IE 175 induced by ts K at the NPT failed to be processed normally to species of lower electrophoretic mobility. Both of these aberrantpropertieswerereversedby transfer of tsK-infected cells from the NPT tothe PT in the absence of further protein synthesis, sug-gestingthat the ts Klesionwas in the Vmw IE 175polypeptideand not in aprocessing enzyme (16).Similardefects in Vmw IE 175 induced by tsTorD at the NPT have also beenobserved (D. MacDonald and C. M. Preston, personal communication).
Marker rescue of ts mutations isa
commonly
150
on November 10, 2019 by guest
http://jvi.asm.org/
151
used technique for locating ts lesions on the physical map of the viral genome (22). In this method, isolated restriction endonuclease frag-ments from wild-type HSV DNA are screened for their ability to rescue selected genetic markers. A number of problems associated with this approach have limited its use. In particular, difficulty in purifying specific DNA fragments, free of contaminating sequences from other re-gions of the viral genome, canlead to spurious rescue of selected markers. Furthermore, the efficiency of rescue may be low, especially when small DNA fragmentsareused to rescue virus, and therefore it can be difficult to map muta-tions which lie close together. In addition, ob-taining large amounts ofsmall fragments may be a problem. All of these difficulties can be overcomebyusing cloned HSV DNAfragments in physical mapping experiments. The greater precision in mapping genetic markers should facilitate theassignment of mutations to specific genes.
In this study, we have used cloned HSV-1 DNA tofine-structuremaptslesions from four mutants, tsT, D,c75,and K.Previousworkhas assignedthemutations in tsT, D,and K tothe shortrepeat sequences ofHSV-1 DNA (22, 23) and the defect in ts c75 totheterminally redun-dant "a" sequences present in both the short and long repeat regions ofthe genome (10). It wastherefore of interesttoknowwhether these mutations were in the genecodingfor Vmw IE 175, known to mapentirelywithin theinternal (IRs) and terminal (TRs) short repeat (30) or within some other gene, forexample, the gene coding forVmw IE 12or68,whose 5'endmaps, in each case, within
IRs/TRs,
and whose 3' terminus maps with the short unique (Us)re-gion.
MATERIALS AND METHODS
Tissue culture cells. BHK-21 clone 13 cells (12)
weregrown inEaglemediumsupplementedwith 10%
tryptosephosphatebroth and 10% calfserum.
Virus. The isolation of HSV-1 strain17andtsT,
D, and K has been described previously (2; I. K.
Crombie, Ph.D. thesis, UniversityofGlasgow, Glas-gow, Scotland, 1975). Nonsyncytial isolates of these
viruseswereusedinallexperiments.tsc75wasderived
from strain 13vB4(7),andbothwerekindly provided
by R. W. Honess. These ts mutants revert to ts+
phenotype at amoderate frequency, suggesting that
thelesionsarepointmutations.
Recombination and complementation tests.
Recombination andcomplementationtestswere
per-fonnedessentiallyasdescribedbyBrownetal.(2).In
therecombinationandcomplementationyieldassays,
cell monolayerswereused insteadofcellsin
suspen-sion.
Preparation ofviral DNA. DNA was obtained
from cell-associated and cell-released virions as
de-scribed by Wilkie (31).
Construction of recombinant plasmid DNA
molecules. BamHI fragments k and y fromwild-type
HSV-1 strain 17 DNA were cloned into the plasmid pAT153 (26) by direct ligation of BamHI cohesive ends (25).
Isolation of plasmid DNA. Cultures of Esche-richia coli strain K-12 HB 101, harboring pAT153 carrying HSV-1 BamHI-y (pGX1) or BamHI-k (pGX2), were grown in Luria broth containing
ampi-cillin (100 tg/ml). Plasmid DNA was amplified by
treating logarithmically growing cells with 25 ytg of
chloramphenicol per ml for 12 h. After harvesting, the cells were suspended in a 25% solution of sucrose in 50
mM Tris-hydrochloride (pH 7.9) and incubated at0°C
inthe presence of lysozyme (1.7mg/ml). After 30min,
EDTAwasadded to a final concentration of 0.05 M
andincubation was continued for a further 30 min at
0°C. NaCl and sodium dodecyl sulfate (SDS) were added to final concentrations of 1.0 M and 1% (wt/
vol), respectively. The lysate was then stored at0°C
overnight before centrifugation for 1 h at 20,000 rpm inaSorvallSS34 rotor. Thesupernatantwasdiluted
with an equal volume of water and extracted twice
with phenol and once withchloroform. The DNA was
concentrated by ethanol precipitation, suspended in
0.01MTris-hydrochloride-0.001 M EDTA (pH 7.9) in
the presence of RNase A (100 ug/ml), and dialyzed
extensively against the same buffer. The DNA solution was extracted twice with phenol and once with
chlo-roform beforebeing precipitatedby isopropanol
treat-ment (13). The pellet was resuspended in a small
volume of0.01 MTris-hydrochloride-0.001 M EDTA
(pH 7.5).
Enzymes. RNase A was obtained from Sigma
Chemical Co. Restriction endonucleases were
pur-chased from New England Biolabs, and the enzyme
reactions werecarried outunder the conditions
rec-ommendedbythesuppliers. ThedigestedDNA used
inmarkerrescueexperimentswasextractedoncewith
phenol and once withchloroform to remove the
re-striction endonuclease.
Purification ofrestrictionendonuclease DNA
fragments. Restriction endonucleasefragmentswere
separated byagarose gel electrophoresis (34) inthe
presenceof ethidium bromide(0.5
,g/ml).
