JOURNALOFVIROLOGY,Nov. 1992,p.6735-6746 Vol.66, No. 11 0022-538X/92/116735-12$02.00/0
Copyright © 1992, AmericanSociety for Microbiology
The
Conserved Helicase Motifs
of
the
Herpes
Simplex Virus
Type 1
Origin-Binding Protein UL9
Are
Important
for
Function
RIK MARTINEZ, LEI SHAO,AND SANDRA K. WELLER* Department ofMicrobiology, The University of ConnecticutHealthCenter,
Farmington,
Connecticut06030Received 5 May1992/Accepted3July 1992
The UL9gene of herpes simplex virus encodes a protein thatspecificallyrecognizes sequences within the viral origins ofreplicationandexhibits helicase and DNA-dependent ATPase activities. The specificDNA binding domain ofthe UL9 protein was localized to thecarboxy-terminalone-third of the molecule (H. M. Weir, J. M. Calder, and N. D. Stow, Nucleic Acids Res.17:1409-1425,1989). The N-terminal two-thirds of the UL9 gene containssixsequence motifs found inallmembers of a
superfamily
of DNA and RNAhelicases,suggesting that this region may be important for helicase activity of UL9. In this report, we examined the functionalsignificance of these six motifs for the UL9 protein through the introduction ofsite-specificmutationsresulting in single amino acid substitutions of the most
highly
conserved residues within each motif. An in vivocomplementationtest wasused tostudy the effect of each mutation on thefunctionof theUL9protein in viral DNA replication. In this assay, a mutant UL9 protein expressed from a transfected plasmid is used to complement areplication-deficientnullmutant in the UL9 gene for theamplification of herpes simplex virus
origin-containingplasmids. Mutations in five of the six conserved motifs inactivated the function of the UL9 protein in viral DNA replication, providing direct evidence for the importance ofthese conserved motifs. Insertion mutants resulting in theintroduction of twoalanines at 100-residue intervals in regions outside the conservedmotifs were also constructed. Three of the insertion mutations were tolerated, whereas the other five abolishedUL9 function.These dataindicate thatother regions of theprotein, in addition to the helicase motifs, areimportantfor
function
in vivo. Several mutations result ininstability
ofthe mutantproducts,presumably
because ofconformational changes in the protein. Taken together, these results suggest that UL9 is very sensitive tomutations withrespect to both
stucture
andfunction,perhaps reflecting itsmultifnctional character.Initiation of DNAreplicationin manyorganismsutilizesa common mechanism involving the specific binding of an
initiator protein to a site within the origin ofreplication. Since binding of an initiator protein represents the first committed step in initiation ofDNA replication, this event
clearly has important regulatory potential. In Escherichia coli, an initiator protein, dnaA, binds DNA and facilitates unwindingattheorigin by recruitingaDNAhelicase, dnaB,
tothe site(4).Inothersystems such as theeukaryoticvirus simian virus 40, the initiator protein, large T antigen, not
only binds the origin in asequence-specificmannerbutalso
hasintrinsic helicaseactivityandiscapable of unwindingthe
duplexDNAin anATP-dependent fashion (16, 17, 19, 64). The herpesvirusesprovide a model for eukaryotic DNA
replication in which many of the replication proteins are
encodedbythevirus.The 152-kb genome ofherpessimplex
virus type 1 (HSV-1) contains three cis-acting origins of DNA replication (63, 65, 77). Two complementary
ap-proacheshave been used toidentifytrans-acting viralfactors
requiredforviralDNAsynthesis.HSV-1 mutants
represent-ing
seven distinctcomplementation
groups which fail to induce viral DNA synthesis under nonpermissive growthconditions have beenidentified(6, 7, 9,11,27, 50, 51,57, 58,
75, 76, 80) (for areview, seereference
74).
Thesecomple-mentation groups represent seven viral genes which have
been shown to be necessary andsufficientfor
origin-depen-dentplasmid
amplification
ina transient transfection assay*
Corresponding
author.(78). Biochemical activities have nowbeen assigned to the
products ofthesegenes:
UL30
andUL42
encode the viralDNA polymerase and its accessory protein (26, 56-58); UL29 encodes the major DNA-binding protein, ICP8 (12, 76); and UL5, UL8,andUL52 genes encode three members
ofa protein complex with helicase-primase activities (13). UL9 encodes the HSV
origin-specific
DNA-binding protein(53).
Purified recombinantUL9proteinhas thefollowing
prop-erties: cooperative origin-specific DNA-binding activity, DNA-dependent nucleoside 5'-triphosphatase, DNA heli-case on
partially
double-stranded substrates, and ability to form dimers in solution (5, 21, 22, 24, 39, 53). Although specific unwinding of HSV origin-containing plasmids by purified UL9hasnotbeen seen(24),severalobservationsof the interactions between UL9and duplexDNAhave been made. When UL9 protein binds to supercoiled plasmids containingons,
the structure intheATregion is distortedandtheregionbecomes sensitivetoagents suchasKMnO4 and
dimethyl
sulfate(38). Fierer and Challberg demonstratedthat UL9 wraps
supercoiled
DNA in asequence-indepen-dent fashion (24). Nucleoprotein complexes consisting of
purified UL9 and
origin-containing
DNAhave been visual-izedbyelectronmicroscopyand form inter- and intramolec-ular interactions, raising the possibility that multimers of UL9 proteinareformed atthe replication origin(59).
How-ever,despitetheseobservationsconcerning
invitro interac-tions of UL9 withduplexDNA, it is still unclear how UL9 functions in vivo during infection. In infected cells, UL9presumablybinds at one of the threeorigins of HSV DNA
6735
on November 9, 2019 by guest
http://jvi.asm.org/
I la 11 111 IV V VI
reldu# 190 290
I la 11
635uas
674sa
III IV V VI
90 390 4959T 6' 790
I Is 11 III IV V VI
190 2e0 390 490 590 690 7v0
I
////
T.
I////
614 aas
FIG. 1. Conserved motifs in a superfamily of helicases. The
number of amino acidresiduesislistedabove each bar. Amino acid
sequencesfrom severalrepresentatives of helicasesuperfamily2are
shown, includingHSVUL9;the CIproteinof thepotyvirustobacco vein mottlingvirus (TVMV) (20); UvrB, anE. coliprotein impli-cated in excision repair(32); RAD3, ayeast(S. cerevisiae) DNA
repair protein (32);ERCC-3,ahumanprotein implicatedin excision
repairand involved in the disorders xerodermapigmentosum and Cockayne's syndrome (72); andp68, ahuman proteinwith RNA
helicaseactivity (35).Thesevenconserved motifsarerepresented by dark bars(30, 31, 36).The motifsarenumberedasindicated above the UL9protein, and the order of the motifsin eachproteinis identical
althoughthespacingvaries within thissuperfamily.
replication and either acts alone to unwind the duplex or
interacts with othercomponents ofthe replication
machin-erytounwind thetemplateattheorigin. Understandingthe precise role of UL9 in the initiationof viral DNAreplication isimportanttoourunderstanding of the mechanismsof viral
DNA replication and to the development of an in vitro systemfor thestudy of HSVDNAreplication.
Proteinsequenceanalysisindicates that UL9 isamember ofasuperfamily of proteins (SF2)which include atleast40 establishedorputative helicases of E. coli,yeasts,insects,
mammals, poxviruses, and herpesviruses and threegroups
ofpositive-strand RNA viruses (31, 32, 37). Seven motifs whicharesharedamongtheseproteinshave been identified. Consistentwith their ability tohydrolyze ATP, these
pro-teins all contain two highly conserved motifs known to defineanucleotide-binding domain (motifsIand IIinFig. 1)
(71). In addition, four other conserved motifs (IIItoVI) have been observed inboth DNAand RNA helicases. Thestrong conservation of these six motifs inalarge number of RNA
and DNA helicases implies that these sequence elements maybe important for helicase function; however, nodirect
evidenceexiststodemonstrate their functionalsignificance. An additional region of sequence conservation (motif Ia) located between motifs I and II is less well conserved betweenmembers ofthesuperfamily and willnotbe consid-ered furtherinthispaper(29, 37, 45). Wereporthereinour
attempts to determine which regions of the UL9gene are
required for its function. A series of mutationswere
intro-duced into theUL9geneinaeukaryotic expression plasmid.
Each mutant plasmidwas tested in a transient replication
851 aas complementation assaydesigned to analyze the replication ability ofmutant UL9 gene products. Our results indicate
thatatleast five of the six conserved motifsareimportantto the function of the UL9geneproduct in HSV DNA replica-tion. In addition, deletion and small insertions were intro-duced into the UL9genetofurthermapimportant functional
domains. It isanticipated that thistypeof structure-function analysis will leadtothe identification ofprotein domains that
IS contribute to the various activities of the protein and will leadtoabetterunderstanding of the mechanism of action of
thisimportant protein.
779aas
783aas
MATERIALS ANDMETHODS
Cells and viruses. African green monkey kidney cells (Vero; American Type Culture Collection, Rockville, Md.)
were propagated and maintained as described previously
(76). The KOS strain of HSV-1 was used as thewild-type
virus. The lacZ insertionmutanthr94 which does not
syn-thesize the UL9 gene product and the cell lines S22 and
2B-11 which support the growth of UL9 mutants are
de-scribed elsewhere(6, 47).
