Copyright© 1976 AmericanSociety for Microbiology Printed inUSA.
Further
Characterization of the Friend Murine Leukemia
Virus Reverse
Transcriptase-RNase
H
Complex
KARIN MOELLING'
Institut fur Virologie, Bereich Humanmedizin, Justus Liebig-Universitdt 6300, Giessen,Germany
Receivedforpublication25November 1975
The purified reverse transcriptase-RNase H complex from Friend murine
leukemiavirus consistsofasinglepolypeptideof 84,000 molecularweight,which
after mild protease treatment in vitroor after intentional
degradation
duringthe purificationprocedure allows the generation ofseveral additional
polypep-tides. Degradation destroys theRNA-dependentDNApolymerase activitywith
native RNA templates and reduces RNase H butdoes not affect response to
synthetic template primers such as poly(rA)oligo(dT). The properties of the
intact murine enzyme consisting of a single polypeptide of84,000 molecular
weight arecomparedtothose ofthe avian asubunit andthe avian a/3enzyme
complex. Theintact murine enzymeresembles the avian /3-containing enzyme
complex andisdifferent from a inthefollowing respects: (i) itbindsto native
RNA templates; (ii) ittranscribesnativeRNAtemplatesinto DNA, areaction
whichcanbe inhibitedby actinomycin D; (iii) RNaseHactivity behaves likea
processive exonuclease; and (iv) analysis of the RNase H digestion products
reveals oligonucleotides approximately four basesin length.
TheRNA-dependent DNApolymerase
activ-ity from mammalian viruses, particularly the
murine ones, have been characterized by
sev-eral laboratories withconflictingresults. As far
asthestructureof theenzyme isconcerned, the
number of subunits identifiedon sodium
dode-cyl sulfate-polyacrylamide gels afterextensive
purification ranges from one to three. The
en-zymesfromFriendmurineleukemiavirus
(Fr-MuLV) and from Moloney murine leukemia
virus(Mo-MuLV) have beendescribedas a
sin-gle polypeptide of 80,000 to 84,000 molecular
weight (8, 12). In other reports the Fr-MuLV
enzyme has been characterized as a
two-poly-peptide complex (15), and the Mo-MuLV
en-zyme has been shown to consist ofthree
sub-units(2).Thetwopolypeptides observed for Fr-MuLV (15)resemble those of the hamster viral enzyme (13).
As the avian viral reverse
transcriptase-RNaseHcomplex consists of twopolypeptides,
aand
/3,
one ofwhich can be shown toarise byproteolytic cleavage from the other
(Moell-ing,Virology,inpress),the questionaroseasto
whether some of themurine viral enzyme sub-units represent degradation products as well. The answer ofthis question wasapproached in two ways: (i) the purified Fr-MuLV enzyme, consisting of a single polypeptide, was
sub-jectedto mild protease treatment in vitro; (ii)
Presentaddress: Max-Planck-Institut fur Molekulare Genetik,D-1Berlin-33 (West),Germany.
an enzyme preparationwasextracted from
Fr-MuLVundernonoptimal conditions for
conser-vation of the enzyme activity. Both experi-ments (trypsin treatmentand intentional
deg-radation) generated several polypeptides.
Structural changeswere correlated with
enzy-matic alterations that reproduce and explain
some of the conflicting data published on the
structure and enzymatic properties of
mam-malianviralreversetranscriptases.
Thesizeof the largestmurine viral enzyme
polypeptideranges between thesizeof thetwo
avian enzymesubunits, leavingopenthe
ques-tionof whetheritcorresponds to aor/3. There
is evidence in favor of either: the ability to
transcribe nativeRNA observed insome cases
(2, 8, 12)resembles thepropertiesof
/,
whereasabsence ofRNA-dependent DNAsynthesis (1,
14) is similarto a. The RNase H activity has beencharacterizedas a randomexonuclease (3)
identicalto thatexhibited by the avian a
frag-ments (3), and theRNase Hdigestionproducts (12) have been described as being larger than
expectedby analogy toa /3-containingenzyme
preparation. Furthermore, it has been observed that protease treatment of the avian
a,8
en-zymecomplex in vitro can completely remove
,/,
whereas a is much more resistant to this treatment. Only more stringent conditionsal-low digestion of a as well (Moelling,
Virol-ogy, in press). The possibility exists, therefore,
thatthelargestmurineviral proteindescribed
418
on November 10, 2019 by guest
http://jvi.asm.org/
sofar is adegradation product ofan evenlarger one.
