Vol.49, No. 1 JOURNALOFVIROLOGY,Jan.1984, p.9-13
0022-538X/84/010009-05$02.00/0
Copyright © 1984, AmericanSocietyforMicrobiology
Binding of Simian Virus 40 A
Protein
to
DNA
with Deletions
at
the
Origin
of
Replication
BETSY A. LEWTON, ANGELO L. DELUCIA, AND PETER TEGTMEYER* DepartmentofMicrobiology, State University ofNew York, Stony Brook, New York 11794
Received 18May 1983/Accepted 23September 1983
Previousstudies with wild-type simian virus 40 DNA have shown that thesequence5'-GAGGC-3'directs
the binding of A protein (T antigen). The functional origin of replication contains four recognition pentanucleotides each of which is separated by a single base pair and arranged as two pairs of direct repetitions that areinverted relative to each other. Analysis of A protein bindingto a series of nonviable
mutanftsprogressively deleting these contactsites leadstothefollowing conclusions: (i) stablebinding of subunits of Aproteintothreeoriginpentanucleotides isnot sufficient for the initiation of DNAreplication,
(ii) the stability of DNA binding dependsoninteractions between boundprotein subunits, and (iii)asingle pentanucleotide is sufficient tobind and orienta subunit ofAprotein.
The origin ofreplication in simian virus 40(SV40) DNA has beenmapped to a 60- to65-base-pair(bp) regioncentered at the unique BglI restriction site (1, 3). Binding ofthe A
protein (T antigen) to the origin regulates the initiation of each round ofDNAreplication (10, 12, 14, 16, 18). Accord-ing to Hay and DePamphilis, RNA
oligonucleotides
atvariouspositionson theearly strand within theorigin prime
thesynthesis ofthe firstforwardarmofDNAreplication (7). Studieswithwild-type SV40DNA show thataconsensus
family of recognition sequences directs the binding of A
protein (2, 17). Different arrangements ofthe
pentanucleo-tide determine different patterns ofprotein binding in three
adjacent regions at theorigin of replication. Inregion I on the early side of the origin, three contact sites arranged as
direct repetitions lead to high-affinity binding but are not essentialfor DNAreplication(2, 3, 10,17).RegionIII on the late side of the origin contains six recognition sequences
arrangedasdirectrepetitions. Bindingtothese sites in vitro
is stable onlyat low saltconcentrations (2, 15).
Region II corresponds well to the functional origin of replication defined by the mapping of viable deletion mu-tants(3). Itcontains fouridealrecognitionpentanucleotides
that are eachseparatedbyabpandarrangedas twopairsof
direct repetitions inverted relative to each other. Base
substitutions between pentanucleotides are associated with wild-type or even increased levels of DNA synthesis. In contrast,
substitutions
within pentanucleotides reduce, but do not eliminate, replication (11). Deletions ofa part of apentanucleotideappear to
eliminate
replication altogether (6, 10).Inthe presentstudyweanalyze thebinding ofAprotein to
nonviable deletion mutants of region II togain insight into theminimalbinding requirementsfor DNA replication. Our
findingsalsoconfirmthe major features of a model for DNA
binding established with wild-typeDNA (15). MATERIALS ANDMETHODS
Proteinpurification. The SV40 A protein was purified from
productively infected CV-1 cells as previously described
(13).
DNA.Wild-typeas wellas mutant DNAs 6-1(dl6) and 6-17
(d19),
cloned inplasmid pMK16, were a gift from Y.Gluz-man(5). The deletion mutant 295
(d118),
cloned in pBR322,*Correspondingauthor.
was given to us by R. Martin.
The
plasmid was grown inEscherichia coli HB101, and the DNA was prepared as
describedby Kahnetal. (8). Forsirnplicitywerefertothese mutants as dl6,
d19,
andd118,
thenumbers of which indicatethenumber ofbp deleted from theoriginofreplication.
DMSfootprinting. SV40 DNA wasdigestedwith NcoI and 3' end labeled with
[32P]deoxyribosylthymine
triphosphate byuseof avianmyeloblastosisvirus reversetranscriptaseas previously described(18). Thetranscriptasewasinactivated byheatingat70°C
for 10min, and the labeledDNA wasthencutwith
Hinfl.