Plasmids. plOO-1 containingoni on a 100-bpMspI
frag-ment was kindly provided by M. D. Challberg, National Institutes ofHealth,Bethesda, Md. (53). p6UL9 and p6UL9-119b, which contain the coding sequences of UL9 down-stream from the ICP6 regulatory sequences, were
con-structedasdescribedpreviously (47). p6UL9 isinthevector Bluescribe(Stratagene, SanDiego, Calif.), and p6UL9-119b is inpUC119 (70); bothvectorscontainanorigin of
replica-tion from the phage M13 and can thus be packaged as
single-stranded DNA in the presence of helper phage M13K07 (60). Recombinant plasmids were propagated in UT481by standard procedures (49).
Site-directed mutagenesis. Single-stranded DNA from p6UL9 or p6UL9-119b was generated following infection
with the helper phage M13K07 by standard procedures (2). Site-directedmutagenesiswasperformedonsingle-stranded
DNA in which uracil residueshad been substitutedby using theMutagene mutagenesis kit (Bio-Rad, Richmond, Calif.) according to the manufacturer's instructions (40, 41). For
some mutants, relevant small fragments were subcloned from p6UL9 into M13mp19 and site-directed mutagenesis
wascarriedoutonsingle strands propagatedin E. coliCJ236 (dut-minus ung-minus). Mutations were confirmed as
de-scribedbelow and cloned back into thep6UL9 or
p6UL9-119bconstructfortesting.Mutagenic oligonucleotideswere
synthesized by usingaCycloneDNAsynthesizer (Biosearch
Inc.,Burlington, Mass.) (Table 1).Inadditionto incorporat-ing thedesired basesubstitution,mostoligonucleotidesalso contained mutations which generateda restriction enzyme
sitepolymorphism without alteringthepredictedgene prod-uct, allowing the identification of mutants by restriction mapping.A deletion(A130-851) removingtheC-terminal721 aminoacid residueswas constructedby completedigestion ofp6UL9withSallandreligation.This deletionremoves527
bp ofthe UL9coding sequence and results ina frameshift
following the noveljunction. An in-frame deletion (A597-703) removing 106 aminoacidresidueswasconstructed by using DNA from two mutants, p6UL9-6i and p6UL9-7i (described in Results), as starting material. p6UL9-6i and p6UL9-7i each containa6-bpinsertionatpositions597 and 703, respectively, resulting in the introduction ofa unique
NotI site into each plasmid. A597-703 was constructed by UL9
I la 11 III IV V VI
1?0 1rST1r
IT
UrV VI Is 11 III IV V VI
1!p 21 v Mr T Or
TVMV
UVRB
RAD3
ERCC3
P68
I Ia 11111 IVY VI
10 094A 9n
V/1
I I I
r/l/AWA
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.55.292.73.299.2]HELICASE MOTIFS OF UL9 6737
TABLE 1. Oligonucleotides used to createmotif and insertionmutants
Mutation Motif no. orposition" Oligonucleotidesequence(5'-3')b Restriction sitechange
Motif
K87A I CGCGGTAGTTQCTCCCGACCCCATgG0CGCGCGGACC Creates NcoI
K87R I CGCGGTAGTTGTTCCCGACCCCATgGGCGCGCGGACC Creates NcoI
E175A II GGCCCAGCGTgGACATAACCACGTCCAGAACGAG AbolishesHinclI
T214S(III) III GCGTTGGCGG0ATGCGTCCAToGCGATGATCC AbolishesNcoI
T214A(IIIa) III GCGTTGGCG0.TGCGTCCATcGCGATGATCC AbolishesNcoI
A213S (IlIb) III TTGGCGGTTG,AGTCCATcGCGATGATCC AbolishesNcoI
F303W IV AGACCGTCGATGACAATGCAGAT AbolishesSail
G354A V GATCGAAGCTtAGCOCCACGGTTAC Creates HindIII
R387K VI GGAGGGTGCGgACTTCCCAGGGACT AbolishesFspI
Insertion
ii i97AA AGAGTGGATCGC CGCTTCCCGCAGCCA Creates NotI
2i i195AA TAGCATTAACGCGGCCGCATCCACGCGGCC Creates NotI
3i i291AA GCCAAGGCGCGCCfl
=TTCCAGCTCCCC
Creates NotI4i i404AA
CTCCGAGCGCOC9CCQCCCGGAGCCOGTC
Creates NotISi i520AA GGCCCTGAG00C.aBQ GTCGAAATGTAC Creates NotI
6i i597AA GCTGCGGGTGGCflCCWCATGGGAACGCG CreatesNotI
7i i703AA GCGCTTGACGGCgCXQ TTCCTCCGAGAG Creates NotI
8i i798AA CTTGAAGCGCGCWWiQGCCCGCAAACTC Creates NotI
a For the insertion mutations, the Ala Ala insertion occurs after the indicated amino acid residue.
bFormotifmutations, the underline indicates the base change to create the amino acid change; lowercase letters indicatesilentchanges to create or abolish
restriction sites. For insertionmutations,theunderline indicates inserted Ala Ala, creating a uniqueNotIsite.
digesting each plasmid with NotIandEcoRI andreligating
the appropriate fragments to generate a plasmid with a deletion between the NotI site atposition 597 in p6UL9-6i and the NotI site at position 703 in p6UL9-7i. All the
mutationswereverifiedbyDNAsequenceanalysis by using
Sequenase and the supplier's instructions (United States Biochemical Corporation, Cleveland, Ohio). Double-strandedplasmidDNA wasdenaturedpriortosequencingas
describedpreviously (10).
Transient replication complementation
(TRC)
assay. Vero cells were transfected with theoris-containing
plasmidplOO-i
by itself orcotransfected with wild-type or mutantp6UL9 by using a modification of the standard calcium
phosphate coprecipitation procedure (33). Exponential-phaseVerocells (1.5 x
106)
weretransfectedin suspension as described previously (81). At 24 h posttransfection, the medium was changed. At 30 h posttransfection, the cells were superinfected with KOS orhr94 at a multiplicity of infection of 10 PFU per cell andincubated at34°C. At48 hposttransfection, cells were harvested and total DNA was isolated as described previously (77). DNA (2
pg)
wasanalyzed by digestion withEcoRIalone or in combination with
lDnI,
followed by agarose gel electrophoresis and Southern blot hybridization. Bluescribe or pBR322 DNA waslabeled with 32P as describedpreviously (23).Transientcomplementation with plasmids. Vero cells were
transfected with2 ,ug ofwild-type or mutantp6UL9and 10 ,ug of salmon sperm DNA. At 30 h posttransfection, cells
were superinfected with hr94 as described above for the TRC assay. At 48 h posttransfection, the cultures were
harvested, frozen and thawed once(-80°C), sonicated, and clarified. Supernatants were assayed on permissive cells
(either S22 or 2B11) for total yield and on Vero cells for recombinants. This assay has been called the transient
complementation(TC)test.
Detection of UL9 proteins inviral infection and transient transfection by immunoblotting. To detect UL9 protein in cells transiently transfected with the expression clone
p6UL9-119b, 1.5 x
106
Verocellsweretransfected with 12,ug of the expression plasmid as described previously (28)
and platedin a 60-mm tissue culturedish. Induction of the
ICP6 promoter was achieved by superinfection at 30 h
posttransfection with hr94. At 48 h posttransfection, the cells were washed in phosphate-buffered saline (PBS) and
resuspended in 400
pl
ofPBSplus200pl
of3 x lysisbuffer(30 mM Tris-HCl
[pH
8.0] 15 mM EDTA [pH 8.0], 3% sodiumdodecyl sulfate[SDSJ)andchilledonicefor 10min. The cells were disrupted by sonication, and the proteins wereprecipitated with4volumes ofacetone at-20°Cfor30 min(69). Proteinswerepelletedat15,000 xg for 15 min at4°C and resuspended in 160 pl of PBS and 40 pl of 5x loading dye (containing0.0625 M Tris [pH 6.8], 2% SDS,
50%
glycerol,
5% 3-mercaptoethanol, and 0.001%bro-mophenolblue) (43).Thesampleswereheated at70°C for 15
min, boiled for10min,centrifugedto removeanyinsoluble debris, and subjected to electrophoresis on SDS-9%
poly-acrylamide gels. Thebandswere
electrophoretically
trans-ferredovernightontopolyvinylidenedifluorideImmobilon-P
membranes(Millipore, Bedford, Mass.) (in 25mMTris[pH 8.3], 192 mM glycine, and 20% methanol) (68). The filters were incubated in TBST (10 mM Tris [pH 8.0], 150 mM
NaCl, 0.1%Tween 20) containing 20% fetal calfserum and
1%bovine serum albumin for 30min,washed inTBST,and
incubatedfor 3 to 4hwithprimaryrabbita-UL9antibody
(generously
provided byM. D.Challberg)diluted1:1,000in TBST. a-UL9 is a rabbit polyclonal antiserum directed against the 10C-terminal amino acid residues of UL9(53).
Followingseveral washes inTBST, the blot was incubated
for 1 hwithsecondary antibody
(goat
anti-rabbit immuno-globulin G-alkaline phosphatase conjugate; Protoblot sys-tem;Promega,Madison,Wis.).All incubations were doneat roomtemperature withshaking.Alkalinephosphatasecolordevelopmentwascarried out asdescribedpreviously (80).
