0022-538X/94/$04.00+0
Copyright (C 1994,American Society for Microbiology
Herpes
Simplex Virus Type 1
Recombination: the
UC-DRI
Region Is Required for High-Level
a-Sequence-Mediated Recombination
REBECCA ELLISDUTCH, BORIS V. ZEMELMAN, ANDI. R. LEHMAN* Department of Biochemistry, Stanford University School of Medicine, Stanford, Califomia 94305
Received 21 December 1993/Accepted 2 March 1994
The a sequences of herpes simplex virus type 1 are believed to be the cis sites for inversion events that generatefourisomericforms of the viral genome. Using an assay that measures deletion of a ,-galactosidase genepositioned between twodirectlyrepeated sequences in plasmidstransientlymaintained in Vero cells, we hadfoundthat the a sequence is morerecombinogenicthan another sequence of similar size. To investigate the basis for theenhanced recombination mediated by the a sequence, we examined plasmids containing direct repeats ofapproximately 350 bp from avarietyofsources and with a wide range of G+C content. We observed that all of these plasmids show similar recombination frequencies (3 to 4%) in herpes simplex virus type 1-infected cells. However, recombination between directly repeated a sequences occurs at twice this frequency (6 to 10%).In addition,we find that insertion of acleavagesite for ana-sequence-specificendonuclease into the repeated sequences does not appreciably increase the frequency ofrecombination, indicating that the presence of endonuclease cleavage sites within the a sequence does not account for its recombinogenicity. Finally, byreplacingsegments of the a sequence with DNAfragments ofsimilar length, we have determined that onlythe95-bp
UC-DR1
segmentisindispensable for high-level a-sequence-mediated recombination.Herpes simplex virustype 1 (HSV-1) is anenveloped virus with a 152-kb linear duplex DNA genome. There are two unique regions in the HSV-1 genome, termed UL (unique long) and
US
(unique short). These are flanked by repeated regions, such that the final structure can be represented as a,,b-UL-ba,,,'c'-Us-ca
(41). Recombinationoccursthroughthe inverted repeats during HSV-1 infection, giving rise to four equimolar isomeric forms that differinthe orientation of their unique regions (18, 23, 45). There is considerable evidence that thisinversion process occursthroughthe HSV-1 a sequences. Insertion of fragments containingana sequence into the viral thymidine kinase gene produces new inversions in regions flanked by the inverted a repeats (32, 50) and deletions of regions with directly repeated a sequences at their ends (32, 50). In addition, the few noninvertingmutantsof HSV-1 that have beenidentified contain large deletionsencompassingthe a sequences attheUL-US
junction (25,37).The HSV-1 a sequencevaries from 200 to500bp in length and is approximately 83% G+C. The a sequence is essential forcleavageandpackaging of HSV-1(35,56)andalso includes the promoter for the ICP34.5 gene (10). The a sequence contains 20-bp direct repeats (DR1) at each end and two
unique segments (Ub and
UJ)
separated by arrays of direct repeats. The composition of the internal directly repeated elements oftheasequencevaries from straintostrain(15,31, 33, 55,56),butallcontainanarrayof12-bpDR2 repeats.The DR2 repeatshaveseveralfeatures that couldpotentially playarole in a-sequence-mediated recombination. They have been shown to adopt a novel DNA conformation under the
influ-ence ofnegative supercoiling (62, 63), and they are sites for cleavage byaHSV-1-induced endonuclease (61).
High-level recombination between either a sequences or other pairs of homologous sequences requires both HSV-1 infectionand replicationof the DNAcontaining the repeated
*Correspondingauthor.
sequences. Recombination between inverted repeats ofthea sequence that had been stably integrated into the cellular genomealong withaflanking HSV-1 origin of replicationwas shown to occur only in conjunction with HSV-1-dependent amplification (34). Similarly, recombination between Tn5 re-peats within plasmids was observed only in association with HSV-1DNAreplication (58).Ourprevious studies of recombination between a sequences inplasmids transiently introduced into Vero cells confirmed that high levels of recombination wereseen only when theplasmids underwentreplicationinHSV-1-infectedcells (20).Analysis of the time courseofHSV-1 recombinationand replication indicated that the two processes parallel each other,and directquantitationof theproductsof recombination by Southern analysis demonstrated that recombination oc-curred during or subsequent to the last round of DNA replication (20).
The exact nature of the high-level a-sequence-mediated recombination remains unclear. HSV-1-infected cells show high levels of homologous recombination, and repeated
re-gions such as the HSV-1 Bam L sequence(38), the HSV-1 b sequences(30),orthe TnS repeats(58) canmediate inversion
events. The a sequence does, however, appear to be more
highly recombinogenic than other sequences: Weber and
co-workersdemonstrated that the3.0-kb b-a-cjunctionwasmore recombinogenic thana3.0-kbplasmidsequence(59), andour
previous work showed that recombination between a
se-quences was twice as efficient as recombination between a
differentset of direct repeats ofthe samesize(20). Recombi-nation betweenasequencescould beenhancedeither because itis ahot spot forhomologousrecombination orbecause itis
actedonbyasite-specific recombinase.The findingthatthea
sequencesofHSV-1 and HSV-2,which have little homology, areincapable ofrecombiningwith each other arguesagainsta site-specific mechanism (51). However, the simple conditions of an in vitro system for a-sequence recombination (7) are
indicative ofasite-specific mechanism.
In thisreport,wedescribe ourcontinuedstudies of
recom-3733
on November 9, 2019 by guest
http://jvi.asm.org/
bination with plasmids transiently introduced into Vero cells.
We find that directrepeatsfrom avarietyofsourcesand with awiderangeof G+Ccontentgive essentiallyidentical recom-bination frequencies. However, directly repeateda sequences
show recombination frequenciesthat are twice thatseenwith
the other sequences. Introduction ofa cleavage site for the
a-sequenceendonuclease identifiedbyWohlrab and coworkers
(61) does not increase the frequency ofrecombination,
indi-cating that the presence of cleavage sites for this enzyme
cannot account for the recombinogenic nature of the a se-quence. Finally, replacement of portions of the a sequence
with DNAsegmentsof similarlengthshowed that theUc-DR1 region contains the cis signals required for high-level
a-se-quence-mediated recombination.
MATERIALS ANDMETHODS
Cells and viruses. All tissue culture experiments were
carried out with Vero cells (African green monkey kidney fibroblasts) obtained from the American TypeCulture
Collec-tion. Cells were propagated in Dulbecco modified Eagle minimum essential mediumsupplementedwith 10% fetal calf
serum,0.1 mM nonessential aminoacids,and 2 mMglutamine. The HSV-1 strain F(A305) (39) was used.
Plasmid construction. All restriction enzymes and linkers
usedinplasmidconstructionwereobtained from NewEngland Biolabsorthe United States BiochemicalCorp.Plasmidswere
prepared either by alkalinelysisfollowed by subsequent CsCl
gradient purification (43) or by the alkaline lysis procedure
followedby purification on Qiagen columns according tothe
manufacturer's instructions. Thetwomethodsgave essentially
thesameresultsin transfectionexperiments.All DNAsamples foranindividual transfectionexperimentwereprepared bythe
samemethod.
