1
Screening for Exo84 Gain-of-Function Mutants that
Mimic Activation of the Exocyst Complex by Sro7/77
By
Sara Elaine Hudson
Senior Honors Thesis
Department of Biology
University of North Carolina at Chapel Hill
May 1, 2020
Approved:
____________________________________
Dr. Patrick Brennwald, Thesis Advisor
Dr. Elaine Yeh, Reader
2 Abstract
Polarized cells, such as the yeast strain Saccharomyces cerevisiae, manifest their cell polarity through the transport, tethering, and fusion of post-Golgi vesicles to specific regions of the cell
membrane. Tethering is achieved in part by activation of an evolutionarily conserved complex of eight proteins called the exocyst. Recent evidence suggests that the S. cerevisiae exocyst is partially activated through the interaction of lethal giant larvae protein homolog, Sro7/77, and a component of the exocyst, Exo84. However, the function and mechanism of this activation is unknown. Our hypothesis was that gain-of-function Exo84 mutants can mimic constitutive activation by Sro7/77, and thereby suppress deleterious phenotypes caused by disruption of Sro7/77 and Exo84 interactions. To test this hypothesis, a library of randomly mutagenized EXO84 alleles was generated and screened at cold temperatures in yeast that were cold-sensitive due to sro7/77 deletions. Although 15 suppressors were identified, none of the extracted EXO84 alleles were shown to be the cause of suppression, suggesting that the sro7/77 deletion causes too severe a phenotype for a mutated EXO84 allele alone to rescue. Consequently, the screen was performed again at restrictive temperatures in a strain of yeast where exocyst activation by Sro7/77 is indirectly disrupted, through the mutation of an upstream regulator, Sec2. The mutated strain, sec2-41, has been shown to exhibit exocytotic defects and causes temperature sensitivity as a result. From this screen, 17 suppressors were identified and one EXO84 allele was shown to suppress temperature
3 Introduction
Cell polarity, or the spatial and temporal asymmetry of cellular components, is essential for the proper growth and functioning of various cell types including the yeast strain Saccharomyces cerevisiae. A critical part of cell polarity involves the transport, tethering, and fusion of post-Golgi vesicles to specific regions of the cell membrane. The exocyst complex has been shown to assist in tethering these post-Golgi vesicles to the plasma membrane. This complex is composed of eight subunits, including Sec15, Exo70, and Exo84, all of which are conserved evolutionarily between yeast and mammals. Evidence suggests that the mechanistic interactions which lead to polarized trafficking are likely similar between yeast and mammals1. Therefore, studying exocyst function in S. cerevisiae can inform us of cell
polarity establishment in mammalian cells, which is essential for the development of many cell types including neurons. Neurological disorders such as schizophrenia and autism, for example, are shown to be linked with dysfunctional cell polarity establishment2.
Recent evidence suggests that activation of the exocyst is required for tethering, although the function of this activation is unknown. Preliminary data from the Brennwald lab suggests that the yeast homologs of tomosyn, Sro7 and Sro773, and the Rho GTPase, Rho3, may both work to activate the
exocyst through different mechanisms, thereby differentially regulating exocytosis4,5. Sro7 and Sro77 are
paralogs derived from whole genome duplication6 with redundant functions and will be referred to
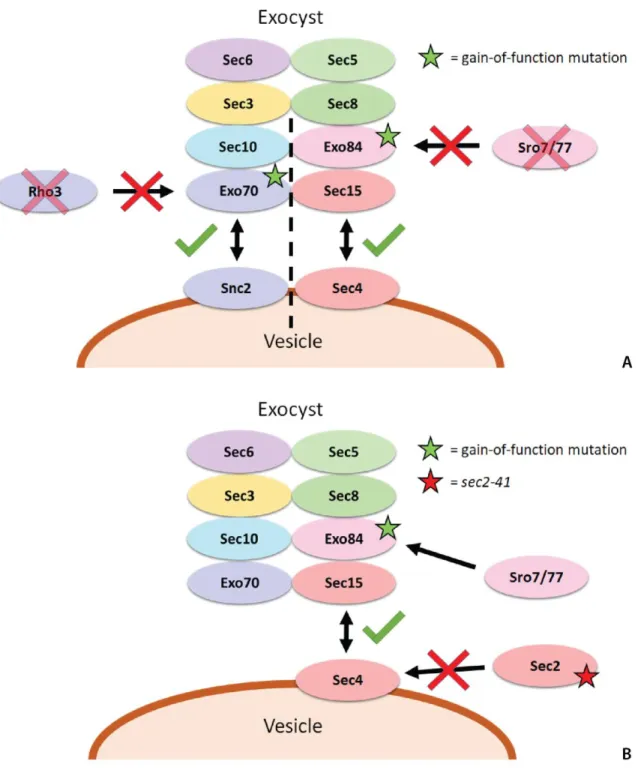
conjointly as Sro7/77. In a recent dominant suppressor screen, gain-of-function Exo70 mutants were shown to suppress cold-sensitive phenotypes due to rho3 deletions in yeast, suggesting that Rho3 activates the exocyst through its interaction with Exo704, and that Exo70 mutants which mimic this