To find out whether or not the 84,000-molecu-lar-weight polypeptide exhibits the properties of a real viral reverse transcriptase-RNase H complex, several of its enzymatic properties werecompared to those of the avian asubunit,
asrepresentative of a degraded enzyme, and to the
a,/
avian enzyme complex. As all previous attempts to isolate /3 alone have failed (Moell-ing, in press), the avian a,3 enzyme complex waschosen for comparison, since it is the best representative of a functioning reversetran-scriptase-RNaseHcomplex presentlyknown.
MATERIALS AND METHODS
Virus. Avianmyeloblastosisviruswasisolatedin partinthis laboratory andwas inpartagenerous gift of J. Beard. Fr-MuLV was grown inthis labo-ratory. Theviruswasoriginallyproduced in STU-mouse cells (10) that have sincebeenestablished as as permanent line (Eveline cells). The cells that were growninsuspension culture wereadapted to substrate-dependent growth by G. Pauliinthis lab-oratory. Approximately 107 cells were seeded in roller bottles (5-cm diameter, 50-cm long) and grown at37C with25mlof Dulbecco modifiedEagle growth mediumin the presence of5% heat-inacti-vated calf serum (56 C, 45 min) and 10% tryptose phosphate broth.
Reagents. 3H-labeled poly(rA) and poly(dT) were obtained from Miles Laboratories. Oligonucleotides (A)A, (A)6A, (A)4A,and(A)2Awerepurchased from Boehringer (Mannheim, WestGermany). Acrylam-ide and methylenebisacrylamide, electrophoresis grade, were bought from Bio-Rad Laboratories
(Richmond, Calif.). Soluene-350 originated from Packard Instrument Co. (Zurich,Switzerland). Dul-becco modified Eagle medium was purchased from Flow Laboratories (Bonn, West Germany), and tryptosephosphate broth was from Difco Laborato-ries (Detroit, Mich.).
Enzymepurification. The reverse transcriptase-RNase H complex from avian myeloblastosis virus and Fr-MuLVwasisolatedbypublishedprocedures (8, 9), anditspurity wasdetermined on SDS-poly-acrylamide gels. Subunit a was recovered from a
phosphocellulose column at 0.11 M KCl. For isola-tionoflarge amountsof a, theenzyme attachedto the DEAE column was left at 4C for 1 to 2 days before elution. The same treatment wasappliedto theFr-MuLVenzymetodeliveradegraded enzyme. Enzyme activity assay. DNA polymerase and RNase H were determined aspreviously described (8, 9). The avianenzyme assayswereperformed in the presence of 8 mM MgCl2, and the Fr-MuLV assays wereperformed in the presence of 0.4 mM MnCl2. Thespecific radioactivity ofonelabeled
de-oxyribonucleotide was 5,000 counts/min per pmol.
Theassayfor RNase Hcontained10,000counts/min ofphage fd DNA-RNAhybrid,withspecificactivity of3,500 counts/min perpmolof[3H]UMP.
Unit of activity. A unit of enzyme activity is
defined as the amount of enzyme required to incor-porate 30pmol of TMP at 39C in 30 min with the synthetic templatepoly(rA)-oligo(dT) inastandard polymerase assay mixture, as this reaction is not affected by any degree of enzymedegradation. Com-parative studies ofenzyme activities were always based on identical units input.
Binding of enzyme to RNAtemplates. In a total volume of 100 ,l, 20 tkl of the murine or avian
/3-containing enzymes were mixed with an excess of RNA (5jigof70S AMV RNA)inthepresence of ions asusedinenzyme assays.Forbinding analysisofa, 100,ul of enzymewasmixed with 5 ,ug of70S AMV RNA under assay conditions in 200 ,l (final vol-ume). Withoutfurther incubation the material was layered on top ofpreformed glycerol density gra-dients (10 to 30% glycerol in TNE [0.01 M Tris-hydrochloride-0.1 MNaCl-1 mM EDTA], pH 7.4-5 mM dithiothreitol-0.2% Nonidet P-40). After cen-trifugation for 90 min at 50,000 rpm and 4 C in a SW50.1Beckman rotor, thegradients were fraction-ated. Position of the enzyme was determined by testing an aliquot (20,u)of eachfraction for enzyme activitybythe addition of fourdeoxyribonucleoside triphosphates and ions toestablish regular enzyme assay conditions. Alternatively, poly(rA) oligo(dT)
andL3H]TTPcan beadded to each fraction for
deter-mination ofresponse tosynthetictemplate-primers (in thecase ofathistesthastobeapplied, since a
does not transcribe native RNA).