RESULTS
Functionalassay forengineeredmutants.Site-directed
mu-tagenesiswas used to determine the functional
significance
VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
of variousregionsin the UL9 gene. To facilitate theanalysis of these engineered mutations, we used an assay inwhich
mutant formsof the UL9 proteins could be tested directly
from clonedexpression plasmids. Theassayis basedonthe demonstration that a plasmid bearing an HSV origin of replication
(ons
ororijL)
can be amplified in a transient transfection experiment if all the necessary trans-acting functions are provided. trans-acting replication functionscanbesupplied by superinfection withHSV(65, 66, 77)or
by cotransfection with a setofsevencloned HSVgenes(8, 78). Viralmutants inany one of the seven required replica-tionproteinsareincapableofamplifyinganorigin-containing plasmid.However,cotransfection withaplasmid expressing
the wild-type version of the mutated gene product can
complementthe defect in the HSV mutant.Thus,thedefect
in two host range mutants ofUL9, hr27and
hrl56
(7), could becomplemented by cotransfection withaplasmidexpress-ing the intact UL9 gene (data not
shown).
This assay waspreviously
describedfor usein theanalysis
ofmutations inthe HSV-1 UL5 gene andwastermed theTRC assay(81).
In this study, a UL9 null mutant,
hr94,
was used as the superinfectingvirus intheTRC assay. Inhr94 the UL9genehasbeen
interrupted
withalacZinsertion, andthevirus is completely defective for viral growth and DNA synthesis(47).
Since no UL9protein
was detected in hr94-infectedcells,
hr94 is consideredanullmutant(47).
Theuseof hr94 avoids any possible interference orcomplication
from the mutantform of theUL9proteinspecified
by hr27orhrl56,
both of which
synthesize
stable forms of UL9containing
point mutations
(47).
The other component of theTRCassay is anexpression plasmid for wild-type andmutantforms of UL9. The UL9geneproduct
ininfectedcellsisexpressed
at lowlevelscompared with otherviral geneproducts,suggest-ingthat theendogenous promoterof the UL9 gene may be
relatively inefficient (54).To increase theexpressionofUL9,
we chose to use the promoter region of ICP6
(the
large subunit of the ribonucleotidereductase)
because the ICP6 proteinis an abundantproteininHSV-infected cellsand theICP6promoter isinducibleuponinfection
(81).
p6UL9was constructed such that thecoding region
of the UL9 genewasplaced within the 5' untranslated
region
of the ICP6 gene(47). Expression
of UL9 fromp6UL9
can be induced intransient transfectionexperiments by providing
transactivat-ing proteins
VPl6and/or
ICPOeitherby
cotransfectionorby
superinfection
withhr94(47).
A
typical
TRC assay is shown inFig.
2. Vero cells or2B-11
cells, containing
thewild-type
version of the UL9 gene,were transfectedwithplOO-1,
containing
oiis,
and in some cases alsowiththe UL9expression
clonep6UL9.
At 30 hposttransfection,
cellsweresuperinfected
with KOSorhr94.At 48 h
posttransfection,
totalDNAwasharvested anddigested
with EcoRI alone(lanes a)
orin combination withDpnI
(lanes b).
EcoRI cleaves plOO-1 once,generating
afragment of 2.8 kb. DpnI recognizes only those cleavage
sites which have been
methylated
bythedammethylation
system of E. coli. Thus, plasmid DNA which has been
propagated in E. coliissensitive toDpnI (Fig. 2, lane Sb),
but DNA which has replicated in a mammalian cell is
expectedtobeDpnIresistant. When Vero cells were trans-fected with
plOO-i
and superinfected with KOS, a DpnI-resistantband of 2.8 kb representinglinearizedplOO-i
wasobserved, indicating
thatKOScansupport thereplicationofthis
origin-containing
plasmid(Fig.
2,lane2b).
The lackofa 2.8-kbDpnI-resistantband in cells transfected withplOO-i
andsuperinfected
with hr94 indicates thatthismutant cannot support thereplicationofplOO-i
(Fig. 2,lanelb).TheDpnI
1 2 3 4
a bab a b a b
5 a b
-*-p6U L9
W smISSums * -_--p100-1
'S
FIG. 2. TRCassay. The TRC assay was performed with either Vero or2B-11cellsasdescribed in Materials and Methods. At 48 h after transfection, total cellular DNA was isolated. DNA was digested with eitherEcoRI alone (lanes a)orEcoRl plus DpnI (lanes b); thus, every sample is represented byapair of lanes (a and b). Lanes1 to4contain 2pgof totalcellularDNAharvested from the TRC assayasfollows: 1, transfection of VerocellswithplOO-iand superinfection withhr94;2, transfection ofVerocells withplOO-i and superinfection with KOS; 3, transfection of 2B-11 cells with
plOO-i and superinfection with hr94; 4, transfection of Vero cells with plOO-i and p6UL9 and superinfection with hr94. Lane 5 contains 5 ng ofplOO-icontainingHSV-1orispropagated in E. coli. After restriction enzyme digestion, the samples were subjectedto 0.8% agarosegel electrophoresis and blottedontoGeneScreenPlus membrane. The blot was hybridized with 32P-labeled Bluescribe plasmid. The positions of the linearized plOO-1 and the 6.3-kb fragment from p6UL9areindicated.
digestion fragments representing input plasmidare not seen inthisexposure but are seen in longerexposures(datanot
shown). If a functional UL9 product is supplied either by cotransfection with theUL9expression clone p6UL9
(lane
4a and b) or by transfecting 2B11 cells which contain a
wild-typeversionofthe UL9 gene(lane3aandb), the defect in hr94 iscomplemented.The6.5-kbband seenwithEcoRI
digestion (lane 4a) represents input p6UL9 plasmid. This plasmid lacks an HSVorigin and cannotreplicate in Vero
cells; therefore, it is sensitive toDpnI digestion (lane 4b).
The 2.8-kbDpnI-resistantbands markedpl00-1in lanes 3b and 4bwhicharepositiveforplasmid amplification migrate
as adoublet. This doublet isroutinely observedwhenhr94 is complemented byafunctionalUL9geneproduct expressed
either from the cotransfected plasmid p6UL9 (lane 4b) or from theintegrated copiesofp6UL9 residentin2B-11 cells (lane 3b); it is not seen in KOS-infected cells(lane 2b). We
speculate that this may be due to an alteration in the conformation of
replicating plasmids
inthe presence ofan excessof UL9protein.
Insummary, the resultspresented
in Fig. 2 indicate that theTRC assay can beused to test the function of UL9 protein expressed from the expression plasmid p6UL9. This assay provides a convenient methodfor testing the function of UL9 protein expressed from
plasmids bearing wild-typeand mutant versions of the UL9 gene.
At least five of the six conserved helicase motifs inUL9 are
importantforfunction. Inthis report, the functional
signifi-canceof the six conservedputative helicase motifs in UL9
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.335.529.72.249.2]HELICASE MOTIFS OF UL9 6739
was examinedby the introduction ofengineered mutations
into themostconserved residue(s) in eachmotif.
Motifs I and H. Oligonucleotide mutagenesis was per-formed on single-stranded DNA from p6UL9 as described in Materials and Methods. Table 1 lists the oligonucleotides used togenerateeachmutation. MotifsI and II are very well conserved among enzymeswhich utilize ATP and/or GTP
(71)(see Table 2). Motif I (also called the "A" site) contain-ing a sequence (G/AX4GKS/T) contains a flexible loop between a beta strand and an alpha helix and is believed to be involved in the binding of the
PPi
moiety of nucleosidetriphosphate (NTP). Through conformational changes in-ducedby ATPbinding, this loop would control access to the
nucleotide-binding domain (34). The conserved lysine resi-dueis thoughttointeractdirectly with one of thephosphate
groups ofthe ATP molecule and therefore be essential for ATPaseactivity.Thislysine is invariant among all members ofsuperfamily 2 (Table 2). To determine the importance of thislysinein UL9, two separate mutations were made at the residue: a conservative change to arginine (K87R) and a change to the small nonpolar amino acid alanine (K87A). Both mutations were constructed to create an NcoI site without altering the UL9 sequence (Table 1). The motif I mutations were tested for their ability tocomplementhr94 in the TRC assay as described above. As shown in Fig. 3, wild-type UL9(p6UL9)can complement hr94 in its ability to support thereplication ofplO0-1in Vero cells (lanemarked
p6UL9); however, mutant K87R and K87A fail to do so
(lanesasmarked).The smallDpnI-sensitive fragments(lanes
binFig.3)represent inputDNA. Theloss of UL9function
in K87R and K87A demonstrates the importance of the
conserved lysinein motif I.
MotifIIcontains one or two negatively charged residue(s)
(D or DE) at the COOH terminus of a beta strand; the
negativelychargedresidue(s)isbelieved to interact with the
Mg2+ associated with the i or y phosphate of a purine nucleotidesubstrate (motif II) (25, 42). Inmolecules whose
structure is known, motif II is believed to be close to the
glycine-rich flexible loop (motif I) and to form the homing
pocket for the hydrophobic adenine moiety of the ATP molecule (3).UL9 and allmembers ofsuperfamily2contain two charged residues, an aspartic acid followed by a
glu-tamic acid (DE) in this motif (Table 2). We targeted this motifbyreplacingtheglutamicacidresidueat position 175 with an alanine (E175A), leaving only one charged amino acidresidue insteadof two(Table 2). The plasmid carrying mutantE175A wasunabletocomplement hr94in the repli-cation assay(Fig.3).