DNAbandswerevisualized underlong-waveUVlightand excised
from thegels.The DNAwaseluted from thegelslice
by high-voltage electrophoresis and purified further byhydroxylapatitechromatographyasdescribed
pre-viously (32). The DNA solution was dialyzed
exten-sively against 0.01 M Tris-hydrochloride-0.001 M EDTA (pH 7.5)and concentratedbyethanol
precipi-tation in the presence of carrier E. coli rRNA(30ytg/
ml). Thepelletwassuspendedin0.01M
Tris-hydro-chloride(pH 7.5).Verysmall restrictionendonuclease
fragmentswereseparated bypolyacrylamide gel
elec-trophoresis (33).Thegelswerestainedwithethidium
bromide (10yg/ml) for 10min, andthe DNAbands
werevisualizedunderlong-waveUVlight.Thebands
were excised from the gel, and the gel slices were
crushedbyDouncehomogenizationin10ml of0.01M
Tris-hydrochloride-0.001M EDTA(pH 7.5).The
sus-pension ofpolyacrylamide was rocked gently
over-nightat4°Cbeforelow-speedcentrifugationtoremove
on November 10, 2019 by guest
http://jvi.asm.org/
152 PRESTON
thepolyacrylamide.The DNAwaspurifiedfurtherby
hydroxylapatite chromatographyasdescribed above.
Marker rescue assay. Marker rescue was
per-formed essentially as described by Stow etal. (22).
Briefly, BHK cellmonolayersin 50-mm dishes were
transfectedwithamixture of mutantviral DNA (0.5
to1
jig)
andaclonedwild-typeHSV-1DNAfragmentand treatedwith 25%(vol/vol) dimethylsulfoxide for
5minat4.25 hpostinfection.Afterincubation for 3or
4daysat31°C,the infectedcells wereharvested,and
the viral yieldwasassayedat31and38.5°C.
SDS-PAGE. Virus-infected cell protein samples
wereanalyzed by SDS-polyacrylamide gel
electropho-resus (PAGE) onagradient gel (6to 15%
polyacryl-amide) witha stacking gelof 5% polyacrylamideas
described by Marsdenetal.(14).
RESULTS
Comparison of polypeptides induced by
tsT, D,K and c75 in BHK cellsunder per-missive and nonperper-missive conditions.
Al-though previous work (6, 7, 11, 14) has shown
thattsT, D, K, andc75 exhibitsimilar,restricted
polypeptide phenotypes at the NPT, no one
study has compared the polypeptides induced
by thesefour mutants. For thisreason,the poly-peptide profilesof BHKcellsinfected withtsT, D, K, and c75 under nonpermissiveand permis-sive conditionswere examined bySDS-PAGE.
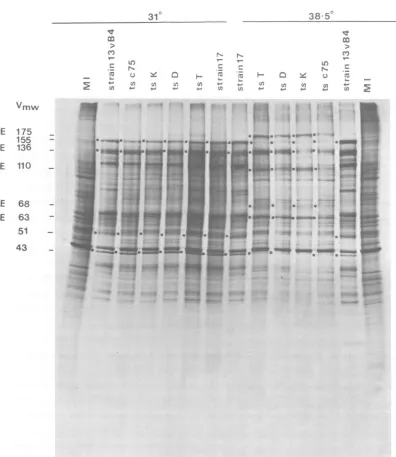
Figure1showsanautoradiogramof electropho-retically separated virus-infected cell polypep-tides labeled with[35S]methioninefrom 6 to 8 h
postinfection.Cells infected with mutantorts+
virusat310Cgavealatepatternofvirus-infected
cellpolypeptide synthesis, asdid cellsinfected
with ts+ virusattheNPT.By comparison, cells
infectedwith tsT, D, K,orc75 underrestrictive
conditionssynthesized increasedamountsof IE
polypeptides (e.g.,Vmw IE175, 110, 68,and63) and reducedamountsofearlyviralpolypeptides (e.g., the major viral capsid polypeptide Vmw
155) and failed tomake detectable amountsof
latepolypeptides (e.g., Vmw 51).The pattem of
virus-specific protein synthesisin tsK-infected
cells at the NPT was more restricted than in
cells infectedwithtsT, D,orc75.Forexample, the Vmw 43polypeptide bandwasnot
detecta-blein ts K-infected cellprofiles, whereastrace
amounts of this species could be seen inother
mutantpolypeptide profiles.
Genetic studies with tsT, D, K, and c75. (i) Complementation. InHSV,themost
com-monly used complementationassayistheyield
test, in which the total amount of infectious
virus produced by cells mixedly infected with
twodifferentmutants atthe NPTiscompared with the yields of virus produced from cells infected with each mutant alone at the NPT
after one growth cycle. Another assay is the
infectiouscenter test,in which the abilityofcells
mixedly infected with two ts mutants to form plaques on uninfected cell monolayers at the NPT is compared with that of cells
singly
in-fected with eachtsmutant.Althoughearly work,onthebasis of the infec-tious center test,
assigned
ts K to a different complementationgroupfrom thatoftsTandD (Crombie,Ph.D. thesis, 1975), subsequent stud-ies with these mutantsraised doubtsaboutthis findingsincenegative resultswereobtained from theyieldtest.For this reason,theability oftsT, D,K, and c75 tocomplement
oneanotherwas checked byusing bothcomplementationassays. Inthe yield test, no complementationwas ob-served betweenpairwise
crosses of these mu-tants(Table
1). This result confirms previous observations from our laboratory (N. D. Stow and D. Dargan,personal
communication) and is consistent withreportsfrom otherlaboratories using mutants of the same phenotype (6, 11). Positivevalues, however,wereobtained fromall but two crosses in the infectious center test. There are twomajor problems associated with this test. First, it involves more manipulations underpermissive conditions than the yieldtest; second,aconsiderableamountof recombination may occur atthesametimeascomplementation
(15). To test the
possibility
that leakiness of virusmayhaveresulted in false-positive values ofcomplementation, viruswasabsorbedtocells in suspension at two different temperatures in theinfectiouscentertest.Byincreasing
the virusabsorption
temperature, the number ofpositivevalues ofcomplementationwas reduced (Table 2).