PlasmidpRD107 (20) wasused asthe parentvectorfor all
constructions. This plasmid contains the ampicillinresistance
gene and the origin ofreplication of pUCl9, permitting its
propagation in Escherichia coli. It also contains the complete lacZ gene with ribosomal bindingsites that allow transcripts madeby runoff transcription tobe translated. Upon transfor-mation with pRD107, E. coli DH5ot (RecA- LacZ-) gives deep blue colonies when plated in the presence ofampicillin and 5-bromo-4-chloro-3-indolyl-fi-D-galactopyranoside (X-Gal). In
addition,pRD107containsan HSV-1 originofreplication, Orin,
which allows theplasmid toreplicate inHSV-1-infected cells. Plasmidscontaining directrepeatsofhomologoussequences were prepared by inserting acopy of the desired insert into
both the XbaI andBamHIsitesofpRD107,sothat the inserts
were positioned in directly repeated orientation. In all cases,
the insertswere prepared for insertion by preliminary
restric-tion digestion, addition of either XbaI or BamHI linkers, digestion withXbaI orBamHI, purification on agarose-TAE
(40mMTris-acetate, 1 mMEDTA) gels, andelution from the
agarosegel with an Elutrap electrophoresis chamber
(Schlei-cher & Schuell). For pRDMS2, the insert was a 374-bp
PstI-to-SacIIfragment from themouseNa+/K+ ATPasegene,
kindly provided by S. Ying Tam, Harvard Medical School. For pRDBR2, the insertwasa378-bp SphI-to-EagI fragment from
pBR322 (nucleotides 562to939). This fragment containedno
partoftheparent vector,pRD107. pRD+X2 contained 400-bp insertsgenerated by SspI-DraI digestion of XX174 (nucleotides 1007to 1406; New England Biolabs). The 390-bp fragment in pRDU32wasfromtheyeastURA3gene,generated byaStuI-NsiI
digestofyRP17 (nucleotides 3556to3945). pRDSVS2 containeda
370-bp insert from aHinclI digest of simianvirus 40
(nucle-otides 2297 to2666). ForpRDUbx2, a380-bp fragmentfrom
an ApaI-PstI digest of
p(UT)2(Hind)D-33/t3CAT
(27)
was used. This insert containsmultiple copies
of the Ubx DNA binding site fromDrosophilamelanogaster.Finally,
pRD1 10,as described previously (20), contains an insert from thed-signaling gene
(dsg)
of Myxococcusxanthus,
provided by
YvonneChengand DaleKaiser,Stanford
University. pRD105,
also described previously
(20),
contains twocopies
of the a sequencefrom HSV-1 strain KOS,provided byJamesSmiley,
McMaster University.
Toconstructplasmidsubstrates
containing cleavage
sites for thea-sequence-specific
endonucleasereported by
Wohlrabetal.
(61), (dG)25
and(dC)25 oligonucleotides
flankedby
XhoIsites were obtained from the PAN
facility,
BeckmanCenter,
StanfordUniversitySchool of Medicine. The
oligonucleotides
wereannealed bymixing equal volumes, heatingto
85°C,
and then cooling to room temperature. The annealedoligonucle-otideswerethendigested overnightwithXholand
purified
on a4% NuSievelow-melting-point
agarosegel
(FMC
BioProd-ucts). The vectors pRDMS1 (pRD107 containing only one mouse Na+/K+ ATPase insert) and pRD111
(pRD107
withonedsg insert) were prepared for insertionby
cleavage
withXhoI,
which cuts once in the mouse and dsg inserts and for which thereare nosites in theparentvectorpRD107.Insertion of the(dG)25
*(dC)25 oligonucleotide
into theprepared
vec-tors resulted inplasmids
msendol and dsgendol. A second copy of the mouse and dsg inserts nowcontaining
the endo-nucleaserecognitionsitewasthen addedtotheBamHI site of thevectorssuch thatthe repeatswerecompletelyhomologous and in the directly repeated orientation, creatingplasmids
msendo2 anddsgendo2. Finally,plasmid dsgmsendowas con-structedbyinsertingacopyof themouseNa+/K+ATPasewith inserted
(dG)25
*(dC)25
into the BamHI site ofdsgendol.
Thus, this plasmid has two nonhomologous inserts, both
containingthe endonuclease site.
Plasmids for analysis of the recombinogenic properties of variousregionsof theasequencewereconstructedby remov-ing portions of the a sequence and replacing them with segmentsof thedsgsequence.Thewild-typea sequencefrom HSV-1 strain KOS is 317 bp in length and contains both
direct-repeatandunique regionssuch thatits finalstructureis
DRl-Ub-(DR2)1,-Uc-DRI
(56).
PlasmidpRDUb-2wasconstructedinthefollowingmanner.
The317-bpa sequencewasremoved frompRD105byBamHI
digestion, isolatedon a1% agarose-TAEgel,and
purified
by Elutrap. This fragmentwasthen digestedwith therestriction enzyme EaeI, and the resulting 222-bp DNAfragment,
with the sequenceEaeI-(DR2-Uc-DR1)-BamHI,
wasgel
purified
and isolated by Elutrap. The required dsg fragment was generated bydigestionofpRD111withCspI,addition ofEagIlinkers(EagIandEaeIproducecompatiblecohesive
ends),
and digestionwith BamHI andEagI.Theresulting 120-bp fragment
was
purified
byusing
a1%agarose-TAEgel
andElutrap.
ThisBamHI-(dsg)-Eagl
fragment was cloned into the Bluescriptcloning
vector(Stratagene)
cutwith BamHI andEagI
toobtain sufficient quantities of DNA and again excised andpurified
priorto use.Theasequenceanddsg fragmentswerejoined by
simultaneous insertion into the Bluescript vector
previously
digested withBamHI,
giving a 342-bp insert of structureBamHI-(dsg-DR2-Uc-DR1)-BamHI.
This insert wasplaced
into the XbaI site of pRD107
(XbaI
linkers added prior to insertion) togivepRDUb-1
and thenintotheBamHI site ofpRDU,-
1 toyield pRDUb-2.Toconstruct plasmid pRDR2-2, twoa-sequence
fragments
were needed. The first, containing the
DR1-Ub portion,
was generated bycleavingthepurified 317-bpasequencewithEaeI andpurifying
theresulting 95-bp fragment through
1%on November 9, 2019 by guest
http://jvi.asm.org/
rose-TAEandElutrap, givingafragment ofstructure
BamHI-(DR1-Ub)-EaeI. The DNA fragment containing the
Uc-DR1
wasgenerated by digestion of pRD105 with DraI andSphI,gel and Elutrap purificationoftheresulting270-bpfragment,and digestion withMnlI for 6 h to remove the DR2 region. The resulting fragmentwasthenfilled in with the Klenow fragment ofDNApolymerase I.XhoI linkerswere added, and productwasdigestedwith BamHIandXhoI. The final 100-bp fragment, with the structure
XhoI-(Uc-DR1)-BamHI,
was gel and Elu-trappurified. Finally,the desireddsg fragmentwasconstructedby cleavageofpRD109 with BamHI andStuI, addition of EagI
linkers, digestion with XhoI and EagI, and gel and Elutrap
purification. This 126-bp fragment, XhoI-(dsg)-EagI, was cloned into Bluescript previously digestedwithXhoIandEagI in order toobtain sufficient quantities of the DNA. Once the three required fragments were obtained, the
BamHI-(DRI-Ub)-EaeI andEagI-(dsg)-XhoI pieceswereligatedby simulta-neous insertion into Bluescript previously digested with BamHIandXhoI. Thisplasmidwasgrownin large quantities, and the221-bp insertwas cut outwithBamHI andXhoI and
purified.Next, this
BamHI-(DR1-Ub-dsg)-XhoI
fragmentand theXhoI-(Uc-DR1)-BamHI
DNA werejoined by double in-sertion into Bluescript previously digested with BamHI. Thisprocedure generatedthe final321-bpinsert,
BamrHI-(DR1-Uh-dsg-Uc-DR1)-BamHI,
whichwassubsequentlycloned into the XbaI site of pRD107(using XbaI linkersonthe insert)togivepRDR2-1and then into theBamHI site of pRDR2-1in direct orientation togivepRDR2 2.
Threefragmentswere alsorequiredtogenerate
pRDUc-2.