activation bypass the need for Rho3 (Figure 1A). Unpublished data from the Brennwald lab has shown that the same gain-of-function Exo70 mutants do not suppress cold-sensitive phenotypes due to sro7/77 deletions (denoted as sro7/77Δ). Additionally, it has been shown that Sro7/77, but not the gain-of-function Exo70 mutants, increases exocyst binding to the Rab GTPase Sec47, suggesting that Sro7/77
4 specifically through its interaction with Exo84. Our hypothesis is that, in a similar dominant suppressor screen, gain-of-function Exo84 mutants can suppress the cold sensitive phenotype from yeast strains with sro7/77Δ by mimicking constitutive exocyst activation by Sro7/77 (Figure 1A).
Additionally, Sro7/77 has been shown to be a direct effector of Sec48, which is activated by the
guanine exchange factor (GEF), Sec29, and subsequently binds to the exocyst via Sec1510.
Loss-of-function SEC2 alleles, such as sec2-41, have been shown to accumulate post-Golgi vesicles8,11 due to a
lack of Sec4 activation and are temperature sensitive as a result. Unpublished data from the Brennwald lab has shown that overexpressing Sro7 suppresses temperature sensitivity due to this allele. It is possible that high copies of Sro7 activate the exocyst such that the complex can bind better to Sec4, bypassing the need for activation by Sec2, which would indicate that binding of Sec15 to Sec4 is mediated through Sro7/77 activation of the exocyst (Figure 1B). Therefore, we also hypothesize that gain-of-function Exo84 mutants can suppress the temperature sensitive phenotype from yeast strains with sec2-41 by mimicking Sro7/77 activation of the exocyst.
To test these hypotheses, a library of randomly mutagenized EXO84 alleles was generated by error-prone PCR and inserted via gap repair into plasmids containing a yeast selection marker. The plasmids were transformed into yeast that were cold sensitive due to sro7/77Δ or were temperature sensitive due to sec2-41 onto media that selected for the plasmid. The transformed yeast were incubated at restrictive temperatures where the wildtype (WT) copy of EXO84 in the chromosome is insufficient to promote growth. Colonies that grew despite this insufficiency could potentially survive because the mutated EXO84 allele is dominant to WT EXO84 and suppresses the sro7/77Δ or sec2-41 allele.
5 From the sro7/77Δ screens, 15 suppressors out of 10,000 total transformant colonies were
identified that rescued cold sensitivity. However, none of the extracted EXO84 alleles from those colonies repeated the suppression phenotype, suggesting that sro7/77Δ may cause too severe a phenotype for a
Figure 1. Schematic Rationale for EXO84 Genetic Screens. (A) Exocyst interactions in the sro7/77Δ
screen for gain-of-function EXO84 alleles (right) as compared to the rho3Δ screen for gain-of-function
EXO70 alleles4 (left) (B) Exocyst interactions in the sec2-41 screen for gain-of-function EXO84 alleles.
6 mutated EXO84 alone to rescue. From the sec2-41 screen, 17 suppressors out of 2,000 total transformant colonies were identified that rescued temperature sensitivity. One of the extracted EXO84 alleles from those colonies repeated the suppression phenotype via the replacement of a leucine with a tryptophan at amino acid 365, indicating that it is possible to rescue sec2-41 by mutating EXO84, perhaps through the creation of a constitutively activated Exo84 protein.
Methods
Strains and Plasmids
A cold sensitive strain of S. cerevisiae, referred to as BY856, was used for the first pilot and large scale screen and a temperature sensitive strain, referred to as BY31, was used for the second pilot screen. Genotypes and phenotypes of the two strains are outlined in Table 1.
