0022-538X/83/050661-06$02.00/0
Copyright © 1983, American SocietyforMicrobiology
The
Major
Herpes
Simplex Virus DNA-Binding
Protein Holds
Single-Stranded
DNA
in
an
Extended
Configuration
WILLIAM T.RUYECHAN
DepartmentofBiochemistry, Uniformed Services UniversityoftheHealth Sciences, Bethesda, Maryland 20814
Received 2 December 1982/Accepted 22 February 1983
Properties of the major DNA-binding protein found in herpes simplex
virus-infected cells were investigated by using a filter binding assay and electron
microscopy. Filter binding indicated that the stoichiometry of binding of the
protein with single-stranded DNA is approximately 40 nucleotides per protein
moleculeatsaturation. Strongclustering of the protein in DNA-protein
complex-es, indicative ofcooperative binding, was seen with the electron microscope.
Measurements ofsingle-stranded fd DNA molecules saturated with protein and
spread for electron microscopy by using both the aqueous and formamide
spreadingtechniques indicated thatthe DNAis held inanextendedconfiguration
witha base spacing of -0.13nmperbase.
Protein synthesis in herpes simplex virustype
1(HSV-1)-infected cells is coordinately
regulat-ed and sequentially ordered. Three groups of
polypeptides have been designated in order of
their temporalappearance. They include the a
group,which doesnotrequire prior viral protein
synthesis for expression, the a group, which
includes proteins involved in viralDNA
metabo-lism, and thefygroup,which is madeup
primari-ly of structuralproteins found intheintact virion
(11). The 1 polypeptidesaresynthesized during
the peak period of viral DNA synthesis and include the viral DNA polymerase and the major herpesvirus DNA-binding protein, which has been designated infected cell polypeptide 8
(ICP8) basedonitselectrophoreticmobility
rela-tivetootherproteinsfound in infected cells (12).
Estimates of the molecular weight of ICP8
have ranged between 128,000 and 135,000.
DNA-cellulosechromatography ofradioactively
labeled infected-cell extracts has shown that
ICP8binds moretightly to single-stranded than
todouble-stranded DNA(4,15). Recent
experi-mentsindicate that thisproteinmaybea
group-specific antigenforherpesviruses,since antisera against ICP8 crossreact with analogous
polypep-tides found in other herpesvirus systems (20).
Finally, ICP8 has been detected in
HSV-trans-formed cells (10),andantibodytoICP8 has been
found in seraobtained from cervical carcinoma
patients (3). This report shows that this
impor-tant herpesvirus-coded protein holds
single-stranded DNAinanextendedconformation.
3H-labeled
HSV-1 DNA was the gift ofMi-chael Bartkoski, as were initial stocks ofVero
cells and HSV-1 strain F (9). Bacteriophage fd
DNA was the gift of Lucy M. S. Chang. Calf
thymus DNA andpancreaticDNase andRNase
were purchased from Sigma Chemical Co. (St.
Louis, Mo.). Cellulosepowderand
DEAE-cellu-losepowderwerepurchased from Bio-Rad
Lab-oratories (Richmond, Calif.). Phosphocellulose
powder (Pl1) was purchased from Whatman,
Inc. (Clifton, N.J.). Preparation ofICP8wasby
theprocedure of Powelletal. (14). Atotalof15
confluent 2.5-liter roller bottles of Vero cells
were infected with HSV-1 strain F at an input
multiplicity of 20 PFU per cell. The cellswere
harvestedat18hpostinfectionandsuspended in
buffercontaining 20mMTris-hydrochloride(pH
7.5) and 0.5mM dithiothreitolata cell
concen-tration of -3 x 107/ml.Aftersonication,DNase
and RNasewereaddedtofinalconcentrations of
20 and 2 ,ug/ml, respectively. After a 30-min
incubation at 4°C, solid NaCl was added to a
concentration of 2.0 M, andtheresulting
precip-itate was removedby low-speedcentrifugation.
Purification of ICP8through chromatographyon
DEAE-cellulose and phosphocellulose was
car-ried out exactly as described by Powell et al.