Chromatography of 3H-labeled poly(rA) diges-tion products. 3H-labeled poly(rA) oligo(dT) (5,000 counts/min per pmol of AMP) was hydrolyzed by viral RNase H activities in standard assay mix-tures.After the reaction themixturewasspottedon strips (50-cm long) of Whatman no. 1 paper and
chromatographed in n-propanol-ammonia-water (55:10:35) for48 h to separate variousoligomers of AMP (5). Portions (50 to 100 ,ug) of (A),A, (A)6A,
(A)4A, and (A)2A were added as standards to the reactionmixturesbefore spotting. The paperwasair dried after chromatography, the position of the standardswaslocatedby UV light, and1-cmstrips were cut. Radioactivity wasdeterminedin toluene-based scintillation fluid. Thestandardsappliedhere donot have terminal phosphates and therefore do notmigrateexactly likephosphorylated productsof RNase H digestion. Therefore no precise assign-ments for thelength of the oligomerswerepossible.
Polyacrylamide gel electrophoresis. Enzyme
polypeptides wereanalyzed by sodiumdodecyl
sul-fate-polyacrylamide gel electrophoresis accordingto amodified method ofShapiroetal. (11)asdescribed byKacian etal. (4). Beforeapplying proteinstothe
gel, concentration was achieved by precipitation
with50%ethanol (for solubilizationof thedetergent NonidetP-40) and 10% trichloroacetic acid (-20C,
10h). Protein waspelletedinaSorvallHB4rotor at
10,000 rpmfor 30 min,dried, andsuspendedin
sam-ple buffer for electrophoresis.
[3H]glucosamine-la-beled virus (Prague A [PR-A] strain ofRous sar-coma virus) was used as internal standard (1,000
counts/min per gel). The so-called gp85 and gp37 glycoproteins migrate more slowly in the buffer system used (13) than their names indicate. The
on November 10, 2019 by guest
http://jvi.asm.org/
420 MOELLING
gels were stained with Coomassie blue, and the positions of thepolypeptides weredeterminedbya Gilford scanningapparatus at 600nm wavelength. Afterwards the gels were frozen, sliced (1 mm),
andincubated in 200
A.l
of soluene for 2 h at 60 C. After the addition of 2 ml of scintillation fluid, radioactivity was determined in a liquid scintilla-tioncounter.RESULTS
Structural properties of the Fr-MuLV
re-versetranscriptase-RNaseHcomplex. The re-verse transcriptase-RNase Hcomplex was
iso-lated from purified Fr-MuLV by
DEAE-cellu-loseandphosphocellulosecolumn
chromatogra-phyaccordingtopublished procedures (8).The
purifiedenzymeconsists ofasinglepolypeptide
with anestimated molecularweightof84,000,
basedonthe sizeof thetwoavianenzyme
sub-units 13andabeing 110,000 and70,000(4)(Fig.
1AandE). Mild proteasetreatmentunder
con-ditions thatallow increase ofa atthe expense
of
(8
inthe avian system(7)generated,fromthe84,000-molecular weight polypeptide,new poly-peptidesof 78,000, 69,000,and60,000molecular
weights, andalargeamountof
low-molecular-weightmaterial. Increasingamountsoftrypsin
(5 to 30
/ig/ml)
predominantly resulted in an increase in theproportion ofthe78,000-molecu-lar-weightpolypeptide (data notshown).