The first,the95-bp
BamHI-(DR1-Ub)-EaeI
fragment,wasthesame asdescribed forpRDR2-2. Thesegmentcontainingthe DR2 region was isolated by cleavage of pRD105 with SstII, purification of the 217-bp fragment generated, cleavage by DraI andEaeI,and isolation of the resulting 122-bp fragment
[EaeI-(DR2)-SstII].Thesetwosegmentswereligated by
simul-taneous cloning into Bluescript previously digested with
BamHI and SstII to give BamHI-(DR1-Ub-DR2)-SstII. Next, the dsg fragment described for pRDR2-2 was inserted by
blunt-end ligation into the Sacl site of the Bluescript vector containing the required portions of thea sequence. Cleavage
of the resulting plasmidwithBamHI and RsaI (which cleaves
near the end ofthe inserted dsg sequence) yielded a 337-bp
insert with structure
BamHI-(DR1-Ub-DR2-dsg)-RsaI
con-taining 12 bp ofBluescript vectorbetween the DR2 and dsg sequences.Thisinsertwascloned into theXbaI siteofpRD107 (usingXbaI linkers) to createpRDU- 1 andinto theBamHI site ofpRDUc-
1 in direct-repeat orientation to givepRDUC-2.
Thecorrect construction of the three replacement constructs was confirmedby DNAsequencing.Transfection, infection,and DNA isolation.Actively growing Verocellsat aconcentration of5 x
106
to7 x106
cells per mlwereelectroporated (12) at220 Vwith20 ,ugof the indicated
plasmid.Cellswereallowed to recoverforapproximately24 h. The transfected cells were then infected with 10 PFU/cell HSV-1
F(A305)
in 3 mlof mediumormock infected with3 ml of medium. Virus and medium were removed after 1 h, the cells were washed twice with phosphate-buffered saline (ob-tained from Gibco), and the mediumwasreplaced.DNA was extracted from the transfected cells using the RAPP procedureas described previously (20).
Transformation, miniprepisolation ofDNA,and restriction analysis. Competent frozen E. coliDH5oc cells (0.2 ml) were prepared and transformed with 20 to 100 ng of DNA bythe method of Hanahan (22) except that the growth medium consistedof5gof Bacto YeastExtract,20 gofBactoTryptone, and 5 gofMgSO4perliter. The cellswere platedwith 100 ,ug
of carbenicillinand 50 Figof X-Gal per ml. Miniprep isolation ofDNA from white colonies wasperformed by the modified boiling procedure(43), with 50 mM Tris-Cl (pH 8.0)-62.5 mM EDTA-0.4% Triton X-100-2.5 M LiCl used as the resuspen-sion buffer. The DNA was digested with Dral for 2 h and electrophoresed through 1% agarosegels in TAE.
Cleavage of plasmid DNA in vitro. A 0.4 M NaCl extract was prepared from nuclei of HSV-1-infected Vero cells, centri-fugedat 100,000 xg,andprecipitated with ammoniumsulfate aspreviously described (7). The dissolvedprecipitate was then dialyzed and applied to a column of heparin-agarose and elutedwitha50ml0.1 to 1.0 MNaClgradient. The0.5 MNaCl heparin-agarose fraction contains the a-sequence-specific cleavage activity.
One microgram of plasmid DNA linearized with SspI was incubated with 500 ngof the heparin-agarose fraction for 10 min at 37°Cin a bufferconsisting of 25 mM N-2-hydroxyeth-ylpiperazine-N'-2-ethanesulfonic acid (HEPES;pH7.6), 1 mM dithiothreitol, 1 mM spermidine,2 mMMgCl2, 1 mM EDTA, and 50 mM NaCl. EDTA (10 mM) was used to stop the reaction, after which theDNA waspurified by using the Magic DNACleanup System (Promega, Inc.).
Two hundred nanograms ofenzyme-treated DNA was di-gested with FspI and Scal, precipitated, and resuspended in 80% formamide-1x Tris-borate-EDTA(TBE) loading buffer. Thesampleswereboiled, chilledon ice, andloaded onto an 8 M urea-1.2X TBE 5% Long Ranger gel (J. T. Baker, Inc.). Thegelwas runwith 0.6x TBEbufferat27Wand50 to55°C. Following electrophoresis, thegelwasequilibrated for20 min in 1xNAQ buffer(80mMTris-HCl, 118 mMborate,2.4 mM EDTA [pH 8.3]). The DNA was then transferred with 1X NAQ buffer onto a 0.22-pLm-pore-size Nytran membrane
(Schleicher&Schuell)at2.5
mA/cm2
for I h, usingaGraphite Electroblotter II (Millipore, Inc.). Following transfer, the membranewas rinsed with 5x SSC(Ix SSC is 0.15 M NaCl plus0.015 M sodium citrate), and theDNA was immobilized onto the damp membrane by cross-linking in a Stratalinker oven (Stratagene, Inc.)with aUV doseof 120 mJ/cm2.Bandswere detected in thefollowing manner. Oligonucle-otides CGCGATCTGGCGACCACACCCGTCC and CCAC AGGACGGGTGTGGTCGCCAGATCGCG were tailed at their 3' ends withdigoxigenin-11-dUTP/dATPby using termi-naltransferaseandusedtoprobe the membrane accordingto instructions provided with Genius 6 kit (Boehringer Mann-heim, Inc.). The membrane was then processed with anti-digoxigenin antibody coupled to alkaline phosphatase and Lumigen PPDchemiluminescentsubstrate, using the Genius7 kit and instructions suggested by the manufacturer (Boehr-inger Mannheim). Followinga2-hincubationatroom temper-ature in the dark, the membrane was exposed to Hyperfilm ECL X-ray film (Amersham, Inc.) for 30 min. The film was developed in an automated film developer (Eastman Kodak, Inc.).
Assay for recombination frequency. The assay used to
measure recombination between sets of direct repeats has been previously described (20). Briefly, plasmids containing
directly repeatedsequencesflankingalacZ genewere electro-poratedinto Verocells,andthe cellswereinfected with HSV-1
ormock infected.Eighteenhours afterinfection,the DNAwas
isolatedbythe RAPPprocedureand thentransformed into E. coliDH5cx.The number of blue andwhitecolonieswasscored
todeterminethefrequencyoflacZdeletion.Sinceeventsother than recombination between the direct repeatscanleadtoloss of a functional lacZ gene, the DNA was isolated from a fraction of thewhite colonies bythe miniprepprocedure and examinedbyrestriction analysis.
on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 1. Comparisonof recombination frequencieswithplasmidscontainingdirectrepeats
Plasmid Source of DNAinsert % G+C HSV-1infection"
%l
White colonies(SDe)
Totaln. Avg whites Expt I Expt2 Expt 3 of colonies coloniespRD105 a sequence 83 - 0.2(0.1) 0.5(0.2) 0.4(0.2) 2,875 0.4
+ 8.7(0.8) 9.8(1.9) 12.6(1.0) 2,433 10.4
pRDMS2 Mouse Na!/K+ 73 - 0.2(0.1) 0.3(0.2) 0.2(0.1) 3,181 0.2
ATPase
+ 4.5 (0.4) 2.5(0.6) 3.9 (0.5) 4,632 3.6
pRD110 M. xanthus dsg 65 - 0.4(0.2) 0.2(0.2) 0.6(0.2) 2,759 0.4
+ 3.4 (0.4) 5.7(1.0) 3.3(0.5) 3,696 4.0
pRDBR2 pBR322 60 - 0.2(0.1) 0.6(0.3) 0.5(0.2) 3,012 0.4
+ 3.7(0.4) 3.1(0.5) 5.9(0.6) 4,926 4.2
pRD+X2 fX174 45 - 0.2(0.1) <0.4 0.4(0.4) 1,996 0.3
+ 3.5 (0.4) 3.5(0.8) 6.7 (0.6) 4,602 4.6
pRDU32 YeastURA3gene 42 - 0.6(0.3) <0.4 0.6(0.3) 2,062 0.5
+ 4.7 (0.6) 2.2(0.6) 3.0((1.7) 2,261 3.3
pRDSVS2 Simian virus40 40 - 0.4(0.2) 0.1 ((.1) 0.2(0.1) 2,704 0.2
+ 3.0(0.5) 3.5(0.5) 4.3(1.0) 2,629 3.6
pRDUbx2 D. melanogaster Ubx 22 - <0.1 <0.1 0.4(0.2) 3,030 0.2
binding sites
+ 3.1(0.6) 2.8(0.5) 4.9(0.6) 3,329 3.6
"Transfectedcells were either infected with HSV-1 (+)or mockinfected(-). "Calculatedas[(1-frequency) (frequency)/numberofcolonies]"2.