Theorigin-containing
fragment, radiolabeledon the 3' late end ofthe early strand, was purified by gel electrophoresis and was ethanolprecipitated. AfterSV40A proteinwas bound to the DNA for 1 h on ice, the guanine residues were methylated by the addition of 1 [L of 10 M dimethyl sulfate
(DMS)
for 10 min. The DMS reaction wasstopped,and theDNAwasrapidly processed bythe Maxam andGilbertDNA sequencingmethodspecificforcleavage at
guanine residues (9).Theresulting fragmentswere runon an
8.3Murea,8%polyacrylamide gel electrophoresissystem at aconstanttemperature of
50°C.
Footprinting
of the late strand was accomplished byradiolabelingthe3' early endof the late strandofthe
origin-containing restriction fragment Hinfl-BstNI as previously
described(2).
DNase footprinting. The BstNI-Hinfl
origin-containing
fragment ofeach DNA was radiolabeled on the 3' end of the
earlyor the late strandbypreviously describedmethods (2). SV40 A protein was bound to the
fragment,
and then 0.0005 units of DNase was added. The reaction was stopped, and the samplewas processed by the method ofDeLucia et al.(2).
Protected
DNA was analyzed by polyacrylamide gelelectrophoresisas describedabove. RESULTS
DMS footprinting. Nucleotides in close proximity to bound protein were
identified
with the DMS protection assay. For convenience we will refer to these as contact sites. In theseexperiments,anorigin-containingSV40DNA fragment was radiolabeled on the 3' end of either the early orthelatestrand. Theguanine residues were then treated with DMS in the presence or absence of A protein at two ionic strengths to determine the bindinglocations and affinities of the mutant DNAs. Because these experiments were
de-9
on November 10, 2019 by guest
http://jvi.asm.org/
Dtein 0 SVASVA
rl i11"..
.005.005 .125
O SVA SVA
I
a..
0*0
G G _
A*: .
G 4[I
G
G
,I*
G G
G a
G mo _ _
G _w -_ _
.-W
G
G 'd_ Amm W
G ._m ...W 4,
....I
... la,
C
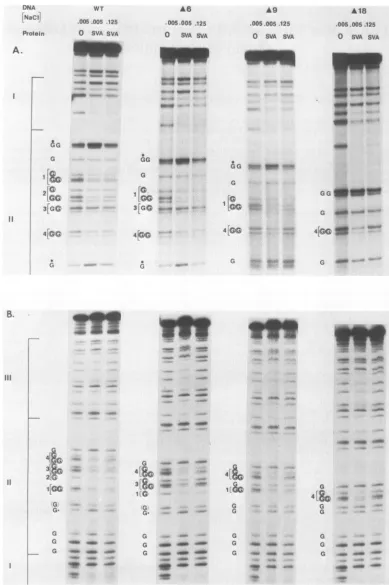
FIG. 1. DMS footprints of theSV40Aproteinboundtoboth strands of the origin regions of wild-type (WT) andmutantDNAs. Restriction
fragmentswereexposedtoDMS in thepresence orabsence ofAprotein in0.005 MNaCIor0.125 MNaCl and processedasdescribed inthe
text.Allguanine (G)residues of region11areplacedonthesamelinewhere guaninesoccuratadjacent positions inthe sequence,andsmall
brackets numbered in an increasing orderidentify recognition pentanucleotides. A circled G representsguanines protected at low ionic
strength, andadottedGrepresentshypersensitiveguanines. (A)NcoI-Hinflorigin fragmentwaslabeledonthe 3' late end of the early strand.
(B) TheBstNI-Hinfl origin fragmentwaslabeledonthe3' early endofthe latestrand.A6,A9,andA18ared16,d19, andd118,respectively.
10
.005.005 .125
0 SVA SVA
.005.005.125
o SVA SVA
V,'A$4." NW-*-*~
*PA
"WV
Pro
A.
II
B.
III
tiG G
1[S
2sa
1..
G
G
[I
.C
;....4
'.'