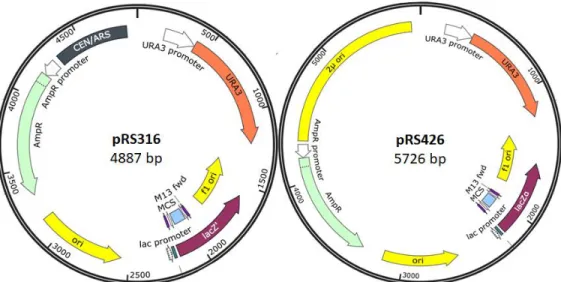
All plasmids used in the screen were designed by the Brennwald lab from the pRS316 or pRS426 vectors, which are capable of being taken up, replicated, and transcribed by both yeast and Escherichia coli. Both plasmids contain URA3 and an ampicillin marker, while pRS316 has a centromere (referred to as CEN) and pRS426 has a 2μ origin of replication, as shown in Figure 2. The ampicillin marker allows for the selection of E. coli DH5α colonies containing the plasmid after transformation. CEN allows for pRS316 to be maintained as a single copy within the yeast cell, replicating alongside the chromosomes. 2μ allows for the pRS426 to be maintained as multiple copies within the yeast cell, facilitating
overexpression of the gene.
Strain BY856 BY31
Genotype ura3-52, his3-Δ200, leu2-3,112,
sro7Δ::LEU2, sro77Δ::HIS3
ura3-52, sec2-41
Required Growth Factors Uracil Uracil
Phenotype Optimal growth at 37 °C on
solid media, 35 °C in liquid culture. Growth deficiency at lower temperatures.
Optimal growth at 25 °C on solid media and in liquid culture. Growth deficiency at higher temperatures.
7 The plasmids based on pRS316 were BB1670, BB2195, and BB1690. BB1670, also referred to as the gap repair plasmid, contained sequences homologous to the yeast chromosome upstream and
downstream of EXO84, including the 5’ and 3’ untranslated regions (UTR) of EXO84. These sequences flanked restriction sites for BamHI and XbaI. By digesting BB1670 with BamHI and XbaI, a linearized plasmid was generated with two sequences of EXO84 homology on either ends. Upon transformation of the digested vector into yeast alongside PCR mutagenized EXO84, the double strand break was repaired by homologous recombination with an EXO84 PCR fragment. This produced an intact plasmid containing mutagenized EXO84, which was subsequently transcribed and translated to produce mutagenized Exo84 proteins. Restriction sites for XhoI and NotI flanked the EXO84 sequence, such that digestion with these enzymes confirmed the presence of EXO84 insert after plasmid extraction. BB2195 contained full length WT SRO7 along with its 5’ and 3’ UTR, which is sufficient to suppress cold sensitivity due to sro7/77Δ. Transforming this plasmid into BY856, consequently, acted as a positive control for suppressing cold sensitivity. BB1690 contained full length WT EXO84 along with its 5’ and 3’ UTR, which is insufficient to suppress cold sensitivity or temperature sensitivity due to sro7/77Δ and sec2-41 respectively, and acted as a negative control for all screens. BB1690 was also used as the source of EXO84 for performing PCR mutagenesis.
8 The plasmid based on pRS426 was BB2311. It contained full length WT SRO7 along with its 5’ and 3’ UTR, which was sufficient to suppress temperature sensitivity due to sec2-41. Transforming this plasmid into BY31 acted as a positive control for suppressing temperature sensitivity.
PCR Mutagenesis and Plasmid Digestion
A forward primer located upstream of the EXO84 coding region and a reverse primer located downstream of the EXO84 coding region were designed and ordered from Eurofins Genomics. WT EXO84 in BB1690 was mutagenized according to the GeneMorph II Kit with 130 ng of template, which produced an approximate mutation frequency of 4-5 mutations per kilobase, using primers at an annealing temperature of 58 °C and extension time of 3 min 20 sec. Two tubes were prepared, one with the DNA polymerase and one without the polymerase, referred to as mut. EXO84 +enzymeand mut. EXO84 – enzyme respectively. BB1670 was digested with BamHI-HF and XbaI restriction enzymes at 37 °C for 2 hours. The PCR mutagenesis products and digestion products (referred to as BB1670 xBX) were stored at -20 °C.