Fur-thermore, the enzyme wasextracted from the
purified virus under conditions that allowed
transfer of the avianenzyme to a preparation
consistingpredominantlyofpure a.Thiscanbe
achieved by binding the enzyme to a DEAE
column for1 to2daysat4Cbefore elution with
acontinuoussaltgradient (Moelling, Virology,
inpress).Suchatreatmentcauseddegradation
of themurine enzyme aswelland givesrise to a
preparation of 84,000-, 78,000-, 69,000-, and
60,000-molecular-weight
polypeptides inaddi-tion to smaller material (Fig. 1C). This poly-peptide patternwas obtainedtwice. Inathird
case the 78,000 polypeptide wasmissing (data
not shown). Storage at -20C increased the
69,000polypeptidepreferentially.Elutionofthe
enzyme from phosphocellulose by means of a continuous salt gradient inaregular
purifica-tion procedure consistently revealed a small
peakof activity at a low salt concentration(0.11
M
KCl)
inadditiontothe main peak at 0.22 MKCl
(notshown). The firstpeak correspondstothe region from which the subunit a in the aviansystem can berecovered (3). Its
polypep-tide composition (Fig. iD) consisted of
78,000-and69,000-molecular-weight bandsinaddition
to the84,000 andsmaller ones. Purified avian
af-containing
enzyme is shown for control.Figure iF shows the isolated avian a subunit
x
C-)
E
0.3-Fr-MuW
"degraded'
0.5
U)
.0
migration
FIG. 1. Scanning profiles of murine and avian viral enzymepolypeptides after gel electrophoresis. (A) PurifiedFr-MuLV reverse transcriptase-RNase H; (B)purified Fr-MuLV enzyme after trypsin treat-ment (25
H.g/ml,
15min,
35 C); (C) purified Fr-MuLV reverse transcriptase after long-lasting col-umn chromatography (attachment to DEAE-cellu-lose for 1 or2days at4 Cbeforeelation);(D)marinematerialelatedat0.11MKC1from the phosphocellu-lose column(corresponding to theelationpoint of the avian asubunit); (E)purifiedavianaf3enzyme prep-aration; (F) avian subunit a aselatedfrom the phos-phocellulose column and also obtained after in vitro protease treatment of the purified af3complex (40pg
oftrypsin, 35 C, 20 mm). The enzymepolypeptides were subjected toelectrophoresis in the presence of [3H]glucosamine-labeled virus (2,000
counts/mmn;
Rous sarcoma virus, Prague A) used as internal standard in all gels except for Fr-MuLV "a." The gels were stained first and subsequently processed for determination ofradioactivity as described. In(A)thedottedline indicates theradioactivemarker
which is onlyshown by arrows in(B)through (F).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.264.447.44.452.2]REVERSE TRANSCRIPTASE-RNase H COMPLEX 421
thatcanberecoveredassuch from the
phospho-cellulose column at low ionicstrength. It can,
however, also be generated from the
a/3-con-tainingenzyme complex by protease treatment,
which completely removes 13 (Moelling,
Vi-rology,inpress).Onlymore stringentdigestion
conditions will also digest a. This effect is
shown since itdemonstratesthat thereis areal
possibility of preferentially losingone subunit
andonlydetectingacomparativelymorestable breakdown product.
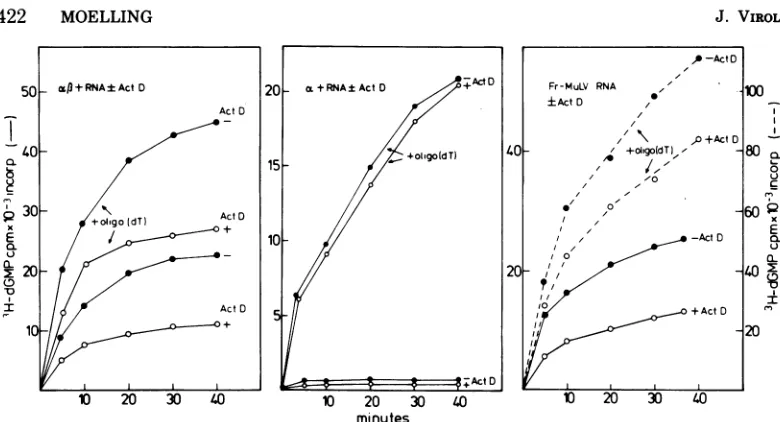
Binding to natural RNA and RNA
tran-scription. The Fr-MuLVreverse
transcriptase-RNase H complex was tested for its ability to
bindtonative RNAtemplates, and itsbinding
propertieswerecomparedtothose of the avian
subunit a and the avian a13 complex (Fig. 2).
The murine enzyme, just like the avian a,8
enzyme, bound to the high-molecular-weight 70S AMV RNA and could be sedimented
through a glycerol density gradient with the
viral RNA. It was then detectable by enzyme
activityassay intheregionof the
high-molecu-lar-weight RNA. Incontrast, acould notbind
toRNA andstayedontopof thegradient under
these conditions. Asexpected from theabilityof
the murine enzyme to bind to native RNA, it
was also able to transcribe native RNA into
DNA (Fig. 3). Addition ofoligo(dT) stimulated
DNA synthesis severalfold. Both reactions
could beinhibited by actinomycin D, with the
reactionwithoutoligo(dT) inhibited about50%.