.,:,::GG
_
_G
I
la,
34[GflC4
4..
a
G G
10
VW_
G .W:--:,- """
C[ 1
4ie:C
.., 'T
_*#._OA
G[
,-ft¢ ''
31& a
:1
IG
4iG
3r
2'1
G
G-_ m
G
G-G G G
.av
I,*T:
*_
s." a ..
G G
C
.... o
.4*
11
.- '.
r
1114b
--.MI,
f,".
-!r 17...
-"Jwk
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.117.506.83.668.2]PROTEIN A BINDING TO SV40 ORIGIN DELETIONS 11
signed to compare the strength of binding with various
origins, A protein concentrations were adjusted to protect 50 to 70% of wild-type origins from DMS. Excess concentra-tions of protein would have decreased the sensitivity of the comparative assay.
Protein contact sites on the early and late strands are shown in Fig. 1A and B, respectively. The large brackets to theleft ofthefigure outline DNase protection
regiQns
I, II, and III to be described below. All of the mutant DNAs maintain the wild-type guanine sequenceand DMSprQtec-tion pattern in regions I andIII. The results are described in detail and correlated with the DNase protection analyses
below. However, it is evident that deletionsof pentanucleo-tides inregion II have little effect on binding to the remaining
recognition sites at 0.005 M NaCl and have variable effects on binding at 0.125 M NaCl, depending on the extent and
location of the deletion.
)Nase footprinting. The limits ofSV40A protein bound to DNAweredefined by the use of DNase footprinting. In this assay each molecule of DNA is cut once producing a ladder
offragments displayed by gel electrophoresis. When the DNAis firstboundwithprotein and thencut, gaps appear in the ladder representing the region of DNA protected by T
A.
DNA
Protein
WT A6 A9 A18
0 SVA 0 SVA 0 SVA 0 SVA
antigen. Double-stranded restriction fragments containing
theoriginofreplicationwereradiolabeledatthe3' endof the early or late strands (Fig. 2A
and
B). The figure showsDNase protection patterns obtained under low salt condi-tions. A protein protects wild-type DNA in the regions
indicatedbythebracketstotheleft of thegels(Fig. 2). Each regionis flankedbyDNasehypersensitive sites and each has
acharacteristic stabilityatdifferentsaltconcentrations(15).
Nevertheless, the boundary between regions I and II is somewhat arbitrary because A protein bound to region I sometimes protects DNA in theearly endofregion IIfrom
DNase(15). As shown inFig. 2, deletions inregion II have little if any effectonbinding ofAproteintoregions IandIII. Protein protects all of the undeleted region II of mutantsd16
andd19 but fails to protecttheearly endof regionIIofd118.
The hypersensitive sites flanking each end of region II are
gradually lost with increasing deletion size. Thus, these sites mustbe determined, in part, by binding to region II.
The DNaseprotection appears to be more complete than DMS protection even though the assays were carried out underthe same conditions. This difference is typical and can be explained in several ways. First, protection from the larger DNase molecule is indeed more efficient because of
B. DNA
Protein
WT *6 *9 A18
0 SVA 0 SVA 0 SVA 0 SVA
I *
I:
_
D
__
Se-e
a-
_OeO
.-0Shrr*
aa"
III
11
r
ma
_0 as
mm _ 1 . S.:"
o _ _ Y:
4* _S
_s
0
*,
do
_0d
* * 4'
[image:3.612.71.528.328.692.2]_ - ...
FIG. 2. DNase footprints ofthe SV40 A protein bound to both strands of the origin regions in wild-type (WT) and mutant DNAs.
Restrictionfragmentswerecutwith DNase in thepresence orabsence of A protein in0.005 M NaCl and processedasdescribed in thetext.
(A)TheBstNI-Hinfl origin fragmentwaslabeledonthe 3' late end of the early strand.(B) Thesamefragmentwaslabeledonthe 3' early end
of thelate strand. *6, *9, and *18ared16, d19, and d118, respectively. II
111I I VOL. 49, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
12 LEWTON, DELUCIA, AND TEGTMEYER
the size of the probe. Second, protection by subunits ofA protein bound to adjacent pentanucleotides provides do-mains of overlapping DNase protection.