(ii) Recombination. The fourmutants were tested for their ability to recombine with one another.Allmutantpairs,except ts T and ts D, produced significant levels ofts+ progeny virus in two-factor crosses. Recombination values ranged from <0.001% (for thecross ts T x ts D) to 17.1% (for the cross tsT x ts K) in a single experiment. By usingrecombination data from three independent experiments, a linkage map of mutationswasconstructed (Fig. 2). It should be notedthatthets c75mutation does not lie to oneend of the genetic map as would be expected if this lesion mapped in the "a" sequence of HSV-1 DNA asreported by Knipe et al. (10).
Fine-structure mappingoftsT, D, K, and c75 mutations. The mutations in ts D and K have been located within the short repeat se-quences (11, 21), whereas the ts c75 lesion has beenreported to lie within theterninally redun-dant "a" sequence ofthe HSV-1 genome (10). Cloned HSV-1BamHI restriction endonuclease fragments fromthis region were therefore tested for their ability to rescue ts+ virus from cells transfectedwith tsmutant DNA. Figure 3 shows
on November 10, 2019 by guest
http://jvi.asm.org/
310 38
50
.)_ N PN- V,
- c
~~~~~N
(UX Q Y a l_- X ~a- I a
4E () (4 (4 U, 4.0 4. (4 U) (4 4-0
U)0 4-0 +.' 40 40 U0 (A 4' 4.' 4' 4.' U,
Vmw
IE 175 155 IE 136
IE
110
IE
68IE 63 51
[image:4.493.47.443.63.520.2]43
FIG. 1. Autoradiogram ofelectrophoretically separated
polypeptides
frommock-infected
cells (MI) andvirus-infectedcells incubated atthePTand NPT. Cellswere labeledwith
['SJmethionine
from6 to 8hpostinfection.
the result ofamarker rescue experiment using clonedBamHI
fragments
k and y from HSV-1 strain 17. Themaplocation of thesefragments
on the HSV-1 genome is also included in the figure. When cells were coinfected with ts K DNA and
pGX1
plasmid
DNA(which
contains theHSV-1 BamHIfragment y),
ahigh
levelof ts+ progeny was obtained. Sincewild-type
se-quences from
BamHI-y
wereableto correctthedefect in the mutant, the ts Klesion must lie withinthisregion.By similar reasoning, ts T, D, andc75lesionswere
mapped
within the BamHI-k sequenceofHSV-1 DNA.Onthe basis of this experiment,the ts Klesionwasseparatedfrom the three other defects. Theseresultswere ob-tained byusing
plasmid DNA digested with BamHI.Ifuncleavedplasmid DNA(about75% supercoiled) wasused, the efficiency ofmarkeron November 10, 2019 by guest
http://jvi.asm.org/
154 PRESTON
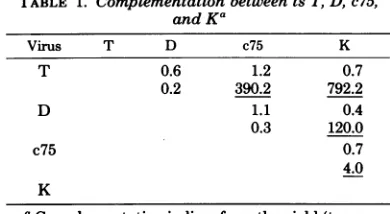
TABLE 1. Complementation betweentsT, D,c75,
andKG
Virus T D c75 K
T 0.6 1.2 0.7
0.2 390.2 792.2
D 1.1 0.4
0.3 120.0
c75 0.7
4.0
K
aComplementation indices from theyield (top
num-ber) and the infectiouscenter(bottomnumber)tests.
Values were calculated from the formulas given in
Brown et al.(2). Indices greater than2.0 were
consid-eredpositive andareunderlined. Anabsorption
tem-peratureof38.5°Cwasused in theyieldtest, whereas
a temperature of 37°C was used in the infectious
[image:5.493.255.448.54.212.2]centertest.
TABLE 2. Complementation indices from infectious
centertestsa
Virus T D c75 K
T 1.3 183.2 408.5
<0.1 38.7 1.8
D 0.7 285.5
0.9 37.9
c75 3.4
1.9
K
aA temperature of 37°C (top number) or
390C
(bottom number) was used.
rescue of ts+ progeny virus was markedly re-duced (Table 3). If, however, cells were coin-fectedwithplasmidDNAcleavedwithHindIII andwholetsmutantDNA, the proportionof ts+ progeny wasonlytwo- tofivefold less than val-ues obtained by using
plasmid
DNA digested withBamHI. These resultssuggest that linear-ized plasmid DNA molecules(generated
by
cleavage with BamHI or
HindIII)
rescue ts+ virus moreefficiently thancircular, supercoiled DNA.Effect of varying the concentration of plasmidDNA on markerrescueofts+virus. Tomaximize the efficiency ofmarkerrescue,a
dose-response curve was obtained by varying the concentration ofplasmid DNA in transfec-tion experiments. Increasing the concentration ofplasmidDNAmixed with aconstant amount ofmutant viral DNA increased the proportion ofts+ progeny (Fig. 4). A plateau level of ts+ progeny was reached when the relative molar ratio ofBamHI-y or -k to viral genomic DNA wasfiveorgreater.
Therefore,
inallsubsequent experiments a 10-foldor greater relative molar ratio of HSV-1BamHI-yor-k(orsubfragmentsTD c75
T ~D --c75 K
-C0001. -C0001, 0
00%
3.86
2-0171 5, 11-2
0.9. 0.7. 1.0 3-1. 10.. 2'4
[image:5.493.51.246.74.181.2]1-6, 0o 1-4
FIG. 2. Linkage mapoftsT, D, c75, and K. Values
represent recombination frequencies (percentage)
from three independentexperiments andwere
calcu-latedbyusingtheformulafromBrownetal.(2). The
toplineofthe map isascalerepresentationof
recom-binationfrequencies (from oneexperiment) between
adjacent markers. The bottom lineofthe map
repre-sents the order of mutations without regardto the
relativephysicalmap distance.
of these sequences) to HSV-1 ts mutant DNA wasused.