RESULTS
Recombination between a sequences occurs at twice the frequencyof recombination betweenany other set of homolo-gous sequences. Wehad previously shown that recombination betweenasequencesinreplicating plasmids is twiceasefficient as that seen between two directly repeated copies of dsg, a
segmentofDNAfromM.xanthus of similar size but with no significant homology to the a sequence (20). To determine whether the high G+C content of the a sequence (83%) is responsible for the increasedfrequency ofrecombination,we constructed six plasmids with the repeats all 370 to 400 bp (compared withthe317-bpasequence) but with G+Ccontent varying from 22 to 73%.
In the absence of superinfection with HSV-1, all of the plasmids showed a low level of recombination (<1% white colonies), consistent withourpreviousfinding (20), and there was nosignificant difference inthe recombination frequencies ofany ofthe plasmids (Table 1).
In the presence of HSV-1 infection, all plasmids showed a greatly increased frequency of recombination. As before, pRD105, containing theasequences, gaveanaverageof10.4% whitecolonies, andpRDI10, the plasmid withdirectlyrepeats dsg sequences, showed a recombination frequency less than halfthatofthea-sequence-containing plasmid (4%).Allother repeated sequences recombined with a frequency similar to thatof thedsgsequence, regardless of thesourceofthe DNA or their G+C content. Restriction digest analysis ofplasmid DNAfrom thewhite colonies ofeach sample showedthat60 to80% containedaplasmid thatwasconsistent with adeletion through the directly repeated sequences. Thus, we can con-clude that the frequency of homologous recombination
be-tween these direct repeats inplasmids replicating in HSV-1-infected cells is 3 to 4% and is unaffected by G+C content rangingfrom 22 to73%. Recombinationbetween a sequences isat least twiceasefficient, confirming the special recombino-genicproperties of thea sequence.
Addition of sites for an a-sequence-specific endonuclease does not significantly increase recombination between direct repeats.An endonuclease that cleaves within the a sequence has been identified and partially purified by Wohlrab and
coworkers (61). It wassuggested that thisenzyme mayplaya
roleina-sequence-mediated recombination bygenerating dou-ble-strand breaks that could behighlyrecombinogenic.To test this hypothesis, we constructed plasmids in which another substrate for the endonuclease, a duplex oligonucleotide, (dG)25*(dC)25 (61),wasinserted into bothrepeatsofpRD110 and pRDMS2. Toverifythat these sequences werecleaved, a partially purified enzyme fraction from HSV-1-infected nuclei abletocleave theDR2 repeatswithin theasequence(datanot
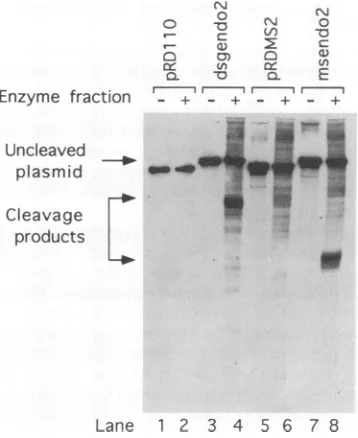
shown) and which is probably identical to the enzyme de-scribedby Wohlrab and coworkers(61)wasincubatedwith the modified plasmid substrates. The DNA wasthen purified and analyzed by modified Southern analysis. In the absence of enzyme,little cleavage of theDNA occurred with anyof the samples (Fig. 1, lanes 1, 3, 5,and7). Whenenzymewasadded, pRD110 andpRDMS2, which lack the endonuclease substrate insert, showed nosignificant cleavage (Fig. 1, lanes2 and 6). However, the two plasmids with the
(dG)25
*(dC)25
insert produceda400-bp product for dsgendo2 anda250-bp product for msendo2, as predicted for cleavage at the inserted endo-nuclease site(Fig. 1, lanes4and8). Theamountofcleavageat the(dG)25
*(dC)25 insertion was similar to that for the asequence(data notshown).
When recombination between directrepeatscontaining the
(dG)25*
(dC)25
was examined in uninfected cells, a low fre-quencyofwhite colonieswasobserved(Table 2).Foursamples gavesignificantly highervalues than normally seen.However, these higher values are randomly dispersed, and restriction analysisofthe DNAisolatedfromthe white coloniesshowed veryfewcorrect products.AfterHSV-1 infection,there was alarge increasein recom-bination frequency (Table 2). pRD105, the a-sequence-con-taining plasmid,showed the highestpercentageof white colo-nies (5.8%). pRD110,with the dsg repeats, again gavewhite colonies at approximately half the frequency seen with pRD105. dsgendo2, formed by addition of the endonuclease substrate site to the dsg sequences, gave recombination fre-quencies thatwereverysimilartothose forpRD11O.Thedata obtained withplasmids pRDMS2 and msendo2are somewhat more complex. pRDMS2 gave 3.0 to 4.4% white colonies in
on November 9, 2019 by guest
http://jvi.asm.org/
cs u) C f en CL 7}a)Q
r---- --ir
_ + _ + _ + _ +
Lane 1 2 3 4 5 6 7 8
FIG. 1. Southern analysis ofendonuclease cleavage of sites
con-taining a (dG)25*(dC)25 insert. The experiment was performed as
described in Materials and Methods. Lanes 1,3,5, and 7,untreated
DNA; lanes 2, 4,6,and8,DNAtreated withenzymefraction. Lanes 1
and 2, pRDI 10; lanes 3 and 4, dsgendo2; lanes 5 and 6, pRDMS2;
lanes 7 and8,msendo2.
five of the six samples. The reason for the high value in
experiment 4 is unclear. The endonuclease insert in msendo
appeared to give significant stimulation in one case (experi-ment 3) but otherwise gave values very similar to those for pRDMS2. Restriction digest analysis showed no significant
difference in the percentage of correct deletion products obtained betweenplasmidswith theendonucleasecleavagesite
andthose lacking this site. Finally, plasmid dsgmsendo, which
contains one dsg-endonuclease cleavage site insert and one
mouse-endonuclease cleavage site insert, gave a very low
frequency of white colonies and nocorrectproducts in restric-tion digest analysis, demonstrating that two homologous se-quences arerequiredevenin thepresenceof thecleavagesites.
We therefore conclude that thepresenceofacleavage sitefor
the a-sequence-specific endonuclease is not sufficient to
ex-plain the consistently high levels of recombination observed between repeateda sequences.