Screen Protocol
BY856 and BY31 were grown in liquid cultures of YPD up to an optical density of 1.0 (λ = 599 nm) at 35 °C and 25 °C respectively. The yeast were pelleted, washed with deionized water and 100 mM lithium acetate, and resuspended in 500 uL of 100 mM lithium acetate. After aliquoting 60 uL of cells into 1.7 mL microcentrifuge tubes, the yeast were pelleted and resuspended in 326 uL of PEG solution. To assist in transformation efficiency, 25 uL of 2 mg/mL single stranded salmon sperm DNA was added to each tube. Then, 5 uL total of plasmid and PCR product was added to each respective tube, as
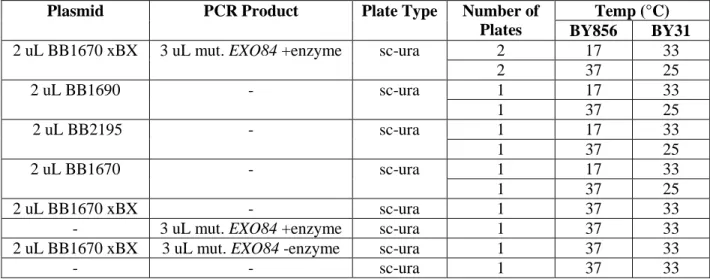
9 Table 2. Pilot Screen with BY856 or BY31 – Combining BB1670 xBX and mut. EXO84+ enzyme produces intact plasmids containing mutagenized EXO84 through homologous recombination. Two plates were placed at 17 °C to screen for potential gain-of-function mutants and at 37 °C to get a count for the total number of transformants. Transforming BB1670 xBX and mut. EXO84 +enzyme alone provided a count for colonies that surfaced due to incomplete digestion and due to BB1690 template in the PCR product, respectively.
Transforming BB1670 xBX and mut EXO84 –enzyme together controlled for transformants due to homologous
recombination between BB1690 template in the PCR product with digested BB1670.
Table 3. Large-Scale Screen with BY856 – Six transformation tubes in total were made of BB1670 xBX plus mut. EXO84 +enzyme, giving 3.6 mL total of cells resuspended in deionized water. 24 plates with 150 uL per plate were prepared, 20 of which were placed at 17 °C to screen for potential gain-of-function EXO84 plasmids, with the other 4 placed at 37 °C to get a count for the total number of transformants.
During the large scale screen with BY856, the 4 plates containing digested gap repair vector and mutagenized EXO84 at 37 °C were incubated at that temperature for 2 days. Afterward, each plate was replica plated first onto a new YPD plate then onto a new sc-ura plate and subsequently placed at 17 °C.
Plasmid PCR Product Plate Type Number of
Plates
Temp (°C)
BY856 BY31
2 uL BB1670 xBX 3 uL mut. EXO84 +enzyme sc-ura 2 17 33
2 37 25
2 uL BB1690 - sc-ura 1 17 33
1 37 25
2 uL BB2195 - sc-ura 1 17 33
1 37 25
2 uL BB1670 - sc-ura 1 17 33
1 37 25
2 uL BB1670 xBX - sc-ura 1 37 33
- 3 uL mut. EXO84 +enzyme sc-ura 1 37 33
2 uL BB1670 xBX 3 uL mut. EXO84 -enzyme sc-ura 1 37 33
- - sc-ura 1 37 33
Plasmid PCR Product Plate Type Number of
Plates
Temp (°C)
2 uL BB1670 xBX 3 uL mut. EXO84 +enzyme sc-ura 20 17
4 37
2 uL BB2195 - sc-ura 1 17
1 37
2 uL BB1670 - sc-ura 1 17
1 37
- 3 uL mut EXO84 +enzyme sc-ura 1 37
2 uL BB1670 xBX 3 uL mut EXO84 - enzyme sc-ura 1 37
10 Plasmid Extraction
Colonies that were observed to grow above background at 17 °C for BY856 or 33 °C for BY31 (referred to as suppressors) were picked and grown in 5 mL of sc-ura liquid culture at 35 °C for BY856 or 25 °C for BY31. After overnight growth, 1.5 mL of each culture was pelleted, washed with 1 mL of deionized water, and pelleted again in 1.7 mL microcentrifuge tubes. The yeast were resuspended in 200uL of TESS Buffer. After adding glass beads to each tube, the yeast were vortexed for 1 minute to begin lysis. 200 uL of 25:24:1 phenol:chloroform:isoamyl alcohol (PCI) was added, and the tubes were vortexed for another minute. After a 5 min spin at 13,000 rpm, the aqueous layer was transferred to a new tube, and 400 uL more of PCI was added before a final 1 minute vortex. After another 5 min spin at 13,000 rpm, the aqueous layers were transferred to Wizard columns for purification. The candidate plasmids were eventually eluted in 50 uL of high-EDTA TE Buffer.