(14). Each fraction from these columns was
assayed for absorbance at 280 nm andanalyzed
by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and subsequent silver staining
(19). Fractions from the phosphocellulose
col-umncontaining ICP8wereadjustedto500,ug/ml
in bovine serumalbumin, dialyzed against
low-salt buffer (50mM KCl, 20 mM
Tris-hydrochlo-ride [pH 7.5], 0.5 mM dithiothreitol, and 20%
glycerol) andapplied to a30-ml single-stranded
DNA-cellulose column (2) which had been
prewashedwithbuffercontaining bovine serum
661
on November 10, 2019 by guest
http://jvi.asm.org/
662 NOTES
a
b
4 92
-466
445
431
FIG. 1. A 9o sodium dodecyl sulfate-polyacryl-amidegel of purified ICP8 peak fractions (a and b) from a single-stranded DNA-cellulose column. The protein was visualized by meansofa silverstaining procedure(10). Molecularweight markers,in descend-ingorder, werephosphorylase B,bovineserum
albu-min, ovalbualbu-min,andcarbonicanhydrase,
albumin. The column was eluted stepwise with
0.05, 0.1, 0.2, 0.4, and 1.0 M KCl. ICP8 was
obtained (-50,ug/109 cells) inthe0.4and1.0 M
fractions,dependingonthebatch of
DNA-cellu-lose. Theproteinwasthendialyzed against0.01
M Tris-hydrochoride (pH 7.5)-0.001 M EDTA (TEbuffer) andquantitated by using a Bio-Rad
proteinassaykit.
ICP8-fd DNA complexes were prepared for
electron microscopy by
mixing
thetwocompo-nents atavarietyofweightratios.Sampleswere
brought to 80 ,ul with TE buffer, against which
the protein and DNA were also dialyzed. The
DNA waspresent at aconcentrationof 1 ,ug/ml
in all samples. ICP8 was present at
concentra-tions of2, 5, and 10,g/ml. The 80-,u mixtures
were incubated at4°Cfor 10minandthen fixed
with 8% glutaraldehyde (1.25
RI/80
IlI)
at roomtemperatureforan additional10min. The
com-plexes were then spread for electron microscopy by either the aqueous ammonium acetate tech-nique or the formamide techtech-nique (7, 17).
Am-moniumacetate was presentin both the
hyper-phase and the hypophase at a concentration of
100 mMintheaqueoustechnique. The
complex-es wereobserved and photographed at
magnifi-cations of9,800x and 16,000x, using a Zeiss
EM 10A high-resolution transmission electron
microscope.
A solution of sonicated 3H-labeled HSV-1
DNA(80,000dpm/,ug)washeatedto100°Cfor5
minand icequenced to obtain a stocksolutionof
single-strandedDNAfor filterbindingassays.A
0.02-pg portion of the DNA was mixed with
increasing amounts of ICP8. The samples were
broughtto100 pI withTEbuffer and incubated
at 4°C for 15 min. After incubation, an 80-pA
aliquotwaswithdrawn and adsorbedtoan
0.45-,um-pore-size nitrocellulose filter witha Hoefer
multichannel filtration manifold. The filtration
speed was 4 ml/min. The filters were then
washed with 5 ml each of cold TE buffer and
90% ethanol,airdried,and counted.
Sodiumdodecyl sulfate-gel electrophoresisof
purified ICP8 and subsequent silver staining
indicated the presence ofa single polypeptide
species with a molecular weight of -128,000
(Fig. 1). Filter binding assays carried out as
^- .015-/
0 4 .010' .
z
.005-.04 .08 .12 .16 .20 .24 .28 .3 2
[image:2.490.121.167.78.248.2]IcP8 pg
FIG. 2. Typicalresults fromafilterbinding assayindicating saturation (maximum retention) ofsonicated,
heat-denatured, 'H-labeledHSV-1 DNA with ICP8ata10:1protein-to-DNA weightratio. The HSV-1 DNA had
anaveragelengthof--400nucleotidesasdeterminedbyelectronmicroscopy(datanotshown).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.490.111.400.469.642.2]b
'4*t{'
;*
'
'
v
'
05
Mm
;. >a¢#to
-0
H
-.