The
UC-DR1
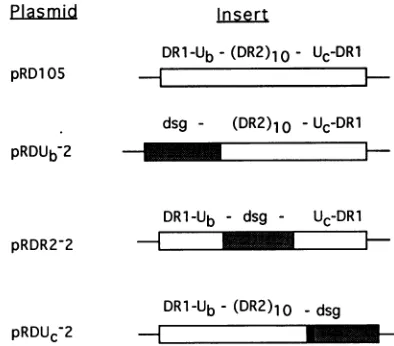
regionof the a sequence is responsible for the high levelof recombination between a sequences. Our studies confirmed thehighlyrecombinogenicnatureof theasequence but indicated that cleavage within the DR2 repeats by the a-sequence endonuclease was not sufficient. We therefore wished to determine which regions of the a sequence were necessary for the high recombinogenicity. Two groups had previouslyperformed deletion analyses of theasequence,with somewhatconflicting results (9, 49), possibly because of vari-ations in the length ofhomologoussequencesexamined. There issignificant evidence that in both prokaryoticandeukaryotic cells, homologous recombination is strongly dependentonthe length of the homologous regions (3, 26, 29, 42, 46, 48,53, 57, 64). Recombination between repeatedsequencesshorter than the minimalefficient processingsegment(approximately220 to 280 bpin mammaliancells) (29, 42) isvery inefficient. Above thissize, there isalineardependenceonlength. Consequently, it is important that the constructs to be tested for their recombination frequency be of similar size. We therefore replaced portions of thea sequence with segmentsofthe dsg sequence such that thelength ofhomologywaskept between 317bp (the size of thewild-typea sequence) and 342 bp. The three constructs (Fig. 2) contain inserts which replace theDRi-Ub
regions (pRDUb-2), the DR2 repeats (pRDR2-2), ortheUc-DR1
regions(pRDUc-2)
ofthe a sequence.Inthepresenceof HSV-1 infection, the frequency of recom-bination with
pRDU,-2,
which lacks theDR1-U1,
region,was similartothatseenwithpRD105, which contains thewild-type asequence(Table3). pRDR2-2, which lacks theDR2 repeats, gave an even higher level of recombination. Replacement of theUc-DR1
region,however, reduced recombinationtohalfof thatobtained with thewild-typeasequence.The reduced level of white colony formationseenwithpRDUc-2
wasconsistent through all of the experiments and was independent of the preparation ofDNA (Table 3,experiment 5). Thus,when the total length ofhomology is keptconstant,eithertheDRi-Ub
region ortheDR2 repeats canbereplaced withnosignificant
decrease in recombination frequency, indicatingthat theyare not responsible forthe increasedfrequency of recombination seenwiththea sequence. Instead,thecis signal forhigh-level a-sequence recombination resides in the
Uc-DR1
segment. Replacement of this segment reduces the recombinationfre-TABLE 2. Effects of insertion ofa-sequence-specific endonuclease substratesonrecombination
HSV-1I White colonies (SD) Totalno. Avg l white
Plasmid infection Expt 1 Expt 2 Expt3 Expt 4 Expt5o
of
colonies coloniespRD110 - 0.3(0.2) 0.6(0.2) 2.8(1.0) 0.1 (0.1) - 4,338 0.9
+ 3.4(0.6) 3.3(0.4) 4.1(1.2) 3.9(0.4) 2.3(0.4),2.8(0.4) 8,985 3.3
dsgendo2 - 3.3(0.7) 0.5(0.2) 0.3(0.2) 2,667 1.4
+ 3.6(0.5) 3.4(0.6) 4.9(0.4) 3.1(0.4),2.9(0.4) 7,856 3.6
pRDMS2 - 1.8(0.6) 0).2(0.1) 0.5(0.2) 0.3(0.1) 4,30)8 0).7
+ 4.4 (0.7) 3.3(0.4) 4.4(0.6) 7.2(0.8) 3.3(0.4), 3.0(0.4) 8,920 4.3
msendo2 - 1.9(0.6) 0.2(0.1) <0.3 0.6(0.2) 3,448 0.8
+ 3.5(0.7) 4.7(0.6) 6.4(0.9) 6.8(0.7) 3.3(0.4) 5,635 4.9
dsgmsendo - 0.4(0.3) 0.3(0.2) 0.1 (0.1) 2,494 0.3
+ 0.5(0.2) 1.6(0.3) 1.4(0.4) 2.2(0.3),0.7(0.2) 6,977 1.3
pRD105 - 0.3(0.2) 0.2(0.1) 0.7(0.4) <0.1 4,176 0.3
+ 5.8(0.6) 4.0(0.6) 6.8(1.3) 8.0(0.8) 5.0(0.3),5.1 (0.4) 8,356 5.8 'HSV-1-infectedsampleswereassayedinduplicate.
notdone.
Enzyme fraction
Uncleaved 4
plasmid
Cleavage products
_-0
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.93.272.80.299.2] [image:5.612.62.562.568.711.2]Plasmid
pRD105
Insert
DR1-Ub-(DR2)10 - Uc-DR1
dsg - (DR2)10 -UC-DR1
mmmmm- L
pRDUb-2
pRDR2-2
DR1-Ub - dsg - UC-DR1
DRi-Ub- (DR2)10 -
dsg
pRDUC-2FIG. 2. Constructs with a-sequence replacements. Inserts are
shown incorrectsize proportion, with total lengths being 317 bp for pRD105, 342 bp forpRDUb7-2,321 bp for pRDR2 2, and 337 bp for pRDUC-2.Constructionsaredescribed in Materials andMethods. OI,
segmentsfrom theHSV-1 strain KOSasequence; ,segmentsfrom
theM.xanthusdsggene;M, 12-bpfragment from the polylinker region of theBluescript cloningvector.
quency to the level observed between any set of non-a-sequencedirect repeats.
DISCUSSION
Ourfindings shed newlighton several aspects of recombi-nation in HSV-1-infected cells. First, the verysimilar recom-bination frequencies observed with direct repeats in which G+Ccontentvaried from 22to 73% indicates that there isa
significant level of homologous recombination in replicating plasmids that is independentofG+Ccontent.Second, recom-bination between a sequences occurs at twice the level ob-served with these othersequences.Third,addition ofa
cleav-age site for an a-sequence-specific endonuclease does not significantly increasethe frequency ofrecombination, indicat-ing that cleavage of the a sequence by this enzyme cannot account for the high recombinogenicity of the a sequence.
Finally, the
Uc-DR1
region is the only segment of the asequence that is indispensable for the enhanced level of recombination.
Severalgroupshavedescribed thehigh frequencyof homol-ogousrecombination thatoccursinHSV-1-infectedcells(5, 6, 8, 14,24, 44, 52, 60).Weber and coworkers demonstrated that Tn5 undergoes significant homologous recombination when present in the HSV-1 genome or on a replicatingplasmid in
HSV-1-infected cells (58). Although recombination could be detected only downto600bp ofhomology,our moresensitive
assayvery likely explains our ability to detect recombination between 350-to400-bp repeats.
Asnotedabove, changes in G+Ccontent donot affect the frequency of recombination.Wewereunabletotestsequences
with G+C content as high as that of the a sequence (83%) becauseof difficultieswithplasmidconstruction. This leftopen
thepossibility thatan extremely high G+Ccontentisneeded forhigh-levelrecombination.However,oursubsequent finding
that large portions of the a sequence can be replaced with
sequences of lower G+C content without affecting recombi-nation frequencyindicates thatthe high G+Ccontentof thea
sequence isnotthe solecauseof its recombinogenicity.
The findingofanHSV-1-induced endonuclease that
specif-ically cleaves within theasequence(61)ledtospeculationthat cleavagebythis enzyme was responsible forthehigh level of recombination. There is considerable evidence that double-strand breaks in DNA can stimulate recombination (for
re-views, see references 21 and 54). However, it is clear that
cleavage by the a-sequence-specific endonuclease is not
re-sponsible fortherecombinogenicity of theasequence.Possibly
recombination occurs with such high frequency in
HSV-1-infected cells that double-strand breaks do not significantly influencethe total signal.It isalsopossible that cleavage by the a-sequence-specific endonuclease, while readily seen invitro,
did not occur within the infected Vero cells. However, our
finding that theDR2region ofthea sequenceisnot required for high-level recombination reinforces our conclusion that
cleavagewithin the DR2 repeats by the a-sequence
endonu-clease isnotthecauseofthehigh level ofa-sequence-mediated recombination.