Fine-structure
physical
mapping
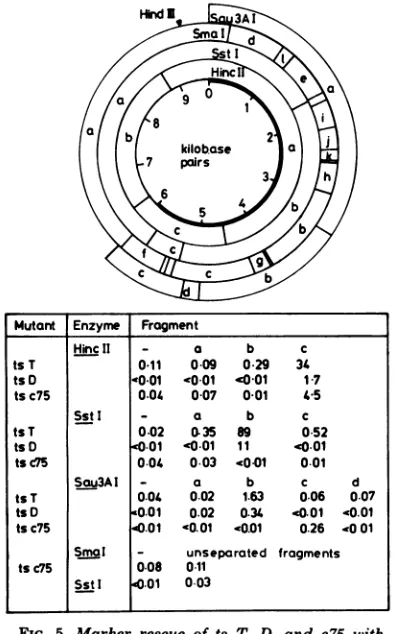
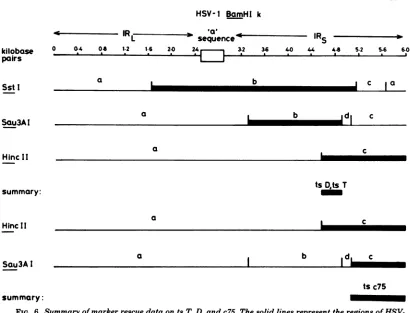
oftsT, D, c75, and K within BamHI-k and-y. Thets mutations were more precisely mapped by
marker rescue with restriction endonuclease fragments within cloned BamHI-k and -y. The results of transfectionexperiments using tsT,D, or c75DNAmixed withspecificsubfragments of plasmid pGX2 DNA (which containsthe HSV-1 BamHI-k fragment) are shown in Fig. 5 and summarized in Fig.6. The
fragments
HincII-c, SStI-b,and Sau3AI-b allrescued thetsT and D lesions. Since the smallest region of BamHI-k common toall threefragmentswasthesequence sharedby Sau3AI-b andHincII-c,
thetsT and D mutations must lie within this DNA. Al-though ts c75 wasrescued by HincII-c, the re-sults ofmarker rescue experiments with sepa-ratedSau3AIfragments from BamHI-ksuggest that this lesion lies in a different region of BamHI-k from ts T and D defects, within the terminal Sau3AI fragment c. No significant levels ofts+ progeny virus were obtained with separated orunseparatedSstIfragmentsin coin-fectionexperiments withts c75 DNA. The ob-servation suggests that the ts c75 lesionmaps closeto an SstIcleavagesite.UnseparatedSmaI fragments from Bam-HI-k also failed to rescue this mutant. Since there is anSmaIsiteadjacent totheSstI sitedelimiting SstI-band c (A. Dav-ison, personal communication), thisresult indi-cates that the ts c75 defect is located close to thisSstIsite.J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.52.248.271.371.2]MAPPING OF HSV-1 ts MUTATIONS 155
[I.
L
IRL
IRS4Us*TRS
~4- S
BamH I
bi
k Y n i I'l x qI R-O 4-IR US -. 4- TR -.
L S S
Viral DNA Cloned HSV-1 fragment Relative eop (38.5° titre/31°titre ) x
103
tsK <0.01
BamHI y 10.3
[| I k <0.01
ts T
_
0.09BamHI y 0.01
BamHl k 203
ts D <0.01
BcmHI y c0.01
BamHIk 31.6
ts c75 <0.01
BamHI y <0.01
BamHI k 3.5
FIG. 3. MarkerrescueoftsT,D, c75, and K. The HSV-1 genome contains two unique regions, UL andUs,
eachflankedbyinverted repeat sequences(TRL, IRL;TRS, IRS)andjoinedatIRL-IRs (20). The S component
of the viral DNA is shown in detail together with theBamHIphysicalmapof this region. Plasmids pGX2 and
pGXI, containingHSV-1 BamHIfragments k and y, respectively, werecleaved with BamHIbefore being
testedforabilityto rescue tsT,D,c75, K.
TABLE 3. Efficiency ofmarkerrescueoftsTandK,
using cloned HSV-1 in linearorsupercoiledform
DNA Plasmid DNA Relative
tsK None <0.001
pGX1 (containsHSV-1 BamHI- 0.04 Y)
pGX1digestedwith BamHI 0.9
pGX1
digestedwithHindIII 0.2tsT None 0.009
pGX2(containsHSV-1 BamHI- 0.8
k)
pGX2digestedwith BamHI 10.5
pGX2digestedwithHindIII 4.3
CValues
represent relative efficiency of plating(EOP), (38.5°C/31°C) x 100.
By using separated restriction endonuclease fragmentsofBamHI-y,the tsKlesionwasalso mapped to a smallregion of the viral genome.
The results ofmarker rescue experiments are
shown in Fig. 7andsummarizedin Fig.8. The
dataareconsistentwith thetsKdefectmapping
in BamHI-y close to the BamHI site which
delimits BamHI-m' and-y.
Figure 9 shows an overall summary of the
fine-map locations oftsT, D, c75, andK
muta-tions.AlthoughthemRNA for Vmw IE 175has
been shown to lieentirelywithinIRs/TRs (30),
the exactmappositionofthe mRNA is not yet
known. The 3' end of themRNA, however,has
been shownbynucleotide sequence analysisof
BamHI-k and nuclease S1 mapping (1) to lie
within BamHI-k, 3.2 kilobase pairs from the
restriction endonuclease site delimitingBamHI
fragments k and m' (A. Davison and F.Rixon, personal communication). Since the size of the
mRNA specifying Vmw IE 175 has been
esti-matedto be 4.7 kilobases (30) and there isno
evidence of splicing of the transcript for this
TRL
:3
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.54.437.63.399.2] [image:6.493.47.241.471.603.2]156
PRESTONWo
i
ac c
0136 076 025 O
ug plasmid DNA ug plasmid DNA
FIG. 4. Relationship betweenefficiency ofmarkerrescueofamutantand the wild-typeDNAfragment
concentration.