Motifsmto VI.The four other motifsconserved among all members of this superfamily of helicases have no known function at present. Our goal was to assess the functional
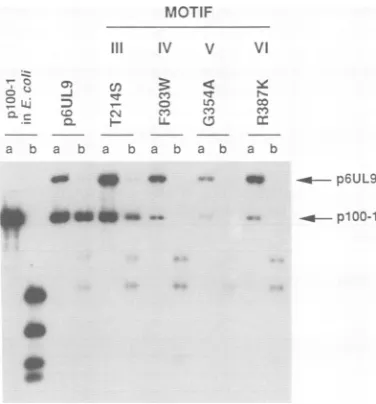
significance of these conserved residues; thus, we have introduced single aminoacid alterations intothemost con-servedresidues in each. Thus, mutations T214S (motif III), F303W(motifIV), G354A (motif V), and R387K (motif VI) were introduced into p6UL9 (Table 2). In each case, a
restriction site was either created or abolished (Table 1).
Figure 4 shows that mutants in motifs IV, V, and VI are unable to complement hr94 in the plasmid amplification
assay, while the T-to-S mutation in motif III resulted in levels of replication which were compromised compared with those seen with wild-typep6UL9. The partial ability of T214S to complement is perhaps not surprising since one other member ofSF2, a vaccinia virus gene product, has a serine in thisposition (32).Tofurtherinvestigatethe impor-tance of motif III, two additional mutations were
con-structed: one in which the threonine at position 214 was
changed to analanine (T214A, also referred to as IIIa) and one in which an alanine at position 213 was changed to a serine (A213S, alsoreferredto asIllb).Figure 5 shows that, as seen above, T214S ispartially compromised for comple-mentation as is A213S; however, mutant T214A totally
abolishes plasmid amplification in this assay. The partial
replication activity of two of the mutations in motif III
(T214SandA213S) were of interest since these represent the first mutations to give a partial phenotype in this type of assay. Since the TRC assay is not quantitative, we used a TC assay to test the ability of the plasmids containing motif mutations to complementhr94 for virusproduction; in this assay, yields of hr94 were measured by plaque assay. Table 3indicates that the results of this assay are quiteconsistent
with the results of the TRC assay. The complementation
index (CI) with no UL9 plasmid isexpectedto be 1. Plasmid
p6UL9-119b bearing wild-type UL9 is capable of comple-menting hr94 (CI = 23). Mutant plasmids which fail to
complement hr94 for plasmid amplification (K87A, K87R, E175A, T214A, F303W, G354A, and R387K) give values very close to 1.0 in the TC assay. The two mutants which
partiallycomplemented hr94 in the TRC assay (T214S and
A213S) generated intermediate but low CIs. These results indicate not only thatresults from the TRC and TC assays are consistent, but also that all threemutationsin motif III impair the function of UL9 to various degrees ofseverity.
The amino acid substitutions which result in loss of
functionof UL9replication activity could, intheory,be due to two different types of changes: a change in protein
conformation or a change in a residue that is involved in
function. Changes in structure often affect the stability of
proteins (55). Toassess whethermutations resultingin loss offunction induce gross loss of stability, UL9 protein in cells
transiently transfectedwith wild-type andmutant versions of UL9 was examined. Transfected cells were induced by infection with
hr94,
and extracts were subjected to SDSpolyacrylamide gel electrophoresis and immunoblotting as
described in Materials and Methods. Transfection with p6UL9 followed by superinfection with hr94 gives higher yields of protein than cells infected with wild-type virus
KOS(Fig. 6).Consideringthattransfectionsusually result in lessthan 5% of the cells taking up DNA and infections can result in100%infection,it isclear that the ICP6promoteris
efficient.Figure 6 alsodemonstratesthat the motifmutations shown, with the exception ofG354A in motif V, result in levels ofUL9comparabletothose in p6UL9, indicating that no loss of stability can be observed this way. Of the mutations in motif III,
T214S,
which results in partial activity in both theTRC and TC assays, is shown in Fig. 6 to induceapproximately
wild-type levels of UL9 protein; mu-tant T214A, which abolished activity in the TRC and TC assays, was not able to induce a stableprotein,asjudged by the absence ofdetectable UL9 protein, and mutant A213S, which alsoresults in partial activity in both assays, was able to induce wild-type levels of UL9 (data not shown). Theabsenceofdetectable UL9 protein inT214A (motif III) and
G354A(motif V) indicates that these mutations very likely confer conformational changes on theprotein which result in instability.
Insertion anddeletion mutations. Althoughtheexistenceof
conserved sequence motifs provides a useful target for mutagenesis, it is also of interest to determine whether other regions of the gene are essential for its function. To this end, a series of linker insertion mutations were introduced into the gene roughly every 100 residues along the UL9 gene. VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
0)
0
0)
bo 00
q
0% 0)
.0
0 02 02 0) 0) 02
0)
C-IO '0 0L)
00
0
..g
0
It 0
2
0.4
_-1!
0->=
0-4.
4-0) C) 0 0) 0' 0) 00
CZ
0.°
0=I
0' 0)
0,0
CA.2
C)00)
A)
00(A4.
0)
0'
0.)
.'
(A0
0
0)
A)
00
0.2
ci
03
00000Cy
GOa o
t4HI- ,. H 00
c
0H0n0n0PH O 00SO 0000
coi c o crooo
r4AWN00
,It
0N QS f4 0
Hpq "N
0C100 r
00
'It 0t-U)
t-ON Noo Noo en
Pq"itt WnPen H0
H
- P
P4H.
00000
00W) 0
0 00
PA
-noo
enenone
t-+~~~
+ p
+ a
+ E
+ -4
0
00
+ E
Y'S~ 00,4n
+ 0>m
0 0
+ 0
000 0
+
0
E-400
0
0
0
0 §,
o2
-00 0
0C '
.) -.0 '0
.0 o
o -0c 0
D '0 '0 =
0) 0
82 i
C)
'0
,_
0e
,~Y
0 .0
o 0. .2 0) 0L Q 1o 0 0
0
00E4
E4 P* r3
0
E4
0
+-+4
03
E-cx C
Eq <
on November 9, 2019 by guest
http://jvi.asm.org/
HELICASE MOTIFS OF UL9 6741
virus:
6 D
RlMotif
I Motif11 0cr,
K87A K87R E175A p6UL9
a b a b a b a b a b
E_ _ _wS US p6UL9
w~~U 4,, * _ p100-i
0
FIG. 3. TRC assayof UL9
proteins
with site-directed mutations in motifsIandIL.The TRC assaywasperformedwithVerocellsasdescribedinMaterials and Methods and in the legendtoFig.2. In each sample, p100-i was used in transfection either alone or in combination with p6UL9 containing
wild-type
UL9 (lane markedp6UL9)orvariousmutant constructsofp6UL9asmarked. hr94was
used forsuperinfection.
hr94 KOS
a,
-4
C& Ill IIIa IIIb
a b a b a b a b a b a b
_
_
aft ot O0
._
a b
-_*--p6UL9
FIG. 5. TRC assayof UL9proteins with additional mutationsin motif III. See legend to Fig. 2 for details of the assay. In each sample,pl00-1wasused in transfection either aloneorin combina-tionwithp6UL9containing wild-typeUL9(lane markedp6UL9)or
variousmutant constructsof p6UL9.Fromthe left, pairs underthe heading hr94 contain wild-type, mutant III (T214S), mutant lIla (T214A),mutantIIIb(A213S), andnoUL9expression plasmid, all superinfected with hr94 as indicated. The pair under the heading KOS represents cells transfected with pl00-1 alone and superin-fectedwith KOS, and the lastpair onthe rightrepresents pl00-1 grownin E. coli.
Specificoligonucleotidesweresynthesizedfor each insertion mutation designed to introduce a unique NotI site and to
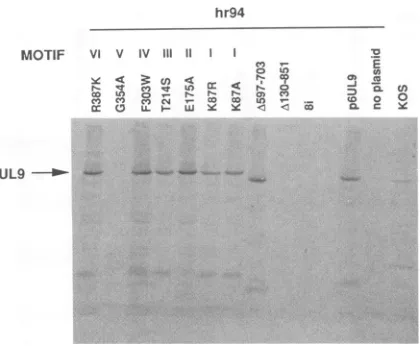
resultin the insertion oftwoalanine residues(Table1).Two alanineswerechoseninanattempttointroducefairly subtle alterationswhich wouldnotbeexpectedtogrossly alter the structure of the UL9 protein. These linker insertion muta-tions havegeneratedvariable results in the TRC assay(Fig.
7 and 8). Three insertions
(ii,
6i, and 7i) are tolerated, asMOTIF
III IV V
-4
a b
cn co
a b
a b
L)
a b
-wa_.
*
Pa e4VI
a b
_ -- p6UL9
- _-.*- pOO-i
judged by the appearance of wild-type levels of
DpnI-resistant plOO-1. Five insertions(2i, 3i, 4i,5i,and8i)result in little or no DpnI-resistant DNA at the position of plOO-1. Faint bandsatthepositionofDpnI-resistant plOO-1in2i, 3i,
4i, and Si(Fig. 7) arebelieved torepresentpartial digestion
products, since they are not seenin duplicate experiments.
Toconfirm that thesemutationsresulted in loss offunction,
the TC assay described above was performed. Table 4 demonstrates that the wild type andinsertions li, 6i and 7i
were able to complement theyieldsofhr94 efficiently; the
otherfive insertionswereunable todo so, thereby
confirm-ing the results ofthe TRCassay. Sincethe insertion muta-tionsare outside the conserved helicase motifs, these data indicate that otherregionsof theproteinarealsoimportant.