DISCUSSION
To understand the process ofinitiation of DNA replica-tion, wechosetofocus onthebindingofAproteintoSV40 DNAdeletedattheBglI restriction site in theorigin. Allthe
mutants are nonviable in permissive host cells. We used
DMS footprinting to define close contacts between origin DNAand A protein and DNasefootprintingtodeterminethe
broader limits of the DNA-protein interaction. The results from both studies aresummarized andinterpretedinFig. 3.
In the case of wild-type DNA, A protein protects all circledguanines (Fig. 3)withinthe fourpentanucleotides of region II and two of three guanines between the
pentanu-cleotides. Weak protectionofanearby guanine isindicated byparentheses andhypersensitive guaninesaredotted(Fig.
3). Protein alsoprotectsthe entire region from DNase. The
arrowbelow each pentanucleotide shows ourinterpretation
of the DMS and DNase protectionresults(15). Inourmodel,
each pentanucleotide directsthebinding ofamonomerofA protein inanorientation thatprotects20bpofDNAatthe 5'
endof the 5'-GAGGC-3' sequence and 10bp at the 3' end
(15).
Mutant dl6 deletes site 2 completely without alteringthe
single-bp spacing
between theremaining sites. Proteinbind-ing
protects all the remaining pentanucleotides from DMS and induceshypersensitivity
oftheoutlyingguanines of themutantDNA in thesamepattern asin thecaseof
wild-type
DNA.
Protein
also protects the entire undeleted portion ofregion
IIfrom DNase. Both DMS(Fig. 1) and DNaseassays(data
notshown)
indicate that binding isonly slightlysensi-tive to0.125 MNaCl. These resultsare consistent withthe
predictions
of ourbinding
model (15). The loss of site 2would decrease the number of protein subunits bound to
region
IIbutwouldnotaffect other interactionswithpentan-ucleotides.
Mutantd19 deletes allofsite2butonly apartof site 3 so
that the
remaining
pentanucleotides are separated by four rather than onebp.
At low salt concentrations, protein protects sites 1 and 4 from DMS and the entire span ofundeleted
region
II from DNase. It is interesting that thisdeletion also creates a new 5'-GAGGC-3' sequence that
overlaps
site1 andis separated fromsite 4 byasingle bpasshown
by
thebracketsinFig.3.The DMS protectionpatternII
-- -- -- -- -- --- -- -- -- -- -- ----
--30 20 10 0 10 20 30
WT I 2 3 4
C T A C T T C T G G A A T A G C T C A@A ( CC A0@0tGI C C T C6@C C T C|T G A T A A A T A A A A A A A A T T A G T GA T G A A G A C C T T A T C O A(G)T|C T C C C T C C CC C A ACO T A T T T A T T T T T T T T A A T C A
I
r
AOk5D1) 1 4 3 4
C T A C T T C T G G A A T A G ,C T C A A(G(GC G|QC C T C C C T C|T G C A T A A A T A A A A A A A A T T A G T
GA TG A A GA C CT TAT CO A(G)T|CT C CC CCCAC CTCAfAA CO TAT T TAT T T T T T T AT C A
A9
*9(6-1)
(6-17)
.
1
1 4...4
C T A C T T CTOGOC AA TAOCCT C A©
AI
C C T Cj©X C C T C:T G C A T A A A T A A A A A A A A T T A G T GAT GA AGA C CTTAT C GATG T cL ATAAGJC:C AACO TAT T TAT T T T T T T AT C AI a -...-.-...
*18
(295)*
~~~~~...
. . .. . . . .. . . .Ala
(295S)
4 .C T A C T T C T 0 0 A A T A 0 C T C©C):VC C T C:T 0 C A T A A A T A A A A A A A A T T A 0 T
O A T 0 A A 0 A C C T T A T C 0 A 0 C*C©©@A©@A C 0 T A T T T A T T T T T T T T A A T C A
FIG. 3. Methylationand DNaseprotectionresults. TheregionIIsequences ofwild-type(WT)andmutantDNAsareshownand numbered
by themethod ofFierset al.(4). Theupper and lower strandsarethe earlyandlate strands,
respectively;
thecurvedarrowaboveeachsequence specifies the location of the deletion. Boxesidentifyrecognitionpentanucleotides numbered inan
early
tolatedirection. Dotted boxes indicatesites thatareincompletely protectedat0.125 M NaCl. Circledguanines(G) indicateDMSprotection,
and theparentheses signifyweakprotection. Largeand small dotsaboveguaninesindicate differentdegreesofhypersensitivity.