.
~~~~~.
;,
C.,~~~~~~V
FI. 3. Eletro
mirorpho
.ICPdcmlxs.prpae ata,\prti/N wegh rai fv: hnonanomditrbuio ofprtin is evdne byth prsec of bot extended, ful sauae cice an
*~ v
*
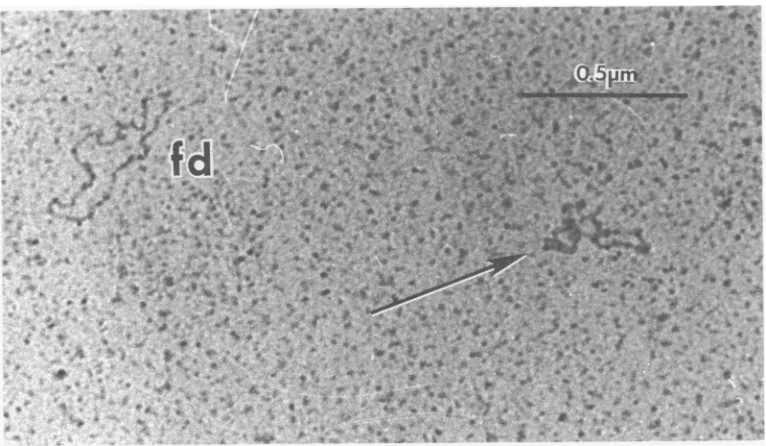
itw@-FIG. 3. Electron micrograph ofICP8-fd complexes prepared at a protein/DNA weight ratio of5:1. The nonrandom distribution of protein is evidenced by the presence of both extended, fully saturated circles and
highly collapsed, irregularly shapedmolecules (arrows) with littleor no proteinbound. Thecomplexeswere
stainedwithuranylacetate(12). Glutaraldehydefixationwasrequiredtomaintain theintegrityof thecomplexes.
described above indicated that saturation of
sonicated,single-strandedHSV-1 DNAby ICP8
occurs ata protein/nucleic acid weight ratio of
10:1 (Fig. 2). This value is in good agreement
with results obtained by Powelletal.(14) which
indicate that ICP8/nucleic acid weight ratios of
10:1aresufficienttoallow thecomplete
denatur-ation ofpolydeoxyadenylic
acid-polydeoxythy-midilic acidduplexes under thermal and solution
conditions where the polynucleotides form a
stable doublehelix. The10:1 ratio indicates that
one molecule of ICP8covers approximately 40
bases. Thisstoichiometry is similartothat of the
Escherichia coli single-strand binding protein
(SSB), whichcoversapproximately 32 basesper
80,000-molecular-weighttetramer(18).
Observation of glutaraldehyde-fixed,
aque-ous-spread ICP8-fd DNA complexes at
subsa-turating levels of protein showsaclearly
nonran-dom distribution of theprotein (Fig. 3). Some of
the DNA molecules appear extended and fully
saturated with protein, whereas others are
col-lapsed irregularstructures typical of naked
sin-gle-strandedDNA under theaqueousmounting
conditions. The frequencyofopen circular and
collapsed structures at several
protein/DNA
weight ratioswasdeterminedby scoringa
mini-mum of 110 structures seen with the electron
microscopeateach ratio. Thefractionof
extend-ed open circular structures increased with
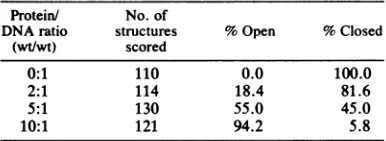
in-creasing amounts of ICP8 (Table 1). The
ob-served increase correlates quite well with the
stoichiometry established in the filter binding
assay andsuggests that theopenstructures are
fully saturated fdDNA-ICP8 complexes
where-asthecollapsedstructures arenaked fd DNA.A
similarclusteringofprotein molecules has been
[image:3.490.50.450.74.361.2]observed withT4gene32proteinandSSB-DNA
TABLE 1. Frequency of open circularstructures at variousprotein/DNA ratios
Protein/ No.of
DNA ratio structures %Open %Closed (wt/wt) scored
0:1 110 0.0 100.0
2:1 114 18.4 81.6
5:1 130 55.0 45.0
10:1 121 94.2 5.8