[image:7.612.329.557.74.235.2]TheN-terminalportionof the genecontainingtheconserved
TABLE 3. Complementation ofhr94 withplasmidscontaining motif mutations inUL9"
Plasmid CIb
None... 1.0
p6UL9-119b... 23.0
K87A... 1.0
K87R... 1.0
E175A... 1.1
T214S(III)... ... 2.9
T214A(IIIa)... ... 1.1
A213S(IIIb)... ... 4.5
F303W... 0.8
G354A... 1.3
[image:7.612.89.277.75.275.2]R387K... 0.9 FIG. 4. TRCassayofUL9proteinswithsite-directed mutations
in motifs III, IV, V, and VI. See the legend to Fig. 2 for the
description of the assay. In each sample, pl00-1 was used in transfection either alone orincombination withp6UL9 containing wild-type UL9 (lanemarkedp6UL9)orvariousmutant constructsof
p6UL9asmarked. hr94wasused forsuperinfection.
aVero cellsweretransfected with theindicatedplasmidand then superin-fected with the UL9 lacZ insertionmutanthr94.Theprogenyof the
trans-fected-infected cells was assayed for PFUs on S22 or 2B-11 cells. This
experimentwasrepeatedthreetimes,and similar resultswereobtainedeach
time.
bCI forhr94 is measuredasthe PFUs of hr94 from cultures transfectedwith
theindicatedplasmid/PFUsofhr94 frommock-transfectedcultures.
Z-o
O X;
0..
a
a b
I
VOL. 66,1992on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.90.278.460.662.2]hr94
MOTIF VI V IV III 11
CO e)
~~~~~~ N~~~~~~o L
ya e' 0 < N I?
cL) 0 ;; r- ra as co
Cl:O .-1 JWi v G
-o
Di cL cn U) a,
0. E
UL9-- _
-oD
6u
CL: 6i 7i 8i p6UL9
a b a b a b a b a b
0 _ W i Nop6UL9p -_- ab
f
FIG. 6. Detection of wild-type and mutant UL9 proteins in transient transfection by immunoblot with a-UL9. Wild-type or mutant versions of p6UL9 (as marked) (12 ±g) were used to transfectVerocellswhich werethensuperinfected with hr94.From lefttoright,mutationsinmotifs VI, V, IV, III,andII;twomotifI mutations; two deletions (A597-703 and A130-851); insertion 8i; wild-type p6UL9; and no plasmidareshown.Inthelane labeled no plasmid,12p.gofsalmon spermDNA wasused.Inthe farright lane, KOS-infectedcellextracts are shown forcomparison. Transfected cellswereprocessedforimmunoblotasdescribedinMaterials and Methods.
helicase motifs appears to be slightly more sensitive to insertions than the Cterminus. Twodeletionsremoving721 and 106 residues (A130-851 and A597-703, respectively)
which also abolish the replication ability of theseplasmids (Fig. 7 and 8, lanesasmarked)werealsoconstructed.
To assess whether the insertion and deletion mutations
described above altertheconformation of the UL9protein drastically enoughto affect stability, Vero cellswere
tran-siently transfected with each construct and superinfected
with hr94 to induce the ICP6 promoterasdescribed above. Extractsweresubjectedtoelectrophoresisand immunoblot-tingasdescribedinMaterialsand Methods. Figures6and9 showthat allinsertionmutations, with the exceptionof8i,
O
6u
a c
a b
CD
6 -J
1i 2i 3j 4i Si .<
a b a b a b a b a b a b a b
! p6UL9
i 4 0 -.---p100-1
0.
- *k
wl*. Dpnl
sensitive
bands
FIG. 7. TRC assayof UL9proteins with insertions li, 2i, 3i, 4i, and 5i and deletion mutation A130-851. See legend to Fig. 2 for details.Pairs ofsamplesaremarked with thenames ofthe p6UL9 constructsused in transient transfections. Smallfragments resulting fromDpnIdigestionaremarked.
FIG. 8. TRC assay of UL9proteins with insertions 6i, 7i, and 8i and deletion mutation A597-703. See legend toFig. 2 for details. Pairs ofsamplesaremarked with thenames ofthep6UL9constructs used in transient transfections.
were capable of producing astable protein. Deletion
A597-703 produced aversion slightly smaller than full length as
expected from the size of the deletion. No band is seen in
A130-851asexpected since thepredictedUL9 geneproduct
wouldnotreactwiththeantibodydirectedtotheCterminus (Fig. 6). In summary, these results suggest that the loss of
functionseeninmostof themutantconstructsismostlikely duetochangesin residues involved in UL9 function rather than disruptionswhich causeinstability; however, the
mu-tationsT214Aand G354A(inmotifs III and V,respectively)
and theinsertionat8iall apparentlyalterthestability of the
resultingUL9 geneproduct.
DISCUSSION
Inthis report, wedescribe theuse ofaninvivo
comple-mentation assay to determine the replication activity of a series ofmutationsin the UL9 gene. The TRC assay is based on the ability of wild-type and mutant forms of UL9
ex-pressed from an expression plasmid to complement a null
[image:8.612.74.284.71.244.2]mutationinUL9foramplificationofanHSV origin-contain-ing plasmid. Of19mutantstested, 14abolished the replica-tionabilityof UL9 and 5weretolerated.
TABLE 4. Complementation of
hr94
withplasmids containing insertion mutations inUL9aPlasmid Cjb
None... 1.0 p6UL9-119b... 106
p6UL9-119b-li... 18
p6UL9-119b-2i... 0.6
p6UL9-119b-3i... 0.9
p6UL9-119b-4i... 1.7
p6UL9-119b-5i... 0.5
p6UL9-119b-6i... 131
p6UL9-119b-7i... 88
p6UL9-119b-8i... 1.3 aVerocellsweretransfected with the indicatedplasmidand then superin-fectedwith theUL9 lacZ insertionmutanthr94.Theprogenyofthe
trans-fected-infectedcellswasassayed forPFUonS22or2B-11 cells.
bCIfor hr94 is measuredas
PFUs
of hr94 from cultures transfected with the indicatedplasmid/PFUsofhr94frommock-transfectedcultured.N
r--42 p6UL9
a b a b
*0Pm1I
lo
-p100-1 -
-n
_~~~s
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.322.547.72.235.2] [image:8.612.68.293.517.670.2]HELICASE MOTIFS OF UL9 6743
KOS
0 0 c C o
hr94
2i 3i 41 51 6i 71
_-- UL9
1 2 3 4 5 6 7 8 9 10
FIG. 9. Detection of wild-type and mutant UL9 proteins in transient transfection by immunoblot with oa-UL9. Wild-type and
mutantversions of UL9areshownasdescribedin thelegendtoFig. 6.KOS-infectedcell extractis shown inlane1.Fromlefttoright (lanes2to10, respectively)areextractsfromcellstransfected with
the indicated plasmid and superinfected with hr94: no plasmid,
wild-typep6UL9 (wt), li,2i,3i, 4i, Si, 6i, and 7i.
Five of six conserved helicase domains as well as other
regions of the UL9 gene are important for function. The
predicted proteinsequenceindicates thatUL9 containsaset ofmotifs conserved among a large number of known and
putative helicases from E. coli, yeasts, eukaryotes, and viruses. These six motifs are located within the first 400
amino acid residues of the UL9 gene (Fig. 1). Certain
residues within these motifsarewellconserved in all
mem-bers ofSF2(Table 2); however, the sequence similarity is
evenmore striking within other UL9 homologs within the
herpesvirus family. Table 5 shows the sequences of the
conserved motifs in varicella-zoster virusgene51(15) and in the humanherpesvirustype6geneCH6R (14). In thispaper,
wehave addressed the functionalsignificance of these
con-servedregions in the UL9geneby introducing single amino
acidsubstitutions into themostconservedresidues of each motif. The results suggest that at least five of the six conserved motifs are importanttothefunction of theUL9 protein in viral DNA replication (see below).
Theresults of insertion and deletionmutagenesis indicate that several other regions outside the conserved helicase motifsarealsorequiredforreplication activity. Insertions 2i,
3i,4i, Si, and 8i all abolished the ability of UL9tofunction in
the TRC assay. Although 8i is probably nonfunctional
be-cause of instability of themutantprotein, itisunclearwhy the other four mutants have abolished activity. None of these mutations coincide exactly with the helicase motifs themselves, although 2i and 3iarenearby motifs II and IV,
respectively. It is possible that the spacing between the various helicase motifs in UL9 is sensitive to insertion because ofrigid requirements in the surrounding secondary and tertiary structures. Another possibility is that the
re-gions outside the motifs which are sensitive to insertion representfunctional regions for other activities displayed by UL9, suchas protein-protein interaction, nuclear
localiza-tion, or a previously unidentified function ofUL9. UL9 is
known to contain a putative leucine zipper, an amino acid sequence motif common to several DNA-binding proteins andbelievedtosignalaregion ofprotein-proteininteractions
(44). The leucine zipper overlaps motif II in the putative helicase domain. Preliminary results with mutations with alterations in the leucines of the leucine zippersuggestthat theseleucines areessential for UL9 function (61). Whether
the leucinezipperrepresents a sitefor dimerization orfor
interactions with other members of the replication complex remainstobe elucidated. Weiretal.demonstrated that the carboxy-terminal 317 amino acids contain all the information
necessaryfor origin-specific DNA binding (73). The
DNA-binding regionwas subsequentlynarrowedto the carboxy-terminal 269 amino acids(residues 564to832) (18). Insertion mutations 6i and 7i at residues 597 and 703, respectively,
weretolerated for UL9 function, but the deletion spanning
thisregion(A597-703) isnottolerated. Since A597-703
spec-ifiesastable protein,weconclude thatanessential region is
located within the region defined by residues 597 and 703. Wespeculate that within this regionaresignals which could
be involved in specific DNA binding. Further biochemical characterization of existingmutants and isolation of addi-tional mutationsacross theUL9genewillberequired fora
morecomplete understanding of the domainstructureof the multifunctional UL9 protein.