After completion of the yeast DNA extraction, 7 uL of each candidate plasmid was transformed into 100 uL of E. coli DH5α. The transformed cells were incubated on ice for 30 minutes, heat shocked in a 42 °C water bath for 45 seconds, and placed back on ice for 5 minutes. 400 uL of SOC was added and the cells were allowed to recover at 37 °C in a 260 rpm shaker for 1 hour. 250 uL of cells were spread onto LB-Amp plates and incubated at 37 °C overnight. Representative colonies were picked the next day and inoculated in 5 mL of TB with approximately 100 ug/mL of ampicillin and incubated at 37 °C overnight. The cultures were subsequently mini-prepped according to Promega Wizard Plasmid Miniprep kit and eluted in 75 uL of low-EDTA TE buffer.
To confirm the presence of an EXO84 insert, digestion with XhoI and NotI was performed with 4 uL of recovered plasmid for 1 hour at 37 °C. The digestion products were then run on a 0.7% agarose gel against either 1 kb or 1 kb + GeneRuler ladder and empty BB1670 vector. The presence of an
11 Confirmation Transformation
To determine a causal link between the mutated EXO84 plasmids and suppression, the candidate plasmids were transformed back into either BY856 or BY31. 2 uL of each candidate plasmid was transformed according to the procedure described previously under Screen Protocol. After resuspension in 1 mL of deionized water, 100 uL of cells were plated on sc-ura. All transformations were recovered at the permissive temperature, BY856 at 37 °C and BY31 at 25 °C, in order to allow growth of all
transformants. After 2-3 days of growth, six representative colonies were picked from each
transformation plate, diluted in 200 uL of deionized water in a 96 well plate, and spotted onto YPD and sc-ura agar plates. The spot plates were incubated at a variety of temperatures to analyze transformed yeast growth between the permissive temperature and the restrictive temperature.
Generating and Testing Point Mutations
Site-directed mutagenesis was performed with BB1690 as template to generate plasmids capable of expressing EXO84 genes containing I28 or W365 point mutations. The PCR tubes were then digested with 2 uL DpnI for 2 hours to destroy template BB1690, and subsequently transformed into E. coli DH5α and mini-prepped according to the procedure previously described in Plasmid Extraction. Point
mutations were confirmed via sequencing. 2 uL of each variant plasmid was transformed into BY31 as described under Screen Protocol and plated as described under Confirmation Transformation.
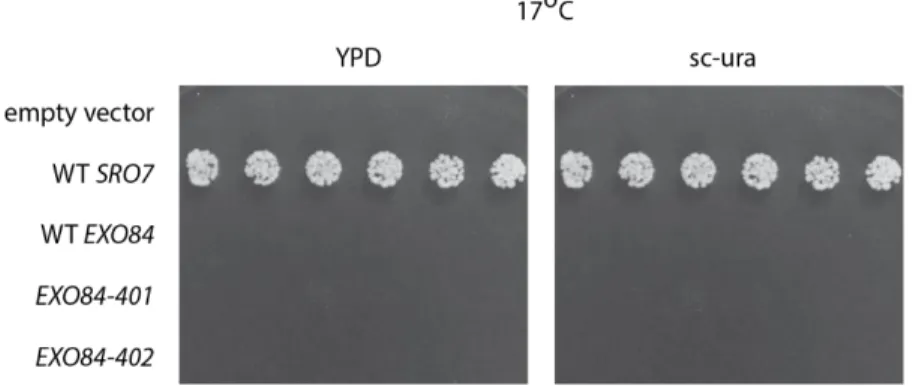
Results
12 the yeast rescues growth at 17 °C, indicating that neither the EXO84-401 or EXO84-402 allele could suppress sro7/77Δ at that temperature.
From the large scale screen with BY856, 13 suppressor colonies were discovered out of 9,000 total transformants. EXO84-403 through EXO84-407 were identified from the replica plates 4 days after they were placed at 17 °C. EXO84-408 through EXO84-415 were discovered from the 20 plates placed directly at 17 °C, ranging from 7 to 14 days after initial transformation.