(V)
MarkerrescueoftsT withpGX2cleavedwithBamHI;(2)markerrescueoftsKwithpGX1cleaved with BamHI.
Mutant Enzyme Fragment
HincII - a b c
ts T 0.11 0-09 0.29 34
tsD0 0-01 -0 01 '0 01 17 tsc75 0.04 0-07 001 45
SstI - a b c
tsT 0.02 035 89 0 52
tsD .001 '0.01 11 0.01
tsc75 0.04 0.03 4001 0.01
Sau3AI - a b c d
ts T 0.04 0.02 1.63 0.06 0.07
ts D <0.01 0.02 0.34 40.01 40.01
tsc75 .01 '0.01 '0.01 0.26 .001
Smal - unseparated fragments tsc75 0.08 0 11
Sst I .01 003
FIG. 5. Marker rescue of ts T, D, and c75 with
restriction endonuclease fragments from pGX2 DNA.
Values represent relative efficiency of plating, 38.5/
31°Cx 103 of progeny virus from transfected cells.
Thephysical maps for pGX2 fragments generated by
SstI,HincII, and HindIII are shown above. Cleavage
polypeptidewithin theregion wherethe lesions map, tsT, D, K, andc75mutationsmustalllie within thecodingsequences of the gene.
Marker rescue ofts c75 with separated HpaI fragments from strain 13vB4. It has previously been reported by Knipe et al. (10) that the ts c75lesionis locatedwithin the "a" sequenceof HSV-1 since the terminaland joint-spanning DNAfragments from ts+ virus DNA rescuethismutant. Becausethese workers had used restriction endonuclease DNA fragments from strain MP andan intratypicrecombinant in markerrescueexperimentswith this virus, we attempted to confirm the original observations by using the parental ts+ virus strain 13vB4 DNAofts c75.Althoughtheresults in Figure 10 show some fragment contamination (for
exam-ple,
HpaI-i
iscontaminated withHpaI-g + h),the data suggest that sequencesfrom IRL/TRL areunabletoconvert tsvirustots+ phenotype. This findingsupports the conclusion that the ts c75 mutationmaps within
IRs/TRs
but not in the"a"sequence.DISCUSSION
Although tsmutationsin atleast29 cistrons of HSV-1 have been identified (3, 19; P. A. Schaffer,
unpublished
data), fewof thesedefects havebeenassignedtospecificgenes. Thispaper describes the fine-structure mapping of four ts mutationswhichliewithin the short repeatse-sites of Sau3AIare given for HSV-1 BamHI-k
se-quencesonly (24, 27; Davison, personal
communica-tion). The solid region of pGX2 refers to HSV-1 BamHI-k sequences, and the open region represents pAT153 DNA sequences.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.493.103.397.61.234.2] [image:7.493.54.256.279.596.2]MAPPING OF HSV-1 ts MUTATIONS 157
HSV-1
BamHI
kIR7
sequence-Rs
0 0.4 04 1.2 1.6 2.0 32 3.6 40 44 44 52 54 6.0
a I
~~~~~b
[ c aa b d c
a _
ts
D*ts
Ta
a b d c
tsc75
summary:
FIG. 6. Summary ofmarkerrescuedataontsT, D, andc75.Thesolid linesrepresenttheregions of
HSV-1BamHI-kwhichrescuetsT, D,orc75.
quences ofthe HSV-1 genome and show that
these lesions mapwithin the coding sequences
of thegenespecifying theimmediate-early
poly-peptide Vmw IE 175 (ICP4).
By using cloned HSV-1 DNA fragments in
marker rescue experiments, I have overcome
manyof theproblemsencounteredwhen using
DNA fragments isolated from viral DNA. The
improved efficiency of rescue of ts+ progeny
from cells coinfected with mutant viral DNA
and ts+ HSV-1 fragment DNA allowed
muta-tionstobe located withinverysmallregionsof the viralgenome.Inthecaseof tsK,rescuewas
obtainedwitha320-base-pairSau3AIfraginent
fromcloned HSV-1 strain 17BamHI-y.Todate,
this is the smallest DNAfragment reported to
rescueatslesion,andclearlymarkerrescuewith this sizeoffragmentisapproachingthe limits of
thetechnique.Theabilitytomapamutation to
averysmallregionof the viralgenomebyusing
clonedfragmentsfromwild-typeHSVDNAnow
makesitpossibletodeterminebyDNA
sequenc-ingmethods theprecisebasepairchangecausing
the defect.
AsimilaranalysisoftsmutationswithinIRs/
TRs
hadbeen made byDixon and Schaffer(6).Theymapped the lesions ofviruses
belonging
tocomplementation group 1-2tothree regions of the short repeat, probably corresponding to
BamHI-yandHSV-1 sequenceswithin
HincII-a and -c in pGX2. Although they assigned ts LB2 mutation to sequences within BamHI-y,
thephysicalmaplocation of thisdefect didnot
correlate withthepositiononthegeneticmap.
Suchanomalousbehaviorwasnotobserved with
the tsKmutation withinBamHI-y. Both tsK
andLB2havemorerestrictedpolypeptide
phe-notypesatthe NPT than othermutantswithin this complementation group, and therefore it would beinterestingto determine whether the
ts LB2 lesion maps in the same region of
BamHI-y as the tsK mutation. In contrast to
resultspresentedin thispaper,Dixon and
Schaf-fer(6)obtainedrescueoftsc75 with two
differ-entfragments from the shortrepeat.This
find-ingled them tosuggestthattsc75mightcontain
two mutations within each short repeat, one
within the"a"sequenceattheIRL/IRs junction
or end of TRs and one within a second "a"
sequencepresentinthe HSV-1 HincII-cregion ofpGX2.Iwasunabletoobtainrescueof tsc75
withthe HincII-afragment (Fig. 5) and
there-kilobase pairs
SstI
Sau3AI
HincII
summary:
Hinc1I
Sau3AI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.493.43.453.57.370.2]Enzyme fragment
Pvul1/BamHI - a b c
0.01 0 01 1536 0*001
Hinfl - a b c.d e f g h.i j.k
<00<004324-001 <0*01 cOOl <001 001 <001 010
SmaI/BamHI - a b. c d e f g
<001 0.02 0.57 <0.01 0.01 <001 003
Sau3AI - a b c d e f
i 006 0o09 172 <0*01 <0.01 <0.01 <0.01
FIG. 7. MarkerrescueoftsKwith restriction endonucleasefragmentsfrom pGX1DNA. Values represent
relativeefficiencyofplating, 38.5/31°Cx103, ofprogenyvirusfrom transfectedcells. Thephysicalmapsfor
pGX1 generated by PvuIIplusBamHI,Hinfl, HindIII, and BamHIplus SmaIare shownabove. Cleavage
sitesof Sau3AIaregivenforHSV-1 BamHI-ysequencesonly(24; M.-J.Murchie,personalcommunication).