Relationship of UL9toothersuperfamilies of helicases. UL9 andother members ofSF2are related tobutdistinct from another superfamily of putative helicases (superfamily 1) characterized bya different setofprokaryotic, eukaryotic, and viral helicases(including HSV-1 UL5 and E. coligenes
uvrD, rep, recB, and recD) (36, 37, 81). The conserved
helicasemotifs in UL5are dispersed throughout the entire
gene(31, 81), whereas in UL9 theyare confinedtothefirst 400 amino acids (see Fig. 1). Many members of SF2appear
tobe multidomain proteins in which the putative helicase motifs are confined to the N-terminal portion ofthe mole-cule. Forinstance, inmanyRNA virusessuchasthe potex-,
tobamo-, flavi-, pesti-, alpha-, and potyviruses, the SF2 family members are multidomain proteins in which the
TABLE 5. Alignmentofputative UL9 homologs in HSV-1, varicella-zoster virus, and human herpesvirus type6
Gene MotifI MotifII MotifIII MotifIV MotifV MotifVI
VVRAPMG8GKTTAL LMDVLVLDEVXBTL IAMDATANAQLVD NICIPB8TV8TAE VVIYTTVVTVGL8FD GPDMV8VYQ8LGBVRTLRK
VVRAPMGBOKTTAL IDBYDVLILDEVKSVI IAMDATVN8QFID NICIF88TL8F8B VLVTITVVTVGLBFD GPDXVBVYQ8LGRVRLLLL
LVRAAMG8GKTTAL TENYDVLILDBIM8II IAMDATLTRHVVZ KLCLFCSTVLAAE VVIYTBVVTVGLBFB GPDLV8VFQBIGRVRBVID HSV UL9
VZV 51
HHV-6CH6R
Consensussequence *VBA*MGSGKTTAL *YDVL*LDE*1S * IAMDAT **h** **C*F*ST** *E V**YT*VvTVGL8F* GPDM8V*QB*GBVB * IAlignment oftheHSV-1origin-binding protein UL9, thevaricella-zostervirus origin-binding protein gene 51(VZV 51) (15)and the putativehuman
herpesvirustype6(HHV-6) origin-binding protein encoded by the open reading frame CH6R (14). Motifsarethosepredictedby Gorbalenyaetal. (31). h representshydrophobicresidues(FYIVLM);*referstoresiduessharedbytwoof thethreegenes.
VOL.66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.69.296.60.279.2]conservedregionofthe
putative
helicases is localizedtothe N terminus and the C-terminalportion
containsregions
conserved among polymerase molecules
(32).
The demon-stration thatatleastfiveofthesixputative
helicasedomains located within the N-terminal 400 residues of UL9 are importantfor UL9function inreplication
oforigin-contain-ing plasmidsstronglysuggeststhatthis regionisinvolved in UL9 helicase activity. Whether this region makes up a
discrete domain which is sufficient for helicase
activity
remains to be determined. As described
above,
theDNA-binding regionhas been locatedto thecarboxy-terminal269
aminoacids(residues564to
832) (18).
Takentogether,
theseresults suggestthat UL9 may containatleasttwofunctional domains, ahelicase domain at the N terminus anda
DNA-binding domain atthe C terminus.
Roles of motifsI toVI.Forsome
time,
motifs I and IIhave been recognized as important forNTPbinding
onthe basisofnotonly nuclear
magnetic
resonanceandX-ray
diffractiondata(25, 42)but also theresultsofsite-directed
mutagenesis
aimed at one or more of the conserved residues. For instance, mutationalanalysisofmotif I in otherATP-binding
proteins showing ATPase
activity
indicates thatsubstitu-tions at the conserved
lysine
residuegenerally destroy
the ATPase activity and in some but not all cases leave ATPbinding intact,
depending
on the nature of thechanged
residue (3). Forinstance, mutation oftheinvariant
lysine
toarginine
inmotifIof theyeastexcisionrepair protein
RAD3(amemberof
SF2)
resultedinaprotein
which could still bind ATP but was defective inhydrolysis
(67).
Some motif Imutations appear to affect
binding
of NTPs per se; forinstance, athreonine-to-serine
replacement
in motifIofthe HSVthymidinekinaseexhibitedanalteredKm forthymidine
and ATP (46). Thus, motifIhas beenstrongly
implicated
inbinding and
hydrolysis
of NTPs in several knownNTP-binding proteins. Our studies are in agreementwith those
previous studies in that the
replacement
of the invariantlysinein motif I of UL9 witheitheran
arginine
or analaninetotally abolishes the
ability
of the mutant form of UL9 to carryoutin vivocomplementation
of HSVDNAreplication.
Motif II has been less well
studied; however,
in two studies, substitution of the invariantAsp
residue in boththe E. coli Fl-ATPase andathermophilic
ATPaseresulted inagreater-than-90%
loss of ATPaseactivity
(1,
79).
InRAD3,
aGlu-to-Lys substitution within motif II resulted in extreme UV
sensitivity
and anincreased rate ofspontaneous muta-tion,indicatingthat thismotifisimportant
forexcisionrepair
and perhapspostreplicative
mismatch correction(62).
InUL9,motif II containsaDE
pair
andsubstitutionof theGlu byanAla(E175A)
wassufficientto abolish UL9 function in the in vivo assay,confirming
theimportance
of this motif.Theimportanceof motifs IIItoVIwithinSF2hasreceived little attention to date, and few mutations have been
re-ported.
Toourknowledge,
theonly
other memberofSF2 in which mutations in motifs III to VI have beenreported
is RAD3.Amongeleven RAD3mutantswhose mutations have beenmapped
whicheitherseverely
ormoderately
affectUVsensitivity, the
positions
ofall elevencorrespond
exactly
to oneof theconservedmotifs asdescribedby
Gorbalenya
et al.(32) (twoin motif1,twoinmotifII,three in motifIII,
two in motif IV, two in motif V, and one in motifVI);
thepositions of the UV-sensitive mutations are indicated in Table 2asunderlinedresidues
(52, 62).
Severalof theseare also associated with an increased spontaneous mutationrate.These results indicate the functional
importance
of theconservedhelicasemotifs of SF2. The mutations
presented
inthis report represent thesecond mutationalanalysis
ofamember of SF2 and suggest that
putative
helicase motifsI,
II,
IV,
and VIareabsolutely
essentialfor UL9function.Theimportance
of motif IVisalsoindicatedby
thefactthattwospontaneously
occurring
mutantsin theUL9gene(hr27
andhr156)
which abolish UL9 function contain identicalsingle
base
pair
substitutionsatcodons309and311 withinmotifIV(see
Table2).
Two mutations in motif III exhibit low butpartial
activity.
This is the first time we have observedpartial activity
in this type of assay, and it will be ofconsiderableinterestto introduce thesemutationsback into the virus and observe the
biological
activity
of theresulting
mutants.Onemutationin motif III
(T214A)
andthemutation inmotifV(G354A)
result intotal loss offunction,
butthesetwomutations alsoresult in
instability
ofUL9;
therefore,
itis not clear whether the actual residues are essential or
whetherthese
regions
areverysensitivetoconformation.It issomewhatsurprising
thatrelatively
conservativesubstitu-tions ofAlafor Thr inmotifIII and Alafor
Gly
inmotifVresult in
protein
instability.
These resultsimply
that the structure of UL9may be very sensitive to certain typesof alterations. Takentogether,
ourresultsindicate thatatleastfive of the sixhelicase motifs are
important
for UL9 func-tion.Thus,
although
unwinding
ofduplex
DNAsequencescontaining
the HSVorigin
ofreplication
has not beendemonstrated with
purified
UL9protein
(24),
thegenetic
data inthisreport indicate
that,
infact,
the helicase motifs areimportant
for itsfunction.Itispossible
that otherviralorcellularfactors will be
required
inorder toobserveduplex
unwinding by
UL9.Our
goal
was to determine the roles of the residues in motifs ItoVI in the overallfunctionof UL9. Inadditionto anNTP-binding fold,
it isanticipated
thata helicasemight
contain several other functionalregions, including
onewhich contacts thenucleic acid substrate. Motif VI ismade up of severalbasicresidues,
and it istempting
tospeculate
thatthis
region
may be involved ineithernuclearlocalizationornonspecific
DNAbinding
orboth. UL9 has beenexpressed
in a functional form in E.
coli
(48),
and the mutationsdescribed in this report will be transferred to the E.
coli
expression
system and tested forvarious biochemicalactiv-ities,
including
NTPbinding,
DNA-dependent
NTPase,
nu-clear
localization, ability
todimerize,
specific
and nonspe-cificDNA-binding,
andhelicaseactivities.Inthisreport,wehave initiateda structure-function
analysis
of the UL9 gene and laid thegroundwork
foramuchmoredetailedanalysis
offunctionaland
biological
aspects ofUL9,
which will leadto the identification of domainsresponsible
for its variousactivities.Itis
anticipated
that the isolation ofmutantswhich can separate thesefunctional domains will facilitate efforts to elucidate theprecise
role of the UL9protein
in the initiation of DNAreplication.