Replacement of portions of the a sequence was used to determine whether any regions within the a sequence were
dispensable. Simple deletionsdecrease the amount of
homol-ogy, and asstated earlier,there is considerable evidencethat
thefrequencyofhomologousrecombination isdirectlyrelated
TABLE 3. Effects ofreplacement of regions of thea sequence onrecombination
HSV-1 % White colonies(SD) Totalno. Avg% white
Plasmid ifcinoinfection Expt 1 Expt 2 Expt 3 Expt4" Expt5" of coloniesoois clnecolonies
pRDUt,h-2 - 0.1(0.1) 0.2 (0.1) 0.1(0.1) -' 3,830 0.1
+ 5.7(0.5) 4.8(0.5) 8.5(0.7) 3.5(0.4), 6.0 (0.5),9.1(0.8) 12,267 6.3 6.2(0.8)
pRDR2-2 - <0.1 0.2(0.1) 0.8(0.2) 4,362 0.4
+ 7.3(0.7) 6.8(0.6) 7.6(0.7) 3.5(0.3) 9.4(0.7), 12.4 (0.7) 11,458 7.8
pRDU,J2 - 0.6(0.2) 0.4(0.2) 0.2 (0.1) 3,950 0.4
+ 3.8(0.4) 3.0(0.4) 4.1 (0.5) 3.0(0.5), 3.0(0.3),2.5(0.4), 19,814 3.0 2.5(0.3) 2.9(0.4), 3.0 (0.5),
1.9(0.2)
pRD105 - 0.2(0.1) <0.1 0.3 (0.1) 5,805 0.2
+ 5.8(0.6) 5.7(0.7) 5.5(0.6) 7.9(0.9), 7.2(0.7),9.7(0.6) 9,206 6.8 5.6(1.0)
'HSV-1-infectedsamples were assayed in duplicate.
"HSV-1-infectedsamples forpRDUb-2,pRDR2-2 and pRD105 were assayed in duplicate;HIV-1-infected samples forpRDUc. 2 wereassayedfive times, using three differentpreparaitionsof DNA.
'- notdone.
J
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.71.268.80.253.2] [image:6.612.57.556.553.694.2]tothe
length
of therepeated regions (3,
26, 29, 42, 46, 48, 53,57, 64).
Moreover,
the317-bp length
of the a sequence from strain KOS isquite
close in size to the minimal efficientprocessing
segmentsize of250 to280 bpseen in mammalian cells(29,
42).
Since recombinationfrequencies
between seg-ments smaller than the minimal efficient processing segment showanexponential decrease,
anysignificant
shorteningofthe a-sequence repeatsmight
be expected to greatly affect thatportion
of thesignal
generated by homologous
recombination. Ourreplacement analysis
indicates that both theDRi-Ub
and DR2
regions
of the a sequence are dispensable forhigh-level
recombination. Infact,
pRDR2-2,which lacks the array of DR2 repeats,gives
thehighest
recombination fre-quency. It ispossible
that these repeats, which are known toadopt
novel DNA structures(62, 63), actually
repressa-se-quence-mediated
recombination. Furtherexperiments
will be neededto test thishypothesis.
Ourmost
significant finding
is that theU,-DRI
region
is theonly
segment of the a sequence that isindispensable
forhigh-level
recombination.Repeats lacking
this regionconsis-tently
recombine at half thefrequency
seen with the full a sequence. The fact that recombination isreducedtothe levelseen with any otherset of
homologous
repeats indicates that removal of theUC-DR1
region
eliminates the specialrecom-binogenicity
associated with the a sequence. The constructpRDUC-2
does contain a short section ofBluescript cloning
vector next to the
dsg region,
but the lack ofsequenceeffectson the level of
homologous
recombination makes itunlikely
that it is ofany
significance.
The
finding
that theUc-DRI region
isindispensable
is at variance with one deletionanalysis.
Chou and Roizman (9),studying
the a sequence from HSV-1 strainF, reported
that deletion of theUb
orUc
domain didnotaffectinversion,
while DR4repeatswereimportant
and DR2 repeatswere essential for recombinationthrough
a sequences. Several factorscom-plicated
theirstudy. First,
use ofthe 153-kb HSV-1 genome makesquantitation
verycomplex.
Moreimportantly,
theseven constructsexamined variedconsiderably
insize,
from less than 100bp
towell over500bp. Thus,
it is difficultto differentiate betweenchanges
inbackground
levels ofhomologous
recom-bination and decreases due to removal of
important
portions of theasequence. Itisinteresting
thatinsertion ofafragment
containing only
theU,-DR1
segment gave little inversion in the ChouandRoizmanstudy.
Itispossible
that the difference in ourfinding
results from the use of a sequences from different strains of HSV-1.Specifically,
the strain KOS a sequenceusedin ourstudy
doesnotcontainthe DR4 repeats, but aportion
ofone DR4 repeat is present inourUc
region.
Furtherexperiments
willbe neededtodetermine whether it is this segment of theUC
that contains the cissignals
forhigh-level
recombination.Smiley
and coworkersalsoperformed
adeletionanalysis
of thea sequence and concluded that the sequencesat the endsweremost
important
for inversion(49).
Though again
compli-cated
by
theuseof the whole viralgenomeanddecreasing
size ofhomology,
their results are consistent with ourfindings.
Deletions from theUc
end of the a sequence decreased recombination muchfasterthan deletionsfrom theUb
termi-nus, and afragment
of less than 100bp
that contained theUC-DR1
(their UbS construct)
wascapable
ofdriving
inver-sions. Both of these
findings
areconsistent withourconclusionthat the
U.-DR1
region
contains the cis site for enhanced asequence recombination.
The
Uc region
ofthea sequence containspac2,
one ofthe two cis sequences essential forcleavage
andpackaging
of HSV-1.However,
it isunlikely
thatcleavage
andpackaging
play arole in recombination. Cleavage and packaging should alsorequire the paclsequencefound in the Ubregion(16,56). Deiss and coworkers (16) found that plasmids with only Uc
could be packaged, but examination of the final products revealed the presenceofUb regions that had been added by
recombination with the helper virus. In our study, miniprep and restriction digest analysis of white colonies from
pRDUbJ2
indicate thatnoadditionsoccurred,demonstratingthat enhanced recombinationcanbeseenin the absenceof the pacl signal found in Ub. In addition, there is considerable evidence that cleavage and packaging are coupled processes. Disruption of packaging, caused by deletions in the packaging signals (17)ormutations inanyof nine differentHSV-1 genes
(1, 2, 4, 19, 36, 40, 47), also prevents processing of high-molecular-weight viral DNA into unit-lengthmonomers.Thus, it islikely thatonce theprocessing of DNA for packaginghas begun, the DNA becomes unavailable for recombination. Finally, we have previously demonstrated that recombination parallels replication through the course of HSV-1 infection, with asignificant signal seenby 6 h postinfection (20). Thus, bothprocessesshouldbe well underwaybeforepackaging has begun.
Several proteins that bind in the a sequence have been identified. Dalziel and Marsden (13) noted the binding of 21-and 22-kDaproteinstotheasequence.Kemble and Mocarski
purified ahost cell protein that bindstothehighly conserved
pac-2element in thecytomegalovirusasequence(28). Finally, Chou and Roizman demonstratedsequence-specificbindingto the
Uc-DR1
region of thea sequencebytwo larger proteins, which theyspeculate tobe ICPI andICP7 (11). Therelation-ship between these proteins and a-sequence recombination remainstobe determined.