ActinomycinDinhibitionhasbeenshown tobe
indicative of synthesis of double-strandedDNA
(6).
Native RNAtranscription was also analyzed
with the degraded murine enzyme (Fig. 1C).
RNA-dependent DNA synthesis with native
RNAwas not seen(Table 1).Onlytheaddition
ofoligo(dT) overcame this defect. Actinomycin
Ddidnot inhibitthis reaction. The same phe-nomena have been observed for the avian a subunit(Moelling, inpress)and arelisted here for comparison. The properties of the
unde-graded murine enzymeand those of the avian
a,/
complex are shown in Table 1 as well.RNase H activity of the degraded murine
en-zyme was reduced by about 30% (not shown)
comparedtothe undegradedone.
Mode of action of murine viral RNase H
activity. Themurine viral RNase H presentin
the 84,000-molecular-weight polypeptide has
been characterizedas anexonuclease (8, 12).If
this RNase H behaves like the one from the
avian a,3 complexit should degrade one RNA
strand completely beforeproceedingtoanother
one. In contrast, the RNase H of the avian a
subunitattacks a new RNA strand after each
bondscission (3).
The RNase H activities of the Fr-MuLV
en-zyme consisting only of the
84,000-molecular-weight polypeptide and of a degraded murine
viralenzyme preparation weretested fora
ran-domorprocessive mode ofaction. Behavior of theavian a-anda/3-containingenzyme prepa-rations are shown forcomparison (Fig. 4).
Ra-dioactively labeled hybrid was hydrolyzed by
RNase H from enzymes of both murine and
avian viral origin. If the radioactively labeled
hybrid was competed for with tenfold molar
20 200 10
E
I~~~~~~~~~~~I
10 20 10 20 10 20
factions
rVIG. 2. Bindingproperties ofthe murine enzyme and avian aand af3 toRNA. Purifiedenzymes were
mixedwith an excessofnativeviral RNAandsedimentedwithoutfurther incubationunderconditions which allowthe RNAtomigratetwo-thirds down thegradient (arrows).Thegradientswerefractionated,and each fractionwastestedfor thepresenceofenzymeactivitybyresponsetosynthetictemplateprimers(seeMaterials andMethods)inthecaseofAMVaandaf3.RNA-dependentDNApolymerase activityinthecaseofFr-MuLV wastestedbytheadditionof fourdeoxyribonucleoside triphosphates (andnoadditionaltemplate).
on November 10, 2019 by guest
http://jvi.asm.org/
x
E
IL
I
0
I
10 20 30 minutes
0
I
-o
v
x
E
[image:5.503.63.453.53.264.2]I
FIG. 3.Kinetics of RNA transcription by purified AMV
a,-containing
enzyme, pure a,andpurified Fr-MuLV reversetranscriptase-RNaseH.The three enzymepreparationswereincubated with2Mgof70S AMV RNAinthe absenceand presenceofoligo(dT)primersunderstandard assay conditions().Furthermore,DNAsynthesis ofthese reactions was investigated in theabsence (-) and presence (+)ofactinomycin D (100 pg/mi). Aliquots were withdrawn from the reaction mixtures at the times indicated andprocessed for
determination of acid-insoluble radioactive material.
TABLE 1. RNA-dependentDNA-synthesisofdegraded and undegradedFr-MuLV reverse transcriptase (RT) compared with that of theavian aandaf3enzymesa
Template Degraded Undegraded Avian a Avian a/3
Fr-MuLV Fr-MuLV
RT RT
RNA 0.4 6 0.05 5.5
RNA +oligo(dT) 18 25 5.5 11
RNA +ActD 0.5 2.5 0.05 2.4
RNA +oligo(dT) +ActD 19 17 5.4 6
aFr-MuLV reversetranscriptase was purified under nonoptimal conditions(long-lasting column
chroma-tography),and its ability to transcribe native RNA was compared with that of undegraded Fr-MuLV reverse transcriptase (see Fig. 1)and to that of avian a- and to aviana,8-containingenzyme. Identical enzymeunits,
defined in Materials and Methods, were applied. A1-Mugsample of70S avian myeloblastosis virus RNA was used astemplateineach reaction, and 1 Mugofoligo(dT) and 100Mugofactinomycin D (ActD) per mlwere
added to the reactions asindicated. The amount of acid-insoluble material was determined and expressed as picomoles of[3H]dGMPincorporated.