VOL.46, 1983 NOTES 663
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.490.255.448.601.672.2]664 NOTES
complexes and isindicative ofcoperative
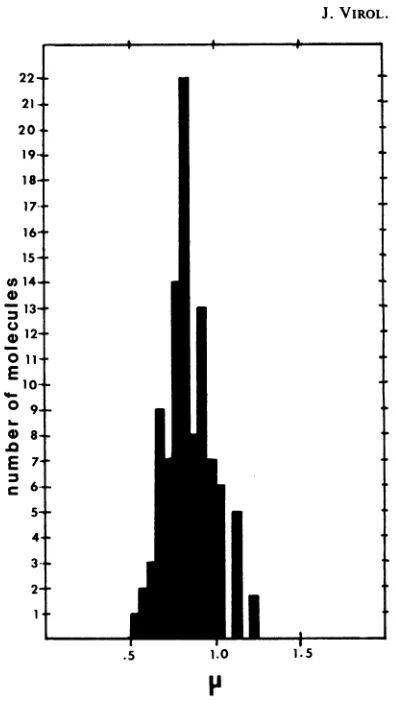
bind-ing(1, 17). Measurementsof 102 extended
ICP8-fd DNA complexes prepared at aprotein/DNA
ratio of10:1yieldedanaverage contour lengthof
0.85 ±0.11 ,um(Fig. 4). This length corresponds
to anucleotide spacing of0.132± 0.017 nm per
nucleotide basedon avalue of6,408 nucleotides
perfd molecule (5). This value is similarto the
base spacing of 0.18nmperbaseseeninSSB-fd
DNAcomplexes.
It is possible that the very close DNA base
spacingof0.132 nmmeasured for the complexes
wasdue to a combinationoftheaction ofICP8
and the aqueous mounting procedure used.
First, since the single-stranded DNA is
visual-izedas ahighly collapsedstructure,itis possible
thatportions of the putative saturated
complex-esdonothaveprotein boundtothem.
Converse-ly, it is possible that a least some ofthe
col-lapsed structures are partially saturated
complexes. Although the results presented in
Table 1 and the tightness of the distribution of
contour lengths in Fig. 4 argue against these
possibilities, they are nonetheless a source of
concern. Second, assuming that allof the open
circularstructures arefully saturated, the ionic
spreading medium used in the aqueous
tech-nique could alter the overall conformation ofthe
complex.
Toexamine thesepossibilities, ICP8-fdDNA
complexes werespread by usingtheformamide
technique, which allows single-strandedDNA to
bevisualizedas anextended smooth filament (7,
17). Procaryotic DNA-binding protein-DNA
complexes have beensuccessfully visualized as
thicker, denser filaments as comparedwith
na-kedDNA.Thus, ifpartially saturated complexes
were present in ICP8-fd DNA mixtures, the
formamidetechniquewould allowdifferentiation
ofprotein-boundandprotein-free regions. Inthe
second case, i.e., if the extended structures
werefully saturated, measurementsof their
con-tourlengths would determinewhetherthe
appar-ent geometry of the complexes was altered by
the spreading medium. fd DNAand ICP8were
mixed in80-,ul aliquots toyieldfinal
concentra-tions of 1 ,gofDNA and -7 ,ugofprotein per
ml. Incubation and fixation of the complexes
werecarriedout asdescribed above.After
fixa-tion,an additional0.1 ,gof fdDNA was added
to ensure thatsomenakedDNA waspresent. A
30-,ul portion ofthis mixture was then spread
from a 40% formamide hyperphase containing
cytochromec(0.3,g/ml)onto a15% formamide
hypophase. The grids were stained with uranyl
acetate, shadowed with gold/palladium, and
photographed at amagnification of16,000x.