Thesolidregion represents HSV-1BamHI-ysequences, andthe openregion referstopA T143DNAsequences.
Note thatpAT153is present in thisplasmidas adimer in head-to-tail arrangement.
HSV-1 BamH I y
0 0.2 0.4 06
[ . a a 0B
1.0 1.2 14 1 6 175
I . *
b
c]~~~Q-
cI
e_
I
f e bI
db
if
I
a
Idjle
[image:9.493.100.445.29.327.2]tsK summary:
FIG. 8. Summary ofmarkerrescuedataontsK. The solid linesrepresenttheregionsof HSV-1BamHI-y
whichrescuetsK.
158
k
ilobase
pairs
Pvu II
Hinf
I
Sma I
Sau3A
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.493.89.417.393.653.2]MAPPING OF HSV-1 ts MUTATIONS 159
1 3 I 4
I
-m a
III
Ie.n 11 v n
I
i5I
35
:
3rS
I
n
'I,
m'
1 7 11
Y'
I
v11
n,DT
K
K
T,D
'c75
c75
08 09
'~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.______os___ 0
fractional
genome length
FIG. 9. Physical map locations of ts T, D, c75, and K mutations. The orientation and map positions of IE
mRNA's in the S componentof HSV-1 DNA are based on data from Watson etal. (30), and Clements etal.
(4)andanunpublished data of Davison and Rixon. IE mRNA's 1, 3, 4, and 5 specify the polypeptides Vmw
110, 175, 68,and 12,respectively.
k u
m o l p ? n i t
HpaI
I I
i
,b h f e qsvrm c g
I
I
I
- a b cde f gh i j I m n k? u?opqr s t v
0.01 7.06 0.50 19.74 0.25 8.60 1.34 0.56
0.15
0.20 0.50 0.20 0D2 0.05 0.04 0.02FIG. 10. Markerrescueofts c75with separated HpaIfragmentsfrom 13vB4 DNA. The HpaIphysical map
ofstrain 13vB4 isalmost identical to that of strain 17 except for the apparent absence of the cleavage site
delimiting HpaI fragments k anduand the presence of a new site within this DNA region. To avoid confusion,
the nomenclatureforthe strain17HpaI digest has been used. The joint-spanning DNA fragments HpaI-a
and darecomposedoffragmentsm+candm +g,respectively. Values represent relative efficiency ofplating,
38.5/310Cx 103, ofprogenyvirusfromtransfected cells. The underlined figures refer to values obtained by
usingfragmentscontaining sequences from the repeated regions of HSV-1 DNA.
forecannotsupporttheirproposal. In addition, Ifailedtoshowconvincingrescueof ts c75 with sequencesfrom thelongrepeatof strain 13VB4 DNA and suggest that this mutant does not haveadefect in the "a"sequence.This conclu-sion is
supported
byDNA sequenceanalysis
of a cloned HSV-1 DNA fragment spanning theIRs/TRs
region of strain 17viralgenome.Thiswork showed that inthe cloned DNA fragment, the 3' endofthe gene for Vmw IE 175lieswithin the short repeat but at least 800 base pairs outside the "a" sequence (Davison and Rixon, personal communication). Hence, iftsc75had a mutation within the "a" sequence, it would not beexpectedtohaveaphenotypelike thegroup 1-2mutants.Themap location of thismutation can therefore nolongerbe used asevidence in
support of the circular
replication
model pro-posed byKnipeetal. (10)in whichthe terminal "appsequencesareregenerated by repair
synthe-sis, using the internal invertedrepeatsas a
tem-plate.
Itshouldbenoted thattsc75wasrescuedless
efficiently by
HSV-1 strain 17 BamHI-kse-quencesthan were tsT and D. Thereason for therelatively poorrescueoftsc75 isnotclear. It ispossiblethat ts c75containstwo tslesions that map close together in BamHI-k. Alterna-tively, viral strain differences may accountfor the inefficient rescue ofts c75. Another
expla-nation is that thereareDNAsequences between the ts T andc75mutations that enhance
recom-bination.