excessof unlabeledhybrid2minafterthestart
of thereaction, therateof RNAdegradation by
theRNaseHfrom theintactFr-MuLV enzyme and the avian a,8 enzyme was not affected, whereas the reactions catalyzed by the
de-gradedmurine enzymeorbythe avian awere
severelyreduced. Addition ofexcess unlabeled
hybrid to the reaction mixtures prior to the
enzymesdidnotreveal any RNase H activities. The intact murine enzyme, once bound to its
substrate, proceeded to hydrolyze its RNA
strand,justlike the aviana,3complex,whereas
a degraded murine enzyme behaved like the
avian asubunit. This reactionwascarried out withasynthetichybrid,
[3H~poly(rA)-poly(dT),
as well as with a phage fd 3H-labeled
RNA-DNA hybrid (not shown), with identical
re-sults.
Murine viral RNase H digestion products. L3H]poly(rA)-poly(dT)wasincubated with iden-tical units ofFr-MuLVenzyme,avian af, and
a under conditions that convert 70% of the
[3H]poly(rA) into acid-soluble material. The
size of the digested products was analyzed by paperchromatography, which allows the sepa-rationofmononucleotides andoligonucleotides
up to eight bases in length, the position of
whichcanbeestimated bytheuse of
fluoresc-ing standards. Fr-MuLV and the avian af3
RNase H activities gave rise to similar
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.60.454.359.432.2]amounts of small-sizedoligomers (possibly tet-ramers; see above), whereas no monomers could be detected (Fig. 5). In contrast, a did not
giverise to smalloligonucleotides but all of the
material remained at the origin, indicating a
sizelarger than eight bases. The untreated
con-trolalso did not migrate. Solubilization of 20 or
90%ofthehybrid or use of the phage fd
DNA-RNA hybrid gave the same results (not shown).
DISCUSSION
The Fr-MuLV reverse transcriptase-RNase
H complex has been previously described as a
single polypeptide of 84,000 molecular weight
(8).The present study shows that this
polypep-tide reveals all the properties known for the
reverse transcriptase-RNase H complex of the
avian viruses. These properties are as follows:
(i) the ability to bind to natural RNA and to
transcribe it without the addition of synthetic primers; (ii) inhibition of DNA synthesis by
actinomycin D, indicating double-stranded
DNAsynthesis;(iii)co-purification of an RNase
H; (iv) exonucleolytic mode of action of the
RNase H activity; and (v) oligomers,
approxi-mately tetranucleotides, as the main RNase H digestion products and the absence of mononu-cleotides.
The Fr-MuLV reverse transcriptase-RNase
Henzyme complex undergoes spontaneous
deg-radation inside the virion due to freezing and thawing or during purification by long-lasting
column chromatography, giving rise to
addi-tional polypeptides of 78,000, 69,000 and 60,000
molecular weight (Fig. 1). Degradation of the
enzyme can also be achieved artificially byin
vitrotreatmentwith trypsin, whichresultsina
similar polypeptide pattern.
Enzyme preparations consisting of two or
threepolypeptides have been described by
oth-ers(2, 13, 15), with sizedeterminationssimilar
totheones observed here. With such degraded
murine viralenzymepreparations it was
possi-ble to reproduce some of the deficiencies
de-scribedfor mammalian viralreverse
transcrip-tases; degradation of an enzyme preparation
causes loss of the ability to transcribe native
RNAtemplates (Table 1). It can, however, be
drastically stimulated to synthesize DNA if oligo(dT) primers are added to the natural RNA
template. These results obtained with a
de-graded enzyme preparation explainpreviously
published results on mammalian reverse
tran-scriptases (1, 13, 14). Response to synthetic
template primers is unaffected by any degree of
enzymedegradation. Reduction of RNaseH
ac-tivity during degradation has been observed
consistently in these studies; however,
com-plete loss of RNase H couldneverbe achieved
with theRNase H assay applied here.
Only two studies have beenpublished
show-minutes
FIG. 4. Competition ofhydrolysis ofradioactively labeled hybrid by the addition ofexcess unlabeled
hybrid.Purifiedreversetranscriptase-RNaseH activities wereincubated with[3H]poly(rA)-poly(dT) under standard assay conditions. At thetimesindicatedportionswerewithdrawnfromthe reaction mixtures and
processedfordeterminationofacid-insoluble material(C).Inaparallelexperimenta10-foldmolarexcessof
unlabeledpoly(rA)-poly(dT)wasadded2 minafterthestartofthe reaction(-).Inacontrol reactiona10-fold
molarexcessofunlabeledhybridwasalreadyaddedtothereaction mixturepriortotheenzyme(A).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.110.397.403.598.2]C13
x 20
AMV a1
20!