Anexamination ofthegridsshowed that only
two types of structure were present:
well-ex-tended naked fd DNAcirclesand much smaller
[image:4.490.253.451.57.409.2].5 1.0 1.5
FIG. 4. Histogram of 102 well-extended ICP8-fd complexes prepared at aprotein/DNA ratio of 10:1. Photographs were takenat amagnificationof9,800x, using a Zeiss EM 10A high-resolution instrument. Negatives were projected onto a blackboard with a
lanternslideprojector, andcontour measurementsof
the molecules were taken with a Keuffel and Esser mapmeasuring device.
circularDNA-protein complexeswith
character-istically thicker, rougher-appearing contours
(Fig. 5). No obviously partial complexes were
seen. Contour measurements of 87 complexes
yieldedanaveragevalueof0.79 + 0.08 ,um.This
contourlengthcorrespondsto abasespacingof
0.125 + 0.012 nm per base. The naked fd in
contrasthad abasespacingof0.27nmperbase,
ingoodagreementwiththepreviously published
value of 0.28 nm per base with the formamide
technique (17).
The contour length and base spacing values
obtained by the two spreading techniques are
the same within experimental error, indicating
that (i) partial saturation of the fd DNA with
ICP8doesnot occurunder theconditionsused,
thus implying a high degree ofcooperativity in
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.46,1983 NOTES 66)
b
~
~
t~~ot7
~
~
~
~~~ *
j
a~~~~~~~~~ ~~~~~~ *..~~~~~~~~~~~~~y4' ~~~~~~~~~~~K~
~44*
4A~ ~ ~ ~ ~ '*
940~~ ~ ~ ~ ~ ~ ~ 4 VA, r
,Av.
14D~~~ ~ ~ ~ ~ ~~a '..$
P
FIG.5Anelctronicrogaphshwing a ICP8 dDNAcomple (arro) and nextndedpotein-ree f
DNAmolcuemuned y uin th frmaidetehniueandshdowdte-hicerwthgol/pllaiumNoe
"grainy"appearance Andrdcdcnorlnt ftecmlx
the binding reaction, and (ii) the tight base
spacing is imposedonthe DNAby ICP8, and the
complexesarestable underwidelyvarying
solu-tionconditions. These results imply that
single-stranded DNA in the aqueous environment of
the infected-cell nucleus can be heldin an
ex-tendedconfiguration by ICP8 witha
characteris-tic base spacing of -0.13nmperbase.
The results described above show that the
interaction of ICP8withsingle-strandedDNA is
similar to that observed with the procaryotic
single-strandbindingproteins T4gene32protein
and SSB. The highly cooperative nature ofthe
interaction seen with the electron microscope
would indicateanapparentassociationconstant
of >107 by analogy with studies on these
pro-caryoticproteins (13, 16). Both SSB andT4gene
32proteinare involved in DNAreplication and
recombination. Recently isolated
temperature-sensitive mutants with lesions in ICP8 have a
DNA-negative phenotype (6;D.Knipepersonal
communication), indicatinganessential role for
ICP8 in HSV DNA replication. Thus, in the
HSV system, ICP8 may play a role similar to
that of T4 gene 32 protein and SSB in the
bacteriophageT4 andE. colisystems.
I thank AnnaC.Weirforexcellent technical assistance and ChristaStanoyevitchfortypingthemanuscript.
This workwas supported bygrant C07114from the Uni-formed ServicesUniversityof theHealth Sciences.
LITERATURE CITED
1. Alberts,B.,andL.Frey. 1970. T4bacteriophagegene
32-astructuralproteinin thereplicationandrecombinationof
DNA. Nature(London) 227:1313-1318.
2. Alberts, B. M., and G. Herrick. 1971. DNA-cellulose chromatography. MethodsEnzymol.21:198-217. 3. Anzai, T.,G. R. Dreesman, R. J. Courtney, E. Adam,
W.E.Rawls,and M.Benyesh-Melnick.1975.Antibodyto herpessimplex virus type 2-inducednon-structural pro-teins inwomenwithcervicalcancerandcontrol groups. J. Natl. CancerInst.54:1051-1059.