Theelongated
arrowsbelowtherecognitionsequencesrepresentourinterpretationoftheDNaseprotectionresults. Eacharrowcorrespondstothe DNase
protection
domain ofamonomerofproteinboundto asinglepentanucleotide (15).Thebracketed pentanucleotidesiteinmutantd19representsanewly
formedcontactsitethatisapparently unused. A6, A9,and A18aredl6,d19,andd118, respectively.
J. VIROL.
A e r -4N
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.562.330.654.2]PROTEIN A BINDING TO SV40 ORIGIN DELETIONS 13
indicates that A protein prefers bindingto site 1 ratherthan
tothe new site. Furthermore, bindingtosite1 is more stable
than binding to site 4 at 0.125 M NaCl. These results are consistent with ourpreviousfindingthatbindingtoregion II occurs in a sequential process in an early to latedirection
(15). We conclude that thehigheraffinity of bindingtosite 1 does not depend onadjacentrecognition pentanucleotidesin
region II.
Mutantd118deletes all of sites 1, 2,and 3 so that site 4 is theonly siteremaining in region II; it occupiesthe original position of site 1 relative to region I but in the opposite orientation. A protein protects this single recognition
se-quence atlow, but not at high, ionic strength. Thus, high-affinitybinding to site 1 maydepend notonly onsequences orprotein-proteininteractionsontheearlyside ofregion II, butalso onthe orientation ofthepentanucleotide. Bindingto
site 4ind118provides the first direct evidence that confirms
ourmodel for the mechanism ofAproteinbinding.Asingle
recognition sequence directs binding of a unit protein and
results in asymmetricDNaseprotection.Theprotectionofa guanineadjacenttosite4raisesaquestionastowhether the
recognition sequence isapentanucleotide or a hexanucleo-tide. We prefer the formeralternative because aguanine in the sixth position is not consistently present and more importantly isnot alwaysused. Forexample,theguanine at thecenteroftheregion II palindrome isnotprotected
by
A protein(Fig. 1Aand 3).The deletions in region II change notonly thenumberof
contact sites but also the distance and rotational angles between remaining sites. Yet, the A proteincan
accommo-date these changes and engage all available sites under low salt conditions. These results can be easily explained ifthe
primary binding form is a monomer. Binding ofpreformed dimersor tetramers toaltered DNAwouldrequire consider-ableflexibility of proteinsubunits. Studiesusingquantitative
transmission electron microscopy (I. A. Mastrangelo, unpub-lished data) demonstrate that A protein monomers are frequent binding forms in region II, although dimer and tetramerforms are also readily seen at higher protein
con-centrations. Even though monomers may be the primary binding form, protein-proteincontactswouldaccountforthe
higher affinity binding evident when recognition sequences
correctly position monomersrelative to oneanother. The 27-bp palindrome in the origin of replication could
have a number of additional functions as aconsequence of
protein binding. For example, binding to the palindrome
could induce a cruciform structure, initiate melting in the
adjacent AT-rich sequences,oralterconformationin
anoth-ermanner.Interestingly, oneofthemajorinitiation sitesfor RNA priming ofDNA replication appears to occur within thethird pentanucleotide in theprotein-binding palindrome (7). Theproduction ofaprimercouldrequire either de novo
synthesis or possibly processing of preexisting early
tran-scripts
by an RNase H-like activity at this site. Thus,deletions in this region could have multiple effects on the
initiation of replication. Mutant
d16
deletes site 2 but maintains a single-bp space between the remaining sites so that the spacing and rotation of bound protein subunits would not be altered. This positioning effect is likely toaccountfor the stablebindingobserved. Because this mutant isnotviable, we conclude that binding of four monomers of
Aprotein in specific positions and orientations is a
require-ment for successful DNA replication. Additional possible functions of the palindrome in DNA replication, if any,
remain tobe demonstrated.