After plasmid extraction, confirmation digestion (see Appendix), and subsequent transformation back into BY856, growth of EXO84-403 through EXO84-415 transformants was compared against WT SRO7, WT EXO84, and empty vector at 25 °C or 30 °C in addition to the restrictive temperature of 17 °C (Figure 4), to determine if any alleles exhibited suppression distinguishable from WT EXO84 at less restrictive temperatures. All transformants grew at the permissive temperature of 37 °C. However, across all confirmation transformations, only WT SRO7 exhibited normal growth at 30, 25, and 17 °C. The deficient growth shown by WT EXO84 at 25 °C was also seen for EXO84-403 through EXO84-409 on both YPD and sc-ura, with neither WT EXO84 nor EXO84-403 through EXO84-409 showing any growth at 17 °C (Fig. 4A). The final five suppressors, EXO84-410 through EXO84-415, were tested at a more permissive temperature, 30 °C, instead of 25 °C, to identify any weak suppression capabilities. However, even at 30 °C, EXO84-410 through EXO84-415 exhibited a growth defect identical to WT EXO84 on YPD, and a slight defect on sc-ura (Fig. 4B). Therefore, none of the 13 extracted EXO84 alleles were capable of suppressing sro7/77Δ at 30, 25, or 17 °C above WT EXO84.
13 From the pilot screen with BY31, 17 total suppressor colonies were identified out of 2,000 total transformants, 2 to 3 days after direct selection at 33 °C. After plasmid extraction, confirmation digestion, and transformation back into BY31, growth of EXO84-501 through EXO84-517 transformants was compared against WT EXO84 and WT SRO7 2μ (Figure 5). The transformants were grown at 30 °C and 31.5 °C in addition to 33 °C, in order to test a range of suppression levels. Except for EXO84-514, all mutant EXO84 transformants failed to grow at 33 °C and showed defective growth at 31.5 °C identical to WT EXO84. EXO84-514, however, did not exhibit any growth defects at 31.5 °C after 2 days of
incubation (Fig. 5C). EXO84-514 also showed normal growth after 3 days at 33 °C when compared to the Figure 4. Retesting
EXO84403 through -415 from the large
14 Figure 5. Retesting EXO84-501 through EXO84-517 from the pilot screen in BY31. Six replicates were tested for each plate, but only three are shown. (A) All plates shown 3 days after growth. (B) All plates shown 2 days after growth. (C) All plates shown 2 days after growth. (D) All plates shown 3 days after growth. A and B indicate multiple replicates of plasmid EXO84-514.
15 permissive temperature of 25 °C (Fig. 5D). This growth was seen for two replicates of EXO84-514, further confirming that suppression is due to the EXO84-514 plasmid. EXO84-514 also exhibited better growth than WT SRO7 2μ at 31.5 °C and 33 °C, indicating that EXO84-514 is a better suppressor of sec2-41 than high copy SRO7. WT SRO7 2μ showed minimal suppression at 33 °C after 3 days (Fig. 5A, 5D) and weak suppression after 2 days (Fig. 5B, 5C), compared to EXO84-514.
After plasmid extraction and sequencing, EXO84-514 was shown to have two non-synonymous substitutions resulting in the replacement of threonine with isoleucine at amino acid 28 (T28I) and the replacement of leucine with tryptophan at amino acid 365 (L365W). To determine which of these two substitutions was required for the sec2-41 suppression phenotype, site-directed mutagenesis was performed to create EXO84-T28I and EXO84-L365W. The plasmids were transformed back into BY31 and compared against WT EXO84, WT SRO7 2μ, and EXO84-514 at 25, 30, 31.5, and 33 °C (Figure 6). EXO84-L365W shows identical suppression to EXO84-514 at all temperatures, with normal growth at 31.5 °C and slightly slower growth at 33 °C. EXO84-T28I shows identical growth to WT EXO84 at all temperatures, with minimal growth at 31.5 °C and no growth at 33 °C.