Precise physical mapping of mutations
en-IEmRNA
BamHI
mutation
.4
-IR
L
--*--I
Rs
-0Lis
it,-tTRs
90I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.493.47.444.51.272.2] [image:10.493.58.436.336.406.2]160 PRESTON
ables recombination
frequencies
to be inter-preted in terms ofphysical distance. Fromthe genetic linkage map of the HSV-1 thymidine kinase locus whichlies withinUL,
Smiley
etal. (21)calculatedaminimum value of 3.2% recom-bination perkilobasepair
of DNA. The maxi-mumdistance betweentsT and K is about 2.14 kilobasepairs.Taking
ageometricmean recom-bination frequencyof11.8% between thesetwo lesions(from threeindependentexperiments),
a value of 5.5% perkilobase pair ofDNA is ob-tained.Although
thisfigure
isconsiderably
higher thantheestimate of
Smiley
etal.(21)
for recombination in theunique
region
of HSV-1 DNA, there is enormous variation in the fre-quency of ts+ recombinantsfromagiven
genetic crossperformedatdifferenttimes,
whichmakes itdifficulttocompare results fromdifferent lab-oratories. Forexample,
the cell type used in geneticexperimentsappearstoinfluence recom-bination values. In thisstudy
and the oneby
Honessetal.(7),significant values ofaround 1% were obtained for thecross ts D x tsc75 per-formed inBHK cells. By contrast,
Knipe
etal. (10) and Dixon and Schaffer (6),using
rabbit skincells and humanembryoniclung
fibroblasts,
respectively, failedtodetect recombination be-tween these two ts mutants. In
calculating
re-combinationfrequencies,
a number of assump-tions aremade,
including
thefollowing:
(i) asingle crossover event is sufficient for the for-mationofvirus withats+phenotype (provided intermolecular recombination occurs between linear andnotcircular DNA
molecules);
(ii) ts+ virushad arelativeefficiency
ofplating (NPT/ PT)of 1; and(iii)thedoubletsmutantreciprocal class arises atthe same frequency as does the ts+class. Although the tsKandTlesionsmap withinadiploid gene,twosourcesofinformation suggest thatonly onefunctional copy is neces-sary forthe ts+phenotype. First, only oneend ofthe S component of HSV-1 strain1061rescuestsD
(11),
andsecond,
HSV-1/HSV-2
recombi-nants, containing one HSV-1 and one HSV-2 short repeat,expressboth VmwIE175 of HSV-1 and the HSV-2 counterpart polypeptide. Therefore, asinglecrossoverevent intheshort repeatsequence shouldbesufficientfor the gen-eration of a ts+ recombinant. On this assump-tion, Dixon andSchaffer (6) suggested thatthe recombination frequency between two muta-tions in adiploid gene shouldbetwicethe fre-quency expected for a haploid gene. Detailed analysisof thegenomestructuresofHSV inter-typicrecombinants withheterologous short re-peats has shownthatthesevirusesareunstable. Recombination between the heterologous re-peats cangenerateDNAmolecules with differ-ent arrangements of HSV-1 and HSV-2
se-quences fromtheparental recombinant genome
(18, 35; Davison, personal
communication).Itistherefore
likely
thatanintratypic recombinant,
heterozygous
for a ts lesion within the geneencoding
Vmw175,
would also be unstable andupon
subsequent
intermolecular recombinationsegregate to
give
virushomozygous
for ts or ts+ markers inadditiontoheterozygous
ts/ts+ virus. For this reason, the recombinationfrequency
between markers in a
diploid
gene would bepredicted
tobelessthan twice thefrequency ofthe samemarkers ina
haploid
gene. Inspiteof the difficulties ininterpreting thegeneticdata,
it is clear that there is a high frequency of recombinationwithin the shortrepeatsof
HSV-1,
but it is still uncertain whether thisregion
undergoes
enhancedrecombinationincompari-sonwith the
unique regions
ofHSV-1 DNA.ACKNOWLEDGMENTS
Iamgrateful to J. H.Subak-Sharpe for his encouragement andvaluablecriticism of the manuscript. I also thank A. J. Davison and M.-J. Murchie forpermission to cite the
unpub-lishedfine-structure restriction endonuclease cleavage maps of HSV-1 strain 17BamHI k andy fragments and M.Buckley
for excellenttechnical assistance. Inaddition, Iamgratefulto B.Honess forproviding stocks of the HSV-1 strain 13vB4 and tsc75 andtoP.A.Schaffer for a copy of themanuscriptby
Dixon and Schaffer beforepublication. LITERATURE CIMD
1. Berk,A.J.,and P. A.Sharp. 1977. Sizing andmapping
ofearlyadenovirus mRNAsby gel electrophoresis ofSi endonuclease-digested hybrids.Cell 12:721-732. 2. Brown,S.M.,D. A.Ritchie, and J. H.Subak-Sharpe.
1973.Genetic studies withherpes simplex virus type1. The isolation oftemperature-sensitive mutants, their arrangement intocomplementation groups and recom-binationanalysisleading toalinkage map. J. Gen. Virol. 13:329-346.
3. Chu, C.-T.,D. S.Parris, R. A. F. Dixon, F. E.Farber, and P. A.Schaffer. 1979.Hydroxylamine mutagenesis ofHSV DNA and DNA fragments: introduction of mutations intoselected regions of the viral genome.
Virology98:168-181.
4. Clements, J. B., J.McLauchlan,and D. J.McGeoch. 1979.Orientation of herpes simplex virus type 1 imme-diateearly mRNA's. Nucleic Acids Res. 7:77-91. 5. Clements, J. B., R. J.Watson, and N. M. Wilkie. 1977.
Temporal regulation of herpes simplex virus type 1 transcription: location of transcripts on the viral ge-nome.Cell 12:275-285.
6. Dixon, R. A. F., and R. A. Schaffer. 1980. Fine-structure mapping and functional analysis of temperature-sensi-tive mutants in the geneencoding the herpes simplex virus type 1immediate early protein VP 175. J. Virol. 36:189-203.
7. Honess,R.W.,A. Buchan, I. W.Halliburton, and D. H. Watson. 1980.Recombination andlinkage between structural andregulatory genes of herpes simplex virus type 1: study of thefunctional organization of the ge-nome. J. Virol.34:716-742.
8. Honess, R. W., and B. Roizman. 1974. Regulation of herpesvirusmacromolecular synthesis. I. Cascade reg-ulation of thesynthesis of three groups of viral proteins. J.Virol. 14:8-19.
9. Jones, P. C., G. S.Hayward, and B. Roizman. 1977. Anatomy of herpes simplex virus DNA VIII. ocRNA is J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 39,1981
homologous to noncontiguous sites in both the L and S components of viral DNA. J. Virol. 21:268-276. 10. Knipe, D. M., W. T.Ruyechan,R.W.Honess, and B.