4. Bayliss, G. J.,H. S.Marsden,andJ.Hay. 1975.Herpes simplex virusproteins:DNAbindingproteinsininfected cells and in the virusstructure.Virology68:124-134. 5. Beck, E.,R. Sommer, E.Auerswald,C. Kurz,B.Zink,
G. Osterburg, H. Schaller,K.Sugimoto,H.Sugiaski, T. Okamoto,and M.Takanami. 1978. Nucleotidesequence ofbacteriophage fd DNA. Nucleic Acids Res. 5:4495-4503.
6. Conley,A.J.,D. M.Knipe,P.C.Jones, and B. Roizman. 1981. Moleculargenetics ofherpes simplex virus. VII. Characterization ofatemperature-sensitive mutant pro-duced by in vitro mutagenesis and defective in DNA synthesisand accumulationof fy polypeptides. J. Virol. 37:191-206.
7. Davis,R.W.,M.Simon,and N.Davidson.1971. Electron microscopeheteroduplexmethods formapping regionsof basesequencehomologyinnucleic acids. Methods Enzy-mol.21:413-428.
8. Dreesman, G.R.,J. Burek, E. Adam, R. H. Kaufman, J. L. Melnick, K. L. Powell, and D. J. Purlfoy. 1980. Expression ofherpesvirus induced antigens in human cervicalcancer. Nature(London)285:591-593. 9. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968.
Characterizationofherpessimplexvirusstrainsdiffering intheir effectonsocialbehavior of infected cells. J. Gen. Virol. 3:357-364.
10. Flannery,V.L.,R.J.Courtney,and P. A.Schaffer.1977. Expressionofanearly,nonstructural antigen ofherpes simplex virus in cells transformed in vitro by herpes simplex virus.J.Virol.21:284-291.
11. Honess, R. W., and B. Roizman. 1974. Regulation of herpesvirusmacromolecularsynthesis.I.Cascade regula-tion ofthesynthesisof three groups of viralproteins.J. Virol. 14:8-19.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.490.57.440.71.294.2]666 NOTES
12. Morse, L.S.,L.Pereira,B.Roizman,andP. A.Schaffer. 1978. Anatomy ofherpes simplex virus (HSV) DNA. X. Mapping of viralgenesby analysis of polypeptides and functionsspecified by HSV-1 x HSV-2 recombinants. J.
Virol. 26:389-410.
13. Newport, J. W.,N.Lonberg, S.C.Kowalczykowski, and P.H.vonHippel.1981. Interactions ofbacteriophage T4-codedgene32protein with nucleic acids.II.Specificity of
bindingtoDNA and RNA. J. Mol. Biol. 145:105-121. 14. Powell, K. L., E. Littler, and D. J. M. Purifoy. 1981.
Nonstructuralproteins of herpes simplex virus.II.Major
virus-specific DNA-binding protein.J. Virol. 39:894-902. 15. Purifoy,D.J.M.,andK.L. Powell.1976.DNA-binding proteins induced by herpes simplexvirus type 2 inHEp-2 cells. J. Virol. 19:717-731.
16. Schneider, R. J.,and J. G. Wetmur. 1982. Kinetics of transferof Escherichiacoli single strand deoxyribonucleic
acidbinding protein between single-stranded deoxyribo-nucleic acid molecules. Biochemistry 21:608-615. 17. Sigal, N.,H.Delius,T.Kornberg,M.L.Gefter,andB. M.
Alberts. 1972. ADNA-unwinding protein isolated from Escherichia coli: its interaction with DNA and DNA polymerases. Proc. Natl. Acad. Sci. U.S.A. 69:3537-3541.
18. Weiner, J. H., L. L. Bertsch,and A.Kornberg. 1975. The deoxyribonucleic acid unwinding protein of Escherichia coli: properties and functions in replication. J. Biol. Chem. 250:1972-1980.
19. Wray, W., T. Boulikas, V. P. Wray, and R. Hancock. 1981. Silver staining of proteins in polyacrylamidegels. Anal.Biochem. 118:197-203.
20. Yeo, J., R. A.Klllington,D. H.Watson, and K. L. Powell. 1981. Studiesoncross-reactive antigens inthe
herpesvi-ruses.Virology 108:256-266.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/