AMVa3 -iContro
10
20
30
40
50
fractions
FIG. 5. Chromatography of digestion products. Fr-MuLVRNaseHdigestion products obtained from
areactionthat transferred70%ofthe
[3H]poly(rA)-poly(dT) hybrid into acid-soluble material were
chromatographedto separateoligomers ofAMP. The
sameexperimentwasperformedwith identicalunits
ofavian aandaf3enzyme inputs. Untreated hybrid
is shown as control. Included standards were
lo-catedbyUVlight.
ingamurine enzymepreparation consistingof a single polypeptide around 80,000 molecular weight (8, 12); in athirdcasethispolypeptide
appearsamongothers (2). In these three
stud-ies natural RNA transcription has been ob-served. Therefore, the84,000-molecular-weight polypeptide appears to be necessary for
tran-scription of native RNA templates-itdoesnot
seemtobe sufficient, however, sinceadegraded
enzyme may retain some of it without being
able to transcribeRNA (Fig. 1C and Table 1).
An analogous lack of RNA transcription was
observed with the 84,000-molecular-weight
en-zymefromreticuloendotheliosisvirus (9). The results obtained here with the
unde-graded and degraded murine viral enzyme
preparations strongly resemble those obtained
with the aviana,3enzymecomplex andisolated
a, respectively. It was shownrecently
(Moell-ing, Virology, in press) that transcription of natural RNAcanonlyoccur inthepresenceof
,8. Subunit a wascharacterized as a
degrada-tion product of /3 that doesnotbindto natural
RNA and is unabletotranscribeit. This defect
canbeovercomeby addition ofexcessoligo(dT)
primers. DNA synthesis, then, appears to be
predominantly single stranded asjudgedfrom insensitivity ofthe reaction to actinomycin D.
It hasbeen shown previously thatthe RNase
H of the a subunit behaves like a random
exonuclease (3) that is in agreement with its
bindingdeficiency. The random mode ofaction
of the Moloney murineviral RNase H activity
and its large digestion products described by
Verma (12) may then be a result of enzyme
degradation as well. The
84,000-molecular-weightpolypeptide doesnotreveal these
prop-erties (Fig. 4 and5) but behaves asone would
expect, in analogy to the 18-containing avian enzyme.
Nolonger does the murineviralreverse
tran-scriptase-RNase Hcomplexappear tobe
excep-tional in its enzymatic and structural
proper-ties; instead, itbehaves identically to whatis
considered to be characteristic ofa real viral
reverse transcriptase-RNase H complex, from
experience with the avian enzyme as it
tran-scribes natural RNA in the absence of
syn-thetic primers and the RNase behaves like a
processive exonuclease. Itrather looksasifthe
avian enzyme is exceptional in respect to its
structure, since itnormallycanonly be isolated
asacomplex of the actualreversetranscriptase
molecule,
/3,
accompanied byusuallyequal mo-lar amounts of its degradationproduct, a. The reason for this effect is notunderstood, but it may reflect a tendency of the enzyme to form dimers or oligomers. Appearance of equal mo-lar amounts of a prevents 8 from decaying further.Careshouldbe taken infurther enzyme
char-acterizations not to analyze and describe the properties of a degraded enzyme preparation.
Justonefreezingandthawing ofthe virus can
be sufficienttodestroytheability of the enzyme
totranscribeRNA and canprevent asuccessful
simultaneous direction assay, as was the case
withreticuloendotheliosis virus (9).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.55.246.54.490.2]It seems verylikely that theundegraded re-verse transcriptase-RNase H enzyme complex exists inall RNAtumorviruseswiththeabove mentioned enzymatic properties, possibly con-sisting of a single polypeptide. It will be of interesttoelucidate itsmode of action.
ACKNOWLEDGMENT
Ithank H. Bauer for his support and interest in these studies.
These studies were supported by the Deutsche For-schungsgemeinschaft (SFB 47). s
LITERATURE CITED
1. Abrell, J. W.,and R. C. Gallo. 1973. Purification, char-acterizationand comparison of the DNApolymerase from two primate RNA tumor viruses. J. Virol. 12:431-439.