ACKNOWLEDGMENTS
We aregrateful to M.
Challberg
forproviding
origin-containing
plasmidsandantiseratoUL9. Wegratefully acknowledge contribu-tionstothisprojectatitsinitialstages fromE.Carmichael.Wealso thankE. Carmichael and D. Dorskyfor
helpful
commentsonthemanuscript.
Thisinvestigationwassupported byPublic Health Servicegrant AI21747 and American Heart Association
grant-in-aid
880731. S.K.W. is therecipientofanAmericanHeart Association-Genen-tech Established InvestigatorAward.REFERENCES
1. Al-Shawi,M.K.,D.Parsonage,and A. E.Senior. 1988. Directed mutagenesis of the strongly conserved aspartate 242 in the
on November 9, 2019 by guest
http://jvi.asm.org/
HELICASE MOTIFS OF UL9 6745 P-subunit ofEscherichia coli proton-ATPase. J. Biol. Chem.
263:19633-19639.
2. Ausubel,F. M., R.Brent, R. E. Kingston,D. D.Moore, J.G.
Seidman, J. A. Smith, and K. Struhl (ed.). 1990. Current protocolsin molecularbiology.JohnWiley&Sons,New York. 3.
Azzaria,
M.,E.Schurr,andP.Gros.1989. Discretemutations introducedin thepredicted nucleotide-bindingsitesofthemdrl geneabolish itsability
toconfermultidrugresistance.Mol.Cell. Biol.9:5289-5297.4.
Bramhill,
D.,and A.Kornberg. 1988. Amodelfor initiationatorigins
ofDNAreplication. Cell 54:915-918.5. Bruckner,R.C.,J. J.Crute,M. S.Dodson,andI. R.Lehman. 1991.The
herpes simplex
virus1origin binding protein:aDNA helicase.J. Biol.Chem. 266:2669-2674.6. Carmichael, E. P., M. J. Kosovsky, and S. K. Weller. 1988. Isolation and characterization ofherpes simplexvirustype 1 host rangemutantsdefectivein viralDNAsynthesis.J. Virol. 62:91-99.
7.
Carmichael,
E.P.,andS. K.Weller.1989.Herpes simplexvirus type 1DNA synthesis requiresthe productoftheUL8 gene: isolationandcharacterizationofanICP6::lacZinsertionmuta-tion.J.Virol.63:591-599.
8.
Challberg,
M.D.1986.A methodforidentifyingthe viralgenesrequired
for herpesvirus DNAreplication. Proc. Natl. Acad. Sci. USA 83:9094-9098.9.
Chartrand,
P., C.S. Crumpacker, P. A. Schaffer,and N. M. Wilkie.1980.Physical
andgenetic
analysisof theherpessim-plex
virusDNApolymeraselocus.Virology103:311-326. 10.Chen,
E.Y.,and P. H. Seeburg.1985. Supercoil sequencing:afast and
simple
method forsequencingDNA. DNA 4:165-170. 11.Coen,
D.M.,D.P.Aschman,P.T.Gelep,M.J.Retondo,S. K.Weller,
andP. A. Schaffer. 1984.Fine mappingandmolecularcloning
ofmutations in theherpes simplexvirusDNApolymer-aselocus. J. Virol. 49:236-247.
12.
Conley,
A.J.,D. M.Knipe, P. C.Jones,andB.Roizman. 1981. Moleculargenetics
ofherpes
simplexvirus.VII. Characteriza-tion of atemperature-sensitive
mutant produced by in vitromutagenesis
anddefective inDNAsynthesisand accumulation of-ypolypeptides.
J.Virol. 37:191-206.13. Crute, J. J., T. Tsurumi, L. Zhu, S. K.Weller, P. D. Olivo, M. D. Challberg, E. S. Mocarski, and I. R. Lehman. 1989.
Herpes simplex
virus 1helicase-primase:
a complexof threeherpes-encoded
geneproducts.
Proc. Natl. Acad. Sci. USA 86:2186-2189.14. Dambaugh,T. R. 1992.Personal communication.
15. Davison, A. J., and J. E. Scott. 1986. The complete DNA sequenceofvaricella-zoster virus. J. Gen. Virol. 67:1759-1816. 16. Dean, F. B., P. Bullock, Y. Murakami, C. R. Wobbe, L.
Weissbach,
andJ.Hurwitz.1987. Simian virus 40(SV40)DNAreplication:
SV40large
TantigenunwindsDNAcontainingthe SV40origin
ofreplication.
Proc. Natl.Acad. Sci. USA 84:16-20.17. Dean, F. B., M. Dodson, H. Echols, and J. Hurwitz. 1987.
ATP-dependent
formationofaspecialized nucleoprotein struc-tureby
simian virus40(SV40) large
tumorantigenatthe SV40replication origin.
Proc. Natl.Acad. Sci.USA84:8981-8985. 18. Deb,S.,
and S. P. Deb. 1991. A269-amino-acid segment withapseudo-leucine zipper
and a helix-turn motif codes for thesequence-specific DNA-binding
domain ofherpes simplexvirus type1origin-binding protein.
J. Virol.65:2829-2838.19.
Dodson,
M.,F. B.Dean,P.Bullock,H.Echols,andJ.Hurwitz. 1987.Unwinding
ofduplex
DNA from the SV40 origin ofreplication by
Tantigen.
Science238:964-967.20.
Domier,
L. L., K. M. Franklin, M. Shahabuddin, G. M.Hell-mann,J.H.
Overmeyer,
S. T.Hiremath, M. F. E. Siaw,G. P.Lomonossoff,
J.G.Shaw,
and R.E. Rhoads.1986. The nucleo-tide sequence of tobacco vein mottling virus RNA. Nucleic Acids Res. 14:5417-5430.21. Elias, P., C. M. Gustafsson, and 0. Hammarsten. 1990. The
origin binding protein
ofherpes
simplexvirus 1bindscoopera-tively
to the viralorigin
ofreplication
oris. J. Biol. Chem. 265:17167-17173.22. Elias, P.,and I. R. Lehman.1988. Interaction oforigin binding
protein withanoriginofreplicationofherpes simplexvirus 1. Proc. Natl. Acad. Sci. USA85:2959-2963.
23. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radiolabelingDNA restrictionfragmentsto high specific activ-ity. Anal. Biochem. 132:6-13.
24. Fierer, D. S., and M. D. Challberg. 1992. Purification and characterizationofUL9,theherpes simplexvirustype1 origin-binding protein. J. Virol.66:3986-3995.
25. Fry,D.C.,S. A.Kuby,and A. S.Mildvan. 1986.ATP-binding site ofadenylate kinase: mechanistic implicationsof its homol-ogy with ras-encoded p21, Fl-ATPase, and other nucleotide-binding proteins. Proc. Natl. Acad. Sci. USA83:907-911. 26. GaHo,M.L.,D. I.Dorsky,C. S.Crumpacker,and D. S.Parris.
1989. The essential 65-kilodalton DNA-binding proteinof her-pessimplex virus stimulates the virus-encoded DNA
polymer-ase.J. Virol. 63:5023-5029.
27. Goldstein,D.J., and S. K. Weler. 1988. An ICP6::lacZ inser-tionalmutagen is usedto demonstratethat theUL52 gene of herpes simplex virus type 1 is required for virus growth and DNAsynthesis.J.Virol. 62:2970-2977.
28. Goldstein, D.J., and S. K. Weller. 1988.Factor(s) present in herpes simplex virustype 1-infected cellscan compensate for the loss of thelarge subunit of the viral ribonucleotide
reduc-tase: characterization of an ICP6 deletion mutant. Virology 166:41-51.
29. Gorbalenya,A.E.,and E. V. Koonin.1988.Onemoreconserved sequencemotif in helicases. Nucleic AcidsRes. 16:7734. 30. Gorbalenya,A.E.,E. V.Koonin, A.P.Donchenko,and V. M.
Blinov. 1988. A conserved NTP-motif in putative helicases. Nature(London)333:22-23.
31. Gorbalenya, A. E., E. V. Koonin, A. P. Donchenko, and V. M. Blinov. 1988. A novelsuperfamily of nucleoside triphosphate-bindingmotifcontainingproteins whichareprobablyinvolved induplex unwindingin DNA and RNAreplicationand
recom-bination. FEBS Lett. 235:16-24.
32. Gorbalenya,A.E., E. V.Koonin,A.P.Donchenko, and V. M. Blinov. 1989. Two related superfamilies of putative helicases involved inreplication, recombination, repair and expressionof DNAandRNA genomes. NucleicAcidsRes. 17:4713-4730. 33. Graham,F.L.,and A.J.vander Eb.1973. Anewtechniquefor
theassayofinfectivity of human adenovirus 5DNA. Virology 52:456-467.
34. Higgins, D. F., M. P.Gallagher, M. L.Mimmack, and S. R. Pearce. 1988.AfamilyofcloselyrelatedATP-binding subunits fromprokaryoticandeukaryoticcells.Bioessays 8:111-116. 35. Hloch, P., G. Scheidner,andH. Stahl. 1990. CompletecDNA
sequenceof the humanp68 protein.NucleicAcidsRes.18:3045. 36. Hodgman, T. C. 1988. A new superfamily of replicative
pro-teins. Nature(London)333:22-23.