Discussion
It has been shown that Sro7/77 activation of the exocyst is necessary for proper exocytosis, but the mechanism of activation remains elusive. The exocyst consists of two tetrameric modules, Sub-complex I, which contains Sec3, Sec5, Sec6, and Sec8, and Sub-Sub-complex II, which contains Sec10, Sec15, Exo70, and Exo8412,13. Sro7/77 has been shown to increase binding of Sec15 to Sec47 and can
16 suppress sec2-41 presumably by activating the exocyst and bypassing the need for Sec2 activation of Sec4. Exo84 has been shown to interact with Sro7/77 in vivo and in vitro5, and thus is a compelling target
for studying the effects of Sro7/77 activation. Identifying gain-of-function Exo84 mutants that mimic Sro7/77 exocyst activation can provide insight into the biochemical and structural properties of an Sro7/77 activated exocyst complex.
We screened for gain-of-function Exo84 mutants from sec2-41 and sro7/77Δ at their respective restrictive temperatures. Out of 2,000 sec2-41 transformants and 10,000 sro7/77Δ transformants, one EXO84 allele, EXO84-514, was identified that could suppress sec2-41 and no EXO84 alleles were isolated that could suppress sro7/77Δ. EXO84-514 contained two non-synonymous mutations, resulting in T28I and L365W. Upon testing the mutations separately, EXO84-L365W showed identical growth to EXO84-514 and EXO84-T28I showed identical growth to WT EXO84, which strongly suggests that L365W is the driver mutation responsible for the suppression of sec2-41, while T28I is a passenger mutation that does not alter the sec2-41 phenotype.
Position 365 is located in the PH domain of Exo84, which is suggested to act as a dock between CorEx II and the CAT-B and CAT-C regions of Sec1012. CorEx II is a structure of four alpha helical
bundles from Sec10, Sec15, Exo70, and Exo84. This structure consists of two dimeric coiled coils, Sec10-Sec15 and Exo70-Exo84, at the core of the exocyst complex, and is likely important in maintaining tight assembly of the four proteins. It is possible that substituting leucine with tryptophan at amino acid 365 adds extra bulk to the PH domain and loosens the hold between Sec10, Exo84, and the rest of CorEx II, thereby increasing protein mobility. This loosening could confer an allosteric change in Sec15 that confers activation, allowing Sec15 to bind more strongly to Sec4. Activation through relaxation of the exocyst subunits is not unprecedented. EXO70-I114F, which suppresses exocytotic defects associated with rho3Δ and cdc42-6, exhibits increased mobility of the Exo70 protein, creating a “hole” next to Sec6 and thereby opening up the binding site for the v-SNARE Snc1/27. The exocyst structural changes caused
17 microscopy of purified EXO84-L365W mutant exocyst complexes, as was done with EXO70-I114F in the gain-of-function Exo70 screen in rho3Δ.
In order to parse the mechanism of suppression, it would be valuable to test the ability of EXO84-L365W to suppress other exocytotic defects. Testing whether EXO84-L365W rescues rho3Δ and cdc42-6 could indicate whether this mutation is specific to activating the exocyst via Sro7 and Exo84 interactions, or if the mutation broadly activates the exocyst complex to overcome defects in Rho3 and Cdc42
activation as well. Yeast strains with sec10-2 have been shown to disrupt Sec10 and Sec15 interactions10,
and are partially suppressed by high copy Sro75. Therefore, sec10-2 could be tested to determine whether
EXO84-L365W impacts Sec10 function and provide evidence for the conformational change of Sec10 CorEx II interactions. Additionally, sec15-1 is a late-acting SEC mutant also shown to be suppressed by high copy Sro75. Testing whether EXO84-L365W suppresses sec15-1 can elucidate how Sro7 exocyst
activation impacts Sec15 function.
High copy Sro7 can suppress sec2-41 presumably by activating the exocyst via Exo84 and bypassing the need for Sec2 activation of Sec4. We demonstrated that EXO84-L365W suppresses sec2-41 better than high copy Sro7, which may be indicative that suppression by L365W does not entirely
correlate with Sro7/77 activation of the exocyst. Consequently, it would be worthwhile to continue refining the sro7/77Δ screen in order to generate Exo84 mutants specific for Sro7/77 activation. However, the sro7/77Δ screen produced only one suppressor colony per 660 transformant colonies compared to the sec2-41 screen which produced one suppressor per 118 transformants. The low rate of suppressor isolation with sro7/77Δ is likely due to the mutation having a much higher selection pressure than sec2-41. Additionally, it is possible that sro7/77Δ causes too severe a phenotype for a mutated EXO84 alone to rescue growth at the highly restrictive temperature of 17 °C. This hypothesis is supported by evidence suggesting other functions for Sro7/77 aside from exocyst regulation, including maintenance of cell proliferation and colony development through the Rho1/Tor1 pathway14. Consequently, the 15
18 To ease the extreme selection pressures associated with sro7/77Δ, the screen could be redone at less restrictive temperatures, such as 25 °C, in which sro7/77Δ strains exhibit suboptimal growth
characterized by smaller colonies and longer growth periods seen in empty vector transformants. Potential gain-of-function EXO84 plasmids could be identified and isolated from colonies that exhibit normal growth similar in morphology and timing to sro7/77Δ colonies transformed with WT SRO7. However, this adjustment could result in more false positives, since non-suppressor colonies are capable of eventual growth.