2. Gerard, G. F., and D. P. Grangenett. 1975. Purification and characterization of the DNA polymerase and RNaseHactivitiesinMoloneymurine sarcoma-leu-kemia virus. J. Virol. 15:785-797.
3. Grandgenett,D.P.,and H. Green. 1974. Different mode of action ofribonucleaseH inpurifiedaand/3 RNA-directed polymerase from AMV. J. Biol. Chem. 249:5148-5152.
4. Kacian,D. L., K.R.Watson, A. Burney,and S. Spie-gelmann. 1971. Purification of the DNApolymerase of avian myeloblastosis virus. Biochim. Biophys. Acta246:365-383.
5. Lapidot,Y., andH.G. Khorana. 1963. Studieson poly-nucleotides. VVIX. Thespecificsynthesis of C'3-C3'-C,'-linked ribonucleotides. J. Am. Chem. Soc. 85:3857-3862.
6. McDonnell, G. P., A.-C.Garapin,W.E.Levinson, N.
Quintrell, L. Fanshier, and J. M. Bishop. 1970. DNA polymerase of RSV: delineation of two reactions with actinomycin. Nature (London) 228:433-435. 7. Moelling, K. 1974. Reverse transcriptase and RNase H:
present in a murine virus and in both subunits of an avian virus.Cold Spring Harbor Symp. Quant. Biol. 39:969-973.
8. Moelling, K. 1974. Characterization of reverse tran-scriptaseand RNase H fromFriend murine leukemia virus.Virology 62:46-59.
9. Moelling, K., H. Gelderblom, G. Pauli, R. Friis, and H. Bauer. 1975. A comparative study on avian reticulo-endotheliosis virus: relationship to murine leukemia virus and viruses of the aviansarcoma-leukosis com-plex. Virology 65:546-557.
10. Schafer, W., andE. Seifert.1968. Production ofa po-tentcomplement-fixing murine leukemia virus anti-serumfrom the rabbit and its reactions with various types of tissue culture cells. Virology 35:323-328. 11. Shapiro, A. L., E. Vinuela, and J. V. Maizel. 1967.
Molecular weight estimation ofpolypeptidechainsby electrophoresis in SDS-polyacrylamide gels. Bio-chem.Biophys. Res. Commun. 28:815-820.
12. Verma, J. 1975. Studies on reverse transcriptase of RNA tumor viruses. III. Properties ofpurified Molo-ney murine leukemia virus DNA polymerase and associated RNase H. J. Virol. 15:843-854.
13. Verma,J. M., N. L. Meuth, H. Fan, and D. Baltimore. 1974. Hamsterleukemia virus: lack of endogenous DNA synthesis of its DNA polymerase. J. Virol. 13:1075-1082.
14. Wang,L.-H.,and P.H.Duesberg.1973.DNA
polymer-aseof murinesarcoma-leukemiavirus:lack of detect-ableRNase H and low activity withavianviral RNA and natural DNAtemplates. J.Virol. 12:1512-1521. 15. Weimann, B. J., J. Schmidt, and D. J. Wolfrun.1974.
RNA-dependent DNA polymerase andribonuclease
HfromFriend virions. FEBS Lett. 43:37-44. on November 10, 2019 by guest
http://jvi.asm.org/
![FIG.1.gelsphocelluloseproteaseavianaration;forstandardH;[3H]glucosamine-labeledMuLVRous(A)(A)loseofviralloseumnwerementmaterialwhich trypsin, (B) Scanning profiles of murine and avian enzyme polypeptides after gel electrophoresis](https://thumb-us.123doks.com/thumbv2/123dok_us/1558781.108485/3.503.264.447.44.452/gelsphocelluloseproteaseavianaration-forstandardh-glucosamine-labeledmulvrous-loseofviralloseumnwerementmaterialwhich-scanning-polypeptides-electrophoresis.webp)
![FIG. 4.processedhybrid.standardunlabeledmolar Competition of hydrolysis of radioactively labeled hybrid by the addition of excess unlabeled Purified reverse transcriptase-RNase H activities were incubated with [3H]poly(rA)-poly(dT) under assay conditions](https://thumb-us.123doks.com/thumbv2/123dok_us/1558781.108485/6.503.110.397.403.598/processedhybrid-standardunlabeledmolar-competition-hydrolysis-radioactively-transcriptase-activities-conditions.webp)