The important questionof whether thea sequence isahot spot for homologous recombination or a cis signal for a site-specific enzyme remains unresolved. However, we have
nownarrowed the regionofimportanceto the95-bp
Uc-DRI
region. It should now be straightforward to examine further the DNA requirementsof the system.
ACKNOWLEDGMENTS
Wethank S. Y.Tamfor thegiftof themouseNa+/K+ ATPase gene used in our construct pRDMS2 and F. Bradley Johnson and Mark Krasnowfor thegiftofthep(UT),(Hind)D-33/t3CATplasmidusedin theconstruction ofpRDUbx2.
R.E.D. isapredoctoralfellow of the National Science Foundation. This workwassupported bygrantsAl 26538 and AG02908 from the National Institutes of Health.
REFERENCES
1. Addison, C.,F.J. Rixon, J.W.Palfreyman,M.O'Hara,andV. G. Preston. 1984. Characterisation ofa herpes simplexvirus type 1
mutantwhich hasatemperature-sensitivedefect inpenetrationof cells andassemblyofcapsids. Virology138:246-259.
2. Addison, C.,F.J.Rixon,and V. G. Preston.1990.Herpes simplex virus type 1 UL28 geneproductisimportantfor theformation of
maturecapsids.J.Gen. Virol. 71:2377-2384.
3. Ahn, B.-Y.,K.J. Dornfeld,T.J. Fagrelius,and D. M.Livingston. 1988.Effect of limitedhomologyongene conversionina Saccha-romycescerevisiaeplasmid recombination system. Mol. Cell. Biol. 8:2442-2448.
4. Al-Kobaisi,M.F., F.J. Rixon, I.McDougall,and V. G.Preston. 1991.Theherpes simplexvirus UL33 geneproductisrequiredfor the assemblyof fullcapsids. Virology 180:380-388.
5. Brown, S. M., D. A. Ritchie, and J. H. Subak-Sharpe. 1973. Genetic studies withherpes simplexvirus type 1. The isolation of
temperature-sensitive mutants, their arrangement into
comple-mentation groups and recombinationanalysis leadingtoalinkage
map.J.Gen.Virol. 18:329-346.
on November 9, 2019 by guest
http://jvi.asm.org/
6. Brown, S. M., J. H.Subak-Sharpe, J. Harland,and A. MacLean. 1992.Analysis of intrastrainrecombination inherpes simplexvirus
type 1 strain 17 andherpes simplexvirustype2 strainHG52 using
restriction endonuclease sitesasunselected markers and
temper-ature-sensitive lesionsasselected markers. J. Gen. Virol. 73:293-301.
7. Bruckner, R.C.,R. E.Dutch,B. V.Zemelman,E. S.Mocarski,and
I. R. Lehman. 1992. Recombination in vitro between herpes
simplex virus type 1 a sequences. Proc. Natl. Acad. Sci. USA 89:10950-10954.
8. Bubley, G. J., and L. E. Schnipper. 1987. Effect of Bloom's syndromefibroblastsongeneticrecombination andmutagenesisof herpes simplexvirustype1. Somatic Cell Mol. Genet. 13:111-117. 9. Chou, J.,and B. Roizman. 1985. Isomerization ofherpes simplex virus Igenome:identification of thecis-actingand recombination siteswithin the domain of theasequence.Cell 41:803-811. 10. Chou, J.,and B. Roizman. 1986. The terminala sequenceof the
herpes simplexvirus genome contains the promoter ofa gene
located in the repeat sequences of the L component. J. Virol. 57:629-637.
11. Chou, J., and B. Roizman. 1989. Characterization of DNA se-quence-common and sequence-specific proteins binding to cis-actingsites forcleavageof the terminala sequenceof theherpes simplexvirus 1 genome.J. Virol. 63:1059-1068.
12. Chu, G.,H.Hayakawa,and P.Berg. 1987.Electroporationfor the efficient transfection of mammalian cells with DNA. Nucleic Acids Res. 15:1311-1326.
13. Dalziel, R. G., and H. S. Marsden. 1984. Identification of two
herpes simplexvirustype1-inducedproteins (21Kand22K)which interactspecificallywith thea sequence ofherpes simplexvirus DNA.J. Gen. Virol. 65:1467-1475.
14. Dasgupta, U. B., and W. C. Summers. 1980. Genetic recombina-tion of herpes simplex virus, the role of the host cell and UV-irradiation of the virus. Mol. Gen. Genet. 178:617-623. 15. Davison, A.,andN. Wilkie. 1981. Nucleotidesequenceof thejoint
between the L and Ssegmentsofherpes simplexvirus types1 and
2.J. Gen. Virol. 55:315-331.
16. Deiss,L.P., J. Chou, and N. Frenkel. 1986. Functional domains within theasequenceinvolved in thecleavage-packagingofherpes simplexvirus DNA. J. Virol. 59:605-618.
17. Deiss,L.P.,and N. Frenkel. 1986.Herpes simplexvirusamplicon: cleavageof concatemeric DNA is linkedtopackagingandinvolves amplification of the terminally reiterated a sequence. J. Virol.
57:922-941.
18. Delius, H.,andJ.B. Clements. 1976. Apartialdenaturationmap
ofherpes simplexvirustype1DNA: evidence for inversion of the
unique DNAregions. J. Gen. Virol. 33:125-133.
19. Desai, P., N. A. DeLuca, J. C. Glorioso, and S. Person. 1993. Mutations inherpes simplexvirustype1genesencoding VP5and VP23 abrogate capsid formation and cleavage of replicated DNA. J. Virol. 67:1357-1364.
20. Dutch,R.E.,R. C.Bruckner,E. S.Mocarski,andI.R. Lehman.
1992. Herpes simplexvirus type I recombination: role of DNA replicationand virala sequences.J. Virol. 66:277-285.
21. Haber, J.E. 1992.Exploringthepathwaysofhomologous
recom-bination. Curr. Opin.Cell Biol. 4:401-412.
22. Hanahan, D. 1985.Techniques for transformation of E. coli, p.
109-135. In D. M. Glover (ed.), DNA cloning: a practical
ap-proach,vol. 1. IRLPressLtd.,Oxford.
23. Hayward, G. S., R. J. Jacob, S. C. Wadsworth, and B. Roizman. 1975.Anatomy ofherpes simplex virus DNA: evidence for four populationsof molecules that differ in the relativeorientations of their long and short components. Proc. Natl. Acad. Sci. USA 72:4243-4247.
24. Honess,R.W.,A.Buchan,I.W. Halliburton, and D. H. Watson.
1980.Recombination andlinkage between structural and
regula-torygenesofherpes simplexvirustype 1:studyof the functional organizationofthegenome. J. Virol. 34:716-742.
25. Jenkins, F. J., and B. Roizman. 1986. Herpes simplex virus 1
recombinants with noninverting genomes frozen in different iso-meric arrangements are capable ofindependent replication. Vi-rology59:494-499.
26. Jinks-Robertson, S., M. Michelitch, and S. Ramcharan. 1993.
Substrate length requirementsfor efficient mitotic recombination inSaccharomycescerevisiae.Mol. Cell. Biol. 13:3937-3950. 27. Johnson,F.B.,and M. A.Krasnow. 1992. Differentialregulation
oftranscription preinitiation complex assembly byactivator and repressorhomeodomainproteins.Genes Dev.6:2177-2189. 28. Kemble, G.W.,andE. S. Mocarski.1989.Ahost cellproteinbinds
to ahighlyconservedsequenceelement(pac-2)with the cytomeg-alovirusa sequence. J.Virol. 63:4715-4728.
29. Liskay, R. M.,A. Letsou, andJ. L. Stachelek. 1987. Homology requirement for efficient gene conversion between duplicated
chromosomal sequences in mammalian cells. Genetics 115:161-167.