37. Hodgman, T. C. 1988. A new superfamily of replicative pro-teins. Nature(London)333:578.(Erratum.)
38. Koff, A., J. F. Schwedes, and P. Tegtmeyer. 1991. Herpes simplexvirus origin-binding protein(UL9) loops and distorts the viralreplication origin.J. Virol. 65:3284-3292.
39. Koff, A., and P. Tegtmeyer. 1988. Characterization of major recognitionsequencesforaherpes simplexvirus type 1 origin-binding protein.J.Virol. 62:4096-4103.
40. Kunkel,T. A.1985.Rapidandefficientsite-specificmutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
41. Kunkel,T.A., J.D.Roberts,and R. A. Zakour. 1987.Rapidand efficientsite-specificmutagenesiswithoutphenotypicselection. MethodsEnzymol. 154:367-382.
42. la Cour,T.F.M.,J. Nyborg, S. Thirup, and B.F. C. Clark. 1985. Structuraldetailsof thebindingofguanosine diphosphate
to elongation factor Tu from E. coli as studied by X-ray crystallography.EMBOJ.4:2385-2388.
43. Laemmli,U. K.1970.Cleavageof structuralproteins during the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
44. Landschulz,W.H.,P. F.Johnson,andS. L. McKnight. 1988. The DNAbindingdomain of theratliver nuclearproteinC/EBP isbipartite.Science 240:1759-1764.
VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
45. Lane, D. 1988. Enlarged family of putative helicases. Nature (London) 334:478. (Letter.)
46. Liu, Q., and W. C.Summers. 1988. Site-directed mutagenesis of anucleotide-binding domaininHSV-1 thymidine kinase: effects on catalytic activity.Virology 163:638-642.
47. Malik, A. K., R Martinez, L. Muncy, E. P. Carmichael, and S.K. Weller.Geneticanalysisofmutations in the HSV-1UL9 origin specificDNAbinding protein: isolation ofanICP6::1acZ insertion mutant.Virology, in press.
48. Malik, A. K., and S. K. Weller. 1992.Unpublished results. 49. Maniatis, T., E. F. Fritsch,andJ.Sambrook. 1982. Molecular
cloning:alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
50. Marchetti, M. E., C. A. Smith, and P. A. Schaffer. 1988. A temperature-sensitive mutation inaherpessimplex virus type1 gene required for viral DNA synthesis maps to coordinates 0.609through0.614 inUL.J.Virol. 62:715-721.
51. Matz, B., J.H.Subak-Sharpe,andV. G.Preston.1983.Physical mapping oftemperature-sensitive mutations of herpes simplex virus type 1 usingcloned restrictionendonucleasefragments.J. Gen. Virol. 64:2261-2270.
52. Naumovski, L., and E. C. Friedberg. 1986. Analysis of the essential and excision repair functions of the RAD3 gene of Saccharomyces cerevisiae by mutagenesis. Mol. Cell. Biol. 6:1218-1227.
53. Olivo,P.D.,N.J.Nelson,and M. D.Challberg. 1988. Herpes simplex virus DNA replication: the UL9 gene encodes an origin-binding protein. Proc. Natl. Acad. Sci. USA 85:5414-5418.
54. Olivo,P.D.,N.J.Nelson,and M. D.Challberg. 1989. Herpes simplex virus type 1geneproductsrequired forDNA replica-tion:identification andoverexpression.J. Virol. 63:196-204. 55. Pakula, A. A., andR.T.Sauer. 1989. Geneticanalysis of protein
stability and function. Annu. Rev. Genet. 23:289-310. 56. Parris, D. S.,A.Cross, L. Haarr, A.Orr, M. C. Frame, M.
Murphy,D.J.McGeoch, and H. S. Marsden. 1988. Identifica-tion of the geneencoding the 65-kilodalton DNA-binding protein ofherpessimplex virus type 1. J. Virol. 62:818-825.
57. Purifoy, D. J. M., R. B. Lewis, and K. L. Powell. 1977. Identification of the herpes simplex virus DNA polymerase gene. Nature(London) 269:621-623.
58. Purifoy, D. J. M., and K. L. Powell. 1981. Temperature-sensitivemutants in two distinct complementation groups of herpes simplex virus type1specify thermolabileDNA polymer-ase.J.Gen. Virol. 54:219-222.
59. Rabkin, S.D., and B. Hanlon. 1991. Nucleoprotein complex formed between herpes simplex virus UL9 protein and the origin of DNA replication: inter- and intramolecular interac-tions. Proc. Natl. Acad. Sci. USA 88:10946-10950.
60. Russell, M., S. Kidd, and M. R Kelley. 1986. An improved filamentoushelper phage forgenerating single-stranded plasmid DNA. Gene 45:333-338.
61. Shao, L.,and S. K.Weller. 1992.Unpublished results. 62. Song, J. M.,B. A.Montelone,W.Siede,and E.C.Friedberg.
1990. Effects of multiple yeast rad3 mutant alleles on UV sensitivity,mutability, and mitotic recombination.J.Bacteriol. 172:6620-6630.
63. Spaete,RR.,and N. Frenkel. 1982. Theherpessimplexvirus amplicon: a new eucaryotic defective-viruscloning-amplifying vector. Cell 30:295-304.
64. Stahl, H., P. Droege, and R. Knippers. 1986. DNA helicase activityofSV40largetumorantigen.EMBO J.5:1939-1944. 65. Stow,N.D. 1982.Localization ofanorigin ofDNAreplication
within the TRS/IRS repeated region of the herpessimplex virus type1genome. EMBOJ. 1:863-867.
66. Stow,N.D.,and E.C.Monagle. 1983. Characterization of the TRS/IRS origin ofDNAreplication ofherpes simplex virus type 1.Virology 130:427-438.
67. Sung, P.,D.Higgins, L.Prakash,and S.Prakash. 1988. Muta-tion oflysine-48toarginine in the yeast RAD3 protein abolishes itsATPase and DNA helicase activities but not the abilityto bind ATP. EMBO J. 7:3263-3269.
68. Towbin, H.,T.Staehelin,andJ. Gordon.1979.Electrophoretic transfer ofproteins frompolyacrylamide gels to nitrocellulose sheets: procedure andsomeapplications. Proc. Natl. Acad. Sci. USA76:4350-4354.
69. Triezenberg,S.J., R. C.Kingsbury,andS. L.McKnight. 1988. Functional dissection of VP16, the trans-activator of herpes simplex virus immediate early gene expression. Genes Dev. 2:718-729.
70. Vieira, J.,andJ. Messing. 1987. Production ofsingle-stranded plasmid DNA.Methods Enzymol. 153:3-11.
71. Walker,J. E.,M.Saraste, M. J. Runswick, and N. J. Gay. 1982. Distantly related sequences in the a- and
P-subunits
of ATP synthase, myosin, kinases and other ATP-requiring enzymes andacommon nucleotidebinding fold. EMBOJ. 1:945-951. 72. Weeda,G.,R.C. A. vanHam, W.Vermeulen, D. Bootsma,A.J.vanderEb, and J. H. J.HoeUmakers. 1990. A presumedDNA helicase encoded by ERCC-3 is involved in the human repair disorders xerodermapigmentosum and Cockayne's syndrome. Cell 62:777-791.
73. Weir, H. M., J.M.Calder,and N. D.Stow. 1989.Binding of the herpessimplex virus type1UL9 geneproduct to an origin of viralDNAreplication.Nucleic Acids Res. 17:1409-1425. 74. Weller, S.K. 1990.Geneticanalysis of HSV genes required for
genomereplication, p. 105-135.In E.Wagner(ed.), Herpesvi-rus transcription and its regulation. CRC Press, BocaRaton, Fla.
75. Weller,S.K.,E. P.Carmichael, D.P.Aschman,D.J.Goldstein, and P. A. Schaffer. 1987.Genetic andphenotypic characteriza-tion ofmutantsinfour essential genes that maptothe left half of HSV-1 ULDNA. Virology 161:198-210.
76. Weller, S.K., K.J. Lee, D. J.Sabounn, and P. A. Schaffer. 1983. Geneticanalysis of temperature-sensitive mutants which define the gene for the major herpes simplex virus type 1 DNA-binding protein.J.Virol.45:354-366.
77. Weller,S.K.,A.Spadaro, J.E.Schaffer,A.W.Murray,A. M. Maxam, and P. A. Schaffer. 1985. Cloning, sequencing, and functionalanalysis ofo0L,aherpessimplex virus type1origin ofDNAsynthesis. Mol. Cell. Biol. 5:930-942.
78. Wu,C.A.,N.J.Nelson,D.J.McGeoch,and M.D.Chaliberg. 1988. Identification ofherpes simplex virus type 1 genes re-quired for origin-dependentDNA synthesis. J. Virol. 62:435-443.
79. Yohda,M., S. Ohta,T. Hisabori, andY.Kagawa. 1988. Site-directedmutagenesis of stable adenosine triphosphate synthase. Biochim.Biophys.Acta933:156-164.
80. Zhu, L., and S. K.Weller. 1988. UL5, aprotein required for HSVDNAsynthesis: genetic analysis, overexpression in Esch-erichiacoli, and generation of polyclonal antibodies. Virology 166:366-378.
81. Zhu, L.,and S. K. Weller. 1992. The sixconserved helicase motifs of the UL5 gene product, a component of the herpes simplex virus type 1 helicase-primase, are essential for its function. J.Virol. 66:469-479.