Despite not having found EXO84 suppressoralleles in the sro7/77Δ screen, fifteen strains of yeast capable of dominantly suppressing the cold sensitive phenotype were identified. Thus, it would be
19 Appendix
20 References
1. Brennwald, P., & Rossi, G. (2007). Spatial regulation of exocytosis and cell polarity: yeast as a model for animal cells. FEBS letters, 581(11), 2119–2124. doi:10.1016/j.febslet.2007.03.043 2. Hakanen, J., Ruiz-Reig, N., & Tissir, F. (2019). Linking Cell Polarity to Cortical Development
and Malformations. Frontiers in cellular neuroscience, 13, 244. doi:10.3389/fncel.2019.00244 3. Lehman, K., Rossi, G., Ardarno, J.E., Brennwald, P. (1999). Yeast Homologues of Tomosyn and
lethal giant larvae Function in Exocytosis and Are Associated with the Plasma Membrane Snare, Sec9. J Cell Bio. 146(1), 125-140. doi: 10.1083/jcb.146.1.125
4. Wu, H., Turner, C., Gardner, J., Temple, B., Brennwald, P. (2009). The Exo70 Subunit of the Exocyst Is an Effector for Both Cdc42 and Rho3 Function in Polarized Exocytosis. Molecular Biology of the Cell, 21(3), 430-442. doi:10.1091/mbc.E09-06-0501
5. Zhang, X., Wang, P., Gangar, A., Zhang, J., Brennwald, P., TerBush, D., Guo, W. (2005). Lethal giant larvae proteins interact with the exocyst complex and are involved in polarized exocytosis. J. Cell Biol. 170(2), 273-283. doi:10.1083/jcb.200502055
6. Byrne, K. P., & Wolfe, K. H. (2005). The Yeast Gene Order Browser: combining curated homology and syntenic context reveals gene fate in polyploid species. Genome research, 15(10), 1456–1461. doi:10.1101/gr.3672305
7. Rossi, G., Lepore, D., Kenner, L., Czuchra, A., Plooster, M., Frost, A., Munson, M., Brennwald, P. (2020) Exocyst structural changes associated with activation of tethering downstream of Rho/Cdc42 GTPases. J. Cell Biol. 219(2). doi:10.1083/jcb.201904161
8. Grosshans, B.L., Andreeva, A., Gangar, A., Niessen, S., Yates, J.R. 3rd, Brennwald, P., Novick, P.
(2006). The yeast lgl family member Sro7p is an effector of the secretory Rab GTPase Sec4p. J. Cell Biol. 172, 55–66. doi:10.1083/jcb.200510016
21 10. Guo, W., Roth, D., Walch-Solimena, C., Novick, P. (1999). The exocyst is an effector for Sec4p,
targeting secretory vesicles to sites of exocytosis. EMBO J. 18(4), 1071-80. doi:10.1093/emboj/18.4.1071
11. Nair, J., Mueller, H., Peterson, M., Novick, P. (1990). Sec2 protein contains a coiled-coil domain essential for vesicular transport and a dispensable carboxy terminal domain. J. Cell Biol. 110(6), 1897-1909. doi: 10.1083/jcb.110.6.1897
12. Mei, K., Li, Y., Wang, S., Shao, G., Guo, W., et al (2018). Cryo-EM Structure of the Exocyst Complex. Nat Struct Mol Biol. 25(2), 139-146. doi:10.1038/s41594-017-0016-2
13. Lepore, D. M., Martínez-Núñez, L., & Munson, M. (2018). Exposing the Elusive Exocyst Structure. Trends in biochemical sciences, 43(9), 714–725. doi:10.1016/j.tibs.2018.06.012 14. Liou, L. C., Ren, Q., Gao, Q., & Zhang, Z. (2014). Sro7 and Sro77, the yeast homologues of the