ACKNOWLEDGMENTS
This work was supported by PublicHealth Service Grants CA-18808 and CA-28146fromtheNationalCancer Institute.
WethankY.Gluzman and R. Martin for theirdeletionmutants. LITERATURECITED
1. Danna,K.J.,andD. Nathans. 1972.Bidirectional replicationof simian virus 40 DNA. Proc. Natl. Acad. Sci. U.S.A. 69:3097-3100.
2. DeLucia,A.L.,B.A.Lewton,R.Tjian,and P.Tegtmeyer.1983. Topography of simian virus 40 A protein-DNA complexes: arrangementofpentanucleotideinteraction sites at theoriginof replication.J. Virol. 46:143-150.
3. DiMaio, D., and D. Nathans. 1980. Cold-sensitive regulatory mutantsofsimian virus 40. J. Mol. Biol. 140:129-142. 4. Fiers, W., R.Contreras, G. Haegeman, R.Rogiers,A. Van de
Voorde,H.VanHeuversewyn, J.VanHerreweghe,G.Volckaert, andM. Ysebaert.1978. Complete nucleotidesequenceof SV40 DNA. Nature(London) 273:113-120.
5. Gluzman, Y.,R.J. Frisque, and J. F. Sambrook. 1980. Origin-defective mutants ofSV40. ColdSpringHarborSymp. Quant. Biol.44:293-299.
6. Gluzman,Y., J. F. Sambrook, and R. J. Frisque. 1980. Expres-sion of earlygenesof origin-defectivemutantsof Simian Virus 40. Proc. Natl. Acad. Sci. U.S.A.77:3898-3902.
7. Hay, R. T., and M. L. DePamphilis. 1982. Initiation ofSV40 DNAreplication in vivo: location and structure of5' endsof DNAsynthesized in theori region. Cell28:767-779.
8. Kahn, M., R. Kolter, C. Thomas, D. Figurski, R. Meyer, E. Remaut, and D. R. Helinski. Plasmid cloning vehiclesderived from plasmids ColEl, F, R6K and RK2. Methods Enzymol. 68:268-280.
9. Maxam, A. M., and W. Gilbert. 1980. Sequencingend-labeled DNA withbase-specific chemicalcleavages.Methods Enzymol. 65:499-560.
10. Myers, R. M., and R.Tjian. 1980.Constructionandanalysisof simian virus 40origins defective in tumorantigen binding and DNAreplication. Proc. Natl. Acad.Sci. U.S.A. 77:6491-6495. 11. Shortle,D., and D.Nathans. 1979. Regulatorymutantsof simian virus 40: constructed mutants with base substitutions at the origin ofreplication. J. Mol. Biol. 131:801-817.
12. Tegtmeyer, P. 1972. Simian virus 40 deoxyribonucleic acid synthesis: the viralreplicon.J. Virol. 10:591-598.
13. Tegtmeyer, P., and B. Andersen. 1981. Partial purification of SV40A protein anda relatedcellularprotein from permissive cells. Virology 115:67-74.
14. Tegtmeyer,P., B.Andersen,S. B.Shaw,and V.G. Wilson.1981. Alternative interactions of the SV40 A protein with DNA. Virology 115:75-87.
15. Tegtmeyer, P.,B. A.Lewton, A. L. DeLucia, V. G. Wilson, and K. Ryder.1983.Topography of simian virus40 Aprotein-DNA complexes: arrangement of protein bound to the origin of replication. J. Virol. 46:151-161.
16. Tenen,D.G.,L. L.Haines,and D. M.Livingston. 1982.Binding ofananalog of the simian virus40 Tantigen towild-type and
mutantviralreplication origins.J. Mol. Biol. 157:473-492. 17. Tjian,R.1978.Protein-DNAinteractionsatthe origin of simian
virus 40 DNA replication. Cold Spring Harbor Symp. Quant. Biol. 43:655-662.
18. Wilson, V. G., M. J. Tevethia, B. A. Lewton, and P. Tegtmeyer. 1982. DNAbinding properties of simian virus40 temperature-sensitive Aproteins.J. Virol.44:458-466.
VOL.49,1984