Roizman. 1979.Molecular genetics of herpes simplex virus: the terminal asequences of the L and S compo-nents areobligatorily identical and constitute a part of astructuralgene mappingpredominantly in the S com-ponent. Proc. Natl. Acad. Sci.U.S.A. 76:4534-4538. 11. Knipe,D. M., W.T.Ruyechan, B.Roizman,andL.W.
Halliburton. 1978. Moleculargeneticsofherpes sim-plex virus:demonstration of regions ofobligatoryand
non-obligatoryidentity within diploid regions of the genome bysequencereplacement and insertion. Proc. Natl. Acad. Sci. U.S.A.76:3896-3900.
12. MacPherson,L,and M. Stoker. 1962. Polyoma trans-formation of hamster cellclones-an investigation of genetic factorsaffecting cellcompetence. Virology 16: 147-151.
13. Marmur, J. 1961. Aprocedure for the isolation of
deoxy-ribonucleicacid frommicro-organisms.J.Mol. Biol. 3: 208-218.
14. Marsden, H. S., L. K. Crombie, and J. H. Subak-Sharpe. 1976.Controlof proteinsynthesis in herpes-virus-infectedcelis:analysis of the polypeptides induced by wild type and sixteen temperature-sensitive mutants of HSV strain 17. J.Gen. Virol.31:347-372.
15.Messer, L I.,andM.C.Timbury.1979.
Complementa-tion with tsmutants of herpes simplex virus. J. Gen. Virol.45:431-439.
16. Preston,C.M.1979.Abnormalproperties of an immedi-ateearlypolypeptideincells infectedwiththeherpes
simplexvirus type1mutanttsK. J.Virol. 32:357-369. 17. Preston,C. M. 1979. Control of herpes simplex virus type 1mRNAsynthesisincellsinfected withwild-typevirus orthetemperature-sensitivemutantts K. J. Virol.29:
275-284.
18. Preston,V.G.,A. J.Davison,H.S.Marsden,M.C.
Timbury,J.H. Subak-Sharpe,and N. M.Wilkie.
1978.Recombinantsbetween herpessimplexvirustypes 1and 2:analysesofgenome structuresand expression
ofimmediateearlypolypeptides.J.Virol.28:499-517.
19. Schaffer,P.A.,V.C.Carter,and M. C.Timbury.1978.
Collaborativecomplementation studyof temperature-sensitive mutants ofherpes simplex virustypes1and2. J.Virol. 27:490-504.
20. Sheldrick,P., andN.Berthelot.1974.Inverted
repeti-tions inthe chromosome ofherpes simplexvirus. Cold
SpringHarborSymp.Quant.Biol.39:667-678.
21. Smiley,J.R., KL J.Wagner,W. P.Summers,and W.
C. Summers.1980.Genetic andphysicalevidence for
thepolarityoftranscription of thethymidinekinase
gene ofherpessimplexvirua.Virology102:83-93. 22. Stow,N.D.,J. ILSubak-Sharpe,and N. M.Wilkie.
1978.Physical mapping of herpes simplex virus type 1
mutationsby marker rescue. J. Virol. 28:182-192. 23. Stow, N. D.,andN. M.Wilkie.1978.Physical mapping
oftemperature-sensitive mutations of herpes simplex virustype 1 byintertypicmarkerrescue.Virology90: 1-11.
24. Sutcliffe, J. G. 1978. pBR322 restriction map derived
from the DNAsequence: accurate DNA sizemarkers up to 4361nucleotide pairs long. Nucleic Acids Res.5: 2721-2728.
25.Tanaka,T.,and B.Weisblum.1975.Construction ofa colicinE1-Rfactor compositeplasmidin vitro: means
foramplificationofdeoxyribonucleicacid. J. Bacteriol.
121:354-562.
26.Twigg,A.J.,and D. Sherrat.1980.
Trans-complement-ablecopy-numbermutantsofplasmid ColEl.Nature
(London)283:216-218.
27.Wagner,M.J.,and W.C.Summers.1978.Structure of thejoint region and the termini of the DNA ofherpes
simplexvirus type1.J.Virol.27:374-387.
28.Watson,R.J., andJ. B.Clements. 1978. Characteri-zation oftranscription-deficienttemperature-sensitive
mutantsof herpessimplexvirus type 1.Virology91: 364-479.
29. Watson, R.J., and J. B. Clements. 1980. Aherpes
simplex virus type1functioncontinuouslyrequiredfor early and late virus RNAsynthesis.Nature(London) 285:329-330.
30. Watson, R.J., C.M. Preston, and J. B. Clements. 1979.Separation andcharacterizationofherpessimplex
virustype 1immediate early mRNA's. J. Virol. 31:42-52.
31.Wilkie, N. M. 1973. Thesynthesisand substructure of herpesvirus DNA: thedistributionof alkali-labile
sin-gle-strandinterruptions in HSV-1 DNA. J. Gen. Virol. 21:453-467.
32.Wilkie, N. M. 1976. Physicalmaps for herpessimplex
virustype1DNA forrestriction endonucleasesHindIII, HpaI,and XbaI. J.Virol.20:222-233.
33. Wilkie,N.K,J. B.Clements,W.Boll,N.Mantei,D.
Lonsdale,and C.Weissmann.1979.Hybridplasmids
containinganactivethymidinekinase gene ofherpes simplexvirus1.Nucleic Acids Res.7:859-877.
34. WilIe,N.M., andR.Cortini.1976.Sequence
arrange-ment inherpessimplexvirustype1DNA:identification
ofterminalfragmentsinrestriction endonuclease di-gests and evidence for inversions in redundant and unique sequences. J. Virol. 20:211-221.
35. Wilkie,N.M.,A.Davison,P.Chartrand,N. D.Stow,
V.G. Preston,andM.C.Timbury.1978.
Recombi-nation inherpessimplexvirus:mappingof mutations
and analysisof intertypic recombinants. ColdSpring HarborSymp. Quant.Biol.43:827-840.