30. Longnecker,R.,and B.Roizman. 1986.Generation ofaninverting
herpes simplex virus I mutant lacking the L-S junction a
se-quences,anoriginof DNAsynthesis,and severalgenesincluding
those specifying glycoprotein E and the (47 gene. J. Virol. 58:583-591.
31. Mocarski, E. S., L. P. Deiss, andN. Frenkel. 1985. Nucleotide sequenceandstructural features ofanovelUs-ajunctionpresent inadefectiveherpessimplexvirusgenome.J.Virol. 55:140-146. 32. Mocarski, E. S., L. E. Post, and B. Roizman. 1980. Molecular
engineering of the herpes simplexvirus genome: insertion of a
second L-Sjunctioninto thegenomecausesadditionalinversions. Cell 22:243-255.
33. Mocarski, E. S., and B. Roizman. 1981. Site-specific inversion sequenceof theherpessimplexvirusgenome: domain and
struc-tural features. Proc. Natl. Acad.Sci. USA 78:7047-7051. 34. Mocarski, E. S., and B. Roizman. 1982. Herpesvirus-dependent
amplification and inversion of cell-associated viral thymidine
kinasegeneflankedbyviralasequencesandlinkedto anoriginof viral DNAreplication.Proc.Natl. Acad.Sci. USA 79:5626-5630. 35. Mocarski,E.S.,and B. Roizman. 1982. Structure and roleofthe
herpessimplexvirusDNAtermini ininversion,circularization and
generationofvirionDNA.Cell 31:89-97.
36. Pertuiset,B.,M. Boccara, J. Cebrian,N.Berthelot,S. Chouster-man, F. Puvion-Dutilleul, J. Sisman, and P. Sheldrick. 1989.
Physical mapping and nucleotide sequence ofa herpes simplex
virustype 1gene requiredforcapsid assembly.J. Virol. 63:2169-2179.
37. Poffenberger, K. L.,E.Tabares, and B. Roizman. 1983. Charac-terization ofaviable, noninverting herpes simplexvirus I genome derivedby insertion and deletion ofsequencesatthejunctionof componentsL and S. Proc. Natl. Acad. Sci. USA80:2690-2694. 38. Pogue-Geile, K. L., G. T.-Y. Lee, and P. G. Spear. 1985. Novel
rearrangementsofherpessimplexvirus DNAsequencesresulting
fromduplicationofasequencewithin theunique regionof theL component. J.Virol. 53:456-461.
39. Post,L.E.,S. Mackem,and B. Roizman. 1981. Regulation of (x genes of herpes simplex virus: expression of chimeric genes produced by fusionofthymidine kinase with et gene promoters.
Cell 24:556-565.
40. Preston,V.G., J.A.V.Coates,and F.J.Rixon.1983.Identification and characterization of a herpes simplex virus gene product requiredforencapsidationof virusDNA. J.Virol. 45:1056-1064. 41. Roizman, B.,and A. Sears. 1990.Herpessimplexviruses andtheir
replication,p. 1795-1843.In B. N. Fields andD.M.Knipe(ed.), Virology. RavenPress,NewYork.
42. Rubnitz, J., and S. Subramani. 1984. The minimumamount of
homologyrequiredforhomologousrecombinationinmammalian cells. Mol. Cell.Biol. 4:2253-2258.
43. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular
cloning:apractical approach,2nd ed. ColdSpringHarbor Labo-ratory, ColdSpring Harbor,N.Y.
44. Schaffer, P.A., M.J. Tevethia,and M. Benyesh-Melnick. 1974. Recombination betweentemperature-sensitive mutantsofherpes simplexvirustype1.Virology58:219-228.
45. Sheldrick, P.,and N.Berthelot. 1975. Inverted repetitions inthe chromosome ofherpessimplexvirus. Cold SpringHarborSymp. Quant. Biol.39:667-678.
46. Shen, P.,and H. V.Huang. 1986. Homologousrecombination in Escherichiacoli: dependenceon substrate length and homology.
Genetics 112:441-457.
47. Sherman, G.,and S. L.Bachenheimer. 1987. DNAprocessingin
on November 9, 2019 by guest
http://jvi.asm.org/
temperature-sensitive morphogenic mutants of HSV-1. Virology 158:427-430.
48. Singer, B. S., L. Gold, P. Gauss, and D. H. Doherty. 1982. Determination of the amount of homology required for recombi-nation inbacteriophage T4. Cell 31:25-33.
49. Smiley, J. R., J. Duncan, and M. Howes. 1990. Sequence require-ments for DNArearrangement induced by the terminal repeat of herpessimplexvirus type 1 KOS DNA. J. Virol. 64:5036-5050. 50. Smiley, J. R., B. S. Fong, and W.-C. Leung. 1981.Construction of
a double-jointed herpes simplex viral DNA molecule: inverted repeats are required for segment inversion, and direct repeats promotedeletions. Virology 113:345-362.
51. Smiley, J. R., C. Lavery, and M. Howes. 1992. Theherpes simplex virus type 1 (HSV-1) a sequence serves as a cleavage/packaging
signal but does not drive recombinationalgenome isomerization
when it is inserted in the HSV-2 genome. J. Virol. 66:7505-7510. 52. Stow, N. D., J. H.Subak-Sharpe, and N. M. Wilkie. 1978. Physical mapping of HSV1 mutations by marker rescue. J.Virol. 28:182-192.
53. Sugawara, N., and J. E. Haber. 1992.Characterizationof double-strandbreak-induced recombination:homology requirementsand single-stranded DNA formation. Mol. Cell. Biol. 12:563-575. 54. Thaler, D. S., and F. W. Stahl.1988. DNA double-chain breaks in
recombination of phage X and of yeast. Annu. Rev. Genet. 22:169-197.
55. Umene, K. 1991. Recombination of the internal direct repeat element DR2 responsible for the fluidity of thea sequence of
herpessimplexvirus type 1. J. Virol. 65:5410-5415.
56. Varmuza, S. L., and J. R. Smiley. 1985. Signals for site-specific cleavageofHSV DNA: maturationinvolvestwoseparatecleavage
events atsitesdistaltotherecognitionsequences.Cell 41:793-802. 57. Watt,V. M.,C.J. Ingles,M. S. Urdea, and W.J.Rutter. 1985.
Homology requirements for recombination in Escherichia coli. Proc. Natl.Acad. Sci.USA 82:4768-4772.
58. Weber,P.C.,M. D.Challberg,N.J. Nelson,M.Levine,andJ. C. Glorioso. 1988. Inversioneventsin the HSV-1 genomearedirectly
mediated by the viral replication machinery and lack sequence
specificity. Cell 54:369-381.
59. Weber,P. C.,M. Levine, and J. C. Glorioso. 1990.
Recombino-genic properties ofherpes simplexvirus type 1 DNA sequences resident in simian virus 40minichromosomes. J. Virol. 64:300-306. 60. Wildy,P.1955. Recombination with herpes simplex virus. J. Gen.
Microbiol. 13:346-360.
61. Wohlrab, F., S. Chatterjee, and R. D. Wells. 1991. The herpes simplexvirus 1 segmentinversion site isspecifically cleaved bya virus-induced nuclear endonuclease. Proc. Natl. Acad. Sci. USA 88:6432-6436.
62. Wohlrab, F.,M.J. McLean,and R.D. Wells. 1987. The segment inversion site ofherpessimplex virus type 1 adoptsanovelDNA structure.J.Biol. Chem.262:6407-6416.
63. Wohlrab,F.,and R.D. Wells. 1989. Slightchanges in conditions influence thefamilyof non-B-DNA conformations of theherpes
simplex virus type 1 DR2 repeats. J. Biol. Chem. 264:8207-8213. 64. Yuan, L.-W., and R. L. Keil. 1990. Distance-independence of
mitoticintrachromosomal recombination in Saccharomyces cerevi-siae. Genetics124